Entropy-Based Estimation of Event-Related De/Synchronization in Motor Imagery Using Vector-Quantized Patterns


Abstract
:1. Introduction
2. Materials and Methods
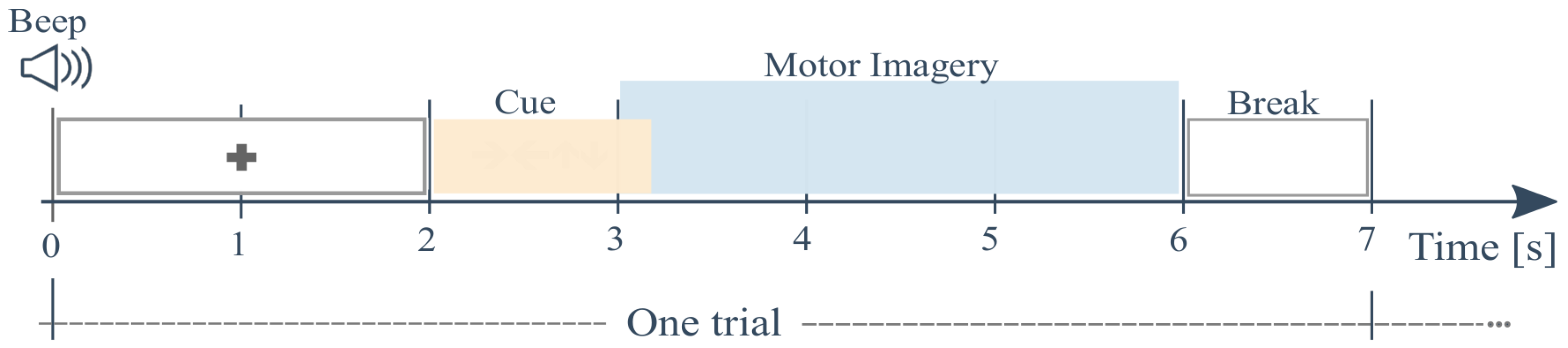
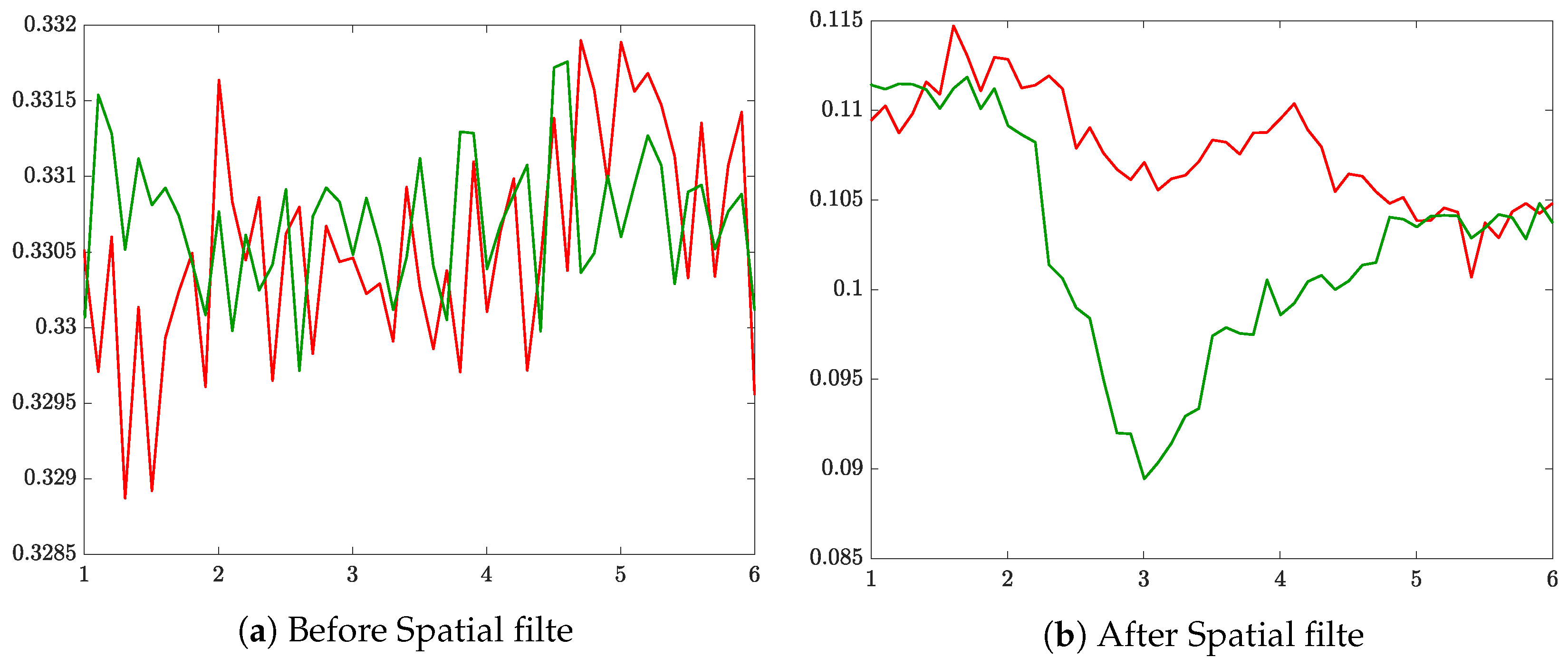
2.1. EEG Recordings and Preprocessing
2.2. Complexity-Based Estimation of Time-Evolving Event-Related De/Synchronization (ERD/S)
2.3. Symbolic Spaces Using Quantized Stochastic Patterns
| Algorithm 1 Building of VQ stochastic patterns. |
|
3. Experiments and Results
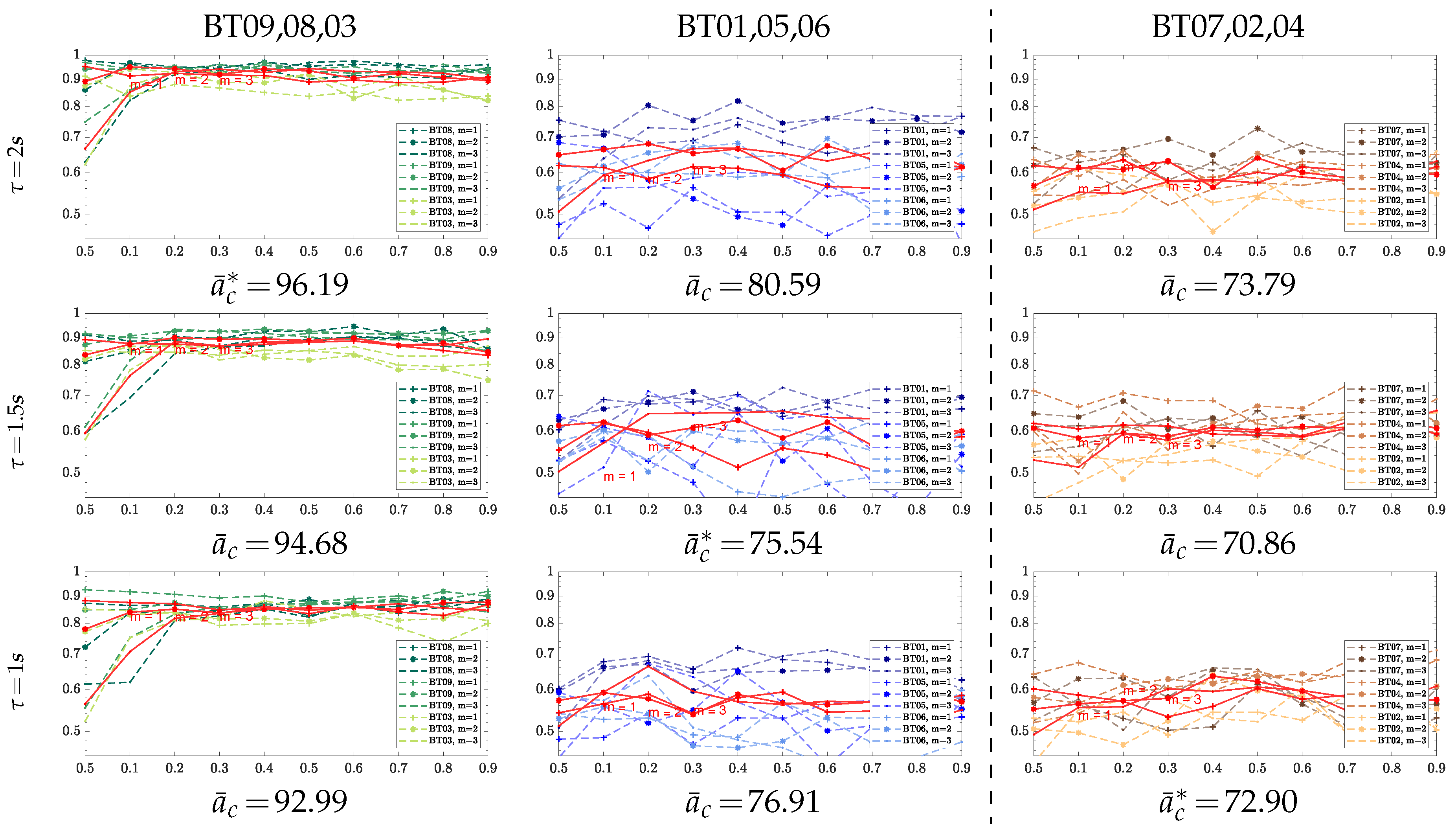
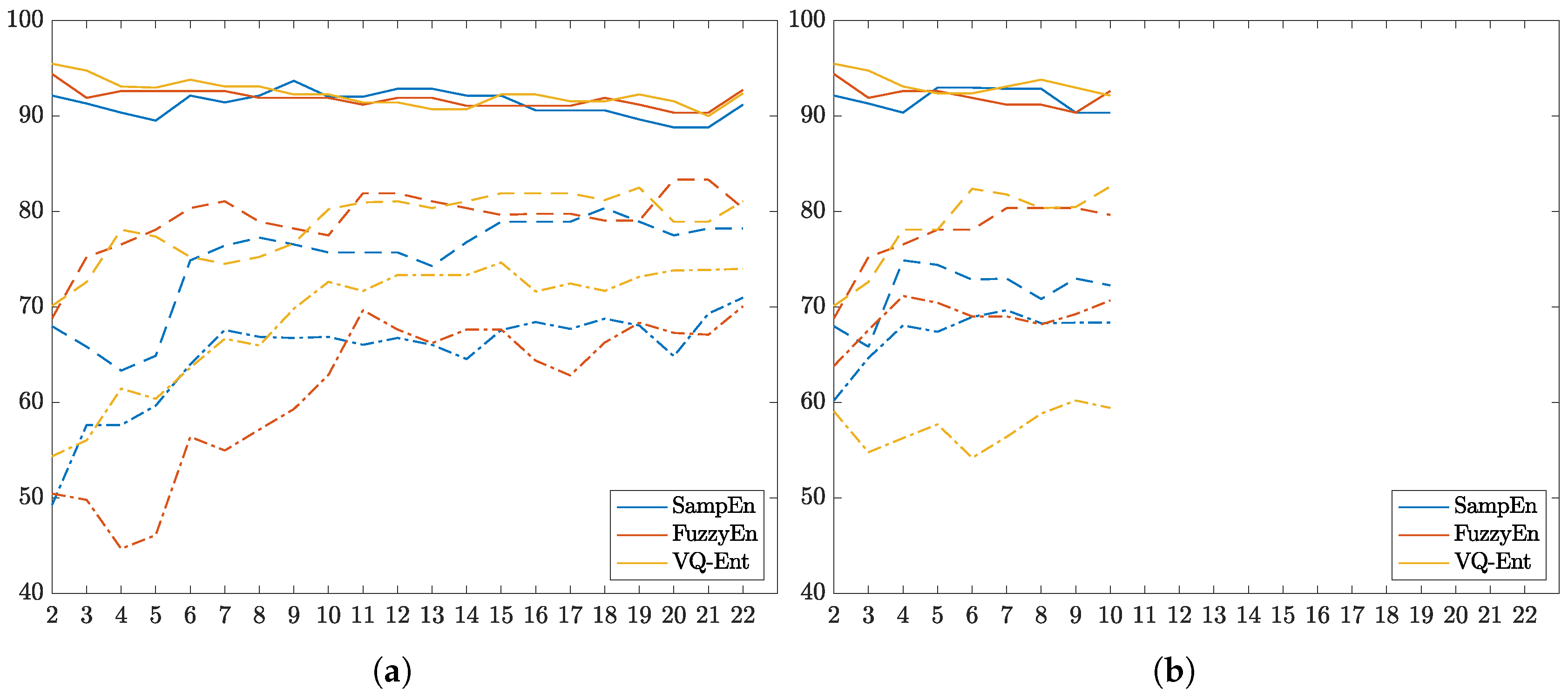
3.1. Parameter Tuning of Compared Entropy-Based Estimators
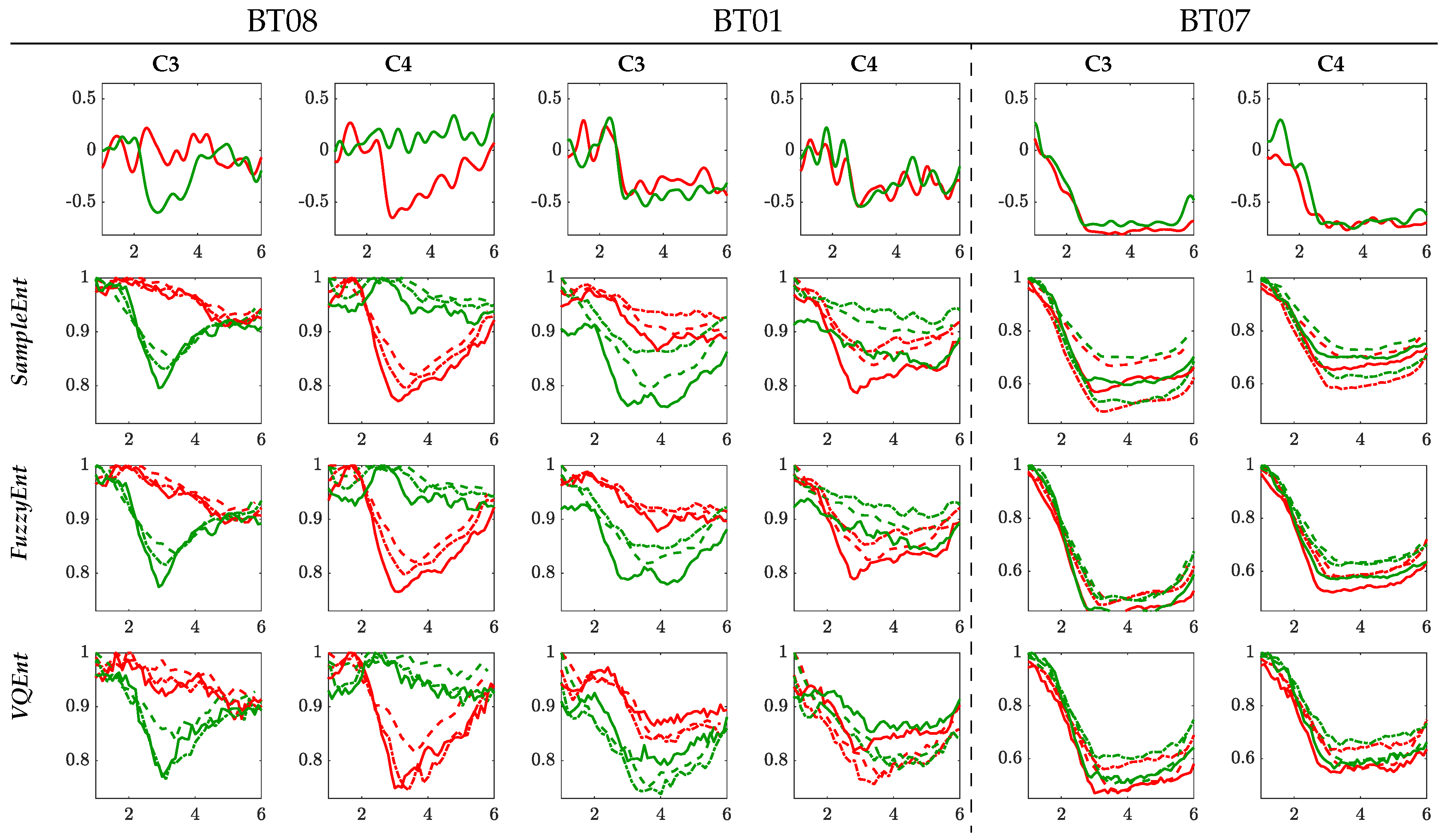
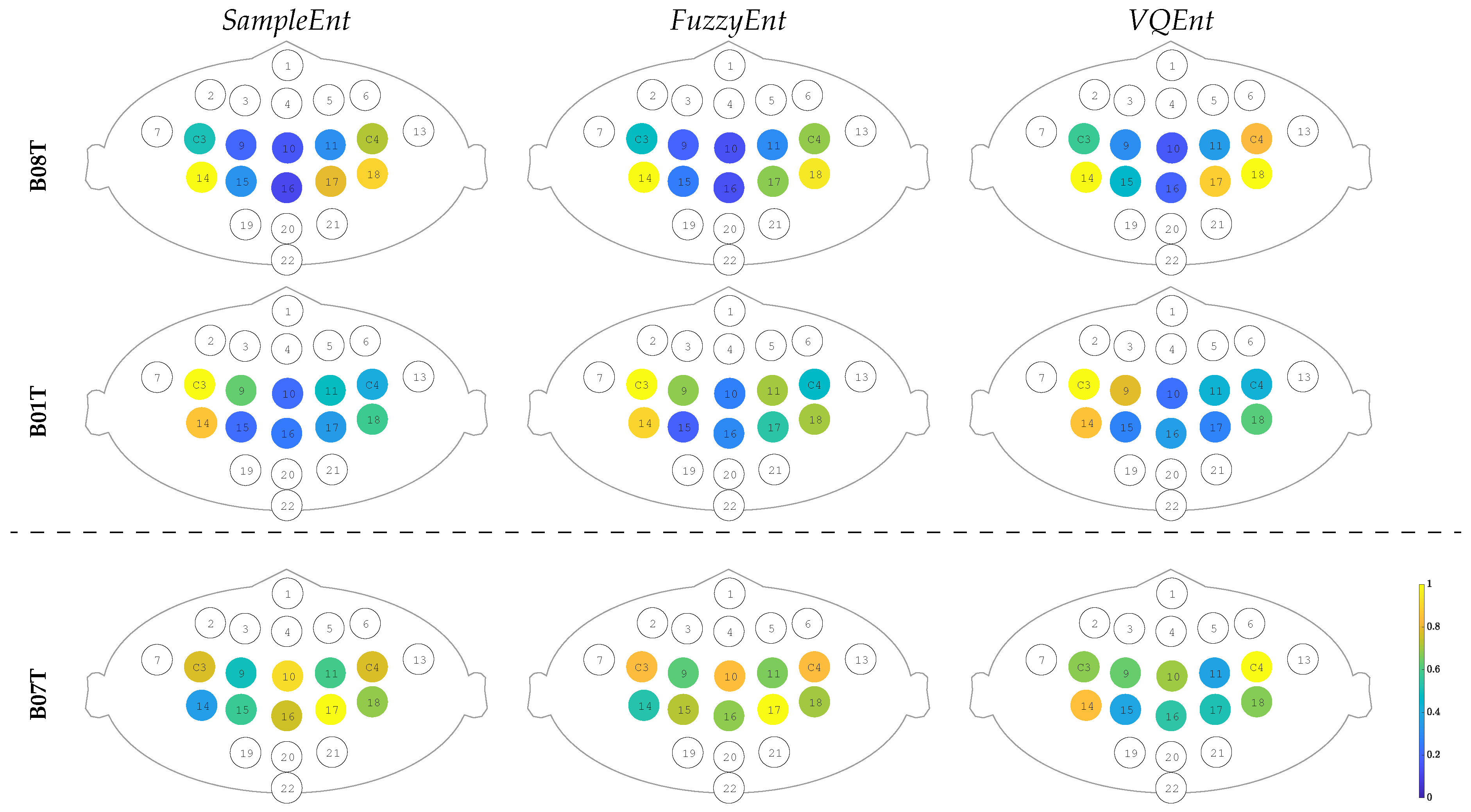
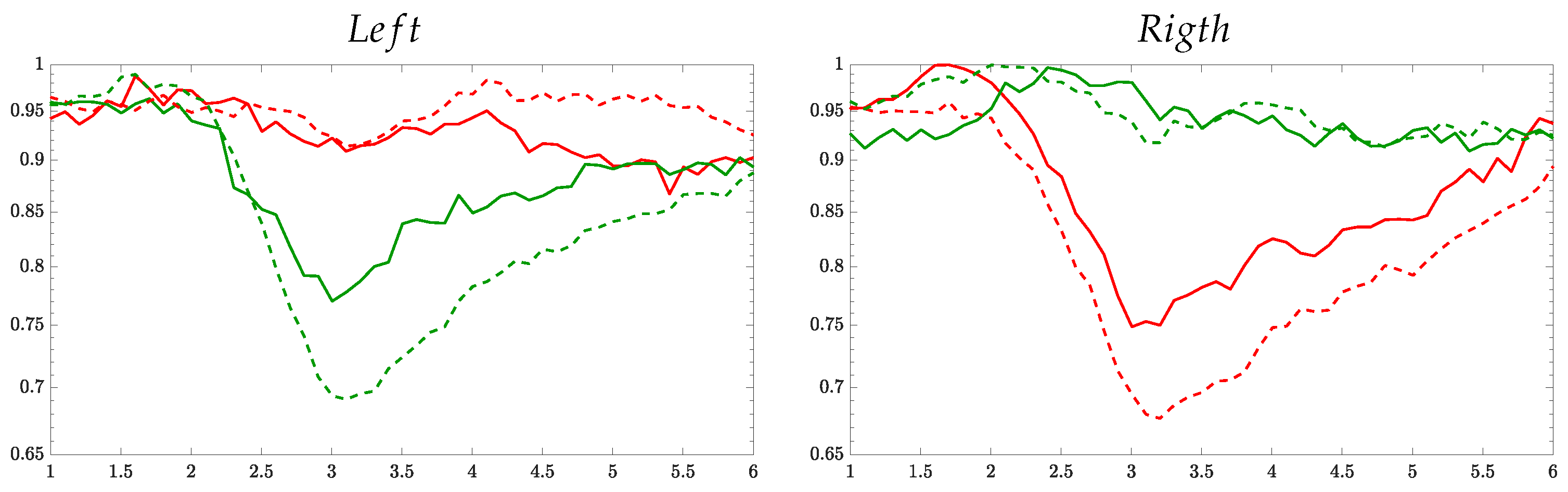
3.2. Interpretability of Time-courses Estimated for Event-Related De/Synchronization
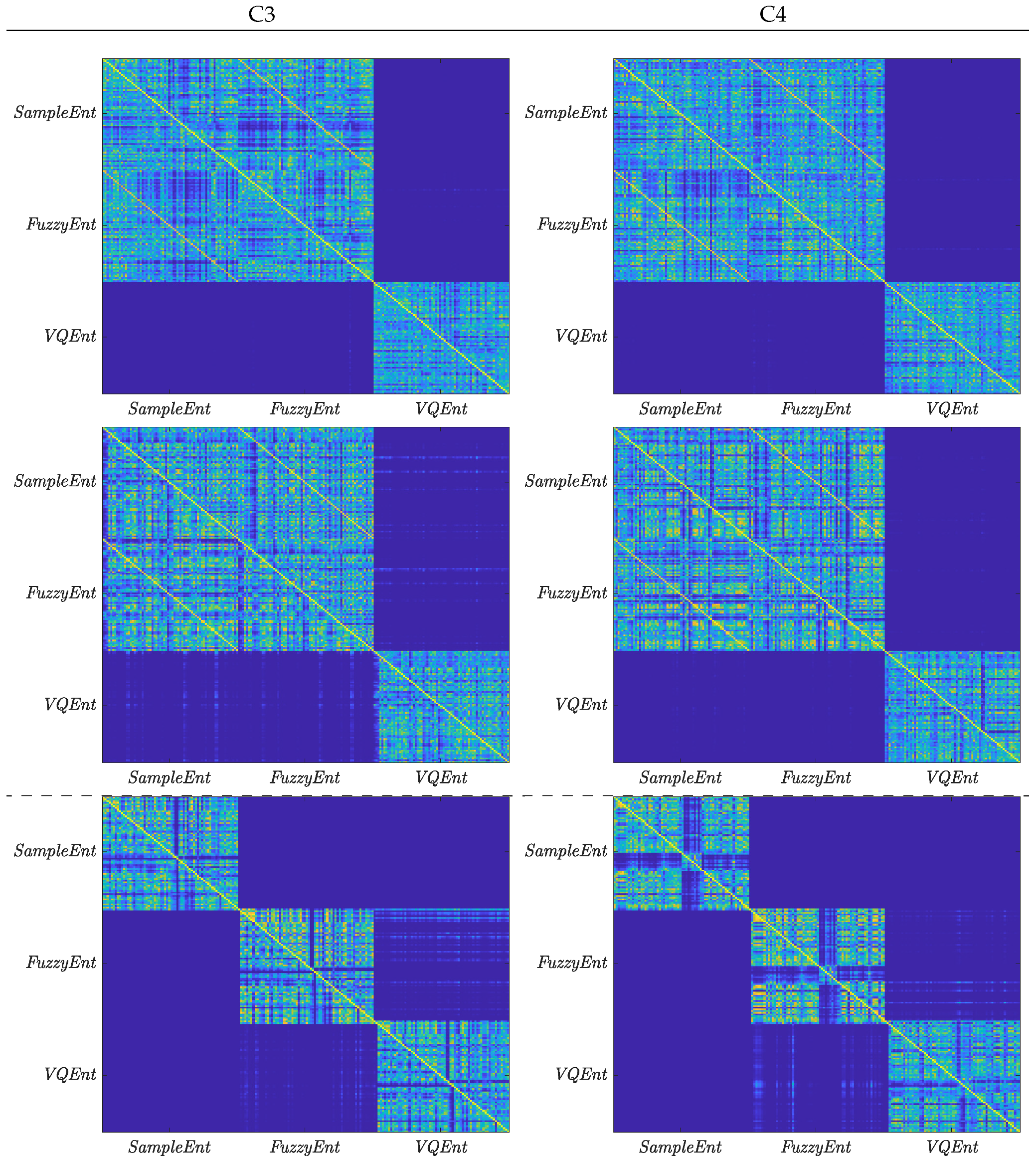
3.3. Statistical Analysis
3.4. Contribution of Sensorimotor Area to Distinguishing between MI Tasks
4. Discussion and Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Opsommer, E.; Chevalley, O.; Korogod, N. Motor imagery for pain and motor function after spinal cord injury: A systematic review. Spinal Cord 2019, 58, 262–274. [Google Scholar] [CrossRef] [PubMed]
- Machado, T.; Carregosa, A.; Santos, M.; da Silva, N.; Melo, M. Efficacy of motor imagery additional to motor-based therapy in the recovery of motor function of the upper limb in post-stroke individuals: A systematic review. Top. Stroke Rehabil. 2019, 26, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Guillot, A.; Debarnot, U. Benefits of Motor Imagery for Human Space Flight: A Brief Review of Current Knowledge and Future Applications. Front. Physiol. 2019, 10, 396. [Google Scholar] [CrossRef]
