
In Vitro Study on Anti-Hepatitis C Virus Activity of Spatholobus suberectus Dunn

 ,
,
Abstract
:1. Introduction
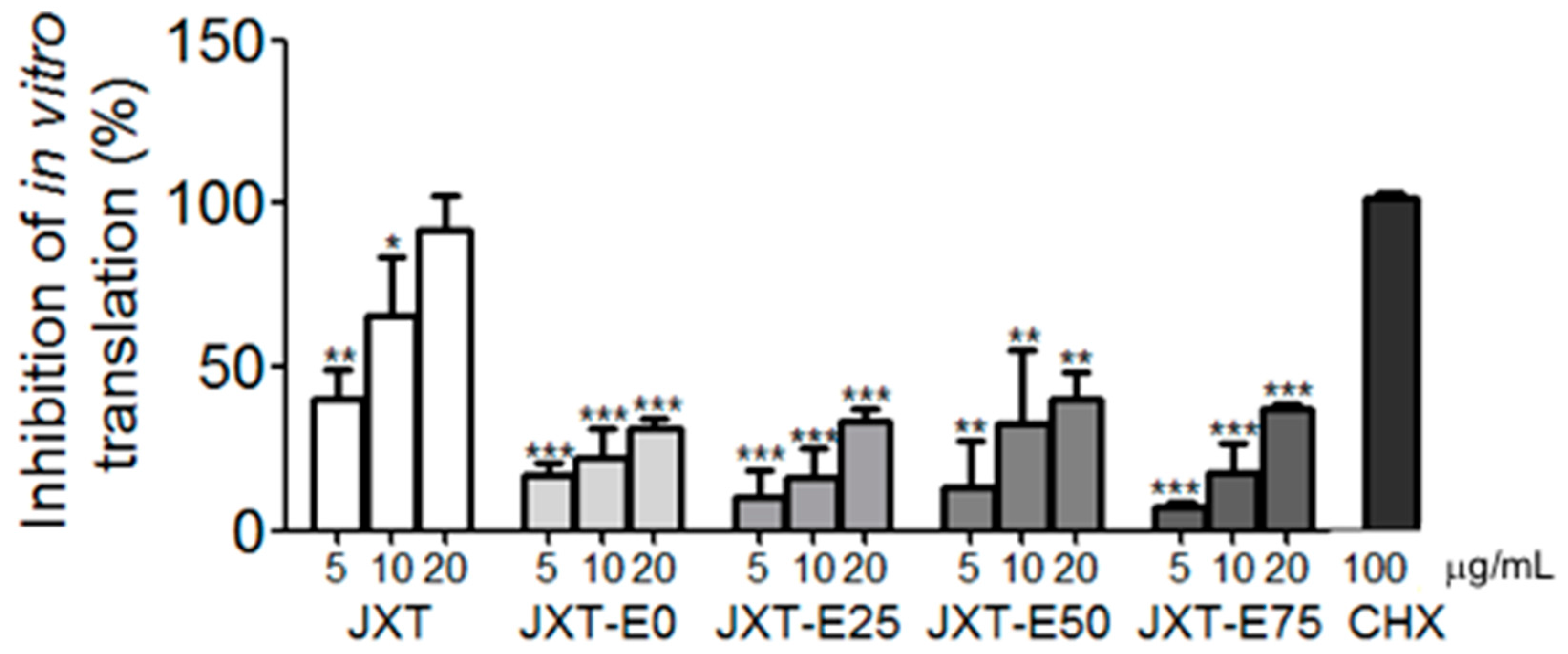
2. Results
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
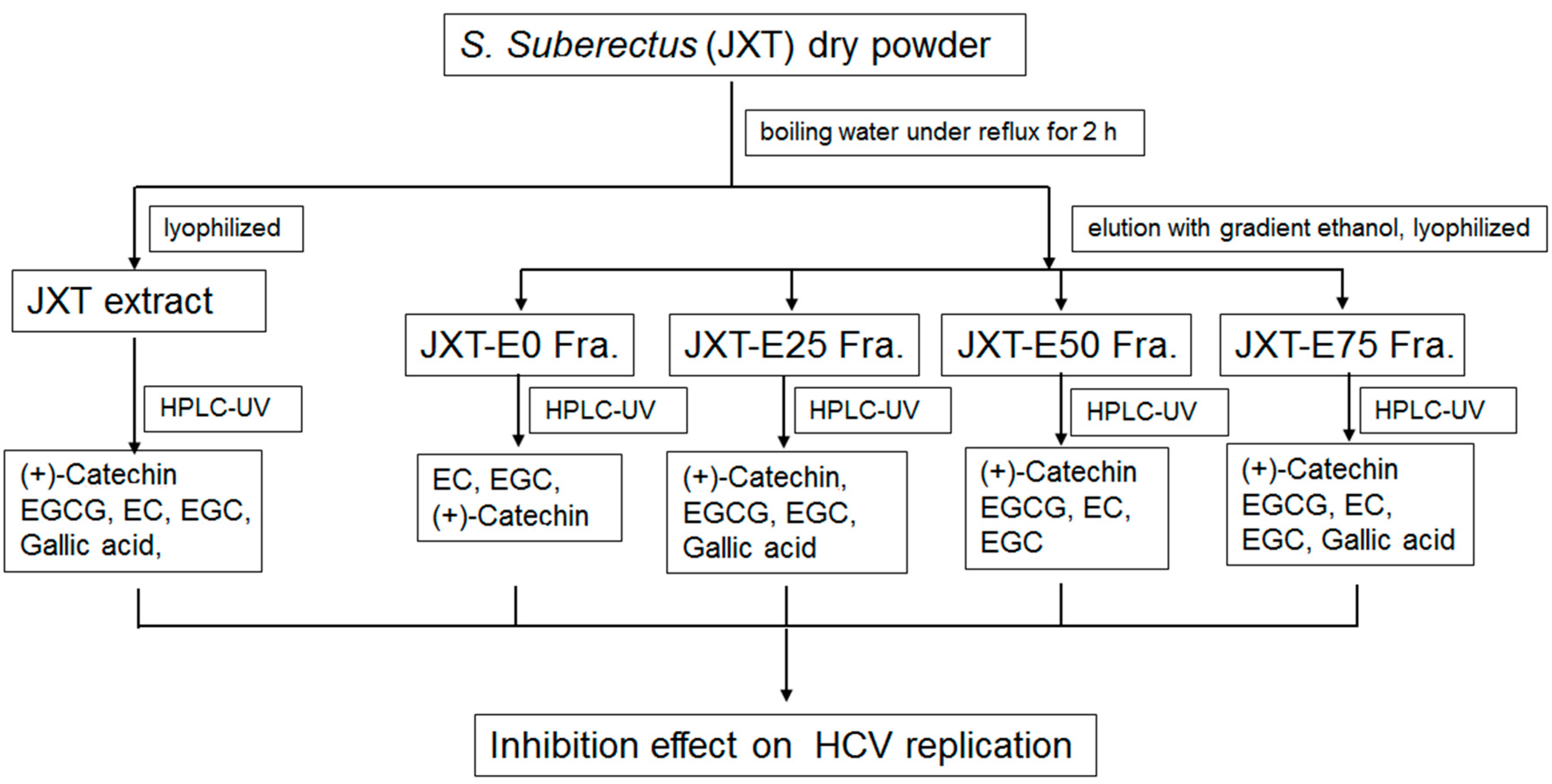
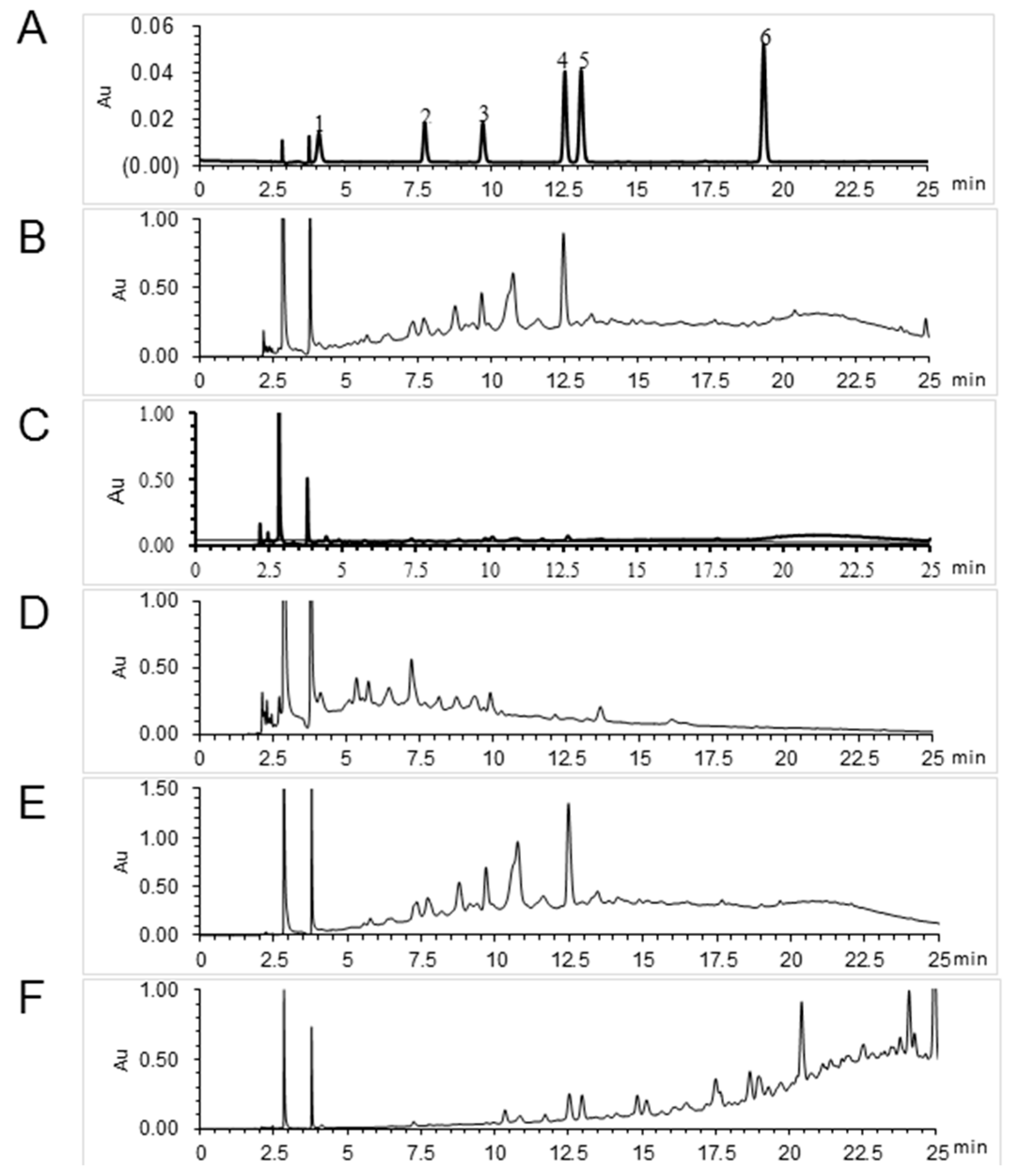
4.2. Extraction and Isolation of Herbs
4.3. Chromatographic Analysis
4.4. Determination of Tannins in JXT Extracts
4.5. Cell Culture Conditions
4.6. Anti-HCV Assay on Huh-luc/neo-ET Cell Line In Vitro
4.7. Cytotoxicity Analysis
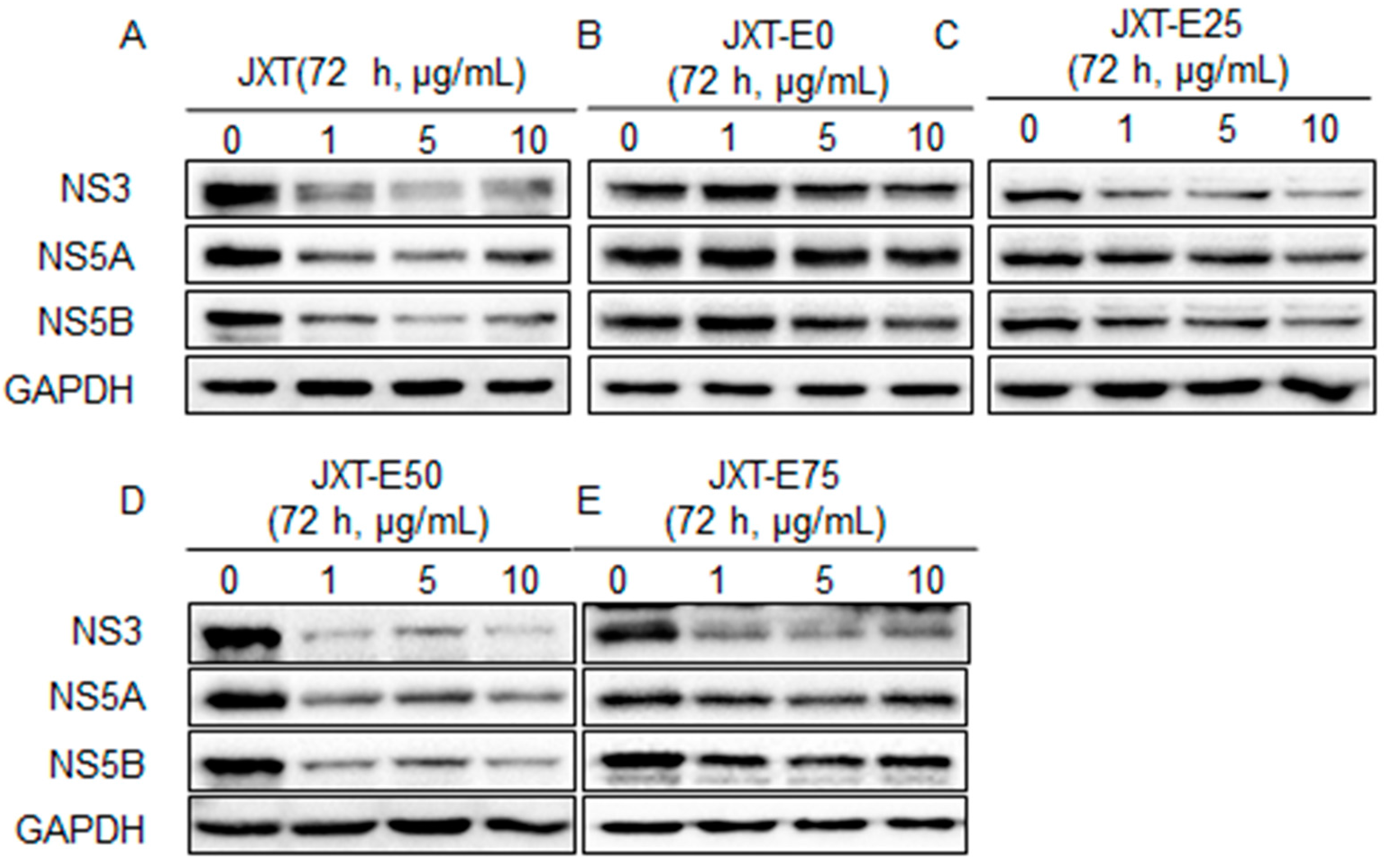
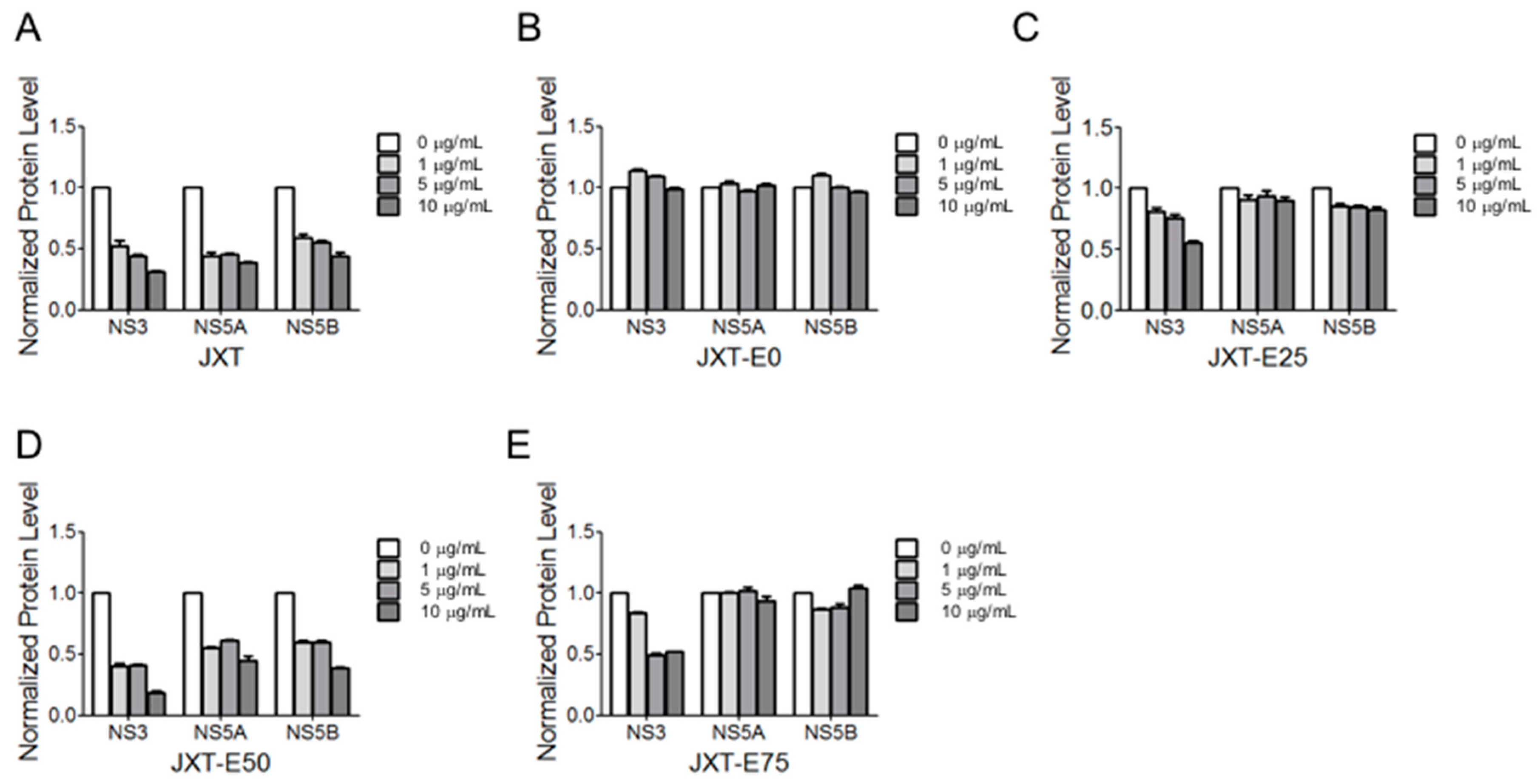
4.8. Western Blot Analysis
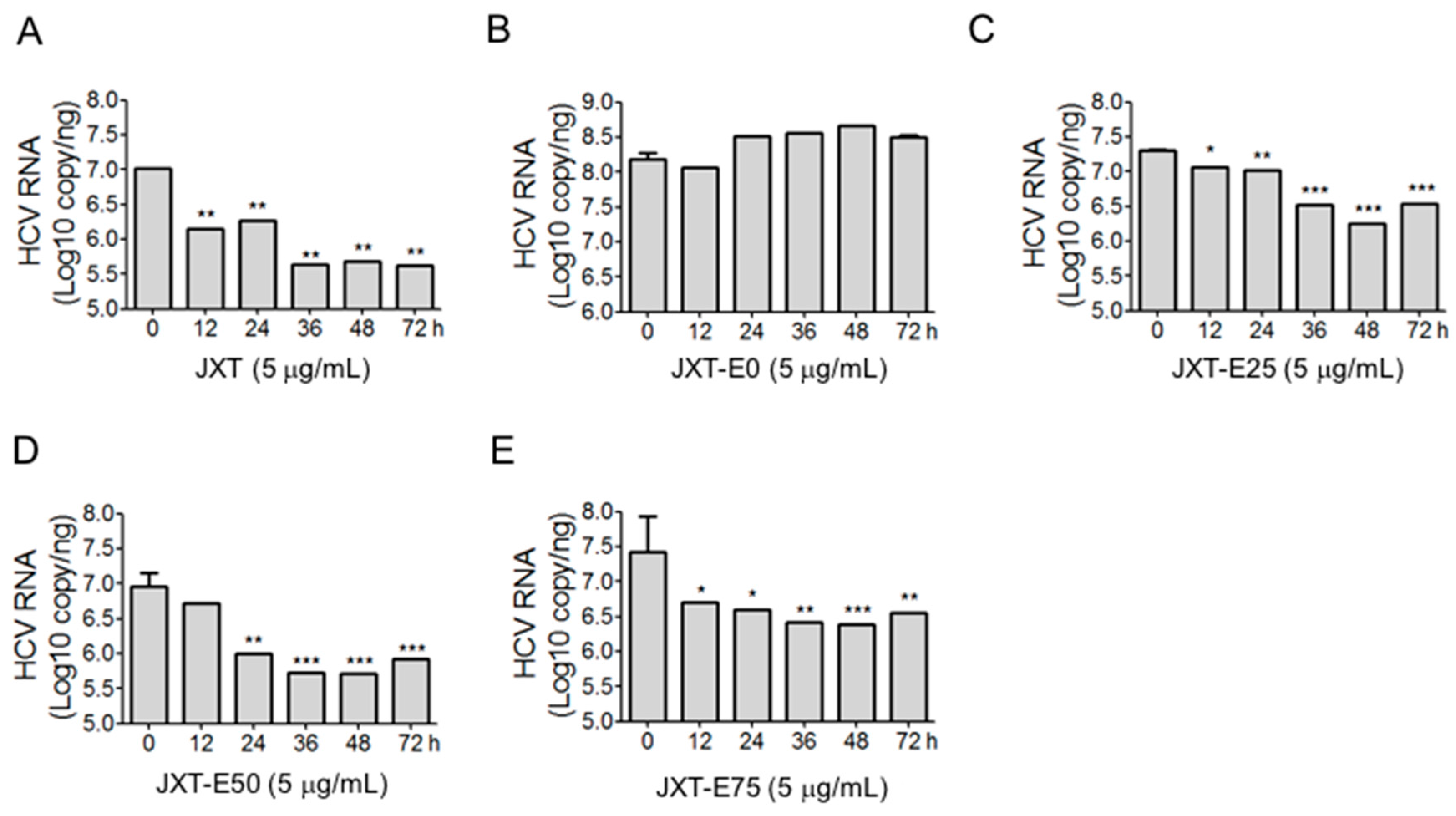
4.9. Extraction of Total RNA and Quantitative Real-Time-PCR (qRT-PCR) Analysis
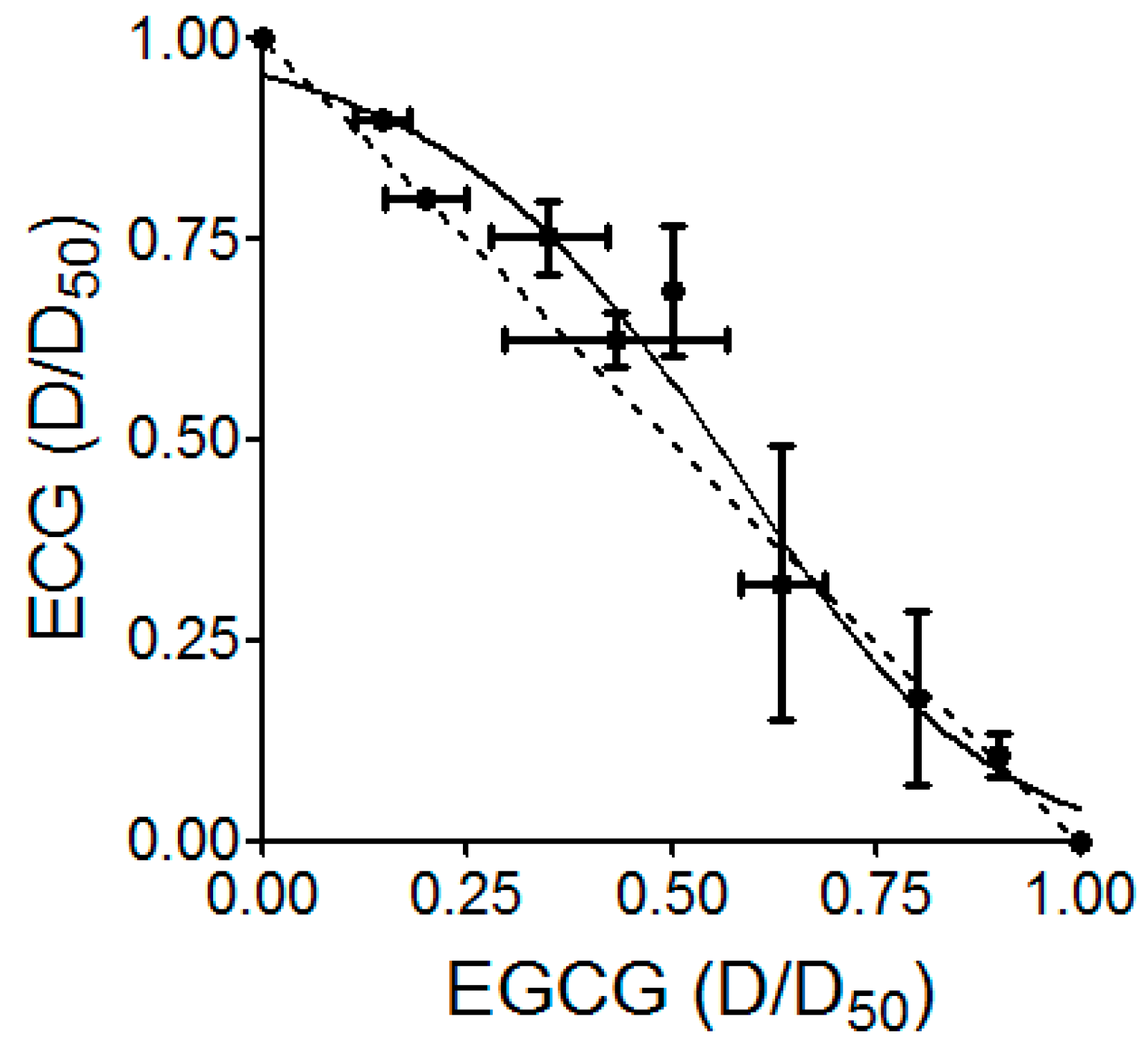
4.10. Combination Study
4.11. In Vitro Transcription and Translation Assay
4.12. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ARDS | Acute respiratory distress syndrome |
| CHX | Cycloheximide |
| C. pinnatifida Bge | Crataegus pinnatifida Bge |
| C. obtusifolia | Cassia obtusifolia |
| C. tinctorius L. | Carthamus tinctorius L. |
| DAAs | Direct anti-viral agents |
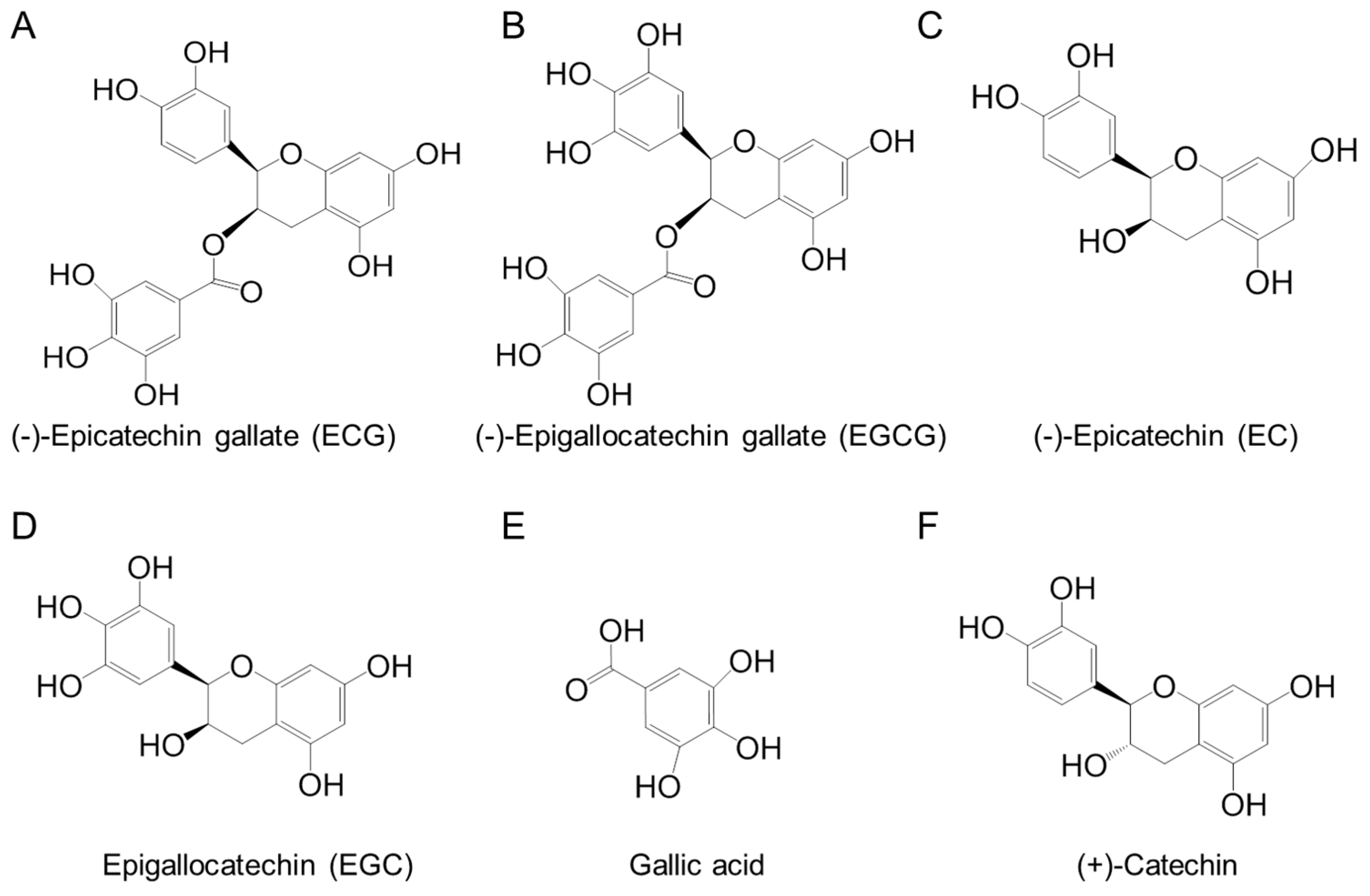
| EC | Epicatechin |
| ECG | Epicatechin gallate |
| EGC | Epigallocatechin |
| EGCG | Epigallocatechin gallate |
| FBS | Fetal bovine serum |
| F. chrysanthemi Indici | Flos chrysanthemi Indici |
| F. lycii | Fructus Lycii |
| F. suspense Thunb | Forsythia suspense (Thunb.) Vahl |
| HCV | Hepatitis C virus |
| JXT | Spatholobus suberectus |
| L. japonicus Houtt | Leonurus japonicas Houtt |
| L. japonica Thunb | Lonicera japonica Thunb |
| MNV-1 | Murine norovirus-1 |
| NEAA | Non-essential amino acid |
| NS proteins | Nonstructural proteins |
| P. cocos | Poria cocos (Schw.) Wolf |
| pegIFNα | Pegylated interferon α |
| qRT-PCR | Quantitative real-time-PCR |
| R. chingii Hu | Rubus chingii Hu |
| RT | Retention time |
| S. suberectus or R. astragali | Radix astragali |
| SVR | Sustained viral response |
References
