
Taxis of Artificial Swimmers in a Spatio-Temporally Modulated Activation Medium


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
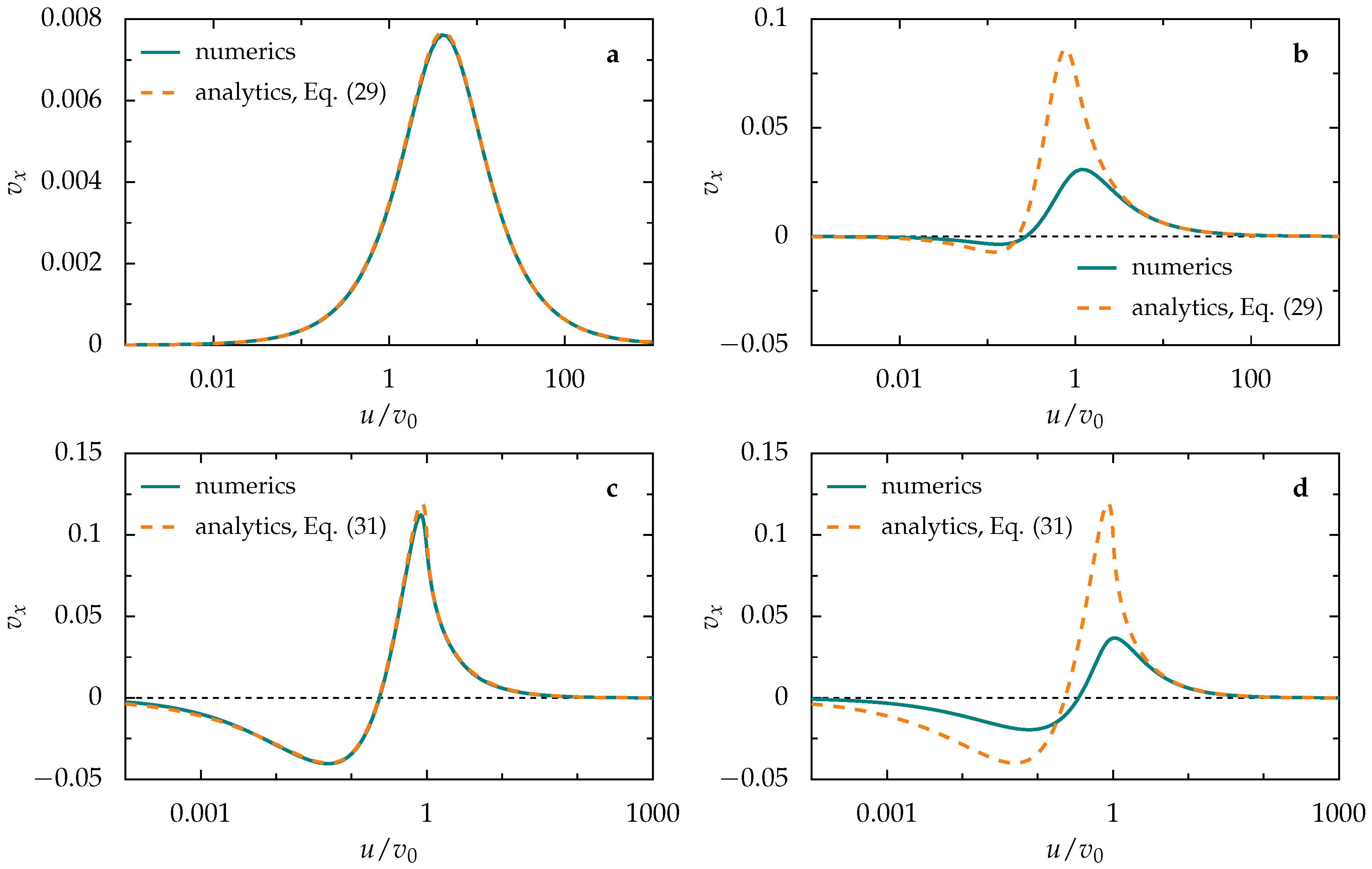{kind=link}
Abstract
:1. Introduction
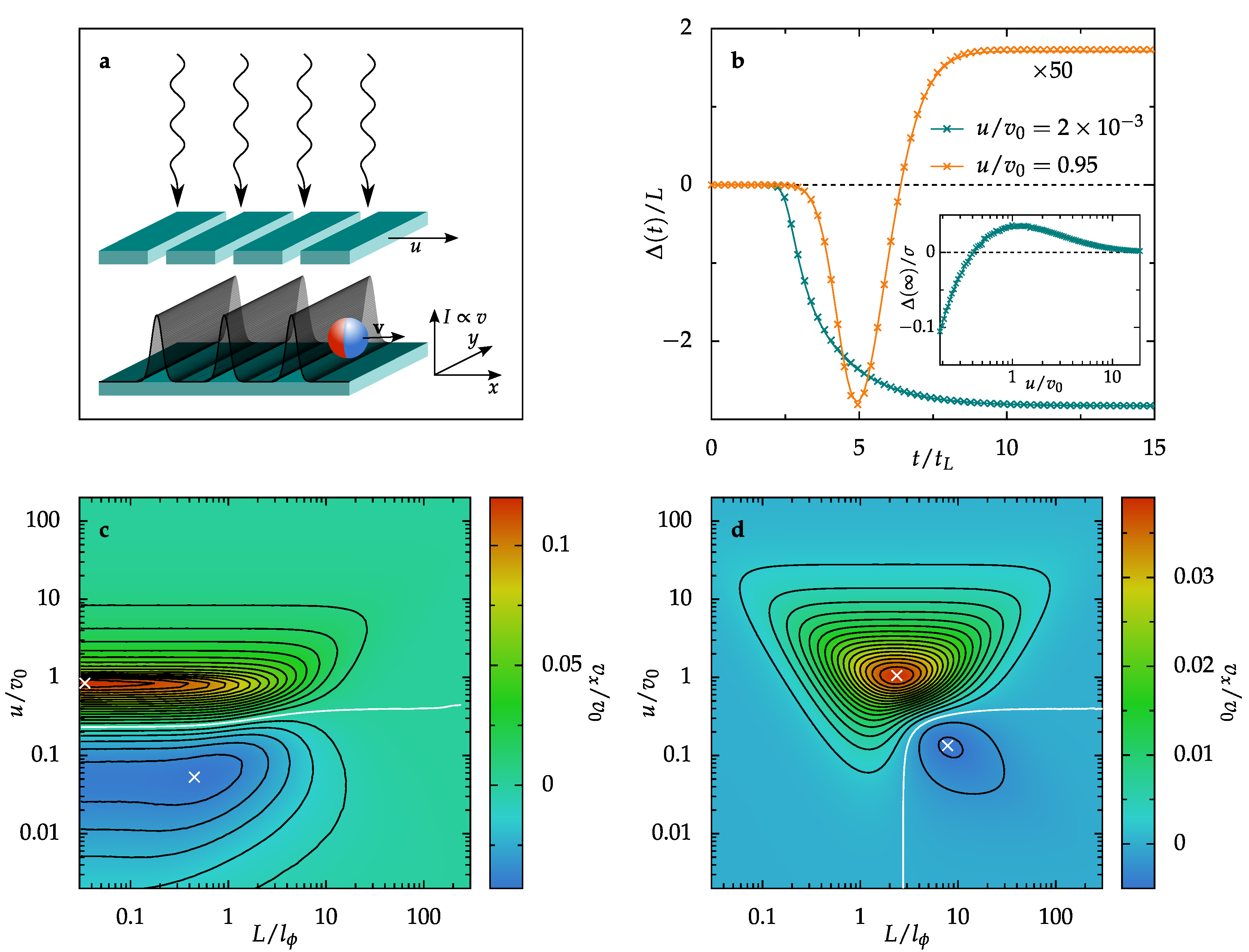
2. Artificial Microswimmers Activated by Traveling Wave Pulses
3. Results and Discussion
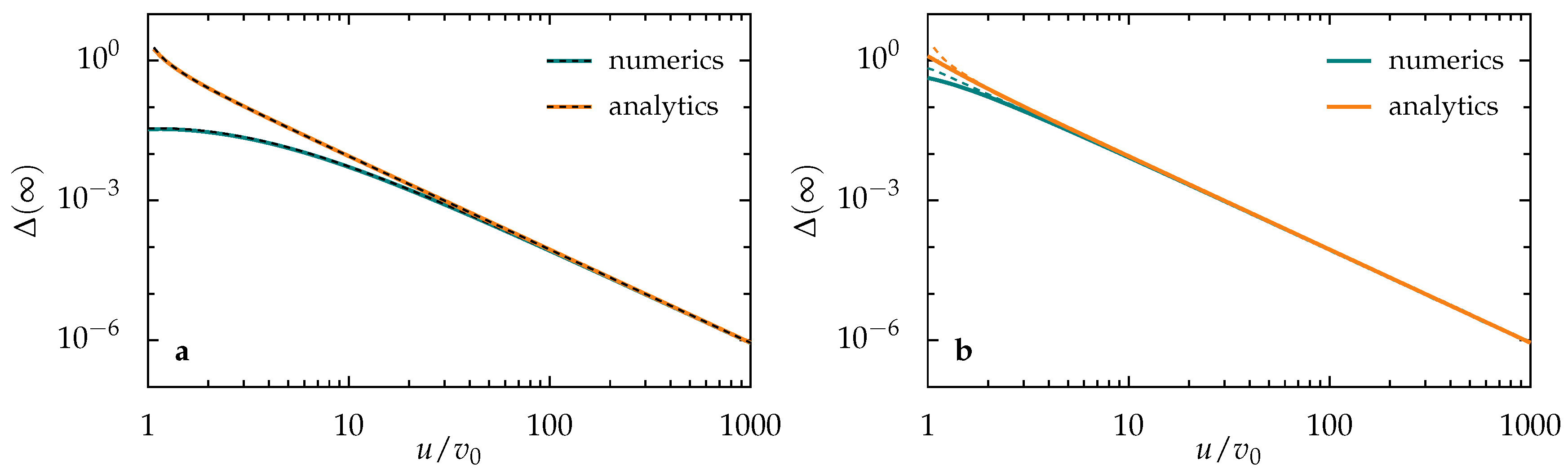
3.1. Diffusive Regime
3.1.1. Single Activation Pulse
3.1.2. Periodic Pulse Train
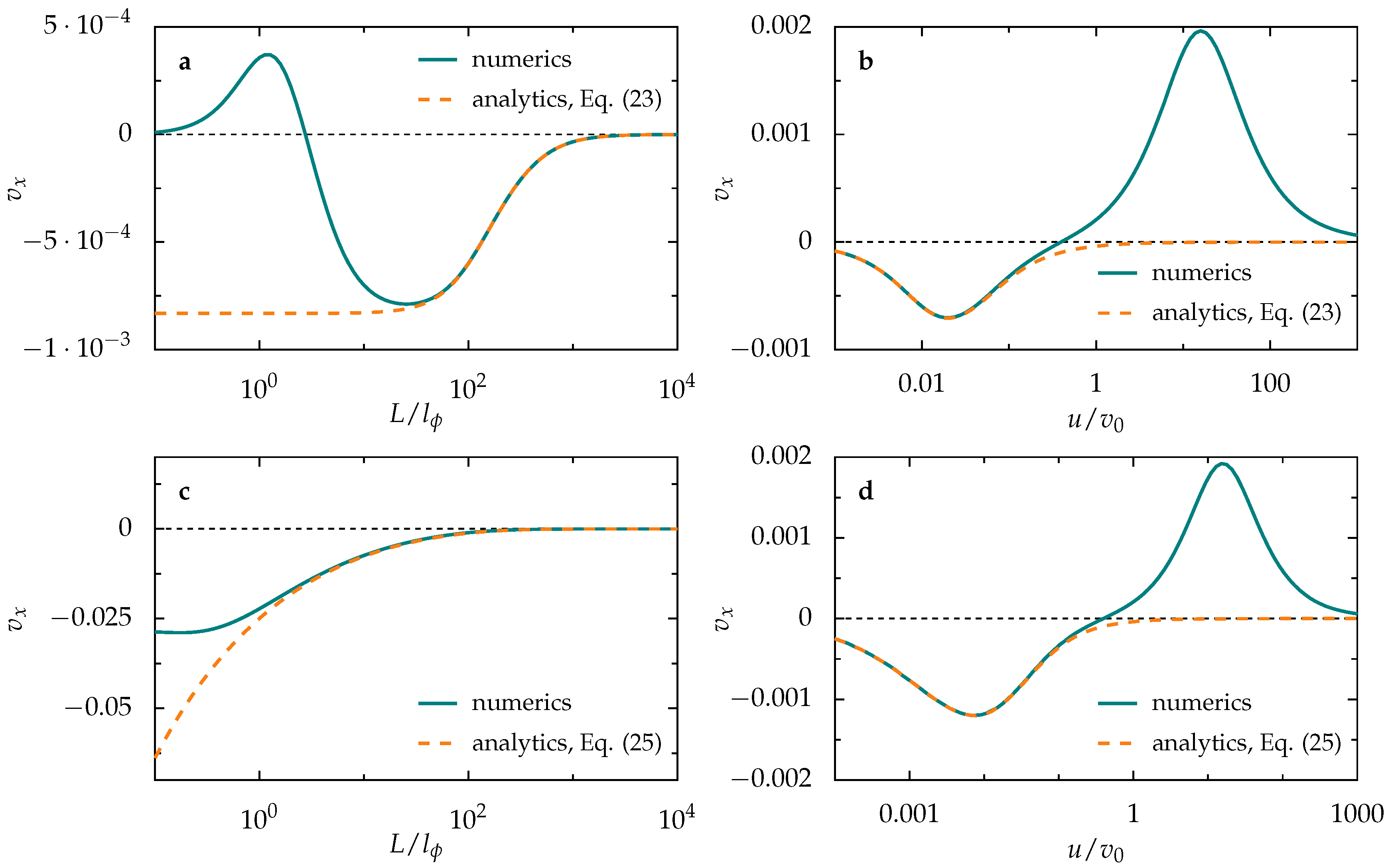
3.2. Ballistic Regime
3.2.1. Single Activation Pulse
3.2.2. Periodic Pulse Train
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
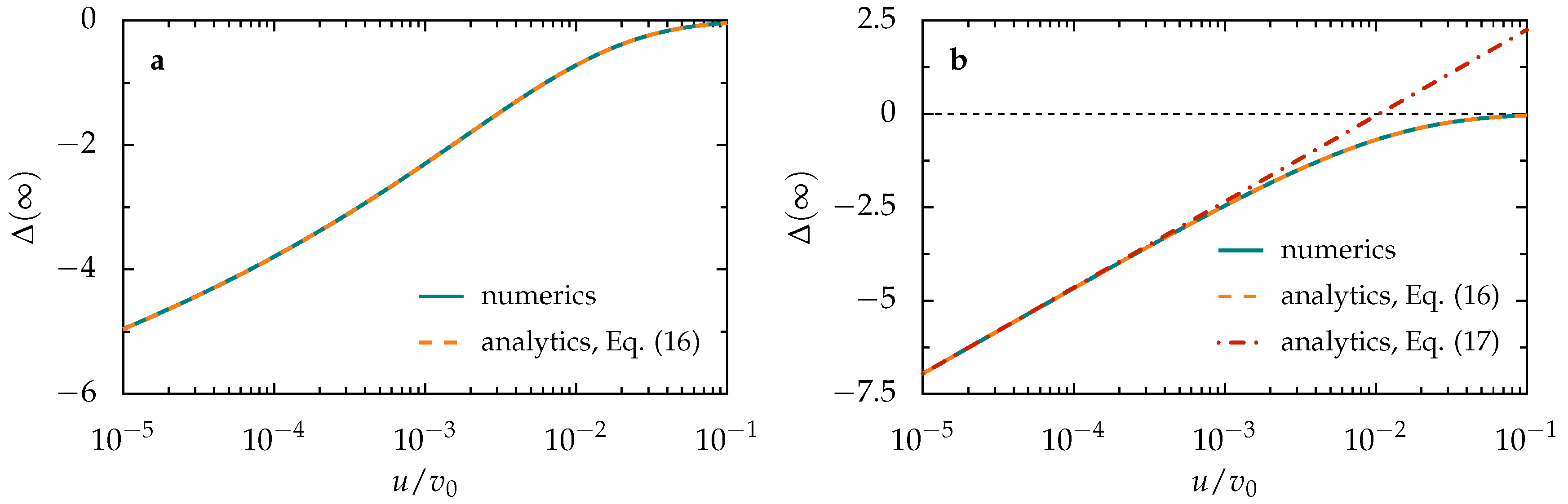
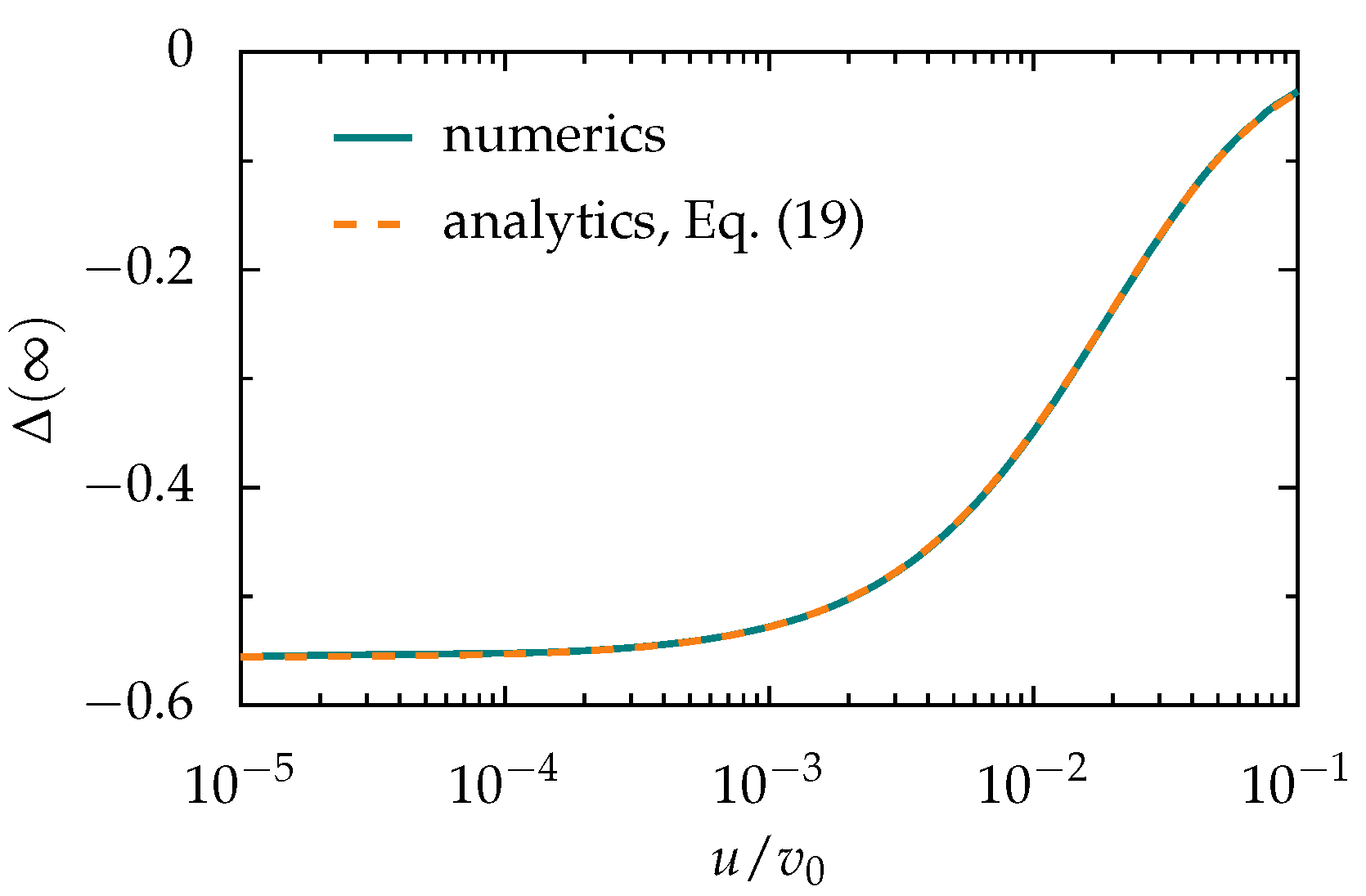
Appendix A. Tactic Shift Induced by a Soliton-Like Pulse
References
- Murray, J.D. Mathematical Biology, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 1993. [Google Scholar]
