Icariin Is A PPARα Activator Inducing Lipid Metabolic Gene Expression in Mice

Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of Icariin on the mRNA of PPARα and Marker Enzymes

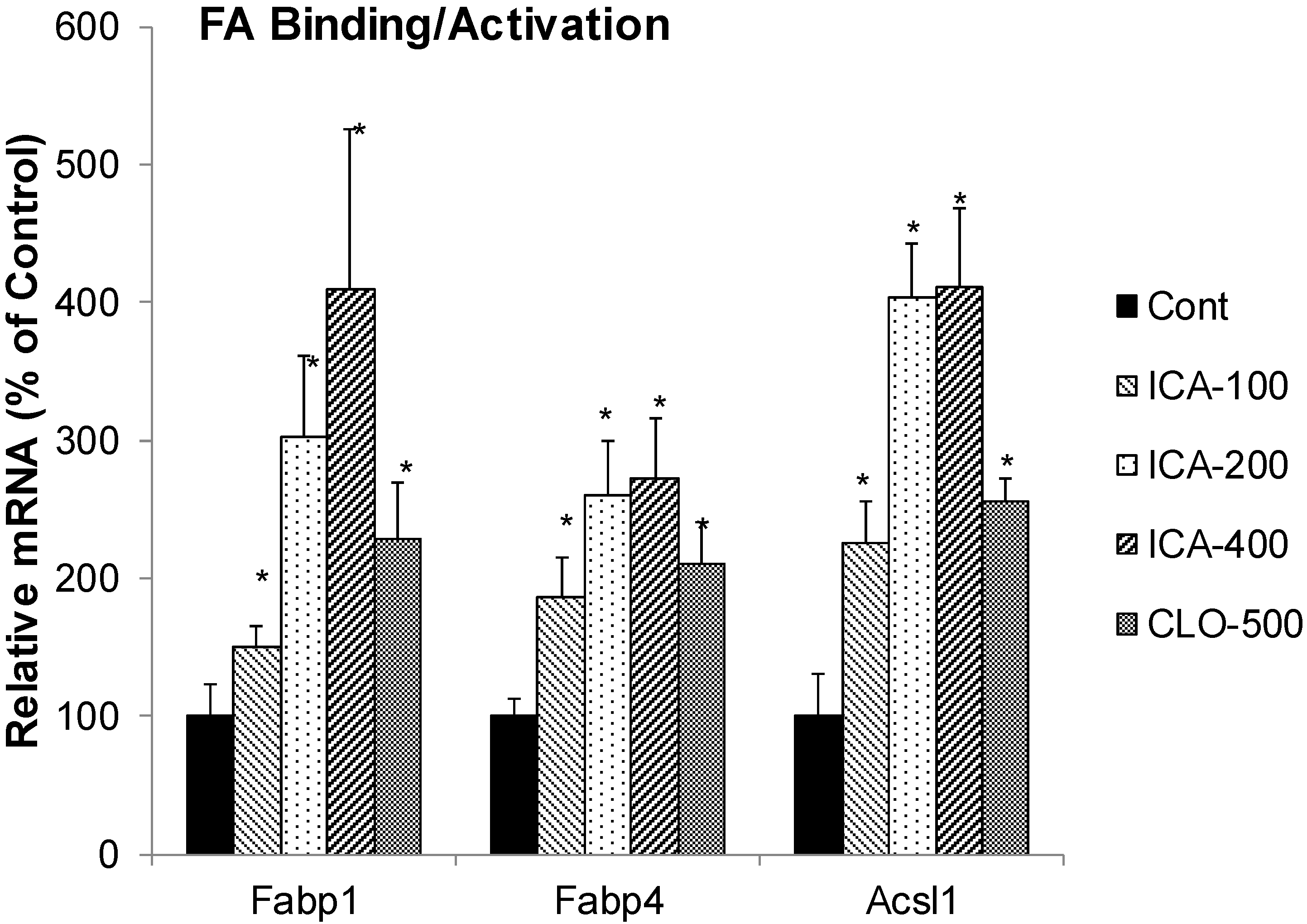
2.2. Effect of Icariin on the mRNA of Fatty Acid Binding Protein and Activation

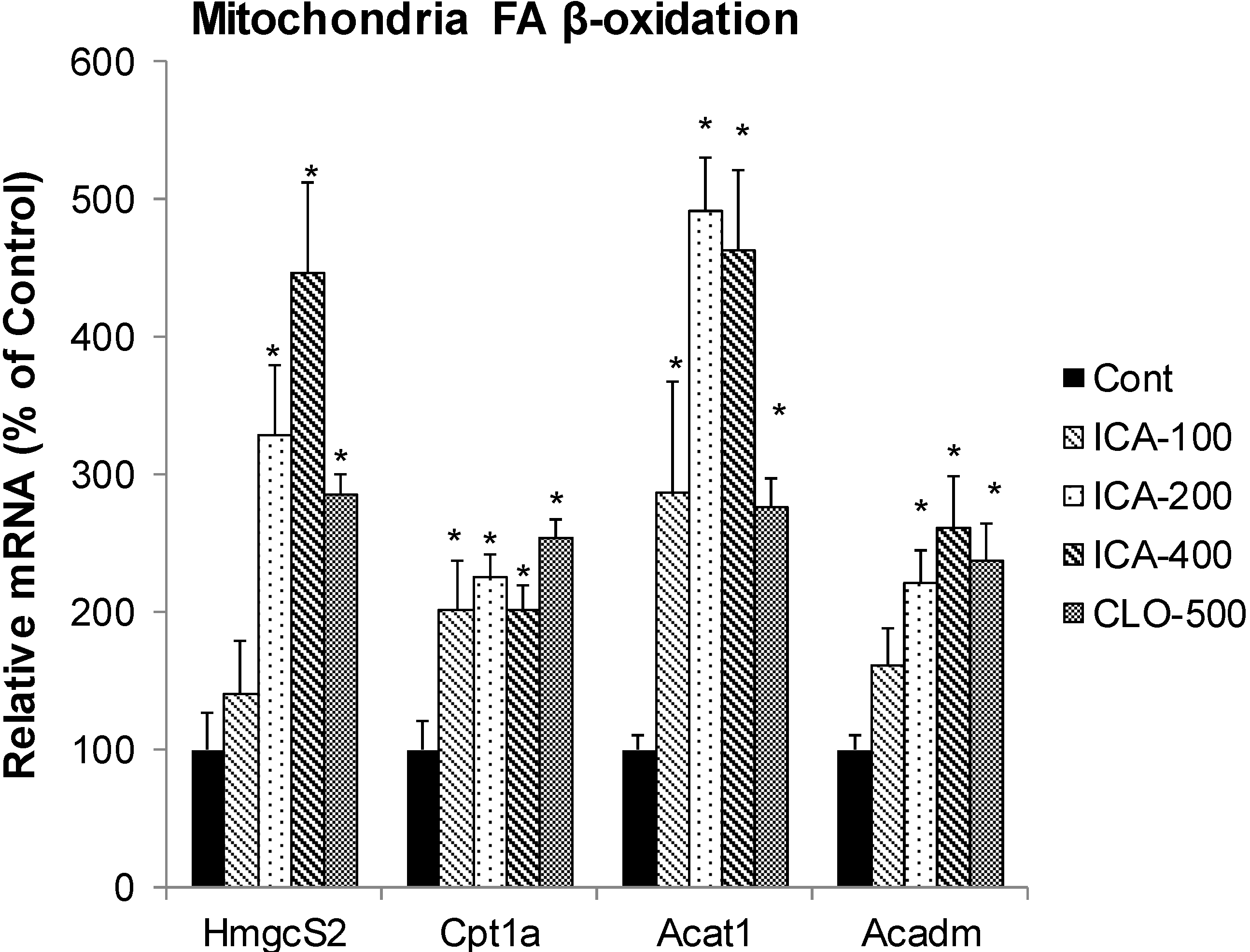
2.3. Effect of Icariin on the mRNA of Mitochondria FA β-Oxidation

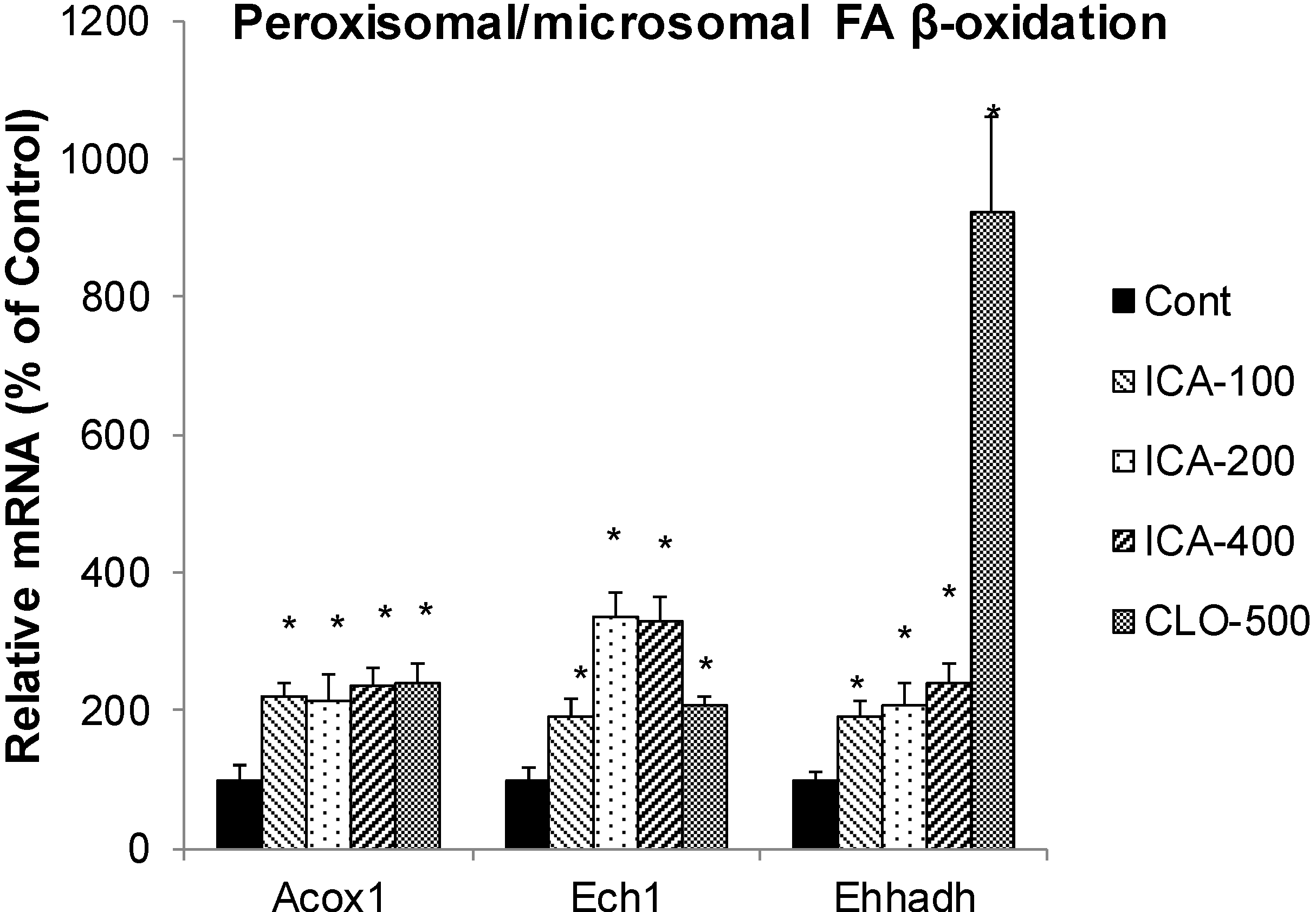
2.4. Effect of Icariin on the mRNA of Peroxisome FA Oxidation

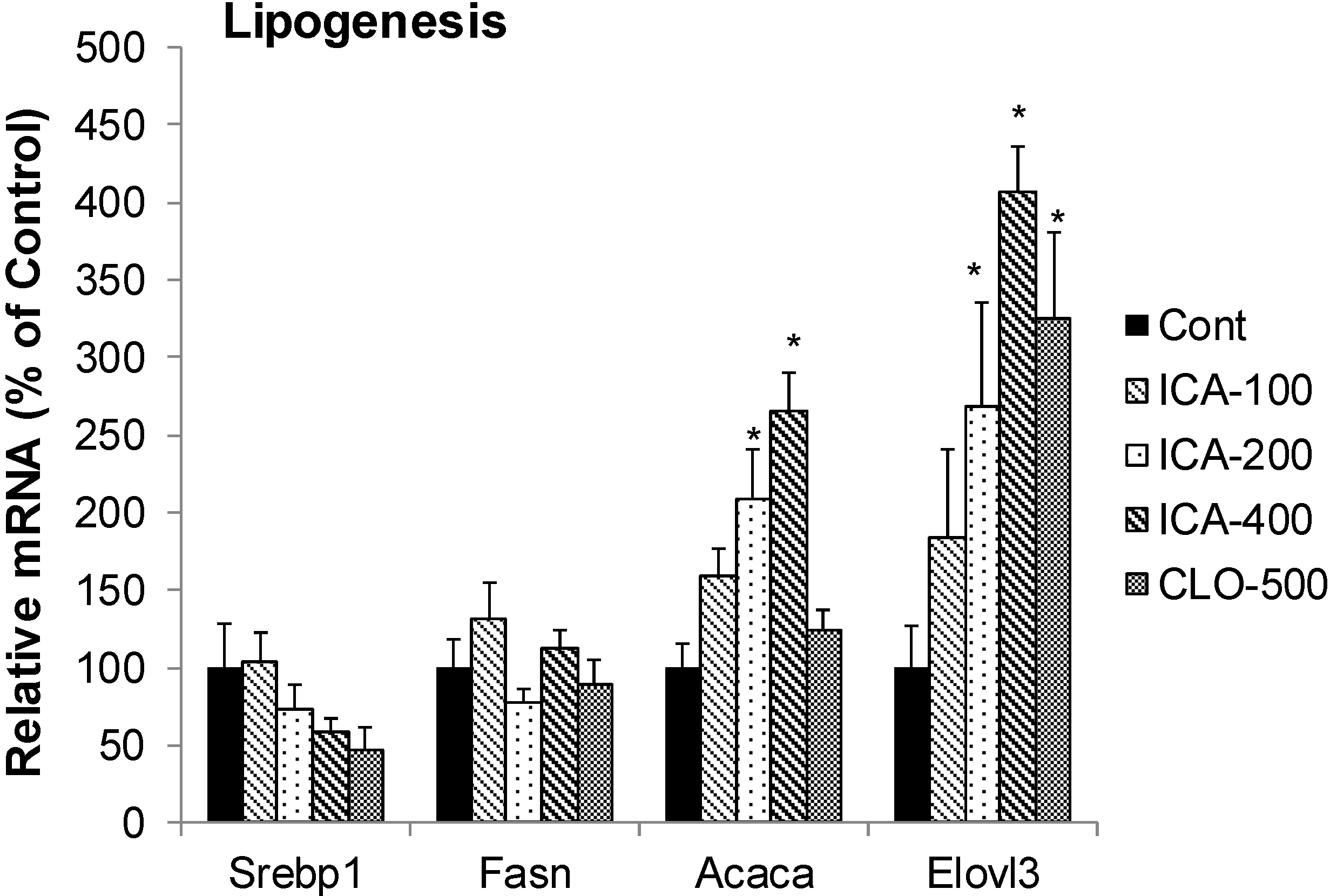
2.5. Effect of Icariin on the mRNA of Lipogenesis

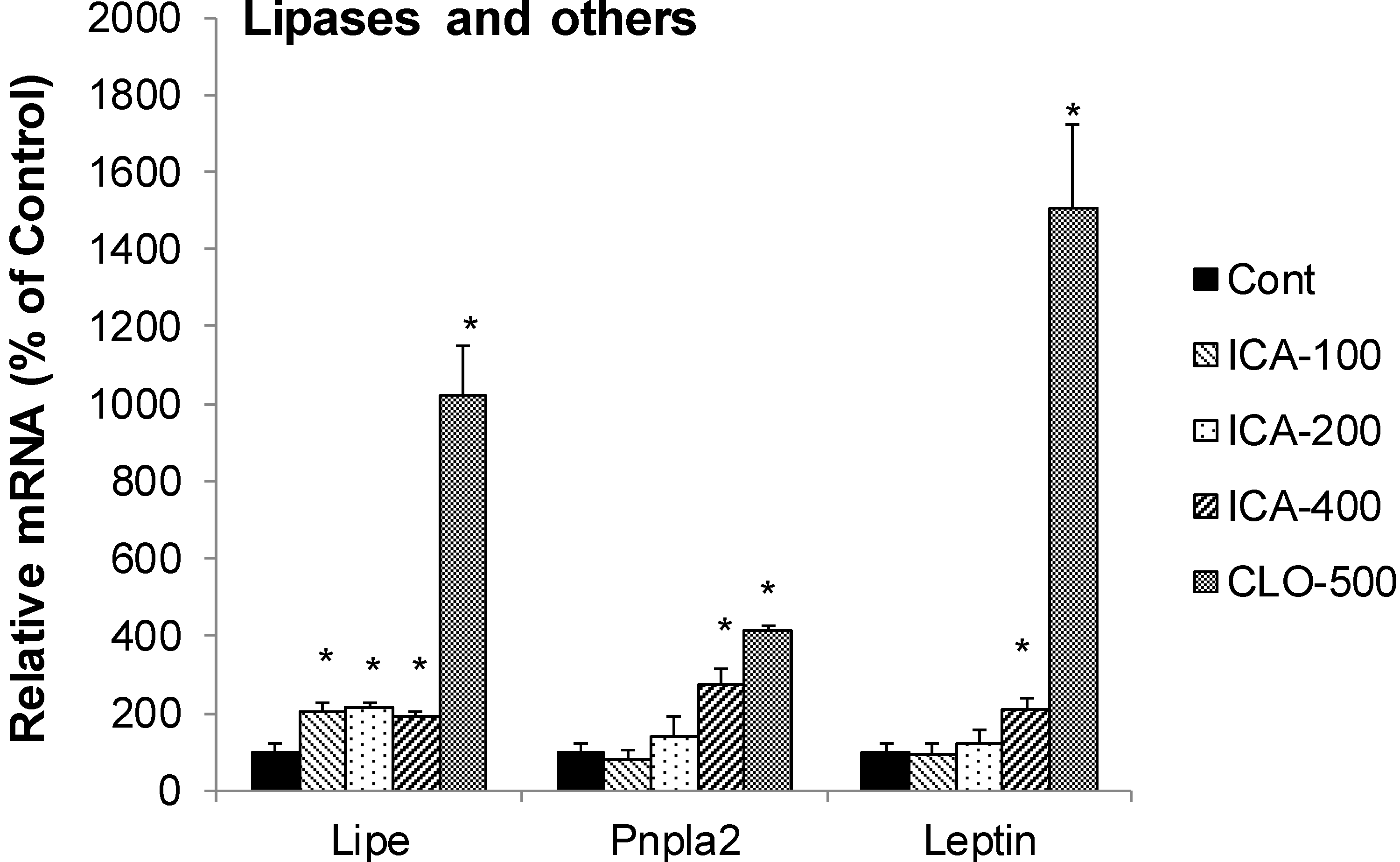
2.6. Effect of Icariin on the mRNA of Lipolysis

3. Experimental Section
3.1. Chemicals
3.2. Animals and Treatments
3.3. Animal General Health and Histopathology
3.4. RNA Isolation
3.5. Real-Time RT-PCR Analysis
3.6. Statistical Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | GenBank No. | Forward | Reverse |
|---|---|---|---|
| Acaca | NM_133360 | AGAAACCCGAACAGTGGAACT | AGGTAGCCCTTCACGGTTAAA |
| Acadm | NM_007382 | GAGCCCGGATTAGGGTTTAG | TCCCCGCTTTTGTCATATTC |
| Acat1 | NM_009230 | TCGATGACTTTGTGACCAACC | TCCACTTCAAACAGCTCGTCT |
| Acox1 | NM_015729 | CTTGGATGGTAGTCCGGAGA | TGGCTTCGAGTGAGGAAGTT |
| Acsl | NM_007981 | CAGGCAAAGCATGTCTTCAA | TCCCAGATTTTTGGCTTGTC |
| Cpt1a | NM_013495 | TGATGACGGCTATGGTGTTTC | CAAACAAGGTGATAATGTCCATC |
| Cyp410 | NM_010011 | CACACCCTGATCACCAACAG | TCCTTGATGCACATTGTGGT |
| Cyp4a14 | NM_007822 | CTGGGTGATGGAACCTCTGT | CATCTGGGAAGGTGACAGGT |
| Ech1 | NM_016772 | CGCGATGACAGTTTCCAGTA | TGTGCAGAGGAGCTCAGAGA |
| Ehhadh | NM_023737 | TGGATGTGGATGACATTGCT | GGGGAAGAGTATCGGCTAGG |
| Elovl3 | NM_007703 | GAGACCCTGCTTCCCTATCC | CTCCTTCCCTTCATCCTTCC |
| Fabp1 | NM_017399 | ATGAACTTCTCCGGCAAGTAC | ACTTTTTCCCCAGTCATGGTC |
| Fabp4 | NM_024406 | TCACCTGGAAGACAGCTCCT | AAGCCCACTCCCACTTCTTT |
| Fasn | NM_007988 | GACCTTCATGGACACAATGCT | ATACCACCAGAGACCGTTATG |
| GAPDH | M32599 | AACTTTGGCATTGTGGAAGG | GGATGCAGGGATGATGTTCT |
