
Life’s a Gas: A Thermodynamic Theory of Biological Evolution

Biosphere Research Institute, 5A The Den, Letham, Angus, DD8 2PY, UK
Entropy 2015, 17(8), 5522-5548; https://0-doi-org.brum.beds.ac.uk/10.3390/e17085522
Submission received: 4 May 2015
/
Revised: 9 July 2015
/
Accepted: 28 July 2015
/
Published: 31 July 2015
(This article belongs to the Special Issue Maximum versus Minimum Entropy Generation: Theoretical Developments and Applications)
Abstract
:This paper outlines a thermodynamic theory of biological evolution. Beginning with a brief summary of the parallel histories of the modern evolutionary synthesis and thermodynamics, we use four physical laws and processes (the first and second laws of thermodynamics, diffusion and the maximum entropy production principle) to frame the theory. Given that open systems such as ecosystems will move towards maximizing dispersal of energy, we expect biological diversity to increase towards a level, Dmax, representing maximum entropic production (Smax). Based on this theory, we develop a mathematical model to predict diversity over the last 500 million years. This model combines diversification, post-extinction recovery and likelihood of discovery of the fossil record. We compare the output of this model with that of the observed fossil record. The model predicts that life diffuses into available energetic space (ecospace) towards a dynamic equilibrium, driven by increasing entropy within the genetic material. This dynamic equilibrium is punctured by extinction events, which are followed by restoration of Dmax through diffusion into available ecospace. Finally we compare and contrast our thermodynamic theory with the MES in relation to a number of important characteristics of evolution (progress, evolutionary tempo, form versus function, biosphere architecture, competition and fitness).

1. Introduction
The nineteenth century saw a remarkable transformation in theories underpinning our understanding of biological evolution (Darwinian theory) and physical chemistry (thermodynamics). These advances occurred in parallel, and in the twentieth century, attempts were made to examine potential crossovers. Could physics inform our understanding of biological evolution or could natural selection represent a fundamental explanation for non-biological phenomena? While most research in this area has been built on the foundations of the modern evolutionary synthesis (MES), this paper presents an attempt at developing a purely thermodynamic theory of evolution. We test a thermodynamic model derived from the first and second laws of thermodynamics, diffusion theory and the maximum entropy production theory, and use this model to explore biological diversity through time. We then examine how thermodynamics impacts upon the structure and function of each level of biological organization, from molecule to biome. Before we do this, we will briefly review the historical contexts of both the MES and thermodynamic theory.
1.1. Biological Evolution
Origins and destinies have framed the human condition from as early as we have evidence, and the origin and evolution of life has been a fundamental issue in science, philosophy and the arts. As Teilhard de Chardin [1] (p. 219) famously observed, “Evolution is a light which illuminates all facts, a curve that all lines must follow”. The importance of origin is emphasised in the beginning of the title chosen by Charles Darwin for his great tome, The Origin of Species. His work is now recognized as the core doctrine of the MES. Baverstock and Rönkkö [2] recently observed that “Darwin’s theory forms the theoretical basis of all biology”.
The history of evolutionary thinking stretches back almost as far as the written record itself. Early thinking represented a materialist approach. Later, this was supplanted by a representational or teleological approach, wherein nature tended towards definite ends and was representative of universal forms or principles. The works of Plato and Aristotle developed much of this thinking, later seen in William Paley’s Benevolent Design and currently represented by the Intelligent Design movement.
The nineteenth century, a period of post-revolutionary expression, where avoidance of church, state and fate made for popular politics, whatever the field of study, would see evolutionary theory move away from a reliance on supernatural intervention, but still maintain some form of teleology. It began with the work of Lamarck [3], who put forward his theory of transmutation or transformisme, focusing upon three areas: constant progression, spontaneous generation, and the theory of acquired trait acquisition. He explained that evolution followed a path of increasing complexity, the progression of which was determined by some vital force or fluid. Changes occurring during the pre-reproductive lifetime of an organism could be passed on to its offspring. He suggested that this same vital force or fluid dictated these changes, promoting development of traits that suited the environmental context of the organism while diminishing other traits that were not useful. His work remained within the teleological approach, given the role of this vital fluid.
It was Charles Darwin who set out the idea of evolution through natural selection most cogently and in most detail. The struggle for existence between individual variants of a species, leading to selection of more successful variants, became the basis for modern evolutionary thinking. Individuals in any large population of replicating organisms differ in terms of their fecundity and mortality. The theory of Darwinian evolution, though named after Charles Darwin, was conceived by a number of gentlemen scientists during the first half of the nineteenth century, including William Wells [4], Patrick Matthew [5], Edward Blyth [6] and Alfred Russel Wallace [7]. This pre-Mendelian work lacked a proper mechanism, and can be referred to as classical Darwinism.
The classical theory was laden with teleological expressions such as natural selection (deliberately derived from the very determinate concept of artificial selection), adaptation (literally, “to fit towards”) and the tree of life. Darwin’s (and Haeckel’s) famous tree of life placed humans at the top, representing a desire to understand evolution as a journey towards our own species. Gould and Lewontin [8] argued that the concept of adaptation was teleological, using the satirical work of Voltaire [9], Candide, to evoke significant discussion amongst evolutionary biology at the time.
The idea of progress, with humans as the perfected state, became married to enlightenment thinking of the time, where education, industry, rationalism and liberty would deliver a utopian outcome. The best, fittest and strongest would emerge from natural selection.
It was the work of Gregor Mendel [10] that would transform Darwinian theory into a science rather than an analogy, demonstrating the inheritance of characteristics. More importantly, it provided a mathematical foundation. This set in place the advent of a new school of evolutionary thinking, statistical Darwinism.
With the concept of alleles and genes now understood if not yet physically proven, mathematical approaches and models of evolution could be attempted. Fisher, Haldane, de Vries and Wright embarked on this work along with others, at the beginning of the twentieth century, following the re-discovery of Mendel’s research, and they would turn to the recently expanding school of thermodynamics to provide a basis for their work, as we shall discuss shortly. It was Fisher’s [11] Genetical Theory of Natural Selection that set out the foundations for the MES.
Julian Huxley [12] introduced the term MES, emphasizing that small genetic changes, acted upon by natural selection, lead to gradual evolution and that macroevolution can be explained by microevolution.
The four central postulates of the MES are:
- (1)
- genetic variation (a combination of mutation, segregation and independent recombination) exists within any given population for almost all traits;
- (2)
- The transfer of genes from one generation to the next occurs independently of other genes;
- (3)
- In general, more offspring are produced than survive to reproduce;
- (4)
- Individuals that do survive and procreate represent a subset of the population of that generation which are genetically best adapted to their environment.
As a result fitness represents the degree to which copies of an individual’s genes are passed into the next generation.
The MES now focuses on the gene as the unit of selection and the central dogma was developed, wherein the biosphere is an outcome of the selfish gene, a view first put forward by Colin Pittendrigh [13]. The gene took on character traits, imbuing it with anthropomorphic selfishness. The selfish gene became an important principle, as advocated by George Williams, William Hamilton and Richard Dawkins. It represented the dominant concept of what became known as neo-Darwinism.
1.2. Thermodynamics
1.2.1. Beginnings
The 19th Century also witnessed a revolution in physical chemistry, with the development of the field of thermodynamics. Similarly to Darwinian evolution, this area arose initially in the form of classical thermodynamics, with the study of heat dissipation and heat conversion to mechanical work in steam engines. Classical thermodynamics is a macroscopic theory of matter. Central to this was the work of Nicolas Carnot. In 1824, Carnot published his book Réflections sur la Puisance Motrice du Feu [17], in which he concluded that:
- -
- Heat could neither be created nor destroyed and that the total heat of the universe was constant;
- -
- When a temperature gradient exists, work can be done;
- -
- There can be no such thing as perpetual motion.
It would be another 30 years before Rudolf Clausius [18] and William Thomson [19] would formally develop the first and second laws of thermodynamics. Clausius [20] gave these laws their simplest and clearest definitions:
First law: The energy of the Universe is constant;
Second Law: the entropy of the universe tends to a maximum (Clausius introduced the term entropy, meaning transformation, which is a measure of the thermal energy of a system per unit temperature that is unavailable for doing work).
Walther Nernst [21] defined the third law of thermodynamics, which states that as the absolute temperature approaches zero Kelvin, the entropy of a perfect crystal of a pure substance approaches zero. The zeroth Law of thermodynamics, considered an essential foundation for the other laws, but developed years later, is attributed to Saha and Srivastva [22] who wrote that “If a body A is in temperature equilibrium with two bodies B and C, then B and C themselves will be in temperature equilibrium with each other”.
The period of classical thermodynamics was followed by that of statistical thermodynamics, based on probabilities of distribution at the microscopic scale. Macroscopic properties were derived from their microscopic constituents. It was Ludwig Boltzmann who developed the concept of the progressive production of molecular randomness. Just as Darwin had focused on heterogeneity between individual organisms as a basis for a mechanistic model of evolution, Boltzmann developed a mechanistic model of macroscopic phenomena focused upon molecular heterogeneity. This field of study became known as statistical mechanics, with contributions from Boltzmann, Maxwell and Gibbs forming the foundation. Boltzmann expressed the molecular heterogeneity in terms of entropy, where S = klogW (where S = entropy, W = the number of energy levels available at a particular temperature and k = Boltzmann’s constant). Importantly, diffusion increases the entropy of a system, as predicted by the Second Law.
With the development of information theory, thermodynamics has been further transformed into a broader set of relations and is being employed across an increasingly broad range of applications. Josiah Gibbs [23] developed an equation expressing entropy as a function of the probability (pi) that a randomly selected particle existed in a particular energy state: S = −k∑pilogpi. This equation was later utilized as the basis of the Shannon Weiner Index used in information theory and, later, in biological diversity. It was not until the middle of the twentieth century that Jaynes [24,25] unified the work of Shannon, Gibbs and Boltzmann, when he showed that thermodynamic entropy is equivalent to the information entropy of a given distribution.
Hatsopoulos and Keenan [26] subsumed the 0th, 1st and 2nd laws into a Unified Principle of Thermodynamics which states that when an isolated system performs a process after the removal of a series of internal constraints, it will reach a unique state of equilibrium: this state of equilibrium is independent of the order in which the constraints are removed.
1.2.2. The Maximum Entropy Production Principle (MEPP)
Early in the development of thermodynamics, the concept of the maximum entropy production principle (MEPP) was germinating. Marcelin Berthelot argued that chemical change moved in the direction of maximum heat production [27]. Following on from Rayleigh’s dissipation function [28], Lars Onsager [29] made the observation that thermodynamic systems reduce barriers to increasing entropy:
(where dS/dt represents the rate of entropy change and I is the impediment to entropy increase). He demonstrated that in Bénard cells, I is minimized and dS/dt is maximized. Hans Zeigler [30] further developed Onsager’s work, and is often credited with formally defining the MEPP. Jaynes’ Entropy Concentration Theory demonstrated that the majority of the distributions allowed by constraints have entropies near the maximum value [31].
dS/dt . I = maximum
In its modern expression, the MEPP states that “non-equilibrium thermodynamic systems are organized in steady state such that the rate of entropy production is maximized ” [32].The MEPP adds to the second law of thermodynamics by not only including the direction of change, but the rate of change.