- Barhoun, P.; Fuelscher, I.; Kothe, E.; He, J.; Youssef, G.; Enticott, P.; Williams, J.; Hyde, C. Motor imagery in children with DCD: A systematic and meta-analytic review of hand-rotation task performance. Neurosci. Biobehav. Rev. 2019, 99, 282–297. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, V.; Watts, N.; Chani, Y.; Keogh, J. Motor imagery training improves balance and mobility outcomes in older adults: A systematic review. J. Physiother. 2019, 65, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Frau-Meigs, D. Media Education. A Kit for Teachers, Students, Parents and Professionals; UNESCO: Paris, France, 2007. [Google Scholar]
- Balamurugan, B.; Mullai, M.; Soundararajan, S.; Selvakanmani, S.; Arun, D. Brain–computer interface for assessment of mental efforts in e-learning using the nonmarkovian queueing model. Comput. Appl. Eng. Educ. 2020. [Google Scholar] [CrossRef]
- Durka, P.; Ircha, D.; Neuper, C.; Pfurtscheller, G. Time-frequency microstructure of event-related electro-encephalogram desynchronisation and synchronisation. Med Biol. Eng. Comput. 2001, 39, 315–321. [Google Scholar] [CrossRef]
- Grandchamp, R.; Delorme, A. Single-Trial Normalization for Event-Related Spectral Decomposition Reduces Sensitivity to Noisy Trials. Front. Psychol. 2011, 2, 236. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; He, B. Brain–computer interfaces using sensorimotor rhythms: Current state and future perspectives. IEEE Trans. Biomed. Eng. 2014, 61, 1425–1435. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Li, W.; Li, X.; Ma, W.; Dang, X. Motor imagery EEG recognition based on conditional optimization empirical mode decomposition and multi-scale convolutional neural network. Expert Syst. Appl. 2020, 149, 113285. [Google Scholar] [CrossRef]
- Gao, L.; Wang, J.; Chen, L. Event-related desynchronization and synchronization quantification in motor-related EEG by Kolmogorov entropy. J. Neural Eng. 2013, 10, 036023. [Google Scholar] [CrossRef]