| Hmgcs2 | NM_008256 | CCTCTGTGAATCCTGGGTGT | CTGTGGGGAAAGATCTGCAT |
| Lipe | NM_010719 | ACGCTACACAAAGGCTGCTT | TCGTTGCGTTTGTAGTGCTC |
| Leptin | NM_008493 | CTATGCCACCTTGGTCACCT | ACCAAACCAAGCATTTTTGC |
| Pnpla2 | NM_025802 | TCATGTGACACAGCGAGTGA | AAAGAGGAGCCAAGCACAAA |
| PPARα | NM_011144 | GTCCTCAGTGCTTCCAGAGG | GGTCACCTACGAGTGGCATT |
| Srebp1 | AF374266 | GTGAGCCTGACAAGCAATCA | GGTGCCTACAGAGCAAGAGG |
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fruchart, J.C. Peroxisome proliferator-activated receptor-alpha (PPARalpha): At the crossroads of obesity, diabetes and cardiovascular disease. Atherosclerosis 2009, 205, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Seok, H.; Cha, B.S. Refocusing peroxisome proliferator activated receptor-α: A new insight for therapeutic roles in diabetes. Diabetes Metab. J. 2013, 37, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Duval, C.; Müller, M.; Kersten, S. PPARalpha and dyslipidemia. Biochim. Biophys. Acta 2007, 1771, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Costet, P.; Legendre, C.; Moré, J.; Edgar, A.; Galtier, P.; Pineau, T. Peroxisome proliferator-activated receptor alpha-isoform deficiency leads to progressive dyslipidemia with sexually dimorphic obesity and steatosis. J. Biol. Chem. 1998, 273, 29577–29585. [Google Scholar] [CrossRef] [PubMed]
- Abdelmegeed, M.A.; Yoo, S.H.; Henderson, L.E.; Gonzalez, F.J.; Woodcroft, K.J.; Song, B.J. PPARalpha expression protects male mice from high fat-induced nonalcoholic fatty liver. J. Nutr. 2011, 141, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Seedorf, U.; Assmann, G. The role of PPAR alpha in obesity. Nutr. Metab. Cardiovasc. Dis. 2001, 11, 189–194. [Google Scholar] [PubMed]