Kleidon and Lorenz [33] and Martyushev and Seleznev [34] concluded that open systems interacting with the external environment in non-linear ways are stabilized in such a way as to maximize entropy production. Harte [35] further showed that the production of information entropy would also be maximized when a system far from equilibrium undergoes a transition from one macrostate to another.
The MEPP was first applied to the earth system in 1975. Paltridge [36] predicted atmospheric and oceanic dissipation, cloud cover, surface temperatures and energy fluxes to a high degree of agreement with observations. Since then, the MEPP has been used in a wide range of non-equilibrium systems, from ecosystem succession [37] to linguistics [38] and from nucleotide sequences [39] to earthquake prediction [40].
The MEPP has been referred to as the fourth law of thermodynamics [41]. It acts locally at each hierarchical level in any complex system, with interactions limiting the entropic production at any given level (i.e., there is sub-optimality), allowing the maximum entropic production possible at the overall level. Serizawa et al. [42] note that open systems existing in a state far from equilibrium become stabilized when entropy production is maximized and this is facilitated by the emergence of dissipative structures. Importantly, dissipative structures with access to free energy will reduce internal entropy and thus become more ordered by externalizing entropic output to the environment. Thus complexity and dissipation are, in effect, two sides of the one coin. The MEPP can be seen as driving complexity.
With thermodynamics and evolution by natural selection firmly established by the end of the nineteenth century, and with both having passed through a classical phase followed by a statistical phase by the middle of the twentieth century, there have been efforts to marry these two fields. We must review these briefly before turning to the aim of this paper: a thermodynamic theory of biological evolution.
1.3. Thermodynamics and Evolution
We have seen that the MES (modern evolutionary synthesis) and thermodynamics have shared similar paths and timelines. Both now model populations rather than precise types [43]. Unsurprisingly, efforts to marry these two approaches began fairly early in their joint histories [44] and indeed, as we have seen, the development of one impacted on the other. Yet there is no definitive theory relating the laws of thermodynamics and biological evolution. There have been four distinct approaches.
Some have argued that the animate and inanimate worlds are so different that the rules applying to each cannot apply to the other. This position reflects the idea of Vitalism, a now discredited concept that assumed a special life force directing organic matter. Even recently, the non-applicability of thermodynamics to biological evolution has been maintained by some. Demetrius [45] wrote that “The science of thermodynamics is concerned with understanding the properties of inanimate matter”. Collier [46] insisted some years ago that there is no evidence that non-equilibrium thermodynamics applies to biological systems in a non-trivial manner. Pross [47] stated that neither the behaviour of a stalking lion nor the single cell phenomenon of chemotaxis is explicable in terms of the second law.
A separate school of thought sought to extend the impact of natural selection to the inanimate world. Molchanov [48] stated: “By now enough facts have been gathered to show that the main principles of Darwinism are applicable to all evolving systems, from elementary particles to galaxies”. Prigogine et al. [49] emphasised that although natural selection can profitably be reformulated in thermodynamic terms, thermodynamics does not offer a rival causal and explanatory principle to it.
In a letter to the marine biologist George Wallich, Darwin stated that “the principle of life will hereafter be shown to be part, or a consequence, of some general law” [50] (p. xiii), an idea re-iterated by Darcy Wentworth Thompson [51] (p. 270) when he wrote: “We rise from the conception of form to an understanding of the forces which gave rise to it”. However it was much later that thermodynamics was suggested as this general principle.
Boltzmann was the first to describe the struggle for existence in thermodynamic terms, when he wrote: “The general struggle for existence of animate beings is therefore not a struggle for raw materials —these, for organisms, are air, water and soil, all abundantly available—nor for energy which exists in plenty in any body in the form of heat (albeit unfortunately not transformable), but a struggle for entropy, which becomes available through the transition of energy from the hot sun to the cold earth” [52] (p. 24).
The idea that thermodynamics might somehow influence biological evolution gained further support from Alfred Lotka [53,54]. He focused on the energy flow in evolutionary processes, claiming that natural selection led to an increase in both energetic efficiency and total energy throughput. Howard Odum and Richard Pinkerton [55] would later set out the maximum power principle as a foundation for systems ecology, claiming that the principle was universally applicable. This states that “during self-organization, system designs develop and prevail that maximize power intake, energy transformation, and those uses that reinforce production and efficiency” [56] (p. 311). Significantly, unlike Lotka’s work, the maximum power principle made no reference to natural selection or competition, breaking the link with these central concepts of the MES.
Schrödinger [57] suggested that lifeforms imported negative entropy (or negentropy) and released positive entropy into their environments. Schrödinger further proposed that biological self-organization and thermodynamics would be reconciled by the study of living systems from a non-equilibrium perspective. Prigogine [58] referred to life forms as dissipative structures, and argued that early organisms evolved from non-living, far-from-equilibrium structures, increasing in complexity through increased entropy production.
Fenchel [59] demonstrated that the increasing complexity of life on our planet followed an increasingly dissipative path, the specific energy needs per unit biomass being higher for multi-cellular organisms than for unicellular organisms, and for homeothermic animals in comparison to poikilothermic animals. Ulanowicz [60] (p. 147) further linked the evolution of diversity and complexity with thermodynamics when he wrote: “In any real process, it is impossible to dissipate a set amount of energy in finite time without creating any structures in the process”.
Wiley and Brooks [61] argued that evolution is itself an entropic phenomenon. Annila and Salthe [62] (p. 301) went further, claiming that “the theory of evolution by natural selection is herein subsumed by the 2nd law of thermodynamics.” Swenson [63] has suggested that a great proportion of the behavior generally believed to be generic to either biological or cultural systems is shown instead to be generic to the physics of the expanding universe.
So does thermodynamics offer some form of unifying theory that envelopes Darwinian theory, or does it represent an alternative theory completely?
2. A Thermodynamic Theory of Evolution
As early as 1908, E. H. Moore [64] discussed the concept of abstraction in terms of unifying fields of study: “The existence of analogies between central features of various theories implies the existence of a general abstract theory which underlies the particular theories and unifies them with respect to those central features”. Much later, E. O. Wilson [65] introduced the concept of consilience, writing that “Units and processes of a discipline that conform with solidly verified knowledge in other disciplines have proven consistently superior in theory and practice to units and processes that do not conform”. We now examine the possibility that the laws of thermodynamics can offer such a superior explanation of biological evolution.
So what should a theory of biological evolution encompass?
- (1)
- An explanation of the known facts relating to the origin, architecture, structure and function of life on earth from an abiotic beginning (be that on this planet or on another astronomical body (e.g., Panspermia));
- (2)
- An explanation for the tempo of biological evolution as observed in the fossil record;
- (3)
- An explanation of the diversity and ecology of life on earth;
- (4)
- Such an explanation should not be contrary to observations made at the molecular, organismal, population nor ecosystem levels of organization.
However, before considering a thermodynamic theory for the evolution of life, we must first define what we mean by life. The classic definition of life involves seven key characteristics: reproduction, a cellular basis, growth and development, biological evolution, regulation, response and metabolism [66]. Viruses are excluded as non-cellular and non-metabolic, and do not appearing in any of the taxonomic kingdoms, while mules could equally be excluded as non-reproductive.
Erwin Schrödinger [57] defined life as that which resists decaying to disorder and to equilibrium. Lovelock [67] (p. 568) further refined this definition, which will be adopted for use in this paper, stating that “Life is one member of the class of phenomena which are open or continuous reaction systems able to decrease their entropy at the expense of substances or energy taken in from the environment and subsequently rejected in a degraded form”.
Life at some point came from non-life. This immediately challenging fact requires at least one moment of abiogenesis. However the thermodynamic definition of life, as a subclass of systems that remain far from thermodynamic equilibrium and convert free energy (or exergy) to less useful energy, allows for this transition more easily than the seven characteristics commonly used, and evades the difficulty of viruses and mules as non-organisms. It also means the transition from what we understand as abiotic to biotic is a gradient rather than a sharp step, moving from one sub-class to another, or, as Pascal et al. [68] put it, life emerged “through states of partial ‘aliveness’”. Life is merely one expression along the matter-energy continuum. Thus any discussion concerning the origin of life in a thermodynamic model need only relate to material availability, since all other considerations are outcomes of the universal laws of physics.
Since life is a thermodynamic concept, then surely the evolution of life should also be best understood through a thermodynamic lens? Experimental and theoretical evidence points to the fact that at every level of organization of the biosphere, thermodynamics plays a central role, acting as the architect of structure, function and change (Table 1). In terms of biological diversification, we theorize that there is a similar drive towards maximum entropy production, represented by change in protein structure and function, and expressed in organismal diversity.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level of organization | Thermodynamic control |
|---|---|
| DNA | Random mutations increase information entropy within genetic material while correction mechanisms increase entropy out with genetic material (i.e. correction consumes free energy) [39,69]. |
| Amino acids | Early abiogenic amino acids shown to form along a thermodynamic gradient [70]. Later biogenic amino acids produce increased entropy of formation. Early coding constrained thermodynamically in terms of concentration and availability of amino acids [71,72,73]. |
| Proteins | Amino acid content often constrained by energetic limitations [72]. Folding and function of proteins thermodynamically determined [74,75]. |
| Cells | Cellularity allows reduction of entropy within cells, providing the stability needed for cellular physiology [76]. Cells also export high levels of entropy [77]. Metabolic networks evolve towards maximum entropy production [78]. |
| Organisms | Increasing complexity brings increasing entropic dissipation. Advent of multicellularity, cellular specialization, increasing size and homeothermy all increased entropy production [76]. Form represents diffusion into ecospace, while function conforms to laws of thermodynamics. |
| Populations | Logistic growth of populations follows MEPP, wherein Krepresents Smax. Increased free energy availability allows population increase within ecosystem constraints [79]. |
| Speciation | Exploration of ecospace through random mutations delivers increased diversity, expressed as speciation if reproductive barriers are in place. Speciation is a eukaryotic trait, since bacteria share DNA plasmids between taxa making the species concept redundant for prokaryotes [80]. |
| Ecosystems | Ecological succession follows the MEPP [81,82,83] with ecosystems moving towards Smax [84]. |
| Biomes | Biome conditions determined by solar radiation density, interacting with the hydrological cycle, and forming the backdrop within which ecosystems develop [85,86]. |
| Earth system | Climate, global circulation patterns and tectonic activity all shown to follow MEPP [36,40,87]. |
We define biological evolution as a thermodynamic process by which maximum entropy production is restored. Evolution can be observed at all levels of organization, including population dynamics (where populations approach the carrying capacity, K) and succession (where ecosystems approach maximum entropic production, Smax). In each of these cases there is a directionality, towards maximum entropy production, constrained by context. The MES and EES cannot account for such directionality, nor can they embrace a systems approach. Indeed early attempts to do so, such as the concept of group selection by V.C. Wynne–Edwards [88] were aggressively attacked.