- Armitage, J.P. Bacterial tactic responses. Adv. Microb. Physiol. 1999, 41, 229–289. [Google Scholar] [PubMed]
- Adler, J. Chemotaxis in bacteria. Science 1966, 153, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Berg, H.C. E. coli in Motion; Springer: New York, NY, USA, 2004. [Google Scholar]
- Wadhams, G.H.; Armitage, J.P. Making sense of it all: Bacterial chemotaxis. Nat. Rev. Mol. Cell Biol. 2004, 5, 1024–1037. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, F. Brownian Agents and Active Particles; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Walther, A.; Müller, A.H.E. Janus particles: Synthesis, self-assembly, physical properties, and applications. Chem. Rev. 2013, 113, 5194–5261. [Google Scholar] [CrossRef] [PubMed]
- Elgeti, J.; Winkler, R.G.; Gompper, G. Physics of microswimmers—Single particle motion and collective behavior: A review. Rep. Prog. Phys. 2015, 78, 056601. [Google Scholar] [CrossRef] [PubMed]
- Bechinger, C.; Di Leonardo, R.; Löwen, H.; Reichhardt, C.; Volpe, G.; Volpe, G. Active particles in complex and crowded environments. Rev. Mod. Phys. 2016, 88, 045006. [Google Scholar] [CrossRef]
- Würger, A. Thermophoresis in colloidal suspensions driven by Marangoni forces. Phys. Rev. Lett. 2007, 98, 138301. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.R.; Yoshinaga, N.; Sano, M. Active motion of a Janus particle by self-thermophoresis in a defocused laser beam. Phys. Rev. Lett. 2010, 105, 268302. [Google Scholar] [CrossRef] [PubMed]
- Buttinoni, I.; Volpe, G.; Kümmel, F.; Volpe, G.; Bechinger, C. Active Brownian motion tunable by light. J. Phys. Condens. Matter 2012, 24, 284129. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Ripoll, M. Thermophoretically induced flow field around a colloidal particle. Soft Matter 2013, 9, 4661–4671. [Google Scholar] [CrossRef]