- Gervois, P.; Fruchart, J.C.; Staels, B. Inflammation, dyslipidaemia, diabetes and PPars: Pharmacological interest of dual PPARalpha and PPARgamma agonists. Int. J. Clin. Pract. Suppl. 2004, 143, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Sze, S.C.; Tong, Y.; Ng, T.B.; Cheng, C.L.; Cheung, H.P. Herba epimedii: Anti-oxidative properties and its medical implications. Molecules 2010, 15, 7861–7670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapoor, S. Icariin and its emerging role in the treatment of osteoporosis. Chin. Med. J. 2013, 126, 400–404. [Google Scholar] [PubMed]
- Li, L.; Zhou, Q.X.; Shi, J.S. Protective effects of icariin on neurons injured by cerebral ischemia/reperfusion. Chin. Med. J. 2005, 118, 1637–1643. [Google Scholar] [PubMed]
- Luo, Y.; Nie, J.; Gong, Q.H.; Lu, Y.F.; Wu, Q.; Shi, J.S. Protective effects of icariin against learning and memory deficits induced by aluminium in rats. Clin. Exp. Pharmacol. Physiol. 2007, 34, 792–795. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.X; Wu, Q.; Luo, Y.; Gong, Q.H.; Yu, L.M.; Huang, X.N.; Sun, A.S.; Shi, J.S. Protective effects of icariin on cognitive deficits induced by chronic cerebral hypoperfusion in rats. Clin. Exp. Pharmacol. Physiol. 2009, 36, 810–815. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Gong, Q.H.; Wu, Q.; Lu, Y.F.; Shi, J.S. Icariin isolated from epimedium brevicornum maxim attenuates learning and memory deficits induced by d-galactose in rats. Pharmacol. Biochem. Behav. 2010, 96, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Nie, J.; Luo, Y.; Huang, X.N.; Gong, Q.H.; Wu, Q.; Shi, J.S. Icariin inhibits beta-amyloid peptide segment 25–35 induced expression of beta-secretase in rat hippocampus. Eur. J. Pharmacol. 2010, 626, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Li, F.; Wu, Q.; Gong, Q.; Lu, Y.; Shi, J. Protective effects of icariin on brain dysfunction induced by lipopolysaccharide in rats. Phytomedicine 2010, 17, 950–955. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.; Gong, Q.H.; Xu, Y.S.; Wang, L.N.; Jin, H.; Li, F.; Li, L.S.; Ma, Y.M.; Shi, J.S. Icariin, a phoshphodiesterase-5 inhibitor, improves learning and memory in APP/PS1 transgenic mice by stimulation of NO/cGMP signalling. Int. J. Neuropsychopharmacol. 2014, 11, 1–11. [Google Scholar]
