The Potential of Tetrandrine as a Protective Agent for Ischemic Stroke

{kind=link}
Abstract
:1. Introduction to Ischemic Stroke
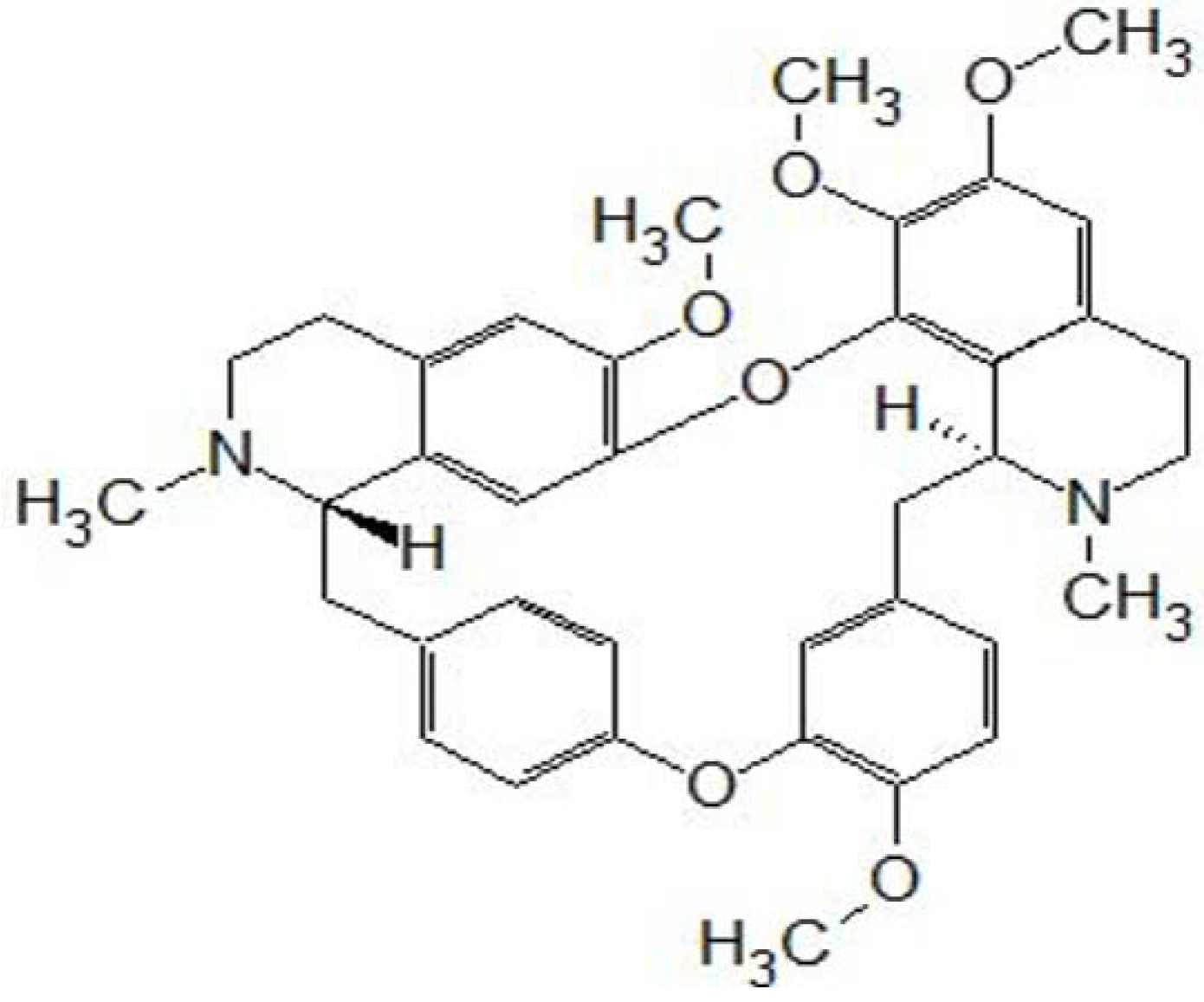
2. Introduction to Tetrandrine

3. Effects of Tetrandrine on the Ischemia-Reperfusion Injury of Various Tissues/Organs
4. Calcium Channel Blocking Effects of Tetrandrine
5. Anti-Inflammatory Effects of Tetrandrine
6. Antioxidative Effects of Tetrandrine
7. Tetrandrine Inhibits Platelet Aggregation
8. Evidence of Tetrandrine against Stroke
9. Toxicity of Tetrandrine
10. Conclusions
Conflict of Interest
References
- Woodruff, T.M.; Thundyil, J.; Tang, S.C.; Sobey, C.G.; Taylor, S.M.; Arumugam, T.V. Pathophysiology, treatment, and animal and cellular models of human ischemic stroke. Mol. Neurodegener. 2011, 6, 11. [Google Scholar] [CrossRef]
- Arumugam, T.V.; Granger, D.N.; Mattson, M.P. Stroke and T-cells. Neuromol. Med. 2005, 7, 229–242. [Google Scholar] [CrossRef]
- Legos, J.J.; Barone, F.C. Update on pharmacological strategies for stroke: Prevention, acute intervention and regeneration. Curr. Opin. Investig. Drugs 2003, 4, 847–858. [Google Scholar]
- Broughton, B.R.S.; Reutens, D.C.; Sobey, C.G. Apoptotic mechanisms after cerebral ischemia. Stroke 2009, 40, E331–E339. [Google Scholar] [CrossRef]
- Zheng, Z.; Yenari, M.A. Post-ischemic inflammation: Molecular mechanisms and therapeutic implications. Neurol. Res. 2004, 26, 884–892. [Google Scholar] [CrossRef]
- Ho, L.J.; Chang, D.M.; Chang, M.L.; Kuo, S.Y.; Lai, J.H. Mechanism of immunosuppression of the antirheumatic herb TWHf in human T cells. J. Rheumatol. 1999, 26, 14–22. [Google Scholar]
- Chen, Y.; Chen, J.C.; Tseng, S.H. Effects of tetrandrine plus radiation on neuroblastoma cells. Anticancer Res. 2009, 29, 3163–3171. [Google Scholar]
- Chen, Y.; Chen, J.C.; Tseng, S.H. Tetrandrine suppresses tumor growth and angiogenesis of gliomas in rats. Int. J. Cancer 2009, 124, 2260–2269. [Google Scholar] [CrossRef]
- Chen, Y.; Tseng, S.H. The potential of tetrandrine against gliomas. Anticancer Agents Med. Chem. 2010, 10, 534–542. [Google Scholar]
- Lieberman, I.; Lentz, D.P.; Trucco, G.A.; Seow, W.K.; Thong, Y.H. Prevention by tetrandrine of spontaneous development of diabetes mellitus in BB rats. Diabetes 1992, 41, 616–619. [Google Scholar]
- Wang, H.L.; Kilfeather, S.A.; Martin, G.R.; Page, C.P. Effects of tetrandrine on growth factor-induced DNA synthesis and proliferative response of rat pulmonary artery smooth muscle cells. Pulm. Pharmacol. Ther. 2000, 13, 53–60. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Fang, L.H. Antagonism of morphine-induced antinociception by tetrandrine is dependent on serotonergic mechanism. Life Sci. 2001, 69, 1429–1439. [Google Scholar] [CrossRef]
