
Carbocations and the Complex Flavor and Bouquet of Wine: Mechanistic Aspects of Terpene Biosynthesis in Wine Grapes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
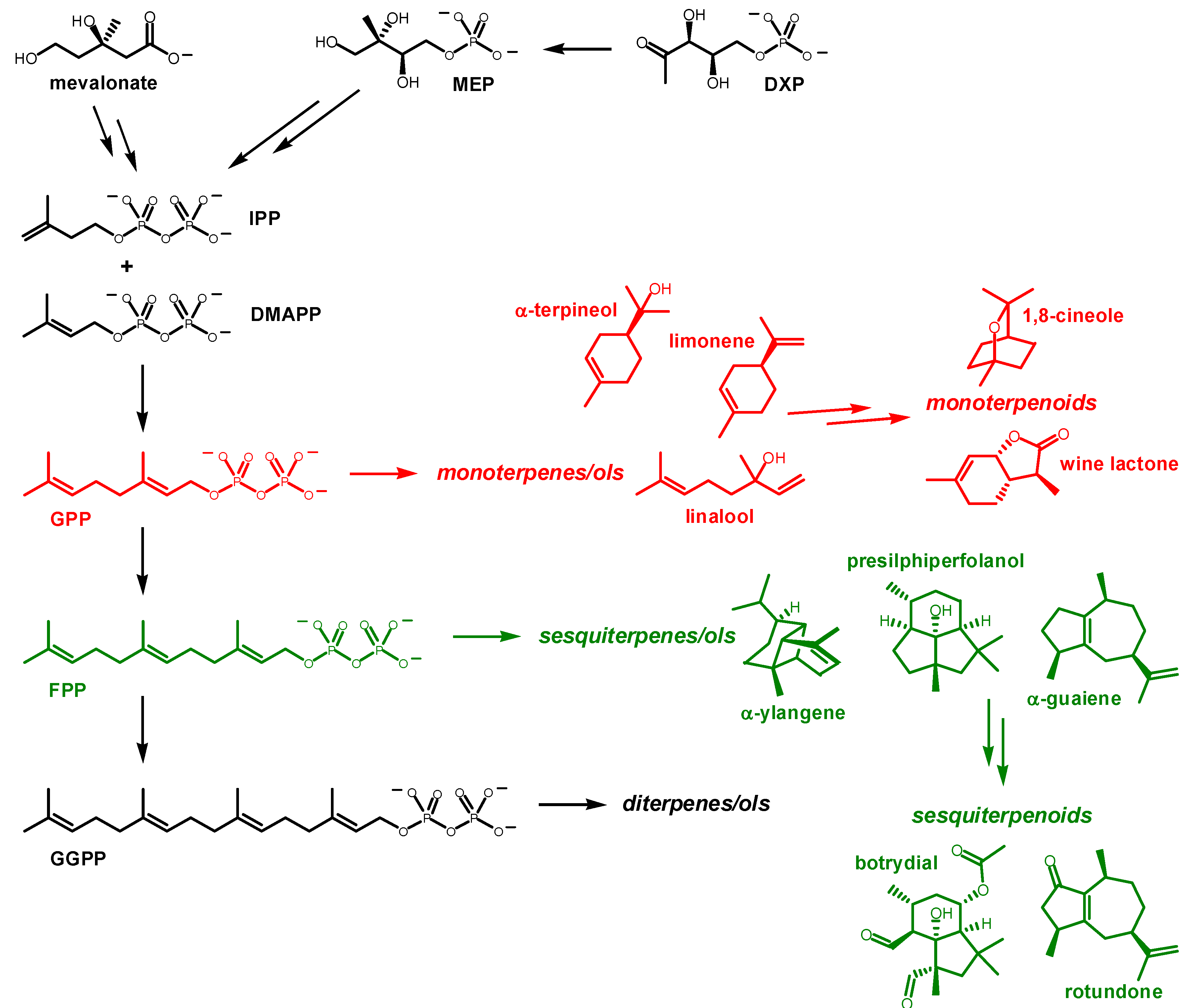
:1. Introduction

2. Representative Examples
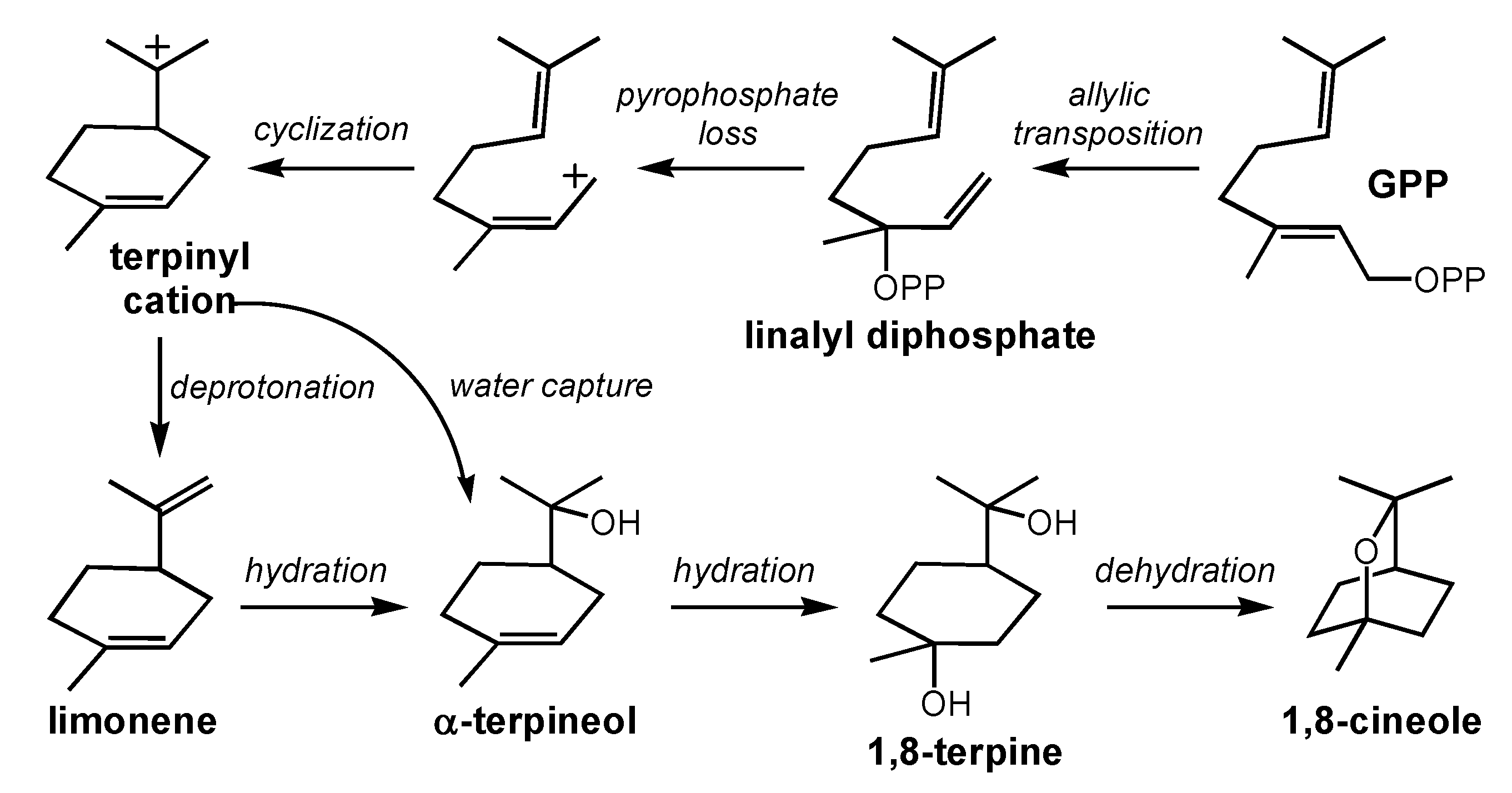
2.1. 1,8-Cineole

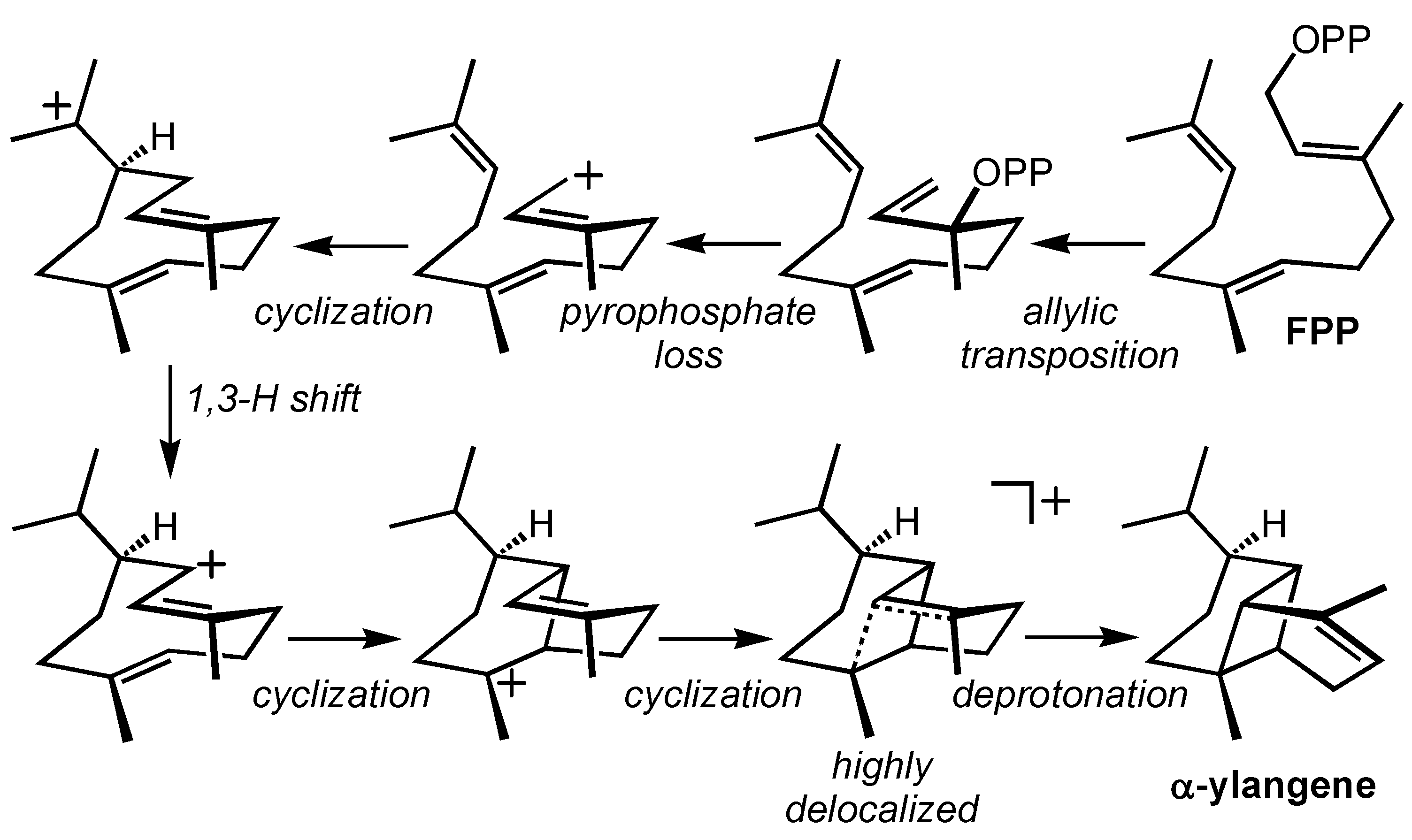
2.2. α-Ylangene

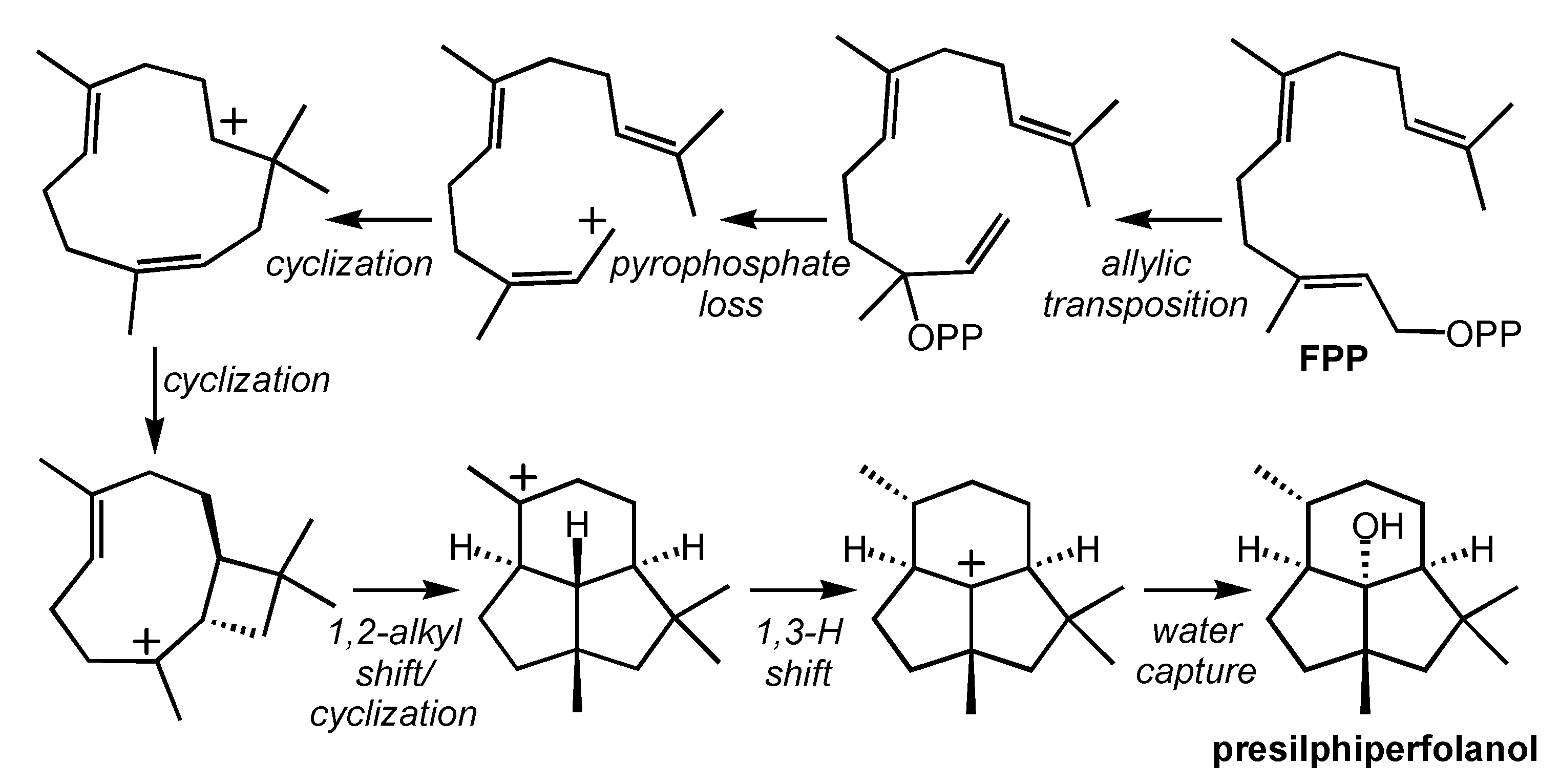
2.3. Botrydial

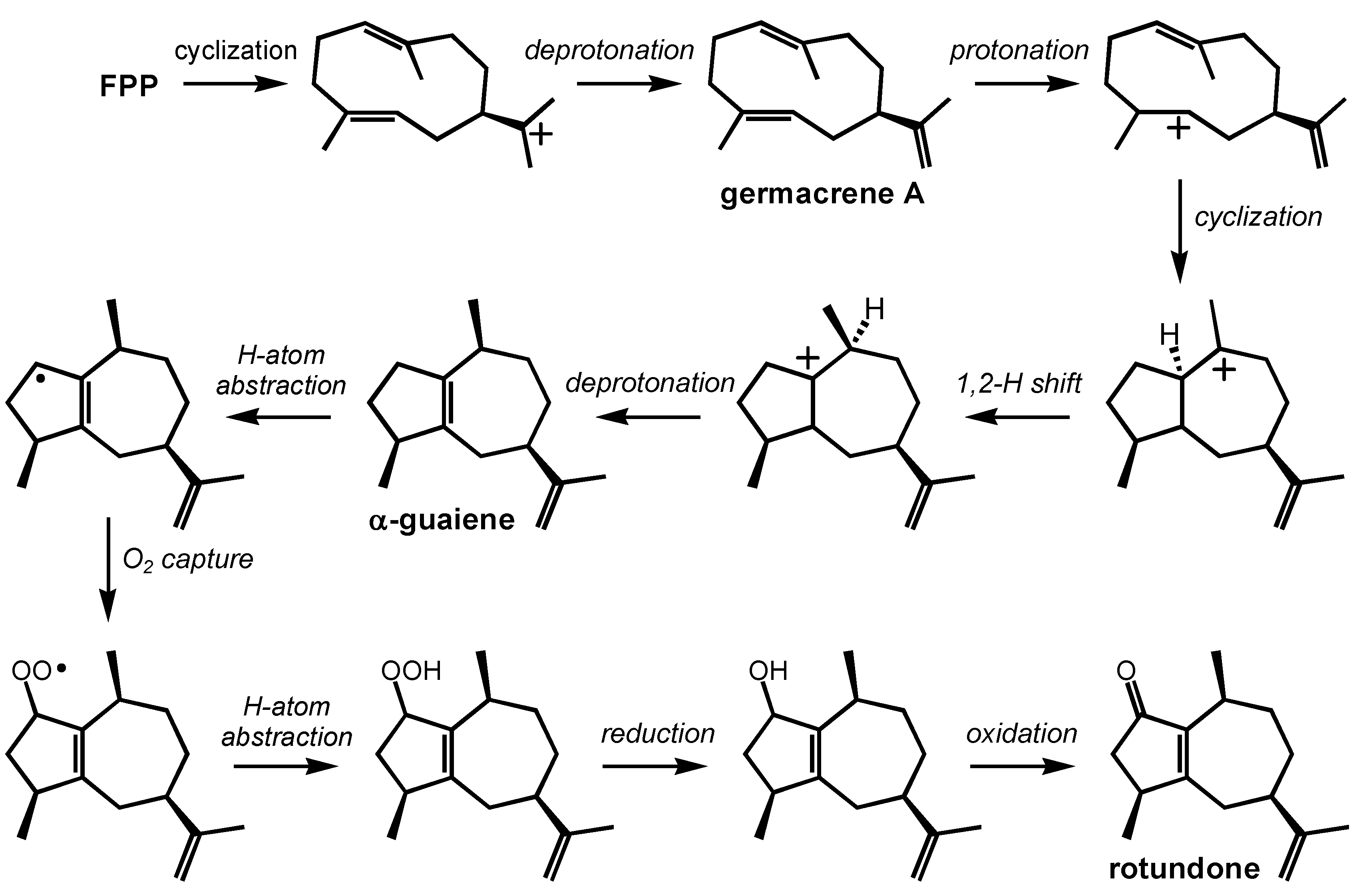
2.4. Rotundone

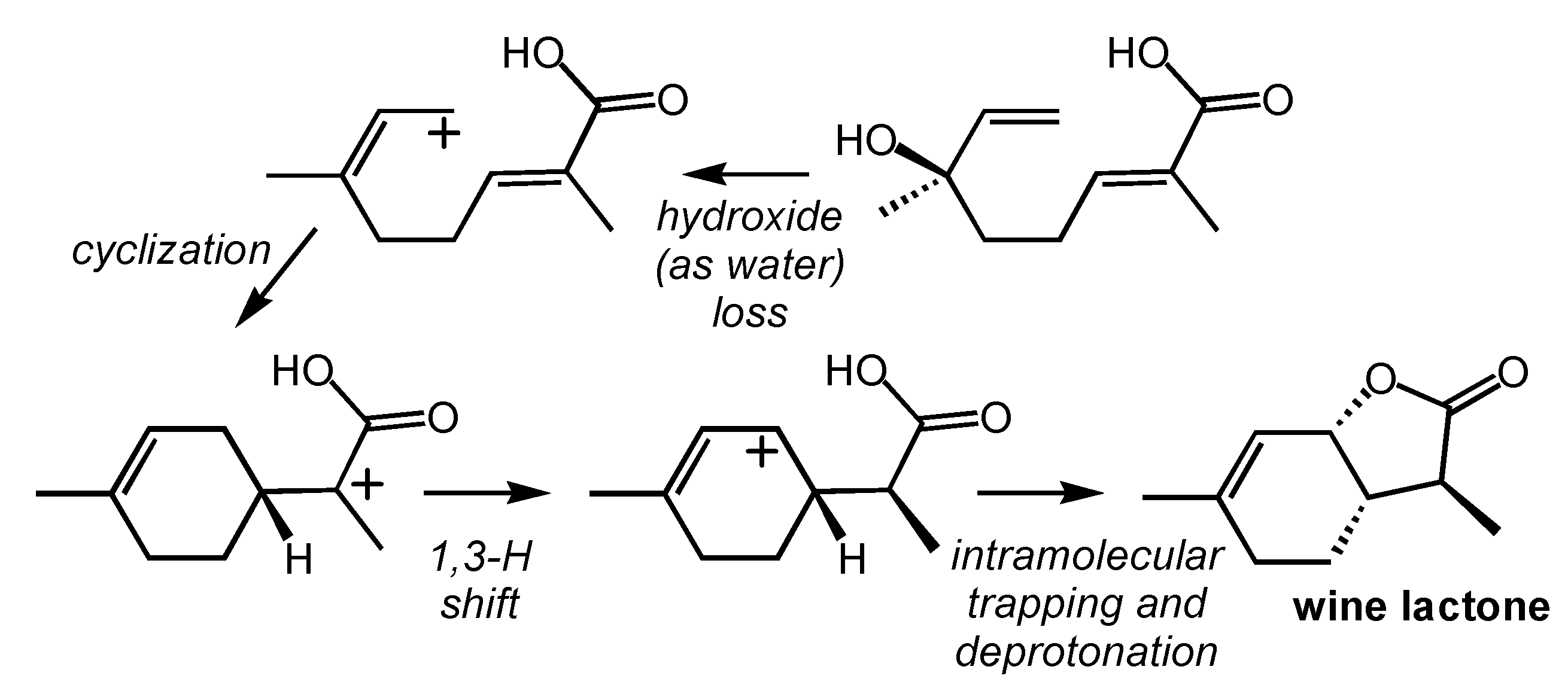
2.5. Wine Lactone

3. Conclusions and Outlook
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tantillo, D.J. Biosynthesis via carbocations: Theoretical studies on terpene formation. Nat. Prod. Rep. 2011, 28, 1035–1053. [Google Scholar] [CrossRef] [PubMed]
- Christianson, D. Structural biology and chemistry of the terpenoid cyclases. Chem. Rev. 2006, 106, 3412–3442. [Google Scholar] [CrossRef] [PubMed]
- Cane, D.E. Isoprenoids Including Carotenoids and Steroids. Compr. Nat. Prod. Chem. 1999, 2, 155–200. [Google Scholar]
- Cane, D.E. Enzymic formation of sesquiterpenes. Chem. Rev. 1990, 90, 1089–1103. [Google Scholar] [CrossRef]