- Gong, Y.; Shi, J.; Xie, G.Y.; Liu, H.R.; Qi, M.Y. Amelioration of icariin for the epididymis impairment induced by streptozocin (STZ) in rats. Chin. J. Appl. Physiol. 2013, 29, 47–50. [Google Scholar]
- Zhang, W.P.; Bai, X.J.; Zheng, X.P.; Xie, X.L.; Yuan, Z.Y. Icariin attenuates the enhanced prothrombotic state in atherosclerotic rabbits independently of its lipid-lowering effects. Planta Med. 2013, 79, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Zhou, F.; Liu, T.; Li, G.Y.; Liu, J.; Gao, Z.Z.; Bai, G.Y.; Lu, H.; Xin, Z.C. Icariin ameliorates streptozotocin-induced diabetic retinopathy in vitro and in vivo. Int. J. Mol. Sci. 2012, 13, 866–878. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.Y.; Chen, K.; Liu, H.R.; Su, Y.H.; Yu, S.Q. Protective effect of Icariin on the early stage of experimental diabetic nephropathy induced by streptozotocin via modulating transforming growth factor β1 and type IV collagen expression in rats. J. Ethnopharmacol. 2011, 138, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Liang, X.G.; Zhu, D.Y.; Lou, Y.J. Icariin promotes expression of PGC-1alpha, PPARalpha, and NRF-1 during cardiomyocyte differentiation of murine embryonic stem cells in vitro. Acta Pharmacol. Sin. 2007, 28, 1541–1549. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.F.; Jin, T.; Lu, Y.F.; Li, L.S.; Liu, J.; Shi, J.S. Effect of icariin on UDP-glucuronosyltransferases in mouse liver. Planta Med. 2014, 80, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Patsouris, D.; Reddy, J.K.; Müller, M.; Kersten, S. Peroxisome proliferator-activated receptor alpha mediates the effects of high-fat diet on hepatic gene expression. Endocrinology 2006, 147, 1508–1516. [Google Scholar] [CrossRef] [PubMed]