- Li, S.Y.; Teh, B.S.; Seow, W.K.; Ling, L.H.; Thong, Y.H. Effect of tetrandrine on immunological response and cardiac transplant rejection in mice. Int. Arch. Allergy Appl. Immunol. 1989, 90, 169–173. [Google Scholar] [CrossRef]
- Chen, Y.; Lai, H.S.; Chiang, B.L.; Tseng, S.H.; Chen, W.J. Tetrandrine attenuated dendritic cell-mediated alloimmune responses and prolonged graft survival in mice. Planta Med. 2010, 76, 1424–1430. [Google Scholar] [CrossRef]
- Wu, J.M.; Chen, Y.; Chen, J.C.; Lin, T.Y.; Tseng, S.H. Tetrandrine induces apoptosis and growth suppression of colon cancer cells in mice. Cancer Lett. 2010, 287, 187–195. [Google Scholar] [CrossRef]
- Liu, Z.; Xu, Z.; Shen, W.; Li, Y.; Zhang, J.; Ye, X. Effect of pharmacologic preconditioning with tetrandrine on subsequent ischemia/reperfusion injury in rat liver. World J. Surg. 2004, 28, 620–624. [Google Scholar]
- Shen, Y.C.; Chen, C.F.; Sung, Y.J. Tetrandrine ameliorates ischaemia-reperfusion injury of rat myocardium through inhibition of neutrophil priming and activation. Br. J. Pharmacol. 1999, 128, 1593–1601. [Google Scholar] [CrossRef]
- Wong, T.M.; Wu, S.; Yu, X.C.; Li, H.Y. Cardiovascular actions of Radix Stephaniae Tetrandrae: A comparison with its main component, tetrandrine. Acta Pharmacol. Sin. 2000, 21, 1083–1088. [Google Scholar]
- Chen, Y.; Wu, J.M.; Lin, T.Z.; Wu, C.C.; Chiu, K.M.; Chang, B.F.; Tseng, S.H.; Chu, S.H. Tetrandrine ameliorated reperfusion injury of small bowel transplantation. J. Pediatr. Surg. 2009, 44, 2145–2152. [Google Scholar] [CrossRef]
- Li, Y.Y.; Li, X.L.; Yang, C.X.; Zhong, H.; Yao, H.; Zhu, L. Effects of tetrandrine and QYT on ICAM-1 and SOD gene expression in pancreas and liver of rats with acute pancreatitis. World J. Gastroenterol. 2003, 9, 155–159. [Google Scholar]
- Mehta, S.L.; Manhas, N.; Rahubir, R. Molecular targets in cerebral ischemia for developing novel therapeutics. Brain Res. Rev. 2007, 54, 34–66. [Google Scholar] [CrossRef]
- Huang, M.H.; Wu, S.N.; Chen, C.P.; Shen, A.Y. Inhibition of Ca2+-activated and voltage-dependent K+ currents by 2-mercaptophenyl-1,4-naphthoquinone in pituitary GH3 cells contribution to its antiproliferative effects. Life Sci. 2002, 70, 1185–1203. [Google Scholar] [CrossRef]
- Jin, Q.; Kang, C.; Soh, Y.; Sohn, N.W.; Lee, J.; Cho, Y.H.; Baik, H.H.; Kang, I. Tetrandrine cytotoxicity and its dual effect on oxidative stress-induced apoptosis through modulating cellular redox states in Neuro 2a mouse neuroblastoma cells. Life Sci. 2002, 71, 2053–2066. [Google Scholar] [CrossRef]
- Wang, G.; Lemos, J.R.; Iadecola, C. Herbal alkaloid tetrandrine: From an ion channel blocker to inhibitor of tumor proliferation. Trends Pharmacol. Sci. 2004, 25, 120–123. [Google Scholar] [CrossRef]
