The Effect of Monoterpenes on Swarming Differentiation and Haemolysin Activity in Proteus mirabilis

Abstract
:Introduction
Results and Discussion

{kind=link}
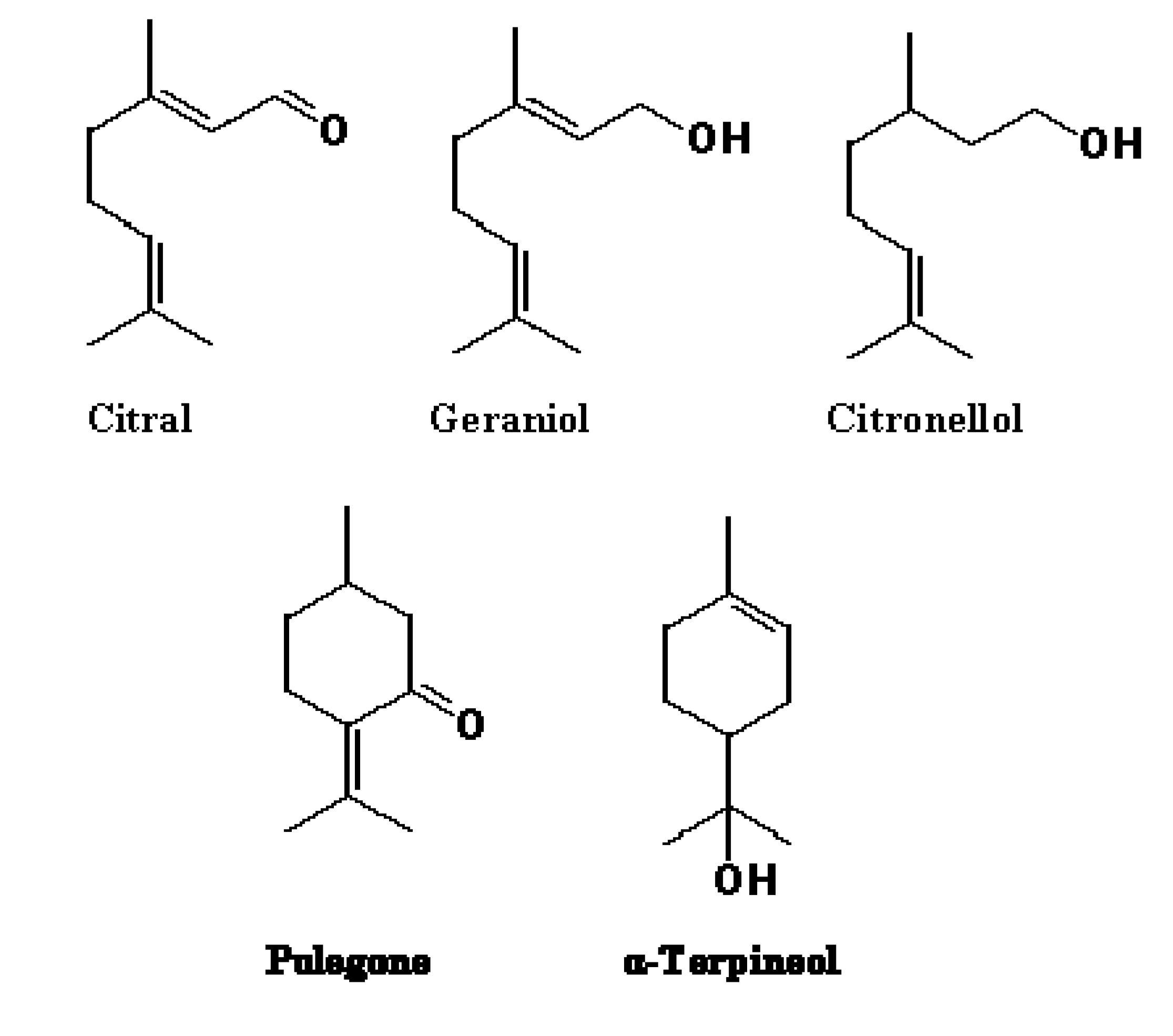{kind=link}
{kind=link}
{kind=link}
{kind=link}
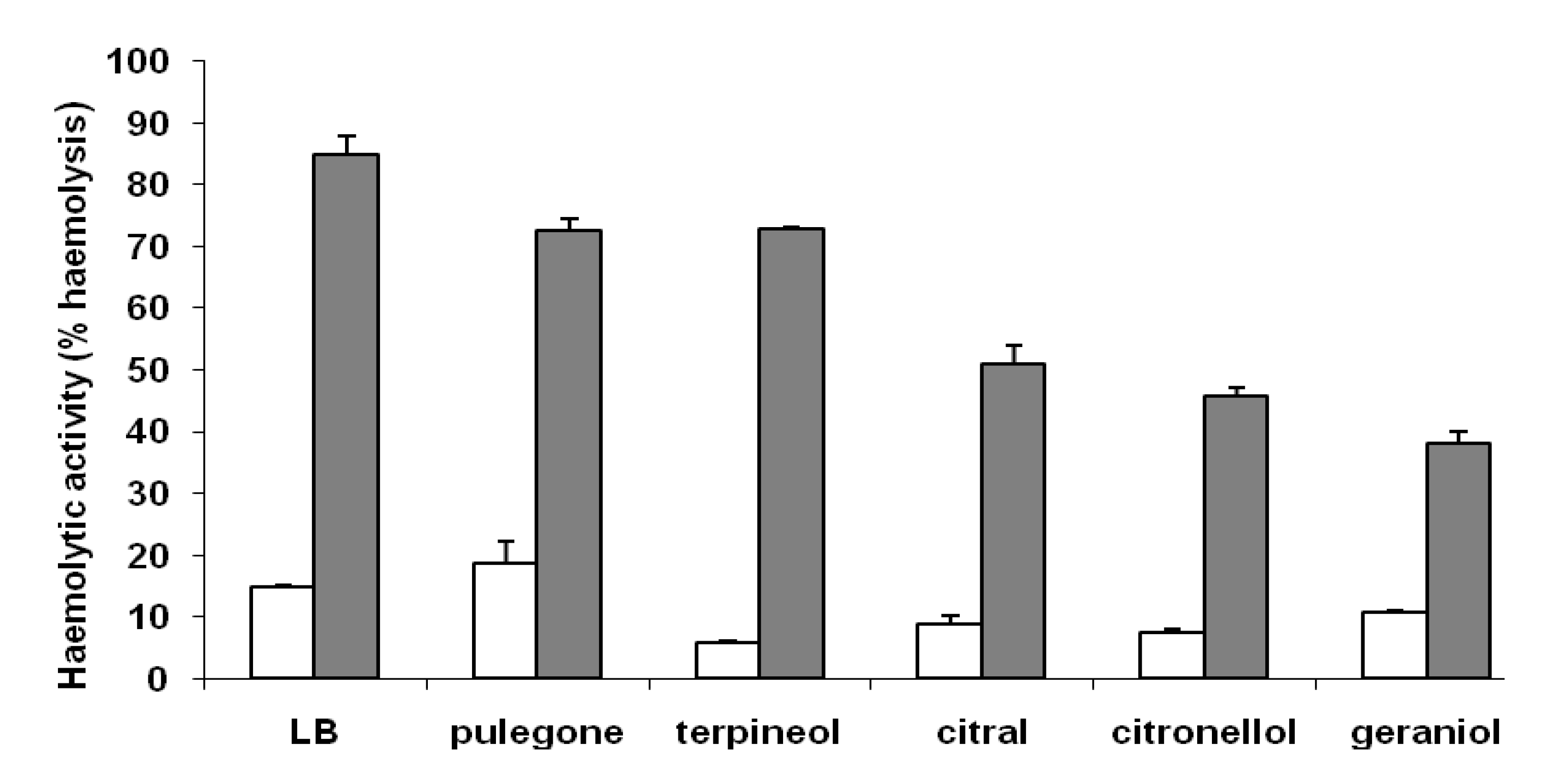{kind=link}
| Treatments | MIC (mg/L) | Colony diameter (mm)*1 | Nº of concentic rings | Second ring width (mm)1 |
|---|---|---|---|---|
| pulegone | 5 | 53.2 ± 1.5e | 5-6 | 3.8 ± 1.4bc |
| citronellol | 3 | 23.7 ± 1.5f | 5-6 | 1.0 ± 0.5d |
| citronellal | 6 | 68.0 ± 0.6b | 6 | 4.3 ± 0.3b |