Yet throughout biology, the underlying physics cannot be ignored, and it is apparent that physics informs biology, not the opposite. We argue that biological diversification can be best understood as a diffusional process, underwritten by the MEPP.
Central to our thinking is the relationship between diffusion and entropy. Toussaint and Schneider [89] (p. 3) have observed that: “As biosystems grow and develop, they should increase their total dissipation, and develop more complex structures with more energy flow, increase their cycling activity, develop greater diversity and generate more hierarchical levels”, while Johnson noted, as early as 1981 [90] (p. 571), that “self-acceleration results from the tendency towards ever-increasing dispersal of the energy within the ecosystem as expressed in ever-increasing diversity; increasing diversity causes an increase in the rate of energy flow”. Diffusion of diversity represents an increase in entropy, moving towards an equilibrium state of maximum entropy. Indeed diversification can be argued to be a consequence of the second law of thermodynamics. But what does life diffuse into?
The volume within which biological diffusion occurs we term ecospace. We define ecospace as the multidimensional space, determined by mass-energy interactions, wherein life can exist. Ecospace does not directly map on to the concept of the niche, but rather is a systems level concept, representing an emergent property informed by all levels of organization and the laws of thermodynamics. Indeed a niche is just one point within ecospace. Ecospace can increase or decrease depending on energetic relations. For example a reduction in incident solar radiation (be it due to changes in axial tilt, axial precession and apsidal precession, increase in dust in the atmosphere from impact or volcanic activity, or seasonal change) may alter ecospace. Ecospace is defined only by the potential limitations imposed by the laws of physics. An increase in energy flux, such as eutrophication, can lead to a decrease in ecospace.
Ecospace is ultimately an emergent thermodynamic outcome, and thus is neither static nor reducible. Biological systems diffuse towards a dynamic equilibrium state within ecospace, called Smax. Diversification is a diffusional process, wherein provided that there is a concentration gradient (i.e., available ecospace), material will move from high to low concentration regions. Thus thermodynamics provides an explanation for the direction and rate of evolution, as it does the direction and rate of succession [84]. Ecospace itself may increase or decrease, leading to, respectively, an increase or a decline in diversity. In terms of species, each species represents a different location within ecospace. Thus the diffusion of life within ecospace is equivalent to the diversification of life within the biosphere. Given that diversity diffuses into ecospace, we would expect the process to continue towards a state of Dmax, wherein maximum entropy production is achieved. Thus thermodynamics provides an explanation for the direction and rate of evolution, as it does the direction and rate of succession [84]. A thermodynamic explanation of evolution represents a “general abstract theory” championed by Moore [64] and with the concept of consilience of Wilson [65].
With the emergence of RNA world, the importance of genetic material as a driver of diffusion was established. Energy lies at the basis of mutation, be it ultraviolet radiation or chemical mutagenesis. Random mutations represent an interesting trend, where information entropy increases within the genetic material, checked by energy-expensive correction mechanisms. It is noteworthy that increased entropy within the genetic material lies at the heart of evolution, whereas living organisms act to reduce entropy within themselves, while increasing entropy within the surroundings. However this increasing entropy must be controlled within limits to prevent malfunction, and there is further control at the physiological level in terms of avoiding excessively rapid burning of chemical energy.
Thermodynamically, changing sequences in proteins lead to diffusion into structural and functional space. Random mutations lead to a form of Brownian motion, where diversity expands into available space, nudged along by random mutations in the coding sequence. The outcome of this diffusive process is observed in increased variation, both genetically and phenotypically. The thermodynamic theory of evolution would posit that increasing functional and structural space, with a concomitant decrease in competitive pressure, will be increasingly filled. In the same way that a gas will expand to fill the space available, so life expands to fill the ecospace available.
At the cellular level, changes in entropy can be attributed to four distinct processes: (a) chemical reactions leading to molecular aggregation; (b) concentration gradients across the membrane produced by mass transport; (c) heat generation from cellular metabolic activity; (d) information stored in terms of genetic code in plasmid, nuclear and mitochondrial DNA [76]. Marína et al. [77] conclude that compartmentalization is the most essential development that significantly decreases the entropy within living cells during biological evolution, thus allowing greater free energy dissipation. Cellular evolution therefore represents a key step in the evolution of life in thermodynamic terms.
3. A Thermodynamic Model of Biological Diversity through Time
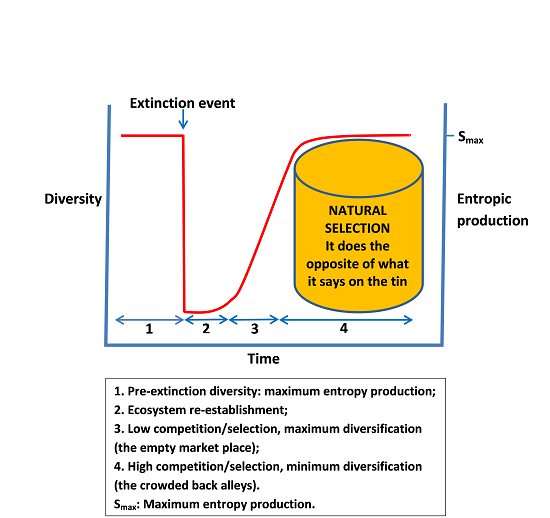
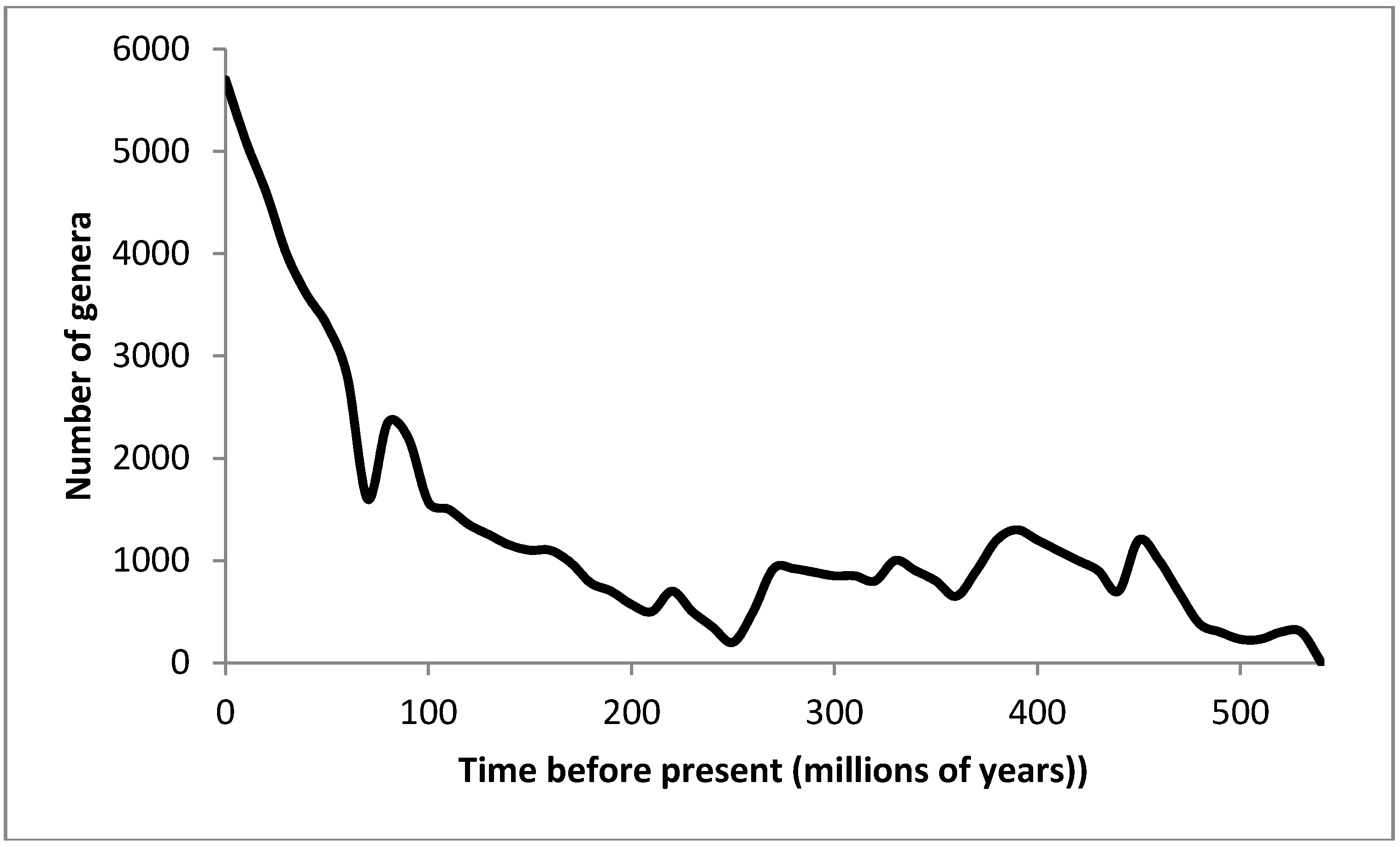
How then would thermodynamics account for the pattern of diversification over geological time? Figure 1 represents a generally accepted record of change in biological diversity, at the level of genus, over the past 500 million years, based on Bambach et al. [91]. In order to provide an adequate theoretical explanation of this pattern, we need to examine the possible contributing elements to evolutionary change through time from a thermodynamic perspective.
Figure 1.
Change in biological diversity at genus level over the last 500 m years. Data from [91].
Figure 1.
Change in biological diversity at genus level over the last 500 m years. Data from [91].

Entropy production would be expected to increase with increasing metabolism, population size, organismal complexity, trophodynamic complexity and ecological succession (see Table 1). The overall direction of evolution encompasses these changes. The driving force is the second law of thermodynamics and the fundamental outcome is maximum entropy production. There are three components to our thermodynamic model of diversification over geological time: increasing diversity towards a dynamic equilibrium of maximum entropy production; recovery from significant extinction events; decreasing likelihood of discovery of fossils with time.
3.1. Increasing Diversity Towards a Dynamic Equilibrium of Maximum Entropy Production
We suggest that diversity will increase logistically to a level representing maximum entropy production. Speciation leads to the diffusion of life throughout the ecosphere, until all available energetic space (ecospace) is filled, driven by the second law of thermodynamics. Localized extinction events lead to an increase in available ecospace, into which new species evolve. Evolution is reliant upon ecospace availability and the process of evolution is thermodynamically one of diffusion into available space. All life forms are part of an ecosystem (or more than one ecosystem), and increasing trophodynamic complexity and ecological complexity occur as part of ecosystem development. Increasing diversity is a cogent aspect of these trends. Diversity therefore moves towards maximum entropy production at both the structural and functional landscapes, and across all levels of organization (Table 1).