- Han, X.; Wang, F.; Yao, W.; Xing, H.; Weng, D.; Song, X.; Chen, G.; Xi, L.; Zhu, T.; Zhou, J.; et al. Heat shock proteins and p53 play a critical role in K+ channel-mediated tumor cell proliferation and apoptosis. Apoptosis 2007, 12, 1837–1846. [Google Scholar] [CrossRef]
- Wang, G.; Lemos, J.R. Tetrandrine blocks a slow, large-conductance, Ca(2+)-activated potassium channel besides inhibiting a non-inactivating Ca2+ current in isolated nerve terminals of the rat neurohypophysis. Pflugers Arch. 1992, 421, 558–565. [Google Scholar] [CrossRef]
- Ishikawa, M.; Zhang, J.H.; Nanda, A.; Granger, D.N. Inflammatory responses to ischemia and reperfusion in the cerebral microcirculation. Front. Biosci. 2004, 9, 1339–1347. [Google Scholar] [CrossRef]
- Hudome, S.; Palmer, C.; Roberts, R.L.; Mauger, D.; Housman, C.; Towfighi, J. The role of neutrophils in the production of hypoxic-ischemic brain injury in the neonatal rat. Pediatr. Res. 1997, 41, 607–616. [Google Scholar] [CrossRef]
- Matsuo, Y.; Onodera, H.; Shiga, Y.; Nakamura, M.; Ninomiya, M.; Kihara, T.; Kogure, K. Correlation between myeloperoxidase-quantified neutrophil accumulation and ischemic brain injury in the rat. Effects of neutrophil depletion. Stroke 1994, 25, 1469–1475. [Google Scholar] [CrossRef]
- Mergenthaler, P.; Dirnagl, U.; Meisel, A. Pathophysiology of stroke: Lessons from animal models. Metab. Brain Dis. 2004, 19, 151–167. [Google Scholar] [CrossRef]
- Yilmaz, G.; Arumugam, T.V.; Stokes, K.Y.; Granger, D.N. Role of T lymphocytes and interferon-gamma in ischemic stroke. Circulation 2006, 113, 2105–2012. [Google Scholar] [CrossRef]
- del Zoppo, G.J.; Milner, R.; Mabuchi, T.; Hung, S.; Wang, X.Y.; Berg, G.I.; Koziol, J.A. Microglial activation and matrix protease generation during focal cerebral ischemia. Stroke 2007, 38, 646–651. [Google Scholar] [CrossRef]
- Tuttolomondo, A.; di Raimondo, D.; di Sciacca, R.; Pinto, A.; Licata, G. Inflammatory cytokines in acute ischemic stroke. Curr. Pharm. Des. 2008, 14, 3574–3589. [Google Scholar] [CrossRef]
- Wang, Q.; Tang, X.N.; Yenari, M.A. The inflammatory response in stroke. J. Neuroimmunol. 2007, 184, 53–68. [Google Scholar] [CrossRef]
- Gelderblom, M.; Leypoldt, F.; Steinbach, K.; Behrens, D.; Choe, C.U.; Siler, D.A.; Arumugam, T.V.; Orthey, E.; Gerloff, C.; Tolosa, E.; et al. Temporal and spatial dynamics of cerebral immune cell accumulation in stroke. Stroke 2009, 40, 1849–1857. [Google Scholar] [CrossRef]
- Jin, R.; Yang, G.J.; Li, G.H. Inflammatory mechanisms in ischemic stroke: Role of inflammatory cells. J. Leukoc Biol. 2010, 87, 779–789. [Google Scholar] [CrossRef]