| citronellyl acetate | >10 | 79.2 ± 3.5b | 6 | 4.5 ± 0.9b |
| geraniol | 3 | 22.5 ± 0.6f | 3-4 | 0.8 ± 0.3d |
| bornyl acetate | 10 | 71.3 ± 1.5b | 6 | 4.3 ± 0.3b |
| α-terpinene | 7.5 | 79.3 ± 3.5ab | 5-6 | 6.1 ± 2.2a |
| α -terpineol | 5 | 49.6 ± 1.0d | 4-5 | 4.2 ± 0.4b |
| terpinene-4-ol | 5 | 62.5 ± 1.0c | 5-6 | 4.5 ± 1.3b |
| linalool | 6 | 72.8 ± 1.7b | 5-6 | 6.2 ± 1.2a |
| linallyl acetate | >10 | 82.1 ± 3.0a | 6 | 4.5 ± 0.1b |
| limonene | 10 | 82.0 ± 2.7a | 6 | 5.4 ± 1.3ab |
| citral | 5 | 41.0 ± 1.5d | 4-5 | 2.8 ± 1.0c |
| mentone | >10 | 66.7 ± 1.7c | 5-6 | 4.7 ± 1.7b |
| β-pinene | >10 | 79.5 ± 4.7ab | 6 | 7.0 ± 3.6a |