- Klaassen, C.D. Learning to program the liver. Annu. Rev. Pharmacol. Toxicol. 2014, 54, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, C.; Benet, M.; Pisonero-Vaquero, S.; Moya, M.; García-Mediavilla, M.V.; Martínez-Chantar, M.L.; González-Gallego, J.; Castell, J.V.; Sánchez-Campos, S.; Jover, R. The human liver fatty acid binding protein (FABP1) gene is activated by FOXA1 and PPARα; and repressed by C/EBPα: Implications in FABP1 down regulation in nonalcoholic fatty liver disease. Biochim. Biophys. Acta 2013, 1831, 803–818. [Google Scholar] [CrossRef] [PubMed]
- Petrescu, A.D.; Huang, H.; Martin, G.G.; McIntosh, A.L.; Storey, S.M.; Landrock, D.; Kier, A.B.; Schroeder, F. Impact of L-FABP and glucose on polyunsaturated fatty acid induction of PPARα-regulated β-oxidative enzymes. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G241–G256. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, A.L.; Atshaves, B.P.; Landrock, D.; Landrock, K.K.; Martin, G.G.; Storey, S.M.; Kier, A.B.; Schroeder, F. Liver fatty acid binding protein gene-ablation exacerbates weight gain in high-fat fed female mice. Lipids 2013, 48, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Krammer, J.; Digel, M.; Ehehalt, F.; Stremmel, W.; Füllekrug, J.; Ehehalt, R. Overexpression of CD36 and acyl-CoA synthetases FATP2, FATP4 and ACSL1 increases fatty acid uptake in human hepatoma cells. Int. J. Med. Sci. 2011, 8, 599–614. [Google Scholar] [CrossRef] [PubMed]
- Thering, B.J.; Bionaz, M.; Loor, J.J. Long-chain fatty acid effects on peroxisome proliferator-activated receptor-alpha-regulated genes in Madin-Darby bovine kidney cells: Optimization of culture conditions using palmitate. J. Dairy Sci. 2009, 92, 2027–2037. [Google Scholar] [CrossRef] [PubMed]
- Rakhshandehroo, M.; Sanderson, L.M.; Matilainen, M.; Stienstra, R.; Carlberg, C.; de Groot, P.J.; Müller, M.; Kersten, S. Comprehensive analysis of PPARalpha-dependent regulation of hepatic lipid metabolism by expression profiling. PPAR Res. 2007, 26839. [Google Scholar] [CrossRef]
- Lee, K.; Kerner, J.; Hoppel, C.L. Mitochondrial carnitine palmitoyltransferase 1a (CPT1a) is part of an outer membrane fatty acid transfer complex. J. Biol. Chem. 2011, 286, 25655–25662. [Google Scholar] [CrossRef] [PubMed]