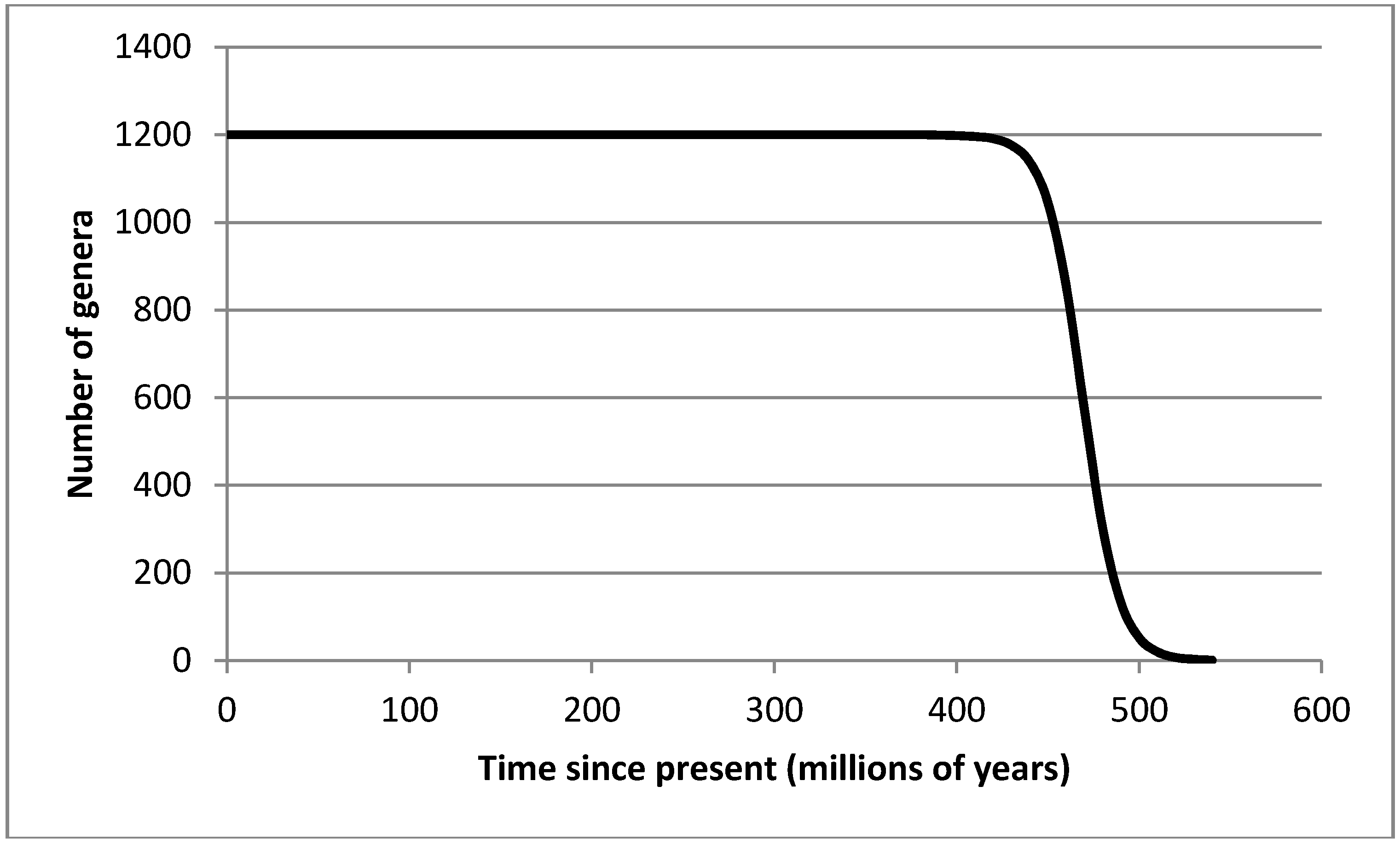
We apply the logistic equation of entropic output, derived from first principles in [84], to diversification over time. Here we view diversity as increasing toward a dynamic equilibrium, Dmax, wherein entropic production is maximized.
(D(t) = diversity at time t; rd = intrinsic capacity for diversification; Dmax = diversity at equilibrium (generating Smax); D0 = initial diversity). Dmax is set at 1200 genera, reflecting the plateau between 460 and 250 million years ago, while D0 = 1. Rd was set, by iteration, at 0.1.
Diversity therefore increases from D0 to a dynamic state of equilibrium, Dmax, following which further diversification relies upon extinction. At global Dmax, the biosphere itself reaches Smax. The same pattern is observed in individual ecosystems, where succession is driven by thermodynamics up to a point approaching Smax [37,84]. This process represents a diffusion of diversity into niche space. The results of this model are shown in Figure 2.
Figure 2.
Diversity against time, as predicted by Equation (1).

3.2. Recovery from Significant Extinction Events
Secondly, mass extinction events occur which dramatically reduce diversity. In our thermodynamic theory of evolution we would expect that following a large extinction event, diversification accelerates, restoring entropic production to Smax. The extinction events result in a geologically immediate reduction in diversity, followed by diffusion into the newly created ecospace. These extinction events represent the biological equivalent of reversed diffusion, wherein ecological simplification reduces the number of species, concentrating life into a smaller number of entities, which then have the potential to diversify, or diffuse, into the newly available space.
Each mass extinction event reduces the diversity, and thus the entropy generated, by an extinction factor that decreases over time, as shown by Alroy [92]. This post-extinction recovery can be modelled by calculating an extinction factor, Fext, and then multiplying (1) by this factor. This is done as follows:
In this case De represents the proportion of pre-existing diversity remaining immediately post extinction. We have used the values for De in Table II. Dmax = 1200 and re = 0.025 (set by iteration). Recovery is rapid (relatively) and the fastest rates of diversification are always observed in the immediate aftermath of mass extinctions, levelling off to an equilibrium level, as noted by Alroy [92]. Importantly, while Fext for each extinction event decreases with time, recovery can overlap with other extinction events if these occur close to previous events, as is the case with the Devonian and Permian extinction series. In these cases, Fext for each event are multiplied together over time. Thus recovery events occur as waves, overlapping through time.
Table 2.
Values for De (the proportion of pre-existing diversity remaining immediately post extinction) for mass extinctions through geological time (based on data from Sepkoski [93]).
| Years before present (×106) | Event | Proportion of pre-extinction diversity remaining (De) |
|---|---|---|
| 440 | Late Ordovician | 0.6 |
| 392 | Devonian 1 | 0.72 |
| 370 | Devonian 2 | 0.65 |
| 360 | Devonian 3 | 0.69 |
| 340 | Devonian 4 | 0.69 |
| 257 | End Permian 1 | 0.525 |
| 250 | End Permian 2 | 0.44 |
| 205 | Late Triassic | 0.57 |
| 183 | Torcian turnover | 0.8 |
| 154 | End Jurassic | 0.8 |
| 90 | Cenomanian | 0.86 |
| 65 | End Cretaceous | 0.6 |
| 34 | End Eocene | 0.84 |
By multiplying the predicted diversity from Equation (1) by the extinction factor in Equation (2), we model the diversity as follows:
Results are shown in Figure 3.
Figure 3.
Logistic model of diversification combined with mass extinction and recovery.

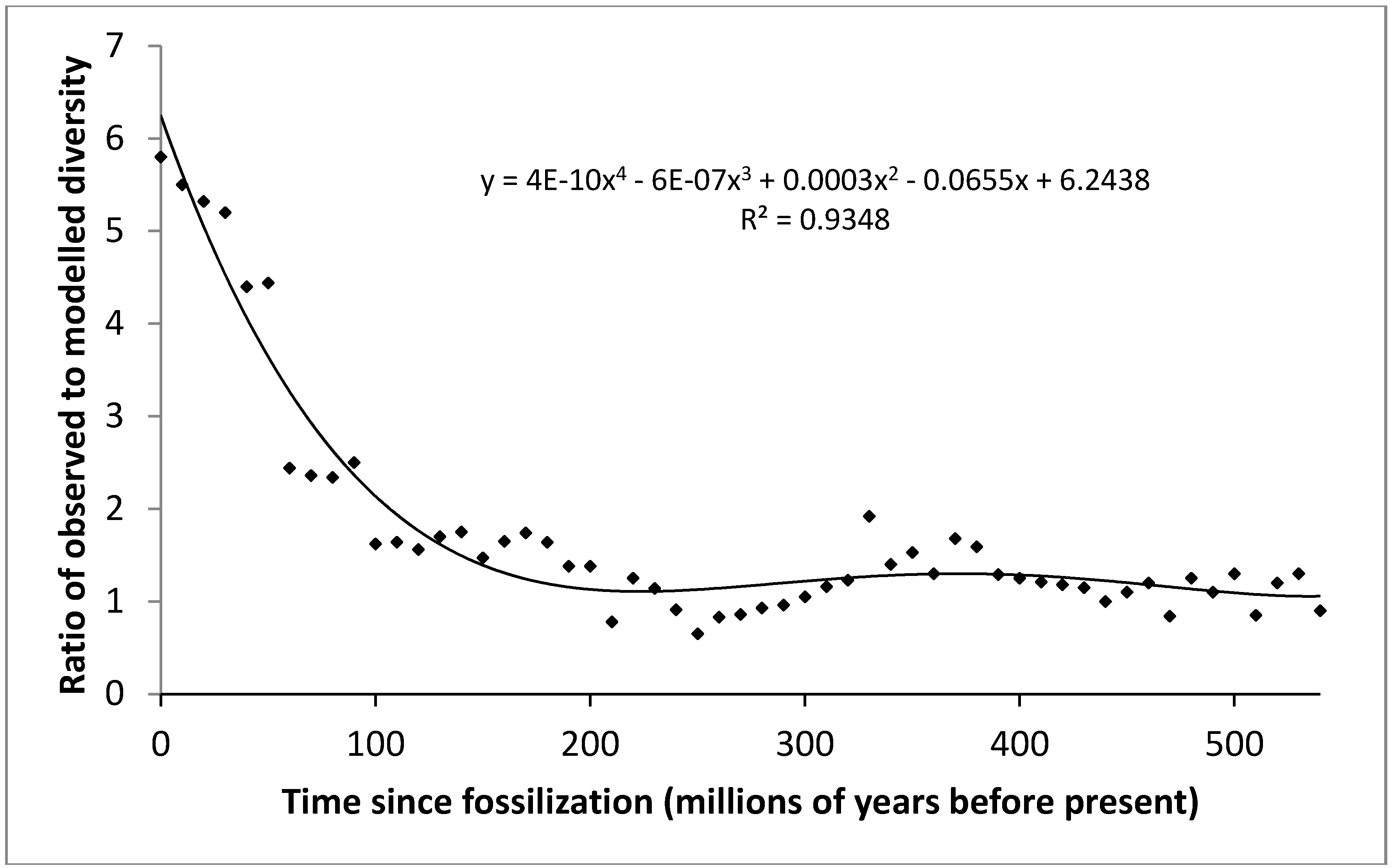
Figure 4.