| 1,8-cineol | 7.5 | 74.6 ± 2.5b | 6 | 3.0 ± 0.9c |
| carveol | 10 | 83.2 ± 1.7a | 5-6 | 7.0 ± 2.3a |
| Control | - | 84.2 ± 3.5a | 6 | 6.5 ± 2.2a |


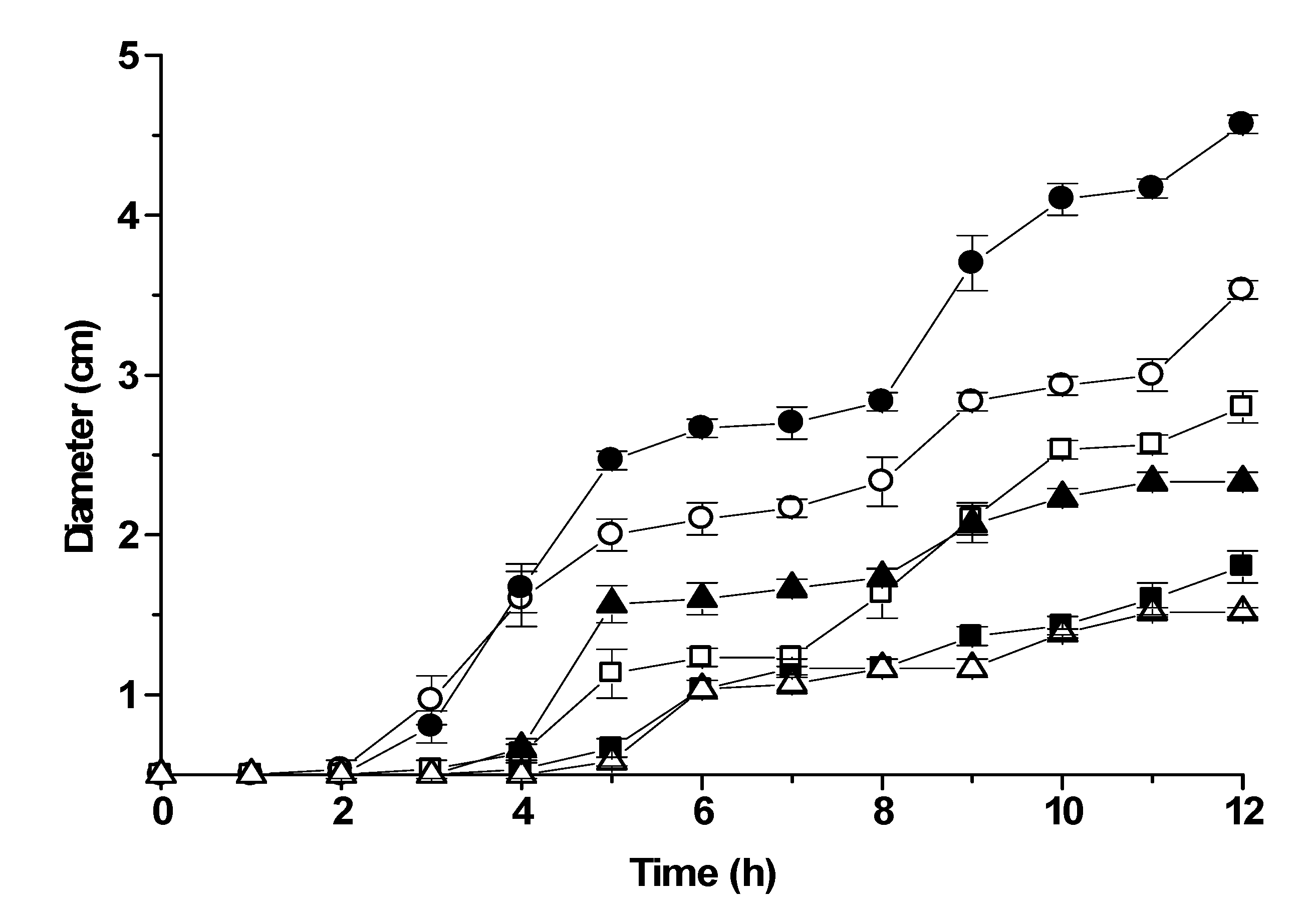
 geraniol (0.3 mg/L), ■ citronellol (0.3 mg/L), □ α-terpineol (0.5mg/L). The data represent the averages of colony diameter of three independent experiments with standard deviations.
geraniol (0.3 mg/L), ■ citronellol (0.3 mg/L), □ α-terpineol (0.5mg/L). The data represent the averages of colony diameter of three independent experiments with standard deviations.
geraniol (0.3 mg/L), ■ citronellol (0.3 mg/L), □ α-terpineol (0.5mg/L). The data represent the averages of colony diameter of three independent experiments with standard deviations.
geraniol (0.3 mg/L), ■ citronellol (0.3 mg/L), □ α-terpineol (0.5mg/L). The data represent the averages of colony diameter of three independent experiments with standard deviations.
 Control;
Control;  0.06 mg/L;
0.06 mg/L;  0.12 mg/L;
0.12 mg/L;  0.25 mg/L;
0.25 mg/L;  0.50 mg/L and
0.50 mg/L and  0.75 mg/L.
Control; 0.06 mg/L; 0.12 mg/L; 0.25 mg/L; 0.50 mg/L and 0.75 mg/L.
0.75 mg/L.
Control; 0.06 mg/L; 0.12 mg/L; 0.25 mg/L; 0.50 mg/L and 0.75 mg/L.