- Moran, J.L.; Wheat, P.M.; Posner, J.D. Locomotion of electrocatalytic nanomotors due to reaction induced charge autoelectrophoresis. Phys. Rev. E 2010, 81, 065302. [Google Scholar] [CrossRef] [PubMed]
- Ebbens, S.; Gregory, D.A.; Dunderdale, G.; Howse, J.R.; Ibrahim, Y.; Liverpool, T.B.; Golestanian, R. Electrokinetic effects in catalytic platinum-insulator Janus swimmers. EPL 2014, 106, 58003. [Google Scholar] [CrossRef]
- Golestanian, R.; Liverpool, T.B.; Ajdari, A. Propulsion of a molecular machine by asymmetric distribution of reaction products. Phys. Rev. Lett. 2005, 94, 220801. [Google Scholar] [CrossRef] [PubMed]
- Howse, J.R.; Jones, R.A.L.; Ryan, A.J.; Gough, T.; Vafabakhsh, R.; Golestanian, R. Self-motile colloidal particles: From directed propulsion to random walk. Phys. Rev. Lett. 2007, 99, 048102. [Google Scholar] [CrossRef] [PubMed]
- Volpe, G.; Buttinoni, I.; Vogt, D.; Kümmerer, H.J.; Bechinger, C. Microswimmers in patterned environments. Soft Matter 2011, 7, 8810–8815. [Google Scholar] [CrossRef]