- Gao, M.; Nettles, R.E.; Belema, M.; Snyder, L.B.; Nguyen, V.N.; Fridell, R.A.; Serrano-Wu, M.H.; Langley, D.R.; Sun, J.H.; O′Boyle, D.R.; et al. Chemical genetics strategy identifies an HCV NS5A inhibitor with a potent clinical effect. Nature 2010, 465, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Ishida, Y.; Takeshita, M.; Kataoka, H. Functional foods effective for hepatitis C: Identification of oligomeric proanthocyanidin and its action mechanism. World J. Hepatol. 2014, 6, 870–879. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, M.; Ishida, Y.; Akamatsu, E.; Ohmori, Y.; Sudoh, M.; Uto, H.; Tsubouchi, H.; Kataoka, H. Proanthocyanidin from blueberry leaves suppresses expression of subgenomic hepatitis C virus RNA. J. Biol. Chem. 2009, 284, 21165–21176. [Google Scholar] [CrossRef] [PubMed]
- Lok, A.S.; Gardiner, D.F.; Lawitz, E.; Martorell, C.; Everson, G.T.; Ghalib, R.; Reindollar, R.; Rustgi, V.; McPhee, F.; Wind-Rotolo, M.; et al. Preliminary study of two antiviral agents for hepatitis C genotype 1. N. Engl. J. Med. 2012, 366, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Kowdley, K.V.; Gordon, S.C.; Reddy, K.R.; Rossaro, L.; Bernstein, D.E.; Lawitz, E.; Shiffman, M.L.; Schiff, E.; Ghalib, R.; Ryan, M.; et al. Ledipasvir and sofosbuvir for 8 or 12 weeks for chronic HCV without cirrhosis. N. Engl. J. Med. 2014, 370, 1879–1888. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.C.; Martin, N.K.; Hickman, M.; Vickerman, P.; Page, E.E.; Everett, R.; Gazzard, B.G.; Nelson, M. Hepatitis C virus reinfection incidence and treatment outcome among HIV-positive MSM. AIDS 2013, 27, 2551–2557. [Google Scholar] [CrossRef] [PubMed]
- Tovo, C.V.; Becker, S.C.; Almeida, P.R.; Galperim, B.; Chaves, S. Progression of liver fibrosis in monoinfected patients by hepatitis C virus and coinfected by HCV and human immunodeficiency virus. Arq. Gastroenterol. 2013, 50, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Mohd Hanafiah, K.; Groeger, J.; Flaxman, A.D.; Wiersma, S.T. Global epidemiology of hepatitis C virus infection: New estimates of age-specific antibody to HCV seroprevalence. Hepatology 2013, 57, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Qin, W.; Cao, H.; Xu, R.; Tan, Y.; Lu, T.; Wang, H.; Tong, W.; Rong, X.; Li, G.; et al. HCV 6a prevalence in guangdong province had the origin from vietnam and recent dissemination to other regions of china: Phylogeographic analyses. PLoS ONE 2012, 7, e28006. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.H.; Yao, Z.H.; Liu, F.L.; Li, H.; Jiang, L.; Zhu, J.W.; Zheng, Y.T. High prevalence of HIV, HCV, HBV and co-infection and associated risk factors among injecting drug users in Yunnan Province, China. PLoS ONE 2012, 7, e42937. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Kang, D.; Tao, X.; Cox, C.; Qian, Y.; Wang, G.; Yang, C.; Zhu, X.; Zhang, N.; Bi, Z.; et al. Syndemics of syphilis, HCV infection, and methamphetamine use along the east coast of china. BMC Public Health 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Wang, Y.; Chow, E.P.; Liang, Y.; Wilson, D.P.; Zhang, L. Risk factors associated with HIV/HCV infection among entrants in methadone maintenance treatment clinics in China: A systematic review and meta-analysis. Drug Alcohol Depend. 2012, 126, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.E.; Waczuk, E.P.; Roversi, K.; da Silva, M.A.; Barros, L.M.; da Cunha, F.A.; de Menezes, I.R.; da Costa, J.G.; Boligon, A.A.; Ademiluyi, A.O.; et al. Polyphenolic composition and evaluation of antioxidant activity, osmotic fragility and cytotoxic effects of raphiodon echinus (Nees & Mart.) schauer. Molecules 2015, 21. [Google Scholar] [CrossRef]
- Ali, F.T.; Ali, M.A.; Elgizawy, M.M.; Elsawy, A.M. Secreted phosphoprotein 1 promoter genetic variants are associated with the response to pegylated interferon α plus ribavirin combination therapy in Egyptian patients with chronic hepatitis C virus infection. Gut. Liver 2015. [Google Scholar] [CrossRef]
- Backus, L.I.; Boothroyd, D.B.; Phillips, B.R.; Belperio, P.; Halloran, J.; Mole, L.A. A sustained virologic response reduces risk of all-cause mortality in patients with hepatitis C. Clin. Gastroenterol. Hepatol. 2011, 9, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Kau, A.; Vermehren, J.; Sarrazin, C. Treatment predictors of a sustained virologic response in hepatitis B and C. J. Hepatol. 2008, 49, 634–651. [Google Scholar] [CrossRef] [PubMed]