- Goussev, A.V.; Zhang, Z.G.; Anderson, D.C.; Chopp, M. P-selectin antibody reduces hemorrhage and infarct volume resulting from MCA occlusion in the rat. J. Neurol. Sci. 1998, 161, 16–22. [Google Scholar] [CrossRef]
- Zhang, R.L.; Chopp, M.; Jiang, N.; Tang, W.X.; Prostak, J.; Manning, A.M.; Anderson, D.C. Antiintercellular adhesion molecule-1 antibody reduces ischemic cell damage after transient but not permanent middle cerebral artery occlusion in the Wistar rat. Stroke 1995, 26, 1438–1443. [Google Scholar] [CrossRef]
- Furuya, K.; Takeda, H.; Azhar, S.; McCarron, R.M.; Chen, Y.; Ruetzler, C.A.; Wolcott, K.M.; DeGraba, T.J.; Rothlein, R.; Hugli, T.E.; et al. Examination of several potential mechanisms for the negative outcome in a clinical stroke trial of enlimomab, a murine anti-human intercellular adhesion molecule-1 antibody–A bedside-to-bench study. Stroke 2001, 32, 2665–2674. [Google Scholar] [CrossRef]
- Schneider, D.; Berrouschot, J.; Brandt, T.; Hacke, W.; Ferbert, A.; Norris, S.H.; Polmar, S.H.; Schafer, E. Safety, pharmacokinetics and biological activity of enlimomab (anti-ICAM-1 antibody): An open-label, dose escalation study in patients hospitalized for acute stroke. Eur. Neurol. 1998, 40, 78–83. [Google Scholar] [CrossRef]
- Chang, D.M.; Chang, W.Y.; Kuo, S.Y.; Chang, M.L. The effects of traditional antirheumatic herbal medicines on immune response cells. J. Rheumatol. 1997, 24, 436–441. [Google Scholar]
- Lin, T.Y.; Tseng, S.H.; Li, S.J.; Chen, J.C.; Shieh, J.S.; Chen, Y. Tetrandrine increased the survival rate of mice with lipopolysaccharide-induced endotoxemia. J. Trauma 2009, 66, 411–417. [Google Scholar] [CrossRef]
- Wong, C.W.; Seow, W.K.; O’Callaghan, J.W.; Thong, Y.H. Comparative effects of tetrandrine and berbamine on subcutaneous air pouch inflammation induced by interleukin-1, tumor necrosis factor and platelet-activating factor. Agents Actions 1992, 36, 112–118. [Google Scholar] [CrossRef]
- Wong, C.W.; Thong, Y.H.; Seow, W.K. Comparative effects of tetrandrine and berbamine on guineapig airway microvascular leakage induced by platelet-activating factor and other allergic mediators. Int. J. Immunopharmacol. 1993, 15, 185–193. [Google Scholar] [CrossRef]
- Shen, Y.C.; Chen, C.F.; Wang, S.Y.; Sung, Y.J. Impediment to calcium influx and reactive oxygen production accounts for the inhibition of neutrophil Mac-1 up-regulation and adhesion by tetrandrine. Mol. Pharmacol. 1999, 55, 186–193. [Google Scholar]
- Allen, C.L.; Bayraktutan, U. Oxidative stress and its role in the pathogenesis of ischaemic stroke. Int. J. Stroke 2009, 4, 461–470. [Google Scholar] [CrossRef]