- Puisac, B.; Ramos, M.; Arnedo, M.; Menao, S.; Gil-Rodríguez, M.C.; Teresa-Rodrigo, M.E.; Pié, A.; de Karam, J.C.; Wesselink, J.J.; Giménez, I.; et al. Characterization of splice variants of the genes encoding human mitochondrial HMG-CoA lyase and HMG-CoA synthase, the main enzymes of the ketogenesis pathway. Mol. Biol. Rep. 2012, 39, 4777–4785. [Google Scholar] [CrossRef] [PubMed]
- Kostiuk, M.A.; Keller, B.O.; Berthiaume, L.G. Palmitoylation of ketogenic enzyme HMGCS2 enhances its interaction with PPARalpha and transcription at the Hmgcs2 PPRE. FASEB J. 2010, 24, 1914–1924. [Google Scholar] [CrossRef] [PubMed]
- Still, A.J.; Floyd, B.J.; Hebert, A.S.; Bingman, C.A.; Carson, J.J.; Gunderson, D.R.; Dolan, B.K.; Grimsrud, P.A.; Dittenhafer-Reed, K.E.; Stapleton, D.S.; et al. Quantification of mitochondrial acetylation dynamics highlights prominent sites of metabolic regulation. J. Biol. Chem. 2013, 288, 26209–26219. [Google Scholar] [CrossRef] [PubMed]
- Harano, Y.; Yasui, K.; Toyama, T.; Nakajima, T.; Mitsuyoshi, H.; Mimani, M.; Hirasawa, T.; Itoh, Y.; Okanoue, T. Fenofibrate, a peroxisome proliferator-activated receptor alpha agonist, reduces hepatic steatosis and lipid peroxidation in fatty liver Shionogi mice with hereditary fatty liver. Liver Int. 2006, 26, 613–620. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, B.; Giancristofaro, A.; Ammazzalorso, A.; D’Angelo, A.; Fantacuzzi, M.; Giampietro, L.; Maccallini, C.; Petruzzelli, M.; Amoroso, R. Discovery of gemfibrozil analogues that activate PPARα and enhance the expression of gene CPT1A involved in fatty acids catabolism. Eur. J. Med. Chem. 2011, 46, 5218–5224. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; McIntosh, A.L.; Martin, G.G.; Petrescu, A.D.; Landrock, K.K.; Landrock, D.; Kier, A.B.; Schroeder, F. Inhibitors of Fatty Acid Synthesis Induce PPAR α-Regulated fatty acid β-oxidative genes: Synergistic roles of L-FABP and glucose. PPAR Res. 2013, 2013, 865604. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Shi, L.; Zhang, H.; Hu, Z.; Wang, C.; Zhang, D.; Song, G. Oxymatrine ameliorates non-alcoholic fatty liver disease in rats through peroxisome proliferator-activated receptor-α activation. Mol. Med. Rep. 2013, 8, 439–445. [Google Scholar] [PubMed]