- Hnatyszyn, H.J. Chronic hepatitis C and genotyping: The clinical significance of determining HCV genotypes. Antivir. Ther. 2005, 10, 1–11. [Google Scholar] [PubMed]
- Berg, T.; Sarrazin, C.; Herrmann, E.; Hinrichsen, H.; Gerlach, T.; Zachoval, R.; Wiedenmann, B.; Hopf, U.; Zeuzem, S. Prediction of treatment outcome in patients with chronic hepatitis C: Significance of baseline parameters and viral dynamics during therapy. Hepatology 2003, 37, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, T.L.; Caetano, A.Z.; Belem, J.M.; Klemz, B.C.; Pinheiro, M.M. Interferon-α induced psoriatic arthritis and autoimmune hemolytic anemia during chronic hepatitis C treatment. Acta Reumatol. Port. 2014, 39, 327–330. [Google Scholar] [PubMed]
- Bartels, D.J.; Sullivan, J.C.; Zhang, E.Z.; Tigges, A.M.; Dorrian, J.L.; de Meyer, S.; Takemoto, D.; Dondero, E.; Kwong, A.D.; Picchio, G.; et al. Hepatitis C virus variants with decreased sensitivity to direct-acting antivirals (DAAS) were rarely observed in daa-naive patients prior to treatment. J. Virol. 2013, 87, 1544–1553. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, D.A.; Neely, R.D.; Bassendine, M.F. Hepatitis C virus and lipids in the era of direct acting antivirals (DAAS). Clin. Res. Hepatol. Gastroenterol. 2013, 37, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Forns, X.; Lawitz, E.; Zeuzem, S.; Gane, E.; Bronowicki, J.P.; Andreone, P.; Horban, A.; Brown, A.; Peeters, M.; Lenz, O.; et al. Simeprevir with peginterferon and ribavirin leads to high rates of SVR in patients with HCV genotype 1 who relapsed after previous therapy: A phase 3 trial. Gastroenterology 2014, 146, 1669–1679. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.S. Sofosbuvir for previously untreated chronic hepatitis C infection. N. Engl. J. Med. 2013, 369. [Google Scholar] [CrossRef]
- Pawlotsky, J.M. Treatment failure and resistance with direct-acting antiviral drugs against hepatitis C virus. Hepatology 2011, 53, 1742–1751. [Google Scholar] [CrossRef] [PubMed]