- Davis, E.; Croteau, R. Cyclization enzymes in the biosynthesis of monoterpenes, sesquiterpenes, and diterpenes. In Biosynthesis; Leeper, F., Vederas, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; Volume 209, pp. 53–95. [Google Scholar]
- Gil, M.; Pontin, M.; Berli, F.; Bottini, R.; Piccoli, P. Metabolism of terpenes in the response of grape (Vitis vinifera L.) leaf tissues to UV-B radiation. Phytochemistry 2012, 77, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Lund, S.T.; Bohlmann, J. The Molecular Basis for Wine Grape Quality-A Volatile Subject. Science 2006, 311, 804–805. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.M.; Toub, O.; Chiang, A.; Lo, B.C.; Ohse, S.; Lund, S.T.; Bohlmann, J. The bouquet of grapevine (Vitis vinifera L. cv. Cabernet Sauvignon) flowers arises from the biosynthesis of sesquiterpene volatiles in pollen grains. Proc. Nat. Acad. Sci. USA 2009, 106, 7245–7250. [Google Scholar] [CrossRef] [PubMed]
- Lange, B.M.; Croteau, R. Isopentenyl diphosphate biosynthesis via a mevalonate-independent pathway: Isopentenyl monophosphate kinase catalyzes the terminal enzymatic step. Proc. Nat. Acad. Sci. USA 1999, 96, 13714–13719. [Google Scholar] [CrossRef] [PubMed]