Experimental
Bacterial strain and growth conditions
Terpenes and terpenoids
MIC assay
Swarming behavior assay
Measurement of cell length
Membrane-associated haemolysin
Acknowledgements
References
- Mobley, H.L.; Belas, R. Swarming and pathogenicity of Proteus mirabilis in the urinary tract. Trends Microbiol. 1995, 3, 280–284. [Google Scholar] [CrossRef]
- Senior, B.W. Proteus morgani is less frequently associated with urinary tract infections than Proteus mirabilis- an explanation. J. Med. Microbiol. 1983, 35, 203–207. [Google Scholar] [CrossRef]
- Fraser, G.M.; Hughes, C. Swarming motility. Curr. Opin. Microbiol. 1999, 2, 630–635. [Google Scholar] [CrossRef]
- Sturgill, G.; Rather, P.N. Evidence that putrescine acts as an extracellular signal required for swarming in Proteus mirabilis. Mol. Microbiol. 2004, 51, 437–446. [Google Scholar] [CrossRef]
- Rather, P.N. Swarmer cell differentiation in Proteus mirabilis. Environ. Microbiol. 2005, 7, 1065–1073. [Google Scholar] [CrossRef]
- Wassif, C.; Cheek, D.; Belas, R. Molecular analysis of a metalloprotease from Proteus mirabilis. J. Bacteriol. 1995, 177, 5790–5798. [Google Scholar]
- Rozalski, A.; Sidorczyk, Z.; Katelko, K. Potential virulence factors of Proteus bacilli. Microbiol. Mol. Biol. Rev. 1997, 61, 65–89. [Google Scholar]
- Fraser, G.M.; Claret, L.; Furness, R.; Gupa, S.; Hughes, C. Swarming-coupled expression of the Proteus mirabilis hpmBA haemolysin operon. Microbiology 2002, 148, 2191–2201. [Google Scholar]
- Mobley, H.L.; Belas, R.; Lockatell, V.; Chippendale, G.; Trifillis, A.L.; Johnson, D.E.; Warren, J.W. Construction of a flagellum negative mutant of Proteus mirabilis: effect on internalization by human renal epithelial cells and virulence in a mouse model of ascending urinary tract infection. Infect. Immunity 1996, 64, 5332–5340. [Google Scholar]
- Walker, K.E.; Moghaddame-Jafari, S.; Locktell, C.V.; Johnson, D.; Belas, R. ZapA, the IgA-degradin metalloprotease of Proteus mirabilis, is a virulence factor expressed specifically in swarmer cells. Mol. Microbiol. 1999, 32, 825–836. [Google Scholar]
- Alwen, J.; Smith, D.G. A medium to suppress the swarming of Proteus species. J. Appl. Microbiol. 1967, 30, 389–394. [Google Scholar]
- William, F.D. Abolition of swarming of Proteus by p-nitrophenyl glycerin: general properties. Appl. Environ. Microbiol. 1973, 25, 745–750. [Google Scholar]
- Hernandez, E.; Ramisse, F.; Carvalho, J.D. Abolition of swarming of Proteus. J. Clin. Microbiol. 1999, 37, 3435. [Google Scholar]
- Liaw, S.J.; Lai, H.C.; Ho, S.W.; Luh, K.T.; Wang, W.B. Inhibition of virulence factor expression ad swarming differentiation in Proteus mirabilis by p-nitrophenylglycerol. J. Med. Microbiol. 2000, 49, 725–731. [Google Scholar]
- Liaw, S.J.; Lai, H.C.; Ho, S.W.; Luh, K.T.; Wang, W.B. Characterization of p-nitrophenylglycerol-resistant Proteus mirabilis super-swarming mutants. J. Med. Microbiol. 2001, 50, 1039–1048. [Google Scholar]
- Liaw, S.J.; Lai, H.C.; Wang, W.B. Modulation of swarming and virulence by fatty acids through the RsbA protein in Proteus mirabilis. Infect. Immunity 2004, 72, 6836–6845. [Google Scholar] [CrossRef]
- Wang, W.B; Lai, H.C.; Hsueh, P.R.; Chiou, R.Y.; Lin, S.B.; Liaw, S.J. Inhibition of swarming and virulence factor expression in Proteus mirabilis by resveratrol. J. Med. Microbiol. 2006, 55, 1313–1321. [Google Scholar] [CrossRef]