- Fried, M.W.; Navarro, V.J.; Afdhal, N.; Belle, S.H.; Wahed, A.S.; Hawke, R.L.; Doo, E.; Meyers, C.M.; Reddy, K.R.; Silymarin, N.; et al. Effect of silymarin (milk thistle) on liver disease in patients with chronic hepatitis C unsuccessfully treated with interferon therapy: A randomized controlled trial. JAMA 2012, 308, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Paddon, C.J.; Westfall, P.J.; Pitera, D.J.; Benjamin, K.; Fisher, K.; McPhee, D.; Leavell, M.D.; Tai, A.; Main, A.; Eng, D.; et al. High-level semi-synthetic production of the potent antimalarial artemisinin. Nature 2013, 496, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Hara, Y. Tea catechins and their applications as supplements and pharmaceutics. Pharmacol. Res. 2011, 64, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Perez, L.B.; Still, P.C.; Naman, C.B.; Ren, Y.; Pan, L.; Chai, H.B.; Carcache de Blanco, E.J.; Ninh, T.N.; van Thanh, B.; Swanson, S.M.; et al. Investigation of vietnamese plants for potential anticancer agents. Phytochem. Rev. 2014, 13, 727–739. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Bae, S.Y.; Oh, M.; Kim, K.H.; Chung, M.S. Antiviral effects of mulberry (Morus alba) juice and its fractions on foodborne viral surrogates. Foodborne Pathog. Dis. 2014, 11, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, Y.; Zheng, Z.; Zhao, S.; Zhao, J.; Lin, Q.; Li, C.; Zhu, Q.; Zhong, N. Antiviral activity of Isatis indigotica root-derived clemastanin b against human and avian influenza A and B viruses in vitro. Int. J. Mol. Med. 2013, 31, 867–873. [Google Scholar] [PubMed]
- Yang, Z.; Wang, Y.; Zhong, S.; Zhao, S.; Zeng, X.; Mo, Z.; Qin, S.; Guan, W.; Li, C.; Zhong, N. In vitro inhibition of influenza virus infection by a crude extract from Isatis indigotica root resulting in the prevention of viral attachment. Mol. Med. Rep. 2012, 5, 793–799. [Google Scholar] [PubMed]