- Eisenreich, W.; Schwarz, M.; Cartayrade, A.; Arigoni, D.; Zenk, M.H.; Bacher, A. The deoxyxylulose phosphate pathway of terpenoid biosynthesis in plants and microorganisms. Chem. Biol. 1998, 5, R221–R233. [Google Scholar] [CrossRef]
- Lange, B.M.; Rujan, T.; Martin, W.; Croteau, R. Isoprenoid biosynthesis: The evolution of two ancient and distinct pathways across genomes. Proc. Nat. Acad. Sci. USA 2000, 97, 13172–13177. [Google Scholar] [CrossRef] [PubMed]
- Sallaud, C.; Rontein, D.; Onillon, S.; Jabès, F.; Duffé, P.; Giacalone, C.; Thoraval, S.; Escoffier, C.; Herbette, G.; Leonhardt, N.; et al. A Novel Pathway for Sesquiterpene Biosynthesis from Z,Z-Farnesyl Pyrophosphate in the Wild Tomato Solanum habrochaites. Plant Cell 2009, 21, 301–317. [Google Scholar] [CrossRef] [PubMed]
- Nagegowda, D.A. Plant volatile terpenoid metabolism: Biosynthetic genes, transcriptional regulation and subcellular compartmentation. FEBS Lett. 2010, 584, 2965–2973. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.; Aubourg, S.; Schouwey, M.; Daviet, L.; Schalk, M.; Toub, O.; Lund, S.; Bohlmann, J. Functional Annotation, Genome Organization and Phylogeny of the Grapevine (Vitis vinifera) Terpene Synthase Gene Family Based on Genome Assembly, FLcDNA Cloning, and Enzyme Assays. BMC Plant Biol. 2010, 10, 226. [Google Scholar] [CrossRef] [PubMed]