- Choi, K.; Kim, J.; Kim, G.W.; Choi, C. Oxidative stress-induced necrotic cell death via mitochondira-dependent burst of reactive oxygen species. Curr. Neurovasc. Res. 2009, 6, 213–222. [Google Scholar] [CrossRef]
- Suh, S.W.; Shin, B.S.; Ma, H.L.; van Hoecke, M.; Brennan, A.M.; Yenari, M.A.; Swanson, R.A. Glucose and NADPH oxidase drive neuronal superoxide formation in stroke. Ann. Neurol. 2008, 64, 654–663. [Google Scholar] [CrossRef]
- Girouard, H.; Iadecola, C. Neurovascular coupling in the normal brain and in hypertension, stroke, and alzheimer disease. J. Appl. Physiol. 2006, 100, 328–335. [Google Scholar] [CrossRef]
- Hall, E.D. Inhibition of lipid peroxidation in central nervous system trauma and ischemia. J. Neurol. Sci. 1995, 134, 79–83. [Google Scholar] [CrossRef]
- Im, J.Y.; Kim, D.; Paik, S.G.; Han, P.L. Cyclooxygenase-2-dependent neuronal death proceeds via superoxide anion generation. Free Radic. Biol. Med. 2006, 41, 960–972. [Google Scholar] [CrossRef]
- Lau, A.T.Y.; Wang, Y.; Chiu, J.F. Reactive oxygen species: Current knowledge and applications in cancer research and therapeutic. J. Cell. Biochem. 2008, 104, 657–667. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Kamendulis, L.M. The role of oxidative stress in carcinogenesis. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 239–267. [Google Scholar] [CrossRef]
- Iwashita, A.; Maemoto, T.; Nakada, H.; Shima, I.; Matsuoka, N.; Hisajima, H. A novel potent radical scavenger, 8-(4-fluorophenyl)-2-((2E)-3-phenyl-2-propenoyl)-1,2,3,4-tetrahydropyrazolo[5,1-c][1,2,4]triazine (FR210575), prevents neuronal cell death in cultured primary neurons and attenuates brain injury after focal ischemia in rats. J. Pharmacol. Exp. Ther. 2003, 307, 961–968. [Google Scholar] [CrossRef]
- Sydserff, S.G.; Borelli, A.R.; Green, A.R.; Cross, A.J. Effect of NXY-059 on infarct volume after transient or permanent middle cerebral artery occlusion in the rat; studies on dose, plasma concentration and therapeutic time window. Br. J. Pharmacol. 2002, 135, 103–112. [Google Scholar] [CrossRef]
- Lees, K.R.; Zivin, J.A.; Ashwood, T.; Davalos, A.; Davis, S.M.; Diener, H.; Grotta, J.; Lyden, P.; Shuaib, A.; Hardemark, H.; et al. NXY-059 for acute ischemic stroke. N. Engl. J. Med. 2006, 354, 588–600. [Google Scholar] [CrossRef]
- Shuaib, A.; Lees, K.R.; Lyden, P.; Grotta, J.; Davalos, A.; Davis, S.M.; Diener, H.; Ashwood, T.; Wasiewski, W.W.; Emeribe, U. NXY-059 for the treatment of acute ischemic stroke. N. Engl. J. Med. 2007, 357, 562–571. [Google Scholar] [CrossRef]
- Koh, S.B.; Ban, J.Y.; Lee, B.Y.; Seong, Y.H. Protective effects of fangchinoline and tetrandrine on hydrogen peroxide-induced oxidative neuronal cell damage in cultured rat cerebellar granule cells. Planta Med. 2003, 69, 506–512. [Google Scholar] [CrossRef]