- Azami, H.; Li, P.; Arnold, S.; Escudero, J.; Humeau-Heurtier, A. Fuzzy Entropy Metrics for the Analysis of Biomedical Signals: Assessment and Comparison. IEEE Access 2019, 7, 104833–104847. [Google Scholar] [CrossRef]
- Wei-Yen, H. Assembling A Multi-Feature EEG Classifier for Left-Right Motor Imagery Data Using Wavelet-Based Fuzzy Approximate Entropy for Improved Accuracy. Int. J. Neural Syst. 2015, 25, 1550037. [Google Scholar]
- Shunfei, C.; Zhizeng, L.; Haitao, G. An entropy fusion method for feature extraction of EEG. Neural Comput. Appl. 2016, 29, 857–863. [Google Scholar]
- Pattnaik, K.; Sarraf, J. Brain Computer Interface issues on hand movement. Comput. Inf. Sci. 2018, 30, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.; Chung, W. Frequency-Optimized Local Region Common Spatial Pattern Approach for Motor Imagery Classification. IEEE Trans. Neural Syst. Rehabil. Eng. 2019, 27, 1378–1388. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Ahmed, K.; Mostafa, R.; Hadjileontiadis, L.; Khandoker, A. Evidence of Variabilities in EEG Dynamics During Motor Imagery-Based Multiclass Brain-Computer Interface. IEEE Trans. Neural Syst. Rehabil. Eng. 2018, 26, 371–382. [Google Scholar] [CrossRef]
- Sri, P.; Yashasvi, K.; Anjum, A.; Bhattacharyya, A.; Pachori, R. Development of an Effective Computing Framework for Classification of Motor Imagery EEG Signals for Brain–Computer Interface. In Advances in Computational Intelligence Techniques; Jain, S., Sood, M., Paul, S., Eds.; Springer: Singapore, 2020; pp. 17–35. [Google Scholar]
- Rostaghi, M.; Azami, H. Dispersion Entropy: A Measure for Time-Series Analysis. IEEE Signal Process. Lett. 2016, 23, 610–614. [Google Scholar] [CrossRef]
- Kuntzelman, K.; Rhodes, L.; Harrington, L.; Miskovic, V. A practical comparison of algorithms for the measurement of multiscale entropy in neural time series data. Brain Cogn. 2018, 123, 126–135. [Google Scholar] [CrossRef]