- Jaillon, O.; Aury, J.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Marais, J. Terpenes in the Aroma of Grapes and Wines: A Review. S. Afr. J. Enol. Vitic. 1983, 4, 49–58. [Google Scholar]
- Villamor, R.R.; Ross, C.F. Wine Matrix Compounds Affect Perception of Wine Aromas. Annu. Rev. Food Sci. Technol. 2013, 4, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Capone, D.L.; Jeffery, D.W.; Sefton, M.A. Vineyard and Fermentation Studies To Elucidate the Origin of 1,8-Cineole in Australian Red Wine. J. Agric. Food Chem. 2012, 60, 2281–2287. [Google Scholar] [CrossRef] [PubMed]
- Fariña, L.; Boido, E.; Carrau, F.; Versini, G.; Dellacassa, E. Terpene Compounds as Possible Precursors of 1,8-Cineole in Red Grapes and Wines. J. Agric. Food Chem. 2005, 53, 1633–1636. [Google Scholar] [CrossRef] [PubMed]
- Park, S.K.; Noble, A.C. Monoterpenes and monoterpene glycosides in wine aromas. In Beer and Wine Production; American Chemical Society: Washington, DC, USA, 1993; Volume 536, pp. 98–109. [Google Scholar]
- Parker, M.; Pollnitz, A.P.; Cozzolino, D.; Francis, I.L.; Herderich, M.J. Identification and Quantification of a Marker Compound for “Pepper” Aroma and Flavor in Shiraz Grape Berries by Combination of Chemometrics and Gas Chromatography-Mass Spectrometry. J. Agric. Food Chem. 2007, 55, 5948–5955. [Google Scholar] [CrossRef] [PubMed]