Observed diversity/predicted diversity vs. time. The polynomial equation describing this relationship, the resultant curve and the coefficient of determination (R2) are shown.
Figure 4.
Observed diversity/predicted diversity vs. time. The polynomial equation describing this relationship, the resultant curve and the coefficient of determination (R2) are shown.

By dividing the observed diversification through time (Figure 1) with that predicted by our model (Figure 3), incorporating only maximum entropic production and extinction/recovery dynamics, we can identify what other changes are occurring through time. The results of this interrogation are displayed in Figure 4.
Here we can see that the results show a long lag phase, where modelled diversification equates to observed diversification (i.e., ratio approximates to one), followed by a sharp increase over the last 130 million years, where observed diversification increases at an increasingly higher rate than modelled levels. We would suggest that this is unlikely to be because of increasing niche availability, as it would be difficult to explain the increasing rate of increase in diversification in more recent geological history (i.e., the last 130 million years), but little change in the preceding 410 million years.
Movement from deep sea to ocean surface and onto land does not explain this pattern either since both events happened long before this recent acceleration. Fossil formation is unlikely to have increased rapidly in the last 130 million years. Impacts of the evolution of silica shells, bone, and other hard parts did not suddenly begin to take effect within the last 130M years, having evolved much earlier. The most obvious change lies in taphonomic integrity. Fossil survival varies inversely with length of time since deposition.
3.2. Likelihood of Discovery of Fossils Decreases with Age
Geological filters impact upon preservation, the proportion of facies and the amount of rock available to sample [94]. Diagenesis, weathering and metamorphosis can all alter sedimentary rock and its fossilized contents. Changing oceanic chemistry (Ca/Mg ratio, pH, silicification, etc.), biomolecular innovation (such as lignin, chitin and bone) and biological context (depth and activity of burrowers, borers, saprobes) have all had some impact on how accurately the fossil record reflects the biodiversity at any point in history [95]. Yet none of these is thought to have had an increasingly exponential impact over the last 130 million years. This leaves erosion, which, as noted by Thomas [96], be it organic or inorganic, has played a particularly significant role.
Hundreds of glaciation events over time have ground down uplifted fossil beds, while wind, rain and wave have all reduced potential fossil evidence to sediment. These processes, along with subduction, accumulate over time. We have incorporated this into our model by simulating the likelihood of discovery, L. With decreasing time since a fossil was laid down, the likelihood of discovery will increase from L0 to Lmax, the latter representing maximum recovery of fossilized material. Of course this situation is unlikely to arise, but we can be certain that it is impossible to exceed Lmax. As time since preservation decreases, the fossil record laid down at any given time will have a likelihood of discovery that is time-dependent and approaches Lmax. Beyond a certain age, little further change will be expected, as there will be a residual survival of all loss-generating events. The rate of change in the likelihood of discovery over time can be calculated using the following equation:
where L = likelihood of discovery and rL = the intrinsic rate of change in likelihood of discovery. Thus:
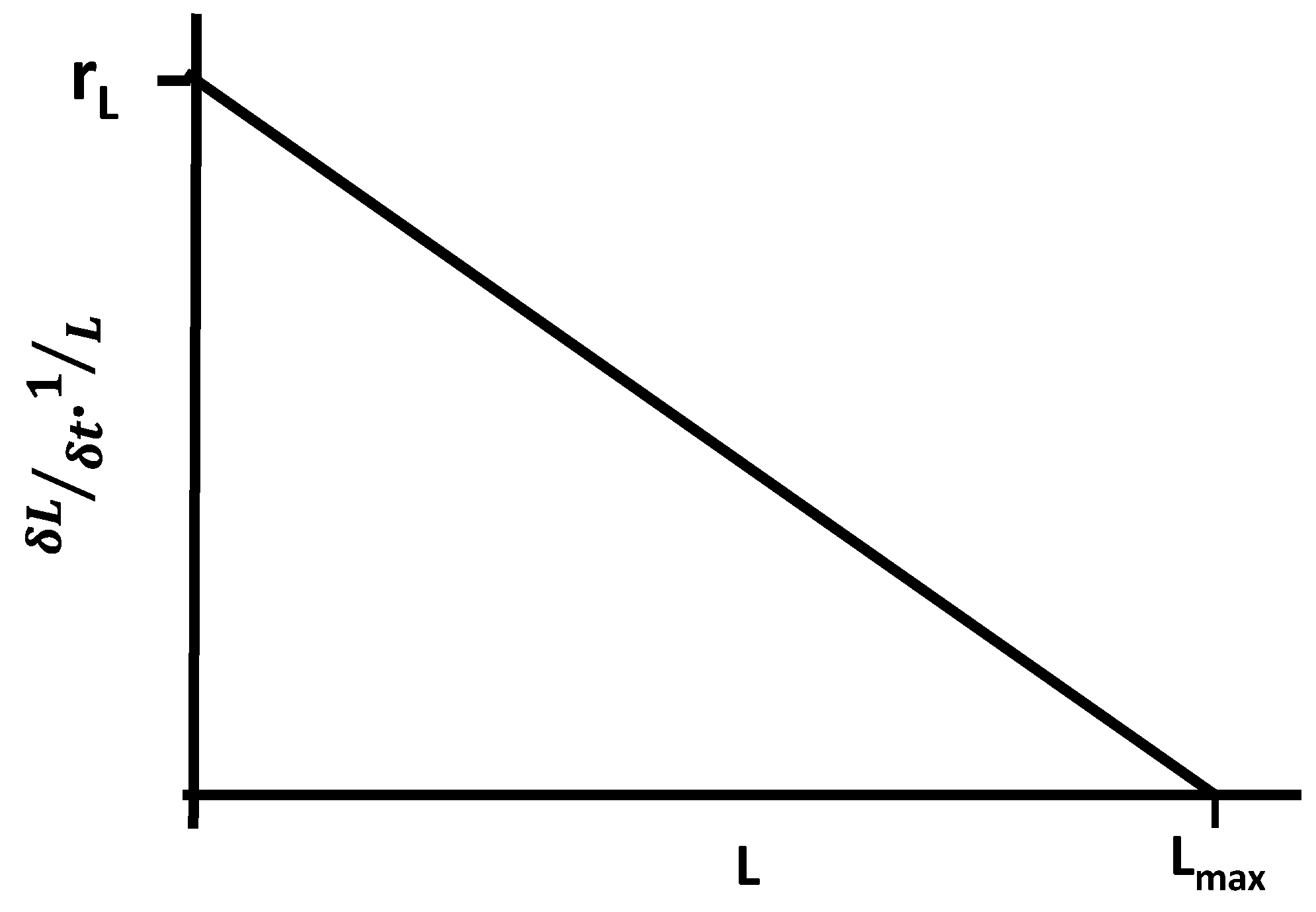
Plotting the rate of increase in discovery likelihood per unit discovery against L (Figure 5), we see that as likelihood of discovery (L) increases, the rate of change decreases towards Lmax.
This straight line relationship is described by:
Multiplying through by L and re-arranging:
Figure 5.
Rate of increase in likelihood of discovery per unit likelihood plotted against likelihood of discovery.
Figure 5.
Rate of increase in likelihood of discovery per unit likelihood plotted against likelihood of discovery.

Dividing (7) through by Lmax:
and thus:
if x = , then:
Thus, solving for L(t):
where L(t) = likelihood of discovery of fossils laid down at time, t.
The following values were used: Lmax = 100, L0 = 1 and rL = 0.0038 (set iteratively).
Thus the longer a fossil has been laid down, the less likely it is to be discovered. Changes in taphonomic catabolism (destruction of the fossil record over time) mean that beyond a certain time, the fossil record will have been exposed to most of the extremes of erosion, leaving a relatively constant basic level of discovery, close to L0. This is represented by the lag phase in Figure 4, over the first 410 million years. In more recent times, it becomes likely that the fossil record remains increasingly intact, as significant, but rarely happening events have occurred less often over increasingly shorter periods of time.
If we combine the three components (diversification to Dmax, mass extinction dynamics and likelihood of discovery) together:
The results are shown in Figure 6 and compared with the observed fossil record. There is a significant correlation coefficient of r = 0.866 (p<0.001). As can be seen, only in the latter stages does the observed diversity increase compared to the modelled diversity, and we expect this to be due to the rapidly increasing impact of increased likelihood of discovery of the fossil record laid down more recently, exceeding the logistic model used.
Figure 6.
A comparison of modelled (solid line) and observed (dashed line) diversity through time adjusted for taphonomic loss. The thermodynamic model has a highly significant correlation with the observed fossil record (r = 0.866, p<0.001).
Figure 6.
A comparison of modelled (solid line) and observed (dashed line) diversity through time adjusted for taphonomic loss. The thermodynamic model has a highly significant correlation with the observed fossil record (r = 0.866, p<0.001).

4. Discussion
The first law of thermodynamics sets in place the fundamentals of life. No life form can create energy, and thus we must obtain it from our environment. The Second Law explains that while energy cannot be destroyed, it can be transformed from useful, free energy, to less useful energy, and that this process is happening constantly. Thus to maintain order, free energy must be acquired. This is the basis of trophic dynamics.
An outcome of the second law is diffusion [97,98], wherein material moves down a concentration gradient in the absence of convective forces. Within our thermodynamic theory of evolution, diffusion is central to any understanding of the tempo of evolution, the properties of the biosphere and to behaviour at physiological and ecological levels. Evolution is a direct outcome of the laws of thermodynamics. In fact we would go as far as to state that evolution may be considered as a diffusional process.
Many of the fundamental processes of both population genetics (for example Fisher’s diffusion approximation to describe the evolution of allele frequencies) and statistical physics are described by diffusion. Fisher went so far as to draw a comparison between the theory of gases and Mendelian genetics (see [99]) reflected in the title of this paper. The reality is that within the Biosphere, the checks, balances and asymptotes act to ensure low competition most of the time. A population explosion is extremely rare and tends to signify ecosystem collapse. More regularly, stochastic events and feedback ensure the survival of the fitting and the diffusion of life into the available space with random leaks (species extinctions) occurring regularly. Occasionally, almost total evacuation of ecospace may occur, during mass extinctions. Mutation, leading to increasing entropy within the genetic material creates new types of particles, which diffuse into the structural and functional space available, a process we refer to commonly as evolution.
Finally, the maximum entropy production principle (MEPP) has a central place in our thermodynamic theory of evolution. Providing important insights into the tempo and direction of evolution, it allows an understanding of the response of the biosphere to significant perturbation, while also underpinning many of the key aspects of growth (both population and individual) and succession. Martyushev and Seleznev [100] (p. 1) observe that “at each hierarchical level of evolution, with pre-set external constraints, a local relationship between the cause and the response of a non-equilibrium system is established in order to maximize the local entropy production”. This is evidenced in Table 1.