- Li, J.; Zhou, B.; Li, C.; Chen, Q.; Wang, Y.; Li, Z.; Chen, T.; Yang, C.; Jiang, Z.; Zhong, N.; et al. Lariciresinol-4-O-β-d-glucopyranoside from the root of Isatis indigotica inhibits influenza a virus-induced pro-inflammatory response. J. Ethnopharmacol. 2015, 174, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.P.; Li, G.; Li, X.; Hu, Q.P.; Liu, J.X.; Zhang, F.X.; Su, Z.R.; Lai, X.P. The roots of Ilex asprella extract lessens acute respiratory distress syndrome in mice induced by influenza virus. J. Ethnopharmacol. 2014, 155, 1575–1582. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, L.; Duan, L.; Dong, X.; Zhou, P.; Liu, E.H.; Li, P. Simultaneous determination of 16 phenolic constituents in spatholobi caulis by high performance liquid chromatography/electrospray ionization triple quadrupole mass spectrometry. J. Pharm. Biomed. Anal. 2015, 102, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Stickel, F.; Hellerbrand, C. Herbs to treat liver diseases: More than placebo? Clin. Liver Dis. 2015, 6, 136–138. [Google Scholar] [CrossRef]
- Wang, H.; Wang, B. Long-term follow-up result of compound dan shen granule (861 Chong Fu Ji) in treating hepatofibrosis. Chin. J. Integr. Tradit. West. Med. 1995, 5, 4–5. [Google Scholar]
- Wang, Z.; Wang, D.; Han, S.; Wang, N.; Mo, F.; Loo, T.Y.; Shen, J.; Huang, H.; Chen, J. Bioactivity-guided identification and cell signaling technology to delineate the lactate dehydrogenase a inhibition effects of Spatholobus suberectus on breast cancer. PLoS ONE 2013, 8, e56631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.L.; Wan, J.Y.; Li, P.; Qi, L.W. Ultrasonic/microwave assisted extraction and diagnostic ion filtering strategy by liquid chromatography-quadrupole time-of-flight mass spectrometry for rapid characterization of flavonoids in Spatholobus suberectus. J. Chromatogr. A 2011, 1218, 5774–5786. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Huang, K.; Liu, W.; Fu, F.; Xu, J. Combined autodock and comparative molecular field analysis study on predicting 5-lipoxygenase inhibitory activity of flavonoids isolated from Spatholobus suberectus dunn. Z. Naturforsch. C 2015, 70, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.; Guo, J.P.; Jin, M.; Chen, Z.Q.; Wang, X.W.; Li, J.W. Antiviral