- Pinedo, C.; Wang, C.M.; Pradier, J.M.; Dalmais, B.; Choquer, M.; le Pêcheur, P.; Morgant, G.; Collado, I.G.; Cane, D.E.; Viaud, M. Sesquiterpene Synthase from the Botrydial Biosynthetic Gene Cluster of the Phytopathogen Botrytis cinerea. ACS Chem. Biol. 2008, 3, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-M.; Hopson, R.; Lin, X.; Cane, D.E. Biosynthesis of the Sesquiterpene Botrydial in Botrytis cinerea. Mechanism and Stereochemistry of the Enzymatic Formation of Presilphiperfolan-8β-ol. J. Am. Chem. Soc. 2009, 131, 8360–8361. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Dunne, K.J.; Evans, K.J.; Barry, K.; Cadle-Davidson, L.; Wilcox, W.F. Optimisation of techniques for quantification of Botrytis cinerea in grape berries and receptacles by quantitative polymerase chain reaction. Aust. J. Grape Wine Res. 2013, 19, 68–73. [Google Scholar] [CrossRef]
- Cadle-Davidson, L. Monitoring Pathogenesis of Natural Botrytis cinerea Infections in Developing Grape Berries. Am. J. Enol. Vitic. 2008, 59, 387–395. [Google Scholar]