Combining these four elements, we have derived a set of equations that represents the first attempt at modelling the evolutionary history of life on earth from a thermodynamic perspective. Importantly, the laws of thermodynamics apply to every level of organization (Table 1) and are central to the development, structure and functioning of these levels [101]. Furthermore, the very fact that there are different levels of organization, each interdependent upon one another, indicates that we expect sub-optimality within each level, rather than optimization, or selfishness [102]. Also important is the fact that the laws of thermodynamics impact on every chemical reaction, be it a biotic or abiotic system, and that respiration, which is fundamental to the supply of the free energy for growth and maintenance, is an entropy-generating process.
4.1. A Comparison of Natural Selection and Thermodynamics as Theories of Biological Evolution.
4.1.1. Progress
The subject of progress has been central to evolutionary and, more broadly, philosophical thinking for centuries, from the tree of life to more recent molecular analysis. Life is seen as moving from prokaryotic to eukaryotic, from unicellular to multicellular and from sea to land. The dominant economic and philosophical model of the current age, Humanism or enlightenment thinking, very much celebrates progress. There is perceived direction to all of this, culminating in Homo sapiens sapiens. Darwin stated, at the end of his greatest work, that “From the war of nature, from famine and death, the most exalted object which we are capable of conceiving, namely the production of the highest animals, directly flows” [103] (p. 429). Yet random mutations lie at the basis of the MES and EES, leaving little explanation space for such progress and direction.
Issues related to progress spill over into ecology, where ecological succession, as its name suggests, moves through a series of stages in a more or less predictable way. Of course differences can occur between similar ecological locations, but generally there are few surprises. The dunes become increasingly occupied and we move from an annual-dominated habitat to a perennial-dominated habitat, with drops in salinity and pH and rises in organic matter, nutrient heterogeneity and complexity [84,104]. There is directionality here, and a species replacement/displacement which cannot be obviously explained by a “selfish gene” argument. Indeed, ecological succession leads to the replacement of gene conglomerates by completely different species.
The point here is that theories related to molecular evolution must also work at the ecosystem level, and so evolution and ecology must both be considered. Furthermore, functionally, ecosystems tend to move along particular functional gradients, as observed by Odum [105], Neutel et al. [106] and van de Koppel et al. [107]. There are limits to the complexity achievable, both in terms of free energy availability (trophodynamics) and from other levels of organization. For example, Lindeman [108] showed that thermodynamic constraints on energy flow through trophic webs led to the reduction in the energy supplied to successive trophic levels.
Within our thermodynamic theory of evolution, there is a clear explanation, which also applies directly to ecology. The biosphere and all of its levels of organization move towards a state of maximum entropy production, clearly exhibited in ecological succession and, we suggest, in evolution.
4.1.2. Tempo
The fossil record also poses challenges in terms of the tempo of evolution. Figure 2 summarizes current knowledge on the number of genera over the last 500 million years. We see that there is neither a gradual increase in numbers of genera nor a constant level of diversity. There are sharp drops, followed by relatively rapid periods of speciation. These rapid phases are then followed by periods of equilibrium, suggesting either no speciation or a dynamic equilibrium of speciation and extinction. However the fastest rates of biological evolution always occur following the sharp declines, which we recognize as being correlated with mass extinction events and other lesser extinction events throughout geological history.
The MES does not attempt to address tempo in evolution and this has led to significant debate over the years. However the thermodynamic model, as presented above, can be seen to adequately account for the observed tempo across evolutionary time. There is no need to revert to Gouldian contingency [109,110]. Neither luck, nor selfish genes nor natural selection explain the pattern of observed tempo in the fossil record. The thermodynamic approach sits much more comfortably with the fact that evolution occurs at its fastest rate during low competition scenarios. As Farnsworth and Niklas [102] (p. 355) observed, “We see evolution of design more as a process of diffusion into a space of possible solutions than as a process of scalar optimization”.
4.1.3. Form vs. Function
While Darwin and Gould mostly focused on form, the thermodynamic theory of evolution holds function as the key issue. If we view evolutionary tempo from a functional point of view, we find restoration of ecosystem function rather than changes in forms as the over-riding pattern. Functionally identical food chains re-emerge, with different players, after every mass extinction. Thus evolution can be viewed as a journey of functional restoration, rather than form displacement/replacement. The direction is towards some type of functional destination, and thermodynamic theory posits that this destination is maximum entropy production.
Ecosystem function is not dependent on specific forms. Rather, forms must evolve into functional space. Thus we see trophic dynamics and entropy production as the key drivers here, rather than competitive form interactions. Function is fundamentally a thermodynamic concept. The restoration of function and functional redundancy as a key feature of resilience can all be understood easily from this perspective. Form, meanwhile, no longer needs a competitive, exclusive explanation. Given the millions of forms that exist on our planet, of every shape and size, a diffusional model provides a simpler explanation of this great diversity.
4.1.4. Biosphere Architecture
Given the reductionist nature of the MES and its extended phenotype approach [111] there is difficulty in understanding the various levels of organization as anything more than the consequences of genetic activity and behaviour, an extended phenotype. However there are clearly emergent properties throughout the biosphere [112] and a system biology approach has been advocated by many workers [113,114,115]. Furthermore, it is clear that unique activities within levels of organization beyond the gene (such as ecological succession) play critical roles in biological evolution, and under such consideration, the gene can neither be the unit of selection nor selfish.
4.1.5. Competition
Although competition is seen as a central factor in the MES, recent research has questioned its role in evolutionary biology. Venditti et al. [116] observed that most biodiversity emerges not from ongoing evolutionary war but from rare and infrequent events. Benton [117] observed that competition-driven displacement probably played a minor role in the history of tetrapods, and that family originations were most often associated with expansion into new niches. Brusatte et al. [118] noted that historical contingency, rather than prolonged competition or general superiority, was the primary factor in the rise of the dinosaurs. Alizon et al. [119] (p. 12382) commented that “under resource competition, there is an exponential slowdown of the apparent rate of evolution” while Mahler et al [120] assert that the rate of phenotypic diversity declines with decreasing opportunity. Phillimore and Prince [121] (p. 1) similarly write, relating to bird evolution, that “speciation slows as the ecological opportunities and geographic space place limits on clade growth”. This paper now provides a thermodynamic explanation for these observations, relating to maximum entropic production.
The three forms of natural selection (stabilizing, directional and disruptive selection) all create further issues. It is widely accepted that stabilizing selection is the most dominant of these three forms of selection. It results in a reduction in variation, as the fittest and most common genotypes outperform the rest of the population. However such reduced variation decreases the likelihood of both directional and disruptive selection. Furthermore, under changing environmental conditions, such as climate disruption, species tend to move rather than evolve [122,123,124]. In the diffusional approach to evolution, competition is not seen as a driver for evolution. Rather, it slows evolution by reducing variation, and thus natural selection and competition are seen as negative feedbacks, indicating the approach towards Smax.
4.1.6. Fitness
The concept of fitness, lying as it does at the heart of the MES, is problematic. It relies on a reductionist argument but has no place in a systems approach, where solutions are more likely to be sub-optimal. This is clearly seen in ecological succession, where species are lost at each stage. Two recent studies further question the significance of fitness. Rodriguez-Munos et al. [125] observed that in cricket populations in the field, sub-ordinate males have the same number of offspring as dominant males. Evans [126] noted that in guppies, less-ornamented individuals produced faster swimming sperm than larger, more decorative fish.
5. Conclusions
A thermodynamic approach can be seen to unify evolution and ecology. The MES, on the other hand, is unable to account for the emergent properties of succession, wherein some gene populations disappear over time, to be replaced with completely different gene populations, in an orderly process of ecosystem development. Genetic fitness and selfish genes appear to have no place in ecological succession, as a gene cannot proliferate beyond the point in time and space that it occurs in. Indeed, genes at one stage of ecological succession contribute towards their local extinction at the next stage in succession, where a different set of genes and organisms replace them. Furthermore, across a wide range of successional studies in different habitats, it has been shown that the same thermodynamic principles hold, again over-arching all other aspects [84]. Collier (1986) asserted that “Natural selection is merely rate-determining, and is best viewed as an extrinsic factor affecting evolutionary dynamics”. Competition, the driver of Darwinian evolution, only comes into play near entropic equilibrium, Smax. Thus, natural selection can be seen as a symptom of approaching a dynamic equilibrium, wherein a reduction in variation through competition reduces diversification, as evidenced by the fossil record following recovery from extinction. In other words, evolution occurs in the empty marketplace, not in the crowded back alleys [80].
Lotka [53] (p. 149) concluded that “evolution proceeds in such direction as to make total energy flux through the system a maximum compatible with the constraints”. Today, we recognize this pattern as the maximum entropy production principle, and this paper has shown that a thermodynamic approach can provide a powerful explanation for biological evolution, while conforming to “verified knowledge in other disciplines” [61].
Acknowledgements
The author thanks three anonymous reviewers for their helpful comments on an earlier draft. Helpful discussions with Seaton Baxter OBE, and Klement Rejsek are gratefully acknowledged.