effects of aqueous extract from Spatholobus suberectus dunn. Against coxsackievirus B3 in mice. Chin. J. Integr. Med. 2011, 17, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Park, K.S.; Choi, Y.J.; Park, J.S. Enterovirus infection in Korean children and anti-enteroviral potential candidate agents. Korean J. Pediatr. 2012, 55, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.P.; Pang, J.; Wang, X.W.; Shen, Z.Q.; Jin, M.; Li, J.W. In vitro screening of traditionally used medicinal plants in china against enteroviruses. World J. Gastroenterol. 2006, 12, 4078–4081. [Google Scholar] [CrossRef] [PubMed]
- Sarwar, S.; Khan, A.A.; Tarique, S. Response guided interferon therapy for genotype 3 of chronic hepatitis C: Compliance and outcome. Pak. J. Med. Sci. 2015, 31, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Waldenstrom, J.; Farkkila, M.; Rembeck, K.; Norkrans, G.; Langeland, N.; Morch, K.; Pedersen, C.; Rauning Buhl, M.; Nieminen, U.; Nuutinen, H.; et al. Short interferon and ribavirin treatment for HCV genotype 2 or 3 infection: Nordynamic trial and real-life experience. Scand. J. Gastroenterol. 2015, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Galani, B.R.; Sahuc, M.E.; Njayou, F.N.; Deloison, G.; Mkounga, P.; Feudjou, W.F.; Brodin, P.; Rouille, Y.; Nkengfack, A.E.; Moundipa, P.F.; et al. Plant extracts from cameroonian medicinal plants strongly inhibit hepatitis C virus infection in vitro. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Wahyuni, T.S.; Tumewu, L.; Permanasari, A.A.; Apriani, E.; Adianti, M.; Rahman, A.; Widyawaruyanti, A.; Lusida, M.I.; Fuad, A.; Soetjipto; et al. Antiviral activities of indonesian medicinal plants in the east Java region against hepatitis C virus. Virol. J. 2013, 10. [Google Scholar] [CrossRef] [PubMed]