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar]
- Trombetta, D.; Castelli, F.; Sarpietro, M.G.; Venuti, V.; Cristani, M.; Daniele, C.; Saija, A.; Mazzanti, G.; Bisignano, G. Mechanisms of antibacterial action of three monoterpenes. Antimicrob. Agents Chemother. 2005, 49, 2474–2478. [Google Scholar] [CrossRef]
- Hammer, K.A.; Carson, C.F.; Riley, T.V. Antimicrobial activity of essential oils and other plant extracts. J. Appl. Microbiol. 1999, 86, 985–990. [Google Scholar] [CrossRef]
- Essawi, T.; Srour, M. Screening of some Palestinian medicinal plants for antibacterial activity. J. Ethnopharmacol. 2000, 70, 343–349. [Google Scholar] [CrossRef]
- Burt, S.A. Essential oils: their antibacterial properties and potential applications in foods: a review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Kotan, R.; Kordali, S.; Cakir, A. Screening of antibacterial activities of twenty-one oxygenated monoterpenes. Z. Naturforsch. C 2007, 62, 507–513. [Google Scholar]
- Mansouri, S.; Amari, A.; Asad, A.G. Inhibitory effect of some medicinal plants from Iran on swarming motility of Proteus rods. J. Med. Sci. 2005, 5, 216–221. [Google Scholar] [CrossRef]
- Naigre, R.; Kalck, P.; Roques, C.; Rocux, I.; Michel, G. Comparison of antimicrobial properties of monoterpenes and their carbonylated products. Planta Med. 1996, 62, 275–277. [Google Scholar] [CrossRef]
- Dahr, P.; Ayala, U.; Andarge, E.; Morisseau, S.; Snyder-Leiby, T. Study of the structural changes on the antimicrobial activity of [3.1.1.]-bicyclics. J. Essent. Oil Res. 2004, 16, 612–616. [Google Scholar] [CrossRef]
- Costa, S.O.P.; Bianco, M.; Bonato, M.C. Instability of protease (gelatinase) trait in Proteus mirabilis. Rev. Microbiol. 1982, 13, 35–45. [Google Scholar]
- Mann, C.M.; Markham, J.L. A new method for determining the minimum inhibitory concentration of essential oils. J. Appl. Microbiol. 1998, 84, 538–544. [Google Scholar]
- Koronakis, V.; Cross, M.; Senior, B.; Koronakis, E.; Hugues, C. The secreted hemolysins of Proteus mirabilis, Proteus vulgaris, and Morganella morganii are genetically related to each other and to the alpha-hemolysin of Escherichia coli. J. Bacteriol. 1987, 169, 1509–1515. [Google Scholar]
- Sample Availability: Requests for samples of the terpenes should be directed to Dr. Sergio Echeverrigaray.
© 2008 by the authors. Licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Echeverrigaray, S.; Michelim, L.; Longaray Delamare, A.P.; Andrade, C.P.; Pinto da Costa, S.O.; Zacaria, J. The Effect of Monoterpenes on Swarming Differentiation and Haemolysin Activity in Proteus mirabilis. Molecules 2008, 13, 3107-3116. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules13123107
Echeverrigaray S, Michelim L, Longaray Delamare AP, Andrade CP, Pinto da Costa SO, Zacaria J. The Effect of Monoterpenes on Swarming Differentiation and Haemolysin Activity in Proteus mirabilis. Molecules. 2008; 13(12):3107-3116. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules13123107
Chicago/Turabian StyleEcheverrigaray, Sergio, Lessandra Michelim, Ana Paula Longaray Delamare, Cristiane Paim Andrade, Sérgio Olavo Pinto da Costa, and Jucimar Zacaria. 2008. "The Effect of Monoterpenes on Swarming Differentiation and Haemolysin Activity in Proteus mirabilis" Molecules 13, no. 12: 3107-3116. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules13123107