- Xie, W.D.; Wang, H.; Zhang, J.F.; Li, J.N.; Can, Y.; Qing, L.; Kung, H.F.; Zhang, Y.O. Enhanced peroxisomal β-oxidation metabolism in visceral adipose tissues of high-fat diet-fed obesity-resistant C57BL/6 mice. Exp. Ther. Med. 2011, 2, 309–315. [Google Scholar] [PubMed]
- Huang, J.; Zhang, Y.; Zhou, Y.; Zhang, Z.; Xie, Z.; Zhang, J.; Wan, X. Green tea polyphenols alleviate obesity in broiler chickens through the regulation of lipid-metabolism-related genes and transcription factor expression. J. Agric. Food Chem. 2013, 61, 8565–8572. [Google Scholar] [CrossRef] [PubMed]
- Houten, S.M.; Denis, S.; Argmann, C.A.; Jia, Y.; Ferdinandusse, S.; Reddy, J.K.; Wanders, R.J. Peroxisomal L-bifunctional enzyme (Ehhadh) is essential for the production of medium-chain dicarboxylic acids. J. Lipid Res. 2012, 53, 1296–1303. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Lee, K.T.; Iseli, T.J.; Hoy, A.J.; George, J.; Grewal, T.; Roufogalis, B.D. Compound K modulates fatty acid-induced lipid droplet formation and expression of proteins involved in lipid metabolism in hepatocytes. Liver Int. 2013, 33, 1583–1593. [Google Scholar] [PubMed]
- Yu, H.L.; Miao, H.T.; Gao, L.F.; Li, L.; Xi, Y.D.; Nie, S.P.; Xiao, R. Adaptive responses by mouse fetus to a maternal HLE diet by downregulating SREBP1: A microarray- and bio-analytic-based study. J. Lipid Res. 2013, 54, 3269–80. [Google Scholar] [CrossRef] [PubMed]
- Jakobsson, A.; Jörgensen, J.A.; Jacobsson, A. Differential regulation of fatty acid elongation enzymes in brown adipocytes implies a unique role for Elovl3 during increased fatty acid oxidation. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E517–E526. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.C.; Chen, Y.F.; Hsu, W.H.; Yang, C.W.; Kao, C.H.; Tsai, T.F. Resveratrol helps recovery from fatty liver and protects against hepatocellular carcinoma induced by hepatitis B virus X protein in a mouse model. Cancer Prev. Res. 2012, 5, 952–962. [Google Scholar] [CrossRef]