Conflicts of Interest
The author declares no conflict of interest
References
- Teilhard de Chardin, P. The Phenomenon of Man; Harper: New York, NY, USA, 1973. [Google Scholar]
- Baverstock, K.; Rönkkö, M. The evolutionary origin of form and function. J. Physiol. Lond. 2014, 592, 2261–2265. [Google Scholar] [CrossRef] [PubMed]
- Lamarck, J.-B. Philosophie Zoologique, ou Exposition des Considérations Relative à l'Histoire Naturelle des Animaux, Volume 1; Librairie P. Savy: Paris, France, 1809. [Google Scholar]
- Wells, W.C. Two Essays: Upon a Single Vision with Two Eyes, the Other on Dew; Constable: London, UK, 1818. [Google Scholar]
- Matthew, P. Naval Timbers and Arboriculture: with Critical Notes on Authors who have Recently Treated the Subject of Planting; Adam Black: Edinburgh, UK, 1931. [Google Scholar]
- Blyth, E. An attempt to classify the “varieties” of animals, with observations on the marked seasonal and other changes which naturally take place in various British species, and which do not constitute varieties. Mag. Nat. Hist. 1835, 8, 40–53. [Google Scholar]
- Wallace, A.R. On the tendency of varieties to indefinitely from the original type. Proc. Linnean Soc. 1858, 3, 53–62. [Google Scholar]
- Gould, S.J.; Lewontin, R.C. The spandrels of San Marco and the Panglossian paradigm: a critique of the adaptationist programme. Proc. R. Soc. Lond. B 1979, 205, 581–598. [Google Scholar] [CrossRef] [PubMed]
- Voltaire. Candide, ou l’Optimisme; Cramer, Marc-Michel Rey, Jean Nourse, Lambert and others: Paris, France, 1759. [Google Scholar]
- Mendel, J.G. Versuche über Pflanzenhybriden Verhandlungen des naturforschenden Vereines in Brünn. Bd. IV für das Jahr, Abhandlungen 1866, 3–47. [Google Scholar]
- Fisher, R.A. The Genetical Theory of Natural Selection; Dover: New York, USA, 1930. [Google Scholar]
- Huxley, J. Evolution: the Modern Synthesis; Allen and Unwin: London, UK, 1942. [Google Scholar]
- Pittendrigh, C. Adaptation, natural selection and behaviour. In Behaviour and Evolution; Roe, A., Simpson, G.G., Eds.; Yale University Press: New Haven, CT, USA, 1958; pp. 390–416. [Google Scholar]
- Jablonka, E.; Lamb, M.J. Soft inheritance: challenging the modern synthesis. Genet. Mol. Biol. 2008, 31, 389–395. [Google Scholar] [CrossRef]
- Pigliucci, M.; Müller, G.B. Evolution: the Extended Synthesis; MIT Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Danchin, E.; Charmantier, A.; Champagne, F.A.; Mesoudi, A.; Pujol, B.; Blanchet, S. Beyond DNA: integrating inclusive inheritance into an extended theory of evolution. Nat. Genet. 2011, 12, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Carnot, S. Réflexions sur la Puissance Motrice du Feu et sur les Machines Propres a Développer Cette Puissance; Chez Bachelier: Paris, France, 1824. [Google Scholar]
- Clausius, R. Über die bewegende Kraft der Wärme, Part I, Part II. Ann. Phys. Berl. 1850, 79, 368–397, 500–524. [Google Scholar] [CrossRef]
- Thomson, W. On the dynamical theory of heat, with numerical results deduced from Mr Joule’s equivalent of a thermal unit, and M. Regnault’s observations on steam. Trans. R. Soc. Edinb. 1851, XX, 261–268. [Google Scholar] [CrossRef]
- Clausius, R. The Mechanical Theory of Heat: With its Applications to the Steam-Engine and to the Physical Properties of Bodies; John van Voorst: London, UK, 1867. [Google Scholar]
- Nernst, W. The New Heat Theorem; Methuen and Co.: London, UK, 1917. [Google Scholar]
- Saha, M.N.; Srivastava, B.N. A Treatise on Heat; Including Kinetic Theory of Gases, Thermodynamics and Recent Advances in Statistical Thermodynamics, the second and revised edition of A Text Book of Heat; The Indian Press: Allahabad and Calcutta, India, 1935. [Google Scholar]
- Gibbs, J.W. Elementary Principles in Statistical Mechanics; Charles Scribner’s Sons: New York, NY, USA, 1902. [Google Scholar]
- Jaynes, E.T. Information theory and statistical mechanics. Phys. Rev. 1957, 106, 620–630. [Google Scholar] [CrossRef]
- Jaynes, E.T. Information theory and statistical mechanics. II. Phys. Rev. 1957, 108, 171–190. [Google Scholar] [CrossRef]
- Hatsopoulos, G.N.; Keenan, J.H. Principles of General Thermodynamics (Vol. 398); Wiley: New York, NY, USA, 1965. [Google Scholar]
- Berthelot, M. Essai de Mecanique Chimique Fondee sur la Thermochimie; Dunod: Paris, France, 1879. [Google Scholar]
- Rayleigh, L. On our perception of the direction of a source of sound. In Proceedings of the Musical Association, 2nd Session; Royal Musical Association: London, UK, 1875; pp. 75–84. [Google Scholar]
- Onsager, L. Reciprocal relations in irreversible processes. II. Phys. Rev. 1931, 38, 2265–2279. [Google Scholar] [CrossRef]
- Ziegler, H. Some extremum principles in irreversible thermodynamics with applications to continuum mechanics. In Progress in Solid Mechanics; Sneddon, I.N., Hill, R., Eds.; North-Holland Publishing Company: Amsterdam, The Netherlands, 1963; pp. 92–193. [Google Scholar]
- Jaynes, E.T. Concentration of distributions at entropy maxima. In E. T. Jaynes: Papers on Probability, Statistics, and Statistical Physics; Rosenkrantz, R.D., Ed.; D. Reidel Publishing Company: Dordrecht, The Netherlands, 1983; pp. 315–336. [Google Scholar]
- Kleidon, A.; Malhi, Y.; Cox, P.M. Maximum entropy production in environmental and ecological systems. Proc. R. Soc. Lond. B 2010, 365, 1297–1302. [Google Scholar] [CrossRef] [PubMed]
- Kleidon, A.; Lorenz, R.D. Entropy production by earth system processes. In Non-Equilibrium Thermodynamics and the Production of Entropy: Life, Earth, and Beyond; Kleidon, A., Lorenz, R.D., Eds.; Springer: Berlin, Germany, 2004; pp. 1–20. [Google Scholar]
- Martyushev, L.M.; Seleznev, V.D. Maximum entropy production principle in physics, chemistry and biology. Phys. Rep. 2006, 426, 1–45. [Google Scholar] [CrossRef]
- Harte, J. Maximum Entropy and Ecology: On the Inference of Patterns in Nature; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Paltridge, G.W. Global dynamics and climate change: a system of minimum entropy exchange. Q. J. R. Meteorol. Soc. 1975, 101, 475–484. [Google Scholar] [CrossRef]
- Stoy, P.C.; Lin, H.; Novick, K.A.; Siqueira, M.; Juang, J.Y. The role of vegetation on the ecosystem radiative entropy budget and trends along ecological succession. Entropy 2014, 16, 3710–3731. [Google Scholar] [CrossRef]
- Berger, A.L.; Pietra, V.J.D.; Pietra, S.A.D. A maximum entropy approach to natural language processing. Comput. Linguist. 1996, 22, 39–71. [Google Scholar]
- Salamon, P.; Konopka, A.K. A maximum entropy principle for the distribution of local complexity in naturally occurring nucleotide sequences. Comput. Chem. 1992, 16, 117–124. [Google Scholar]
- Dong, W.M.; Bao, A.B.; Shah, H.C. Use of maximum entropy principle in earthquake recurrence relationships. Bull. Seismol. Soc. Am. 1984, 74, 725–737. [Google Scholar]
- Morel, R.E.; Fleck, G. A fourth law of thermodynamics. Chemistry 2006, 15, 305–310. [Google Scholar]
- Serizawa, H.; Amemiya, T.; Itoh, K. Tree network formation in Poisson equation models and the implications for the maximum entropy production principle. Nat. Sci. 2014, 6, 514. [Google Scholar] [CrossRef]
- De Vladar, H.P.; Barton, N.H. The contribution of statistical physics to evolutionary biology. Trends Ecol. Evol. 2011, 26, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Depew, D.J.; Weber, B.H. Darwinism Evolving: Systems Dynamics and the Genealogy of Natural Selection; Bradford Books/MIT Press: Cambridge, MA, USA, 1995. [Google Scholar]
- Demetrius, L. Thermodynamics and evolution. J. Theor. Biol. 2000, 206, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Collier, J. The dynamics of biological order. In Entropy, information and Evolution; Weber, B.H., Depew, D.J., Smith, J.D., Eds.; MIT Press: Cambridge, MA, USA, 1988; pp. 227–242. [Google Scholar]
- Pross, A. Toward a general theory of evolution: Extending Darwinian theory to inanimate matter. J. Syst. Chem. 2011, 2, 1–14. [Google Scholar] [CrossRef]
- Molchanov, A.M. Thermodynamics and evolution. In Oscillatory Processes in Biological and Chemical Systems; Frank, G.M., Ed.; Nauka: Moscow, USSR, 1967; pp. 292–305. [Google Scholar]
- Prigogine, I.; Nicolis, G.; Babloyantz, A. Thermodynamics of evolution. Phys. Today 1972, 25, 23–28. [Google Scholar] [CrossRef]
- De Beer, G. Some unpublished letters of Charles Darwin. Notes Rec. R. Soc. 1959, 14, 12–66. [Google Scholar] [CrossRef]
- Thompson, D.W. On Growth and Form; Cambridge University Press: Cambridge, UK, 1916. [Google Scholar]
- Boltzmann, L. The second law of thermodynamics. In Theoretical Physics and Philosophical Problems: Selected Writings; McGinness, B., Ed.; D. Reidel: New York, NY, USA, 1974; pp. 13–32. [Google Scholar]
- Lotka, A.J. Contribution to the energetics of evolution. Proc. Natl. Acad. Sci. USA 1922, 8, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Lotka, A.J. Natural selection as a physical principle. Proc. Natl. Acad. Sci. USA 1922, 8, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Odum, H.T.; Pinkerton, R.C. Time’s speed regulator: The optimum efficiency for maximum output in physical and biological systems. Am. Sci. 1955, 43, 331–343. [Google Scholar]
- Odum, H.T. Self-organization and maximum empower. In Maximum Power: The Ideas and Applications of H.T. Odum; Hall, C.A.S., Ed.; Colorado University Press: Colorado, CO, USA, 1995; pp. 311–330. [Google Scholar]
- Schrödinger, E. What is Life? The Physical Aspect of the Living Cell; Cambridge University Press: Cambridge, UK, 1944. [Google Scholar]
- Prigogine, I. Order through fluctuation: self-organization and social system. In Evolution and Consciousness; Jantsch, E., Waddington, C.H., Eds.; Addison-Wesley: London, UK, 1976; pp. 93–133. [Google Scholar]
- Fenchel, T. Intrinsic rate of natural increase: the relationship with body size. Oecologia 1974, 14, 317–326. [Google Scholar] [CrossRef]
- Ulanowicz, R.E. Ecology, the Ascendent Perspective: Robert E. Ulanowicz; Columbia University Press: New York, NY, USA, 1997. [Google Scholar]
- Wiley, E.O.; Brooks, D.R. Victims of history—a nonequilibrium approach to evolution. Syst. Biol. 1982, 31, 1–24. [Google Scholar] [CrossRef]
- Annila, A.; Salthe, S. 2010. Physical foundations of evolutionary theory. J. Non-Equilib. Thermodyn. 2010, 35, 301–321. [Google Scholar] [CrossRef]
- Swenson, R. Emergent attractors and the law of maximum entropy production: foundations to a theory of general evolution. Syst. Res. 1989, 6, 187–197. [Google Scholar] [CrossRef]
- Moore, E.H. On a form of general analysis with application to linear differential and integral equations. In Proceedings of Atti del IV Congresso Internazionale dei Matematici, Rome, Italy, 6–11 April 1908; Volume 2, pp. 98–114.