- Li, Y.; Gao, X.; Wang, L. Reverse Dispersion Entropy: A New Complexity Measure for Sensor Signal. Sensors 2019, 19, 5203. [Google Scholar] [CrossRef] [Green Version]
- Kafantaris, E.; Piper, I.; Lo, M.; Escudero, J. Augmentation of Dispersion Entropy for Handling Missing and Outlier Samples in Physiological Signal Monitoring. Entropy 2020, 22, 319. [Google Scholar] [CrossRef] [Green Version]
- Pitsik, E.; Frolov, N.; Hauke, K.; Grubov, V.; Maksimenko, V.; Kurths, J.; Hramov, A. Motor execution reduces EEG signals complexity: Recurrence quantification analysis study<? A3B2 show [feature]?>. Chaos Interdiscip. J. Nonlinear Sci. 2020, 30, 023111. [Google Scholar]
- Miao, M.; Zeng, H.; Wang, A.; Zhao, C.; Liu, F. Discriminative spatial-frequency-temporal feature extraction and classification of motor imagery EEG: An sparse regression and Weighted Naïve Bayesian Classifier-based approach. J. Neurosci. Methods 2017, 278, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; McFarland, D.; Wolpaw, J. Adaptive Laplacian filtering for sensorimotor rhythm-based brain–computer interfaces. J. Neural Eng. 2012, 10, 016002. [Google Scholar] [CrossRef] [Green Version]
- Sannelli, C.; Vidaurre, C.; Müller, K.; Blankertz, B. Common spatial pattern patches-an optimized filter ensemble for adaptive brain-computer interfaces. In Proceedings of the 2010 Annual International Conference of the IEEE Engineering in Medicine and Biology, Buenos Aires, Argentina, 31 August–4 September 2010; pp. 4351–4354. [Google Scholar]
- Delgado-Bonal, A.; Marshak, A. Approximate Entropy and Sample Entropy: A Comprehensive Tutorial. Entropy 2019, 21, 541. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.; Nguyen, T. Entropy-Constrained Maximizing Mutual Information Quantization. arXiv 2020, arXiv:abs/2001.01830. [Google Scholar]