- Quan, H.Y.; Kim, D.Y.; Kim, S.J.; Jo, H.K.; Kim, G.W.; Chung, S.H. Betulinic acid alleviates non-alcoholic fatty liver by inhibiting SREBP1 activity via the AMPK-mTOR-SREBP signaling pathway. Biochem. Pharmacol. 2013, 85, 1330–1340. [Google Scholar] [CrossRef] [PubMed]
- Murugesan, S.; Goldberg, E.B.; Dou, E.; Brown, W.J. Identification of diverse lipid droplet targeting motifs in the PNPLA family of triglyceride lipases. PLoS One 2013, 8, e64950. [Google Scholar] [CrossRef] [PubMed]
- Jha, P.; Claudel, T.; Baghdasaryan, A.; Mueller, M.; Halilbasic, E.; Das, S.K.; Lass, A.; Zimmermann, R.; Zechner, R.; Hoefler, G.; et al. Role of adipose triglyceride lipase (PNPLA2) in protection from hepatic inflammation in mouse models of steatohepatitis and endotoxemia. Hepatology 2014, 59, 858–869. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the icarriin are available from the authors as well as from commercial resources (Sigma or Chinese Pharmaceutical companies).
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.-F.; Xu, Y.-Y.; Jin, F.; Wu, Q.; Shi, J.-S.; Liu, J. Icariin Is A PPARα Activator Inducing Lipid Metabolic Gene Expression in Mice. Molecules 2014, 19, 18179-18191. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules191118179
Lu Y-F, Xu Y-Y, Jin F, Wu Q, Shi J-S, Liu J. Icariin Is A PPARα Activator Inducing Lipid Metabolic Gene Expression in Mice. Molecules. 2014; 19(11):18179-18191. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules191118179
Chicago/Turabian StyleLu, Yuan-Fu, Yun-Yan Xu, Feng Jin, Qin Wu, Jing-Shan Shi, and Jie Liu. 2014. "Icariin Is A PPARα Activator Inducing Lipid Metabolic Gene Expression in Mice" Molecules 19, no. 11: 18179-18191. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules191118179