- Fernandes, M.A.; Custodio, J.B.; Santos, M.S.; Moreno, A.J.M.; Vicente, J.A.F. Tetrandrine concentrations not affecting oxidative phosphorylation protect rat liver mitochondria from oxidative stress. Mitochondrion 2006, 6, 176–185. [Google Scholar] [CrossRef]
- Ye, J.; Ding, M.; Zhang, X.; Rojanasakul, Y.; Shi, X. On the role of hydroxyl radical and the effect of tetrandrine on nuclear factor-kappaB activation by phorbol 12-myristate 13-acetate. Ann. Clin. Lab. Sci. 2000, 30, 65–71. [Google Scholar]
- Shi, X.; Mao, Y.; Saffiotti, U.; Wang, L.; Rojanasakul, Y.; Leonard, S.S.; Vallyathan, V. Antioxidant activity of tetrandrine and its inhibition of quartz-induced lipid peroxidation. J. Toxicol. Environ. Health 1995, 46, 233–248. [Google Scholar] [CrossRef]
- Cao, Z.F. Scavenging effect of tetrandrine on active oxygen radicals. Planta Med. 1996, 62, 413–414. [Google Scholar] [CrossRef]
- Castranova, V. Generation of oxygen radicals and mechanisms of injury prevention. Environ. Health Perspect. 1994, 102, 65–68. [Google Scholar]
- Castranova, V.; Kang, J.H.; Moore, M.D.; Pailes, W.H.; Frazer, D.G.; Schwegler-Berry, D. Inhibition of stimulant-induced activation of phagocytic cells with tetrandrine. J. Leukoc. Biol. 1991, 50, 412–422. [Google Scholar]
- Chen, F.; Sun, S.; Kuhn, D.C.; Lu, Y.; Gaydos, L.J.; Shi, X.; Demers, L.M. Tetrandrine inhibits signal-induced NF-kB activation in rat alveolar macrophages. Biochem. Biophys. Res. Commun. 1997, 231, 99–102. [Google Scholar] [CrossRef]
- Shen, Y.C.; Chou, C.J.; Chiou, W.F.; Chen, C.F. Anti-inflammatory effects of the partially purified extract of radix stephaniae tetrandrae: Comparative studies of its active principles tetrandrine and fangchinoline on human polymorphonuclear leukocyte functions. Mol. Pharmacol. 2001, 60, 1083–1090. [Google Scholar]
- Sekiya, N.; Hikiami, H.; Yokoyama, K.; Kouta, K.; Sakakibara, I.; Shimada, Y.; Terasawa, K. Inhibitory effects of Stephania tetrandra S. Moore on free radical-induced lysis of rat blood cells. Biol. Pharm. Bull. 2005, 28, 667–670. [Google Scholar] [CrossRef]
- Zhou, Y.D.; Fang, X.F.; Cui, Z.J. UVA-induced calcium oscillation in rat mast cells. Cell Calcium 2009, 45, 18–28. [Google Scholar] [CrossRef]
- Steg, P.G.; Dorman, S.H.; Amarenco, P. Atherothromobosis and the role of antiplatelet therapy. J. Thromb. Haemost. 2011, 9, 325–332. [Google Scholar]
- Nieswandt, B.; Pleines, I.; Bender, M. Platelet adhesion and activation mechanisms in arterial thrombosis and ischemic stroke. J. Thromb. Haemost. 2011, 9, 92–104. [Google Scholar]