- Jardim, A.C.; Igloi, Z.; Shimizu, J.F.; Santos, V.A.; Felippe, L.G.; Mazzeu, B.F.; Amako, Y.; Furlan, M.; Harris, M.; Rahal, P. Natural compounds isolated from Brazilian plants are potent inhibitors of hepatitis C virus replication in vitro. Antivir. Res. 2015, 115, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Afdhal, N.; Reddy, K.R.; Nelson, D.R.; Lawitz, E.; Gordon, S.C.; Schiff, E.; Nahass, R.; Ghalib, R.; Gitlin, N.; Herring, R.; et al. Ledipasvir and sofosbuvir for previously treated HCV genotype 1 infection. N. Engl. J. Med. 2014, 370, 1483–1493. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.P.; Ramirez, S.; Jensen, S.B.; Purcell, R.H.; Gottwein, J.M.; Bukh, J. Highly efficient full-length hepatitis C virus genotype 1 (strain TN) infectious culture system. Proc. Natl. Acad. Sci. USA 2012, 109, 19757–19762. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.Q.; Lin, X.P.; Wang, Z.; Zhou, X.F.; Qin, X.C.; Kaliyaperumal, K.; Zhang, T.Y.; Tu, Z.C.; Liu, Y. Asteltoxins with antiviral activities from the marine sponge-derived fungus Aspergillus sp. SCSIO XWS02F40. Molecules 2015, 21. [Google Scholar] [CrossRef] [PubMed]
- Reddy, B.U.; Mullick, R.; Kumar, A.; Sudha, G.; Srinivasan, N.; Das, S. Small molecule inhibitors of HCV replication from pomegranate. Sci. Rep. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.T.; Wang, F.; Wang, G.X.; Han, J.; Wang, Y.; Wang, Y.H. In vivo anthelmintic activity of crude extracts of radix angelicae pubescentis, fructus bruceae, caulis spatholobi, semen aesculi, and semen pharbitidis against Dactylogyrus intermedius (monogenea) in goldfish (Carassius auratus). Parasitol. Res. 2010, 106, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, J.; Chen, J.; Zhu, D.; Zhou, H.; Wang, X. A study on anticancer activity of Caulis spatholobi extract on human osteosarcoma SAOS-2 cells. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.G.; Ruan, J.S.; Zhang, L.; Fan, F.T.; Zhang, F.; Wang, A.Y.; Zheng, S.Z.; Zeng, L.; Li, W.L.; Lu, Y. Effect of aqueous extracts of several kinds of herbs on human platelet aggregation and expression of p-selectin in vitro. Chin. J. Integr. Med. 2015, 21, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; He, H.; Wu, B.; Yao, S. Cytotoxic effect on cancer cells and structural identification of phenols from spatholobi caulis by HPLC-ESI-MS(n). Nat. Prod. Commun. 2009, 4, 809–812. [Google Scholar] [PubMed]
- Xia