- Zhao, S.; Chen, B.; Zhu, P.; Príncipe, J. Fixed budget quantized kernel least-mean-square algorithm. Signal Process. 2013, 93, 2759–2770. [Google Scholar] [CrossRef]
- Cardenas-Pena, D.; Tobar-Rodriguez, A.; Castellanos-Dominguez, G. Adaptive Bayesian label fusion using kernel-based similarity metrics in hippocampus segmentation. J. Med. Imaging 2019, 6, 1–8. [Google Scholar] [CrossRef]
- Zhang, Y.; Nam, C.S.; Zhou, G.; Jin, J.; Wang, X.; Cichocki, A. Temporally constrained sparse group spatial patterns for motor imagery BCI. IEEE Trans. Cybern. 2018, 49, 3322–3332. [Google Scholar] [CrossRef]
- Latchoumane, C.; Chung, D.; Kim, S.; Jeong, J. Segmentation and Characterization of EEG During Mental tasks using Dynamical Nonstationarity. In Proceedings of the Computational Intelligence in Medical and Healthcare (CIMED 2007), Plymouth, UK, 25–27 July 2007. [Google Scholar]
- Ma, M.; Guo, L.; Su, K.; Liang, D. Classification of motor imagery EEG signals based on wavelet transform and sample entropy. In Proceedings of the 2017 IEEE 2nd Advanced Information Technology, Electronic and Automation Control Conference (IAEAC), Chongqing, China, 25–26 March 2017; pp. 905–910. [Google Scholar]
- Ahn, M.; Cho, H.n.; Ahn, S.; Jun, S. High theta and low alpha powers may be indicative of BCI-illiteracy in motor imagery. PLoS ONE 2013, 8, e80886. [Google Scholar] [CrossRef] [Green Version]
- Collazos-Huertas, D.; Caicedo-Acosta, J.; Castaño-Duque, G.; Acosta-Medina, C. Enhanced Multiple Instance Representation Using Time-Frequency Atoms in Motor Imagery Classification. Front. Neurosci. 2020, 14, 155. [Google Scholar] [CrossRef] [PubMed]
- Pfurtscheller, G. EEG event-related desynchronization (ERD) and synchronization (ERS). Electroencephalogr. Clin. Neurophysiol. 1997, 1, 26. [Google Scholar] [CrossRef]