- Siebert, T.E.; Wood, C.; Elsey, G.M.; Pollnitz, A.P. Determination of Rotundone, the Pepper Aroma Impact Compound, in Grapes and Wine. J. Agric. Food Chem. 2008, 56, 3745–3748. [Google Scholar] [CrossRef] [PubMed]
- Caputi, L.; Carlin, S.; Ghiglieno, I.; Stefanini, M.; Valenti, L.; Vrhovsek, U.; Mattivi, F. Relationship of Changes in Rotundone Content during Grape Ripening and Winemaking to Manipulation of the “Peppery” Character of Wine. J. Agric. Food Chem. 2011, 59, 5565–5571. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.-C.; Burrett, S.; Sefton, M.A.; Taylor, D.K. Production of the Pepper Aroma Compound, (−)-Rotundone, by Aerial Oxidation of α-Guaiene. J. Agric. Food Chem. 2014, 62, 10809–10815. [Google Scholar] [CrossRef] [PubMed]
- Wood, C.; Siebert, T.E.; Parker, M.; Capone, D.L.; Elsey, G.M.; Pollnitz, A.P.; Eggers, M.; Meier, M.; Vössing, T.; Widder, S.; et al. From Wine to Pepper: Rotundone, an Obscure Sesquiterpene, Is a Potent Spicy Aroma Compound. J. Agric. Food Chem. 2008, 56, 3738–3744. [Google Scholar] [CrossRef] [PubMed]
- Barter, S.; Siebert, T.; Krstic, M.; Herderich, M.J.; Francis, L. Mapping the Origins of High Rotenone and Pepper Flavour in Shiraz. Available online: http://www.awri.com.au/information_services/ebulletin/2015/02/03/technical-review-february-2015/ (accessed on 10 June 2015).