- Wilson, E.O. Consilience: The Unity of Knowledge; Knopf: New York, NY, USA, 1998. [Google Scholar]
- McKay, C.P. What is life—and how do we search for it in other worlds? PloS Biol. 2004, 2, 1260–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovelock, J.E. A physical basis for life detection experiments. Nature 1965, 207, 568–570. [Google Scholar] [CrossRef] [PubMed]
- Pascal, R.; Pross, A.; Sutherland, J.D. Towards an evolutionary theory of the origin of life based on kinetics and thermodynamics. Open Biol. 2013, 3, 130156. [Google Scholar] [CrossRef] [PubMed]
- Tessera, M.; Hoelzer, G.A. On the thermodynamics of multilevel evolution. BioSystems 2013, 113, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Higgs, P.G.; Pudritz, R.E. A thermodynamic basis for prebiotic amino acid synthesis and the nature of the first genetic code. Astrobiology 2009, 9, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Akashi, H.; Gojobori, T. Metabolic efficiency and amino acid composition in the proteomes of Escherichia coli and Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2002, 99, 3695–3700. [Google Scholar] [CrossRef] [PubMed]
- Seligmann, H. Cost-minimization of amino acid usage. J. Mol. Evol. 2003, 56, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Trifonov, E.N. The triplet code from first principles. J. Biomol. Struct. Dyn. 2004, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Anfinsen, C.B. Principles that govern the folding of protein chains. Science 1973, 181, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Lazaridis, T.; Karplus, M. Thermodynamics of protein folding: a microscopic view. Biophys. Chem. 2002, 100, 367–395. [Google Scholar] [CrossRef]
- Davies, P.C.; Rieper, E.; Tuszynski, J.A. Self-organization and entropy reduction in a living cell. Biosystems 2013, 111, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Marína, D.; Martín, M.; Sabater, B. Entropy decrease associated to solute compartmentalization in the cell. Biosystems 2009, 98, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Unrean, P.; Srienc, F. Metabolic networks evolve towards states of maximum entropy production. Metab. Eng. 2011, 13, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Aoki, I. Entropy production in living systems: from organisms to ecosystems. Thermochim. Acta 1995, 25, 359–370. [Google Scholar] [CrossRef]
- Skene, K.R. Shadows on a Cave Wall: A New Theory of Evolution; Ard Macha Press: Angus, UK, 2009. [Google Scholar]
- Aoki, I. Entropy balance in Lake Biwa. Ecol. Model. 1987, 37, 235–248. [Google Scholar] [CrossRef]
- Ludovisi, A. Biotic and abiotic entropy production in lake ecosystems. Ecol. Model. 2004, 179, 145–147. [Google Scholar] [CrossRef]
- Meysman, F.J.R.; Bruers, S. A thermodynamic perspective on food webs: Quantifying entropic production within detrital-based ecosystems. J. Theor. Biol. 2007, 249, 124–139. [Google Scholar] [CrossRef] [PubMed]
- Skene, K.R. The energetics of ecological succession: A logistic model of entropic output. Ecol. Model. 2013, 250, 287–293. [Google Scholar] [CrossRef]
- Kleidon, A. Nonequilibrium thermodynamics and maximum entropy production in the Earth system. Naturwissenschaften 2009, 96, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Westhoff, M.C.; Zehe, E.; Schymanski, S.J. Importance of temporal variability for hydrological predictions based on the maximum entropy production principle. Geophys. Res. Lett. 2014, 41, 67–73. [Google Scholar] [CrossRef]
- Lucarini, V.; Pascale, S. Entropy production and course graining of the climate fields in a general circulation model. Clim. Dyn. 2014, 43, 981–1000. [Google Scholar] [CrossRef]
- Wynne-Edwards, V.C. Animal Dispersion in Relation to Social Behavior; Oliver & Boyd: London, UK, 1962. [Google Scholar]
- Toussaint, O.; Schneider, E.D. The thermodynamics and evolution of complexity in biological systems. Comp. Biochem. Physiol. 1998, 120, 3–9. [Google Scholar] [CrossRef]
- Johnson, L. The thermodynamic origin of ecosystems. Can. J. Fish. Aquat. Sci. 1981, 38, 571–590. [Google Scholar] [CrossRef]
- Bambach, R.K.; Knoll, A.H.; Wang, S.C. Origination, extinction, and mass depletions of marine diversity. Paleobiology 2004, 30, 522–542. [Google Scholar] [CrossRef]
- Alroy, J. Dynamics of origination and extinction in the marine fossil record. Proc. Natl. Acad. Sci. USA 2008, 105, 11536–11542. [Google Scholar] [CrossRef] [PubMed]
- Sepkoski, J.J., Jr. A compendium of fossil marine families. Contrib. Biol. Geol. 1982, 51, 1–125. [Google Scholar]
- Smith, A.B.; McGowan, A.J. The ties linking rock and fossil records and why they are important for palaeobiodiversity studies. Geol. Soc. Lond. Spec. Publ. 2011, 358, 1–7. [Google Scholar] [CrossRef]
- Allison, P.A.; Bottjer, D.J. Taphonomy: bias and process through time. In Taphonomy: Bias and Process through Time; Allison, P.A., Bottjer, D.J., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 1–17. [Google Scholar]
- Thomas, R.D.K. Taphonomy: Ecology’s loss is sedimentology’s gain. Palaios 1986, 1, 206. [Google Scholar]
- Bryan, G.H. Thermodynamics of diffusion. Nature 1907, 76, 637–638. [Google Scholar] [CrossRef]
- Truesdell, C. Thermodynamics of diffusion. In Rational Thermodynamics; Truesdell, C., Ed.; Springer: New York, NY, USA, 1984; pp. 219–236. [Google Scholar]
- Kimura, M. Diffusion models in population genetics. J. Appl. Probab. 1964, 1, 177–232. [Google Scholar] [CrossRef]
- Martyushev, L.M.; Seleznev, V.D.; Maximum entropy production principle (MEPP). Comment about restrictions and typical misconceptions of critics of MEPP. 2013; arXiv:1311.2068. [Google Scholar]
- Aoki, I. Entropy and exergy in the development of living systems: A case study of lake-ecosystems. J. Phys. Soc. Jpn. 1998, 67, 2132–2139. [Google Scholar] [CrossRef]
- Farnsworth, K.D.; Niklas, K.J. Theories of optimization, form and function in branching architecture in plants. Funct. Ecol. 1995, 9, 355–363. [Google Scholar] [CrossRef]
- Darwin, C.R. The Origin of Species by Means of Natural Selection or the Preservation of Favoured Races in the Struggle for Life; Senate: London, UK, 1994. [Google Scholar]
- Averiss, R.J.; Skene, K.R. Changes in nutrient heterogeneity in a sand dune succession at Tentsmuir Point, Eastern Scotland. Bot. J. Scotl. 2001, 53, 45–56. [Google Scholar] [CrossRef]
- Odum, E.P. The strategy of ecosystem development. Science 1969, 164, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Neutel, A.M.; Heesterbeek, J.A.P.; van de Koppel, J.; Hoenderboom, G.; Vos, A.; Kaldeway, C.; Berendse, F.; Ruiter, P.C. Reconciling complexity with stability in naturally assembling food webs. Nature 2007, 449, 599–602. [Google Scholar] [CrossRef] [PubMed]
- Van de Koppel, J.; Rietkerk, M.; Weissing, F.J. Catastrophic vegetation shifts and soil degradation in terrestrial grazing systems. Trends Ecol. Evol. 1997, 12, 352–356. [Google Scholar] [CrossRef]
- Lindeman, R.L. The trophic-dynamic aspect of ecology. Ecology 1942, 23, 399–418. [Google Scholar] [CrossRef]
- Eldredge, N.; Gould, S.J. Punctuated equilibria: An alternative to phyletic gradualism. In Models in Paleobiology; Schopf, T.J.M., Ed.; Freeman Cooper: San Francisco, CA, USA, 1972; pp. 82–115. [Google Scholar]
- Gould, S.J. Wonderful Life: The Burgess Shale and the Nature of History; Norton: New York, NY, USA, 1989. [Google Scholar]
- Dawkins, C.R. The Extended Phenotype: The Long Reach of the Gene; Oxford University Press: Oxford, UK, 1982. [Google Scholar]
- Higgins, P.A.; Mastrandrea, M.D.; Schneider, S.H. Dynamics of climate and ecosystem coupling: Abrupt changes and multiple equilibria. Philos. Trans. R. Soc. Lond. B 2002, 357, 647–656. [Google Scholar]
- Von Bertalanffy, L. General system theory. Gen. Syst. 1956, 1, 11–17. [Google Scholar]
- Wolkenhauer, O. Systems biology: the reincarnation of systems theory applied in biology? Brief. Bioinform. 2001, 2, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, K. Life: an Introduction to Complex Systems Biology; Springer: London, UK, 2006. [Google Scholar]
- Venditti, C.; Meade, A.; Pagel, M. Phylogenies reveal new interpretation of speciation and the Red Queen. Nature 2010, 463, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Benton, M.J. On the non-prevalence of competitive replacement in the evolution of tetrapods. In Evolutionary Palaeobiology; Jablonski, D., Erwin, D.H., Lipps, J., Eds.; Chicago University Press: Chicago, IL, USA, 1996; pp. 185–210. [Google Scholar]
- Brusatte, S.L.; Benton, M.J.; Ruta, M.; Lloyd, G.T. Superiority, competition, and opportunism in the evolutionary radiation of dinosaurs. Science 2008, 321, 1485–1488. [Google Scholar] [CrossRef] [PubMed]
- Alizon, S.; Kucera, M.; Jansen, V.A.A. Competition between cryptic species explains variations in rates of lineage evolution. Proc. Natl. Acad. Sci. USA 2008, 105, 12382–12386. [Google Scholar] [CrossRef] [PubMed]
- Mahler, D.L.; Revell, L.J.; Glor, R.E.; Losos, J.B. Ecological opportunity and the rate of morphological evolution in the diversification of Greater Antillean anoles. Evolution 2010, 64, 2731–2745. [Google Scholar] [CrossRef] [PubMed]
- Phillimore, A.B.; Price, T.D. Density-dependent cladogenesis in birds. PLoS Biol. 2008, 6, e71. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Perry, A.L.; Low, P.J.; Ellis, J.R.; Reynolds, J.D. Climate change and distribution shifts in marine fishes. Science 2005, 308, 1912–1915. [Google Scholar] [CrossRef] [PubMed]
- Archibald, S.B.; Johnson, K.R.; Mathewes, R.W.; Greenwood, D.R. Intercontinental dispersal of giant thermophilic ants across the Arctic during early Eocene hyperthermals. Proc. R. Soc. Lond. B 2011, 278, 3679–3686. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez Muñoz, R.; Bretman, A.; Slate, J.; Walling, C.A.; Tregenza, T. Natural and sexual selection in a wild insect population. Science 2010, 328, 1269–1272. [Google Scholar] [CrossRef] [PubMed]
- Evans, J. Quantitative genetic evidence that males trade attractiveness for ejaculate quality in guppies. Proc. R. Soc. Lond. B 2010, 277, 3195–3201. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Skene, K.R. Life’s a Gas: A Thermodynamic Theory of Biological Evolution. Entropy 2015, 17, 5522-5548. https://0-doi-org.brum.beds.ac.uk/10.3390/e17085522
AMA Style
Skene KR. Life’s a Gas: A Thermodynamic Theory of Biological Evolution. Entropy. 2015; 17(8):5522-5548. https://0-doi-org.brum.beds.ac.uk/10.3390/e17085522
Chicago/Turabian StyleSkene, Keith R. 2015. "Life’s a Gas: A Thermodynamic Theory of Biological Evolution" Entropy 17, no. 8: 5522-5548. https://0-doi-org.brum.beds.ac.uk/10.3390/e17085522