- Hashizume, T.; Yamaguchi, H.; Sato, T.; Fujii, T. Suppressive effect of biscoclaurine alkaloids on agonist-induced activation of phospholipase A2 in rabbit platelets. Biochem. Pharmacol. 1991, 41, 419–423. [Google Scholar]
- Kim, H.S.; Zhang, Y.H.; Fang, L.H.; Yun, Y.P.; Lee, H.K. Effects of tetrandrine and fangchinoline on human platelet aggregation and thromboxane B2 formation. J. Ethnopharmacol. 1999, 66, 241–246. [Google Scholar] [CrossRef]
- Teh, B.S.; Ioannoni, B.; Seow, W.K.; McCormack, J.G.; Thong, Y.H. Suppression by tetrandrine of human platelet aggregation induced by platelet-activating factor and other stimulants. Int. Arch. Allergy Appl. Immunol. 1989, 88, 267–272. [Google Scholar] [CrossRef]
- Kim, H.S.; Zhang, Y.H.; Yun, Y.P. Effects of tetrandrine and fangchinoline on experimental thrombosis in mice and human platelet aggregation. Planta Med. 1999, 65, 135–138. [Google Scholar] [CrossRef]
- Chang, K.H.; Chen, M.L.; Chen, H.C.; Huang, Y.W.; Wu, T.Y.; Chen, Y.J. Enhancement of radiosensitivity in human glioblastoma U138MG cells by tetrandrine. Neoplasma 1999, 46, 196–200. [Google Scholar]
- Sun, F.; Liu, T.P. Tetrandrine vs nicardipine in cerebral ischemia-reperfusion damages in Gerbils. Acta Pharmacol. Sin. 1995, 16, 145–148. [Google Scholar]
- Gralla, E.J.; Coleman, G.L.; Jonas, A.M. Toxicology studies with d-tetrandrine (NSC-77037), a plant alkaloid with vascular and lymphotoxic effects in dogs and monkeys. Cancer Chemother. Rep. Part 3 1974, 5, 79–85. [Google Scholar]
- Herman, E.H.; Chadwick, D.P. Cardiovascular effects of d-Tetrandrine. Pharmacology 1974, 12, 97–109. [Google Scholar] [CrossRef]
- Hui, S.C.G.; Chan, T.Y.; Chen, Y.Y. Tetrandrine inhibits lipid peroxidation but lacks reactivity towards superoxide anion and hydrogen peroxide. Pharmacol. Toxicol. 1996, 78, 200–201. [Google Scholar] [CrossRef]
- Liu, S.J.; Zhou, S.W.; Xue, C.S. Effect of tetrandrine on neutrophilic recruitment response to brain ischemia/reperfusion. Acta. Pharmacol. Sin. 2001, 22, 971–975. [Google Scholar]
- Sample Availability: Not available.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, Y.; Tsai, Y.-H.; Tseng, S.-H. The Potential of Tetrandrine as a Protective Agent for Ischemic Stroke. Molecules 2011, 16, 8020-8032. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules16098020
Chen Y, Tsai Y-H, Tseng S-H. The Potential of Tetrandrine as a Protective Agent for Ischemic Stroke. Molecules. 2011; 16(9):8020-8032. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules16098020
Chicago/Turabian StyleChen, Yun, Ya-Hui Tsai, and Sheng-Hong Tseng. 2011. "The Potential of Tetrandrine as a Protective Agent for Ischemic Stroke" Molecules 16, no. 9: 8020-8032. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules16098020