- Hong, Y.; Blackman, N.M.K.; Kopp, N.D.; Sen, A.; Velegol, D. Chemotaxis of nonbiological colloidal rods. Phys. Rev. Lett. 2007, 99, 178103. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, P.K.; Li, Y.; Marchesoni, F.; Nori, F. Pseudochemotactic drifts of artificial microswimmers. Phys. Rev. E 2015, 92, 012114. [Google Scholar] [CrossRef] [PubMed]
- Ten Hagen, B.; Kümmel, F.; Wittkowski, R.; Takagi, D.; Löwen, H.; Bechinger, C. Gravitaxis of asymmetric self-propelled colloidal particles. Nat. Commun. 2014, 5, 4829. [Google Scholar] [CrossRef] [PubMed]
- Uspal, W.E.; Popescu, M.N.; Dietrich, S.; Tasinkevych, M. Rheotaxis of spherical active particles near a planar wall. Soft Matter 2015, 11, 6613–6632. [Google Scholar] [CrossRef] [PubMed]
- Lozano, C.; ten Hagen, B.; Löwen, H.; Bechinger, C. Phototaxis of synthetic microswimmers in optical landscapes. Nat. Commun. 2016, 7, 12828. [Google Scholar] [CrossRef] [PubMed]
- Armitage, J.P.; Lackie, J.M. (Eds.) Biology of the Chemotactic Response; Cambridge University Press: Cambridge, UK, 1990.
- Wessels, D.; Murray, J.; Soll, D.R. Behavior of Dictyostelium amoebae is regulated primarily by the temporal dynamic of the natural cAMP wave. Cell Motil. Cytoskelet. 1992, 23, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Stokes, G.G. On the theory of oscillatory waves. Trans. Camb. Philos. Soc. 1847, 8, 441–473. [Google Scholar]
- Van den Broeck, C. Stokes’ drift: An exact result. Europhys. Lett. 1999, 46, 1–5. [Google Scholar] [CrossRef]
- Höfer, T.; Maini, P.K.; Sherratt, J.A.; Chaplain, M.A.J.; Chauvet, P.; Metevier, D.; Montes, P.C.; Murray, J.D. A resolution of the chemotactic wave paradox. Appl. Math. Lett. 1994, 7, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, R.E. Traveling-wave chemotaxis. Phys. Rev. Lett. 1996, 77, 775–778. [Google Scholar] [CrossRef] [PubMed]
- Geiseler, A.; Hänggi, P.; Marchesoni, F.; Mulhern, C.; Savel’ev, S. Chemotaxis of artificial microswimmers in active density waves. Phys. Rev. E 2016, 94, 012613. [Google Scholar] [CrossRef] [PubMed]
- Serdyuk, I.N.; Zaccai, N.R.; Zaccai, J. Methods in Molecular Biophysics: Structure, Dynamics, Function; Cambridge University Press: New York, NY, USA, 2007. [Google Scholar]
- Ten Hagen, B.; van Teeffelen, S.; Löwen, H. Brownian motion of a self-propelled particle. J. Phys. Condens. Matter 2011, 23, 194119. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Velegol, D.; Chaturvedi, N.; Sen, A. Biomimetic behavior of synthetic particles: From microscopic randomness to macroscopic control. Phys. Chem. Chem. Phys. 2010, 12, 1423–1435. [Google Scholar] [CrossRef] [PubMed]
- Kapral, R.; Showalter, K. (Eds.) Chemical Waves and Patterns; Springer: Dordrecht, The Netherlands, 1995.