- Ahn, M.; Jun, S.C. Performance variation in motor imagery brain–computer interface: A brief review. J. Neurosci. Methods 2015, 243, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Velasquez-Martinez, L.; Arteaga, F.; Castellanos-Dominguez, G. Subject-Oriented Dynamic Characterization of Motor Imagery Tasks Using Complexity Analysis. In Proceedings of the International Conference on Brain Informatics, Haikou, China, 13–15 December 2019; Springer: Cham, Switzerland, 2019; pp. 21–28. [Google Scholar]
- Maris, E.; Oostenveld, R. Nonparametric statistical testing of EEG-and MEG-data. J. Neurosci. Methods 2007, 164, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Zich, C.; Debener, S.; Kranczioch, C.; Bleichner, M.G.; Gutberlet, I.; De Vos, M. Real-time EEG feedback during simultaneous EEG–fMRI identifies the cortical signature of motor imagery. Neuroimage 2015, 114, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Ahn, M.; Ahn, S.; Hong, J.H.; Cho, H.; Kim, K.; Kim, B.S.; Chang, J.W.; Jun, S.C. Gamma band activity associated with BCI performance: Simultaneous MEG/EEG study. Front. Hum. Neurosci. 2013, 7, 848. [Google Scholar] [CrossRef] [Green Version]
- Giusti, R.; Batista, G.E. An empirical comparison of dissimilarity measures for time series classification. In Proceedings of the 2013 Brazilian Conference on Intelligent Systems, Fortaleza, Brazil, 19–24 October 2013; pp. 82–88. [Google Scholar]
- Xygonakis, I.; Athanasiou, A.; Pandria, N.; Kugiumtzis, D.; Bamidis, P.D. Decoding motor imagery through common spatial pattern filters at the EEG source space. Comput. Intell. Neurosci. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, M.; Iso, N.; Fujiwara, K.; Moriuchi, T.; Tanaka, G.; Honda, S.; Matsuda, D.; Higashi, T. Cerebral haemodynamics during motor imagery of self-feeding with chopsticks: Differences between dominant and non-dominant hand. Somatosens. Mot. Res. 2020, 37, 6–13. [Google Scholar] [CrossRef]