- Pedersen, D.S.; Capone, D.; Skouroumounis, G.; Pollnitz, A.; Sefton, M. Quantitative analysis of geraniol, nerol, linalool, and α-terpineol in wine. Anal. Bioanal. Chem. 2003, 375, 517–522. [Google Scholar] [PubMed]
- Martin, D.; Chiang, A.; Lund, S.; Bohlmann, J. Biosynthesis of wine aroma: Transcript profiles of hydroxymethylbutenyl diphosphate reductase, geranyl diphosphate synthase, and linalool/nerolidol synthase parallel monoterpenol glycoside accumulation in Gewürztraminer grapes. Planta 2012, 236, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Bonnländer, B.; Baderschneider, B.; Messerer, M.; Winterhalter, P. Isolation of Two Novel Terpenoid Glucose Esters from Riesling Wine. J. Agric. Food Chem. 1998, 46, 1474–1478. [Google Scholar] [CrossRef]
- Guth, H. Determination of the Configuration of Wine Lactone. Helv. Chim. Acta 1996, 79, 1559–1571. [Google Scholar] [CrossRef]
- Tantillo, D.J. Recent excursions to the borderlands between the realms of concerted and stepwise: carbocation cascades in natural products biosynthesis. J. Phys. Org. Chem. 2008, 21, 561–570. [Google Scholar] [CrossRef]
- Tantillo, D.J. The carbocation continuum in terpene biosynthesis-where are the secondary cations? Chem. Soc. Rev. 2010, 39, 2847–2854. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.J.; Tantillo, D.J. How cyclobutanes are assembled in nature—Insights from quantum chemistry. Chem. Soc. Rev. 2014, 43, 5042–5050. [Google Scholar] [CrossRef] [PubMed]
- Siebert, M.R.; Zhang, J.; Addepalli, S.V.; Tantillo, D.J.; Hase, W.L. The Need for Enzymatic Steering in Abietic Acid Biosynthesis: Gas-Phase Chemical Dynamics Simulations of Carbocation Rearrangements on a Bifurcating Potential Energy Surface. J. Am. Chem. Soc. 2011, 133, 8335–8343. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.J.; Tantillo, D.J. Biosynthetic consequences of multiple sequential post-transition-state bifurcations. Nat. Chem. 2014, 6, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Pemberton, R.P.; Hong, Y.J.; Tantillo, D.J. Inherent Dynamical Preferences in Carbocation Rearrangements Leading to Terpene Natural Products. Pure Appl. Chem. 2013, 85, 1949–1957. [Google Scholar] [CrossRef]
- Pemberton, R.P.; Tantillo, D.J. Lifetimes of carbocations encountered along reaction coordinates for terpene formation. Chem. Sci. 2014, 5, 3301–3308. [Google Scholar] [CrossRef]
- Pemberton, R.P.; Ho, K.C.; Tantillo, D.J. Modulation of inherent dynamical tendencies of the bisabolyl cation via preorganization in epi-isozizaene synthase. Chem. Sci. 2015, 6, 2347–2353. [Google Scholar] [CrossRef]
- Zu, L.; Xu, M.; Lodewyk, M.W.; Cane, D.E.; Peters, R.J.; Tantillo, D.J. Effect of Isotopically Sensitive Branching on Product Distribution for Pentalenene Synthase: Support for a Mechanism Predicted by Quantum Chemistry. J. Am. Chem. Soc. 2012, 134, 11369–11371. [Google Scholar] [CrossRef] [PubMed]
- Tantillo, D.J. Walking in the woods with quantum chemistry—Applications of quantum chemical calculations in natural products research. Nat. Prod. Rep. 2013, 30, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Capone, D.L.; Van Leeuwen, K.; Taylor, D.K.; Jeffery, D.W.; Pardon, K.H.; Elsey, G.M.; Sefton, M.A. Evolution and Occurrence of 1,8-Cineole (Eucalyptol) in Australian Wine. J. Agric. Food Chem. 2011, 59, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Saliba, A.J.; Bullock, J.; Hardie, W.J. Consumer rejection threshold for 1,8-cineole (eucalyptol) in Australian red wine. Food Qual. Prefer. 2009, 20, 500–504. [Google Scholar] [CrossRef]
- Ebeler, S.E. Analytical Chemistry: Unlocking the Secrets of Wine Flavor. Food Rev. Intern. 2001, 17, 45–64. [Google Scholar] [CrossRef]
- Wise, M.L.; Urbansky, M.; Helms, G.L.; Coates, R.M.; Croteau, R. Syn Stereochemistry of Cyclic Ether Formation in 1,8-Cineole Biosynthesis Catalyzed by Recombinant Synthase from Salvia officinalis. J. Am. Chem. Soc. 2002, 124, 8546–8547. [Google Scholar] [CrossRef] [PubMed]