- Thakur, S.; Chen, J.X.; Kapral, R. Interaction of a chemically propelled nanomotor with a chemical wave. Angew. Chem. Int. Ed. 2011, 50, 10165–10169. [Google Scholar] [CrossRef] [PubMed]
- Löber, J.; Martens, S.; Engel, H. Shaping wave patterns in reaction-diffusion systems. Phys. Rev. E 2014, 90, 062911. [Google Scholar] [CrossRef] [PubMed]
- Navarro, R.M.; Fielding, S.M. Clustering and phase behaviour of attractive active particles with hydrodynamics. Soft Matter 2015, 11, 7525–7546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bickel, T.; Zecua, G.; Würger, A. Polarization of active Janus particles. Phys. Rev. E 2014, 89, 050303. [Google Scholar] [CrossRef] [PubMed]
- Geiseler, A.; Hänggi, P.; Marchesoni, F. Self-polarizing microswimmers in active density waves. Sci. Rep. 2017, 7, 41884. [Google Scholar] [CrossRef] [PubMed]
- Kalinay, P. Effective transport equations in quasi 1D systems. Eur. Phys. J. Spec. Top. 2014, 223, 3027–3043. [Google Scholar] [CrossRef]
- Geiseler, A.; Hänggi, P.; Schmid, G. Kramers escape of a self-propelled particle. Eur. Phys. J. B 2016, 89, 175. [Google Scholar] [CrossRef]
- Redner, S. A Guide to First-Passage Processes; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Goel, N.S.; Richter-Dyn, N. Stochastic Models in Biology; Academic Press: New York, NY, USA, 1974. [Google Scholar]
- Risken, H. The Fokker-Planck Equation, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 1989. [Google Scholar]
- Burada, P.S.; Schmid, G.; Reguera, D.; Rubí, J.M.; Hänggi, P. Biased diffusion in confined media: Test of the Fick-Jacobs approximation and validity criteria. Phys. Rev. E 2007, 75, 051111. [Google Scholar] [CrossRef] [PubMed]
- Burada, P.S.; Schmid, G.; Hänggi, P. Entropic transport: A test bed for the Fick-Jacobs approximation. Philos. Trans. R. Soc. A 2009, 367, 3157–3171. [Google Scholar] [CrossRef] [PubMed]
- Abramowitz, M.; Stegun, I.A. (Eds.) Handbook of Mathematical Functions, 10th ed.U.S. Government Printing Office: Washington, DC, USA, 1972.
- Mijalkov, M.; McDaniel, A.; Wehr, J.; Volpe, G. Engineering sensorial delay to control phototaxis and emergent collective behaviors. Phys. Rev. X 2016, 6, 011008. [Google Scholar] [CrossRef]
- Prudnikov, A.P.; Brychkov, Y.A.; Marichev, O.I. Integrals and Series. Volume 2: Special Functions; Gordon and Breach: New York, NY, USA, 1992. [Google Scholar]
- Dingle, R.B. Asymptotic expansions and converging factors. II. Error, Dawson, Fresnel, exponential, sine and cosine, and similar integrals. Proc. R. Soc. A 1958, 244, 476–483. [Google Scholar] [CrossRef]
- Gradshteyn, I.S.; Ryzhik, I.M. Table of Integrals, Series, and Products, 7th ed.; Academic Press: Amsterdam, The Netherlands, 2007. [Google Scholar]






© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geiseler, A.; Hänggi, P.; Marchesoni, F. Taxis of Artificial Swimmers in a Spatio-Temporally Modulated Activation Medium. Entropy 2017, 19, 97. https://0-doi-org.brum.beds.ac.uk/10.3390/e19030097
Geiseler A, Hänggi P, Marchesoni F. Taxis of Artificial Swimmers in a Spatio-Temporally Modulated Activation Medium. Entropy. 2017; 19(3):97. https://0-doi-org.brum.beds.ac.uk/10.3390/e19030097
Chicago/Turabian StyleGeiseler, Alexander, Peter Hänggi, and Fabio Marchesoni. 2017. "Taxis of Artificial Swimmers in a Spatio-Temporally Modulated Activation Medium" Entropy 19, no. 3: 97. https://0-doi-org.brum.beds.ac.uk/10.3390/e19030097