- Allison, B.Z.; Neuper, C. Could anyone use a BCI? In Brain-Computer Interfaces; Springer: London, UK, 2010; pp. 35–54. [Google Scholar]
- Tian, Y.; Xu, W.; Yang, L. Cortical classification with rhythm entropy for error processing in cocktail party environment based on scalp EEG recording. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Blankertz, B.; Tomioka, R.; Lemm, S.; Kawanabe, M.; Muller, K.R. Optimizing spatial filters for robust EEG single-trial analysis. IEEE Signal Process. Mag. 2007, 25, 41–56. [Google Scholar] [CrossRef]
- Kaffashi, F.; Foglyano, R.; Wilson, C.; Loparo, K. The effect of time delay on Approximate and Sample Entropy calculations. Phys. D Nonlinear Phenom. 2008, 237, 3069–3074. [Google Scholar] [CrossRef]
- Thompson, M.C. Critiquing the concept of BCI illiteracy. Sci. Eng. Ethics 2019, 25, 1217–1233. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | SampleEnt | FuzzyEnt | VQEnt | ||||||
|---|---|---|---|---|---|---|---|---|---|
| [s] | 2 | 1.5 | 1 * | 2 | 1.5 * | 1 | 2 | 1 * | |
| B09T | 94.9 ± 8.3 | 95.7 ± 6.8 | 94.1 ± 5.21 | 94.2 ± 7.1 | 95.1 ± 7.2 | 95.0 ± 5.4 | 96.8 ± 5.2 | 96.6 ± 6.7 | 97.4 ± 4.0 |
| B08T | 94.4 ± 8.9 | 94.3 ± 8.3 | 92.0 ± 10.0 | 96.9 ± 3.8 | 96.1 ± 5.4 | 92.7 ± 8.7 | 97.6 ± 3.6 | 95.4 ± 6.2 | 92.4 ± 3.2 |
| B03T | 94.9 ± 3.4 | 91.3 ± 7.0 | 88.2 ± 6.4 | 89.7 ± 5.9 | 88.9 ± 6.8 | 86.1 ± 6.5 | 94.1 ± 5.4 | 92.0 ± 6.1 | 89.2 ± 8.6 |
| B01T | 81.2 ± 12.4 | 80.2 ± 14.7 | 78.2 ± 11.1 | 79.6 ± 11.1 | 81.1 ± 8.7 | 80.4 ± 11.6 | 81.9 ± 7.9 | 80.4 ± 9.2 | 81.1 ± 7.5 |
| B05T | 71.7 ± 11.4 | 73.7 ± 12.9 | 74.8 ± 12.4 | 73.0 ± 10.7 | 79.3 ± 6.9 | 75.5 ± 8.6 | 68.4 ± 10.2 | 71.4 ± 15.2 | 72.1 ± 10.3 |
| B06T | 70.3 ± 16.3 | 75.4 ± 12.8 | 72.9 ± 10.9 | 69.5 ± 11.2 | 73.9 ± 13.8 | 75.9 ± 6.2 | 69.6 ± 14.1 | 74.8 ± 12.2 | 77.5 ± 7.3 |