- Herderich, M.J.; Siebert, T.E.; Parker, M.; Hayasaka, Y.; Curtin, C.; Mercurio, M. Metabolomics and the quest for understanding quality in flavor chemistry and wine research. In Recent Advances in the Analysis of Food and Flavors; American Chemical Society: Washington, DC, USA, 2012; Volume 1098, pp. 1–11. [Google Scholar]
- Lodewyk, M.W.; Gutta, P.; Tantillo, D.J. Computational studies on biosynthetic carbocation rearrangements leading to sativene, cyclosativene, alpha-ylangene, and beta-ylangene. J. Org. Chem. 2008, 73, 6570–6579. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.C.; Tantillo, D.J. Prediction of a New Pathway to Presilphiperfolanol. Org. Lett. 2008, 10, 4827–4830. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.J.; Irmisch, S.; Wang, S.C.; Garms, S.; Gershenzon, J.; Zu, L.; Köllner, T.G.; Tantillo, D.J. Theoretical and Experimental Analysis of the Reaction Mechanism of MrTPS2, a Triquinane-Forming Sesquiterpene Synthase from Chamomile. Chem. Eur. J. 2013, 19, 13590–13600. [Google Scholar] [CrossRef] [PubMed]
- Luan, F.; Degenhardt, A.; Mosandl, A.; Wüst, M. Mechanism of Wine Lactone Formation: Demonstration of Stereoselective Cyclization and 1,3-Hydride Shift. J. Agric. Food Chem. 2006, 54, 10245–10252. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, O.; Harrison, J.G.; Felix, R.J.; Cortes Guzman, F.; Gagne, M.R.; Tantillo, D.J. Carbonium vs. carbenium ion-like transition state geometries for carbocation cyclization—How strain associated with bridging affects 5-exo vs. 6-endo selectivity. Chem. Sci. 2013, 4, 3894–3898. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.J.; Tantillo, D.J. Perturbing the Structure of the 2-Norbornyl Cation through C−H···N and C−H···π Interactions. J. Org. Chem. 2007, 72, 8877–8881. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.C. The energy of the transition states and the intermediate cation in the ionization of 2-norbornyl derivatives. Where is the nonclassical stabilization energy? Acc. Chem. Res. 1983, 16, 432–440. [Google Scholar] [CrossRef]
- Grob, C.A. Inductivity and bridging in carbocations. Acc. Chem. Res. 1983, 16, 426–431. [Google Scholar] [CrossRef]
- Walling, C. An innocent bystander looks at the 2-norbornyl cation. Acc. Chem. Res. 1983, 16, 448–454. [Google Scholar] [CrossRef]
- Schreiner, P.R.; Severance, D.L.; Jorgensen, W.L.; von Schleyer, P.; Schaefer, H.F. Energy Difference between the Classical and the Nonclassical 2-Norbornyl Cation in Solution. A Combined ab Initio-Monte Carlo Aqueous Solution Study. J. Am. Chem. Soc. 1995, 117, 2663–2664. [Google Scholar] [CrossRef]
- Sieber, S.; von Ragué Schleyer, P.; Vančik, H.; Mesić, M.; Sunko, D.E. The Nature of the 7-Norbornyl Cation and its Rearrangement into the 2-Norbornyl Cation. Angew. Chem. Int. Ed. 1993, 32, 1604–1606. [Google Scholar] [CrossRef]
- Laube, T. X-ray Crystal Structures of Carbocations Stabilized by Bridging or Hyperconjugation. Acc. Chem. Res. 1995, 28, 399–405. [Google Scholar] [CrossRef]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wedler, H.B.; Pemberton, R.P.; Tantillo, D.J. Carbocations and the Complex Flavor and Bouquet of Wine: Mechanistic Aspects of Terpene Biosynthesis in Wine Grapes. Molecules 2015, 20, 10781-10792. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules200610781
Wedler HB, Pemberton RP, Tantillo DJ. Carbocations and the Complex Flavor and Bouquet of Wine: Mechanistic Aspects of Terpene Biosynthesis in Wine Grapes. Molecules. 2015; 20(6):10781-10792. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules200610781
Chicago/Turabian StyleWedler, Henry B., Ryan P. Pemberton, and Dean J. Tantillo. 2015. "Carbocations and the Complex Flavor and Bouquet of Wine: Mechanistic Aspects of Terpene Biosynthesis in Wine Grapes" Molecules 20, no. 6: 10781-10792. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules200610781