| B07T | 66.9 ± 11.9 | 67.7 ± 14.7 | 71.0 ± 10.7 | 67.8 ± 14.9 | 70.0 ± 14.3 | 70.1 ± 13.1 | 72.7 ± 13.8 | 71.9 ± 16.5 | 74 ± 10.1 |
| B02T | 59.4 ± 13.8 | 61.3 ± 8.7 | 68.5 ± 11.7 | 56.6 ± 7.7 | 60.9 ± 10.9 | 67.5 ± 16.8 | 65.7 ± 12.2 | 67.5 ± 11.0 | 73.5 ± 11 |
| B04T | 60.5 ± 11.8 | 62.1 ± 15.5 | 62.9 ± 11.0 | 58.1 ± 10.9 | 64.2 ± 6.5 | 65.1 ± 8.9 | 65.8 ± 14.3 | 73.2 ± 12.0 | 71.2 ± 10.7 |
| Mean | 77.1 ± 10.9 | 78.0 ± 11.3 | 78.0 ± 9.9 | 76.2 ± 9.3 | 78.8 ± 9.0 | 78.7 ± 9.5 | 79.2 ± 9.7 | 80.4 ± 10.6 | 80.9 ± 8.1 |
| SampleEnt | FuzzyEnt | VQEnt | |||||
|---|---|---|---|---|---|---|---|
| # | |||||||
| B09T | 2 | 0.9 | 2 | 0.3 | 2 | 0.3 | 83 |
| B08T | 1 | 0.9 | 1 | 0.3 | 3 | 0.6 | 47 |
| B03T | 3 | 0.9 | 3 | 0.6 | 2 | 0.1 | 116 |
| B01T | 1 | 0.8 | 1 | 0.2 | 2 | 0.2 | 86 |
| B05T | 1 | 0.8 | 3 | 0.6 | 2 | 0.1 | 110 |
| B06T | 3 | 0.9 | 1 | 0.9 | 2 | 0.6 | 47 |
| B07T | 1 | 0.5 | 1 | 0.6 | 3 | 0.9 | 32 |
| B02T | 2 | 0.8 | 1 | 0.05 | 2 | 0.3 | 72 |
| B04T | 2 | 0.6 | 1 | 0.5 | 3 | 0.9 | 30 |
| Median | 1 | 0.8 | 2 | 0.5 | 2 | 0.3 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velasquez-Martinez, L.; Caicedo-Acosta, J.; Castellanos-Dominguez, G. Entropy-Based Estimation of Event-Related De/Synchronization in Motor Imagery Using Vector-Quantized Patterns. Entropy 2020, 22, 703. https://0-doi-org.brum.beds.ac.uk/10.3390/e22060703
Velasquez-Martinez L, Caicedo-Acosta J, Castellanos-Dominguez G. Entropy-Based Estimation of Event-Related De/Synchronization in Motor Imagery Using Vector-Quantized Patterns. Entropy. 2020; 22(6):703. https://0-doi-org.brum.beds.ac.uk/10.3390/e22060703
Chicago/Turabian StyleVelasquez-Martinez, Luisa, Julián Caicedo-Acosta, and Germán Castellanos-Dominguez. 2020. "Entropy-Based Estimation of Event-Related De/Synchronization in Motor Imagery Using Vector-Quantized Patterns" Entropy 22, no. 6: 703. https://0-doi-org.brum.beds.ac.uk/10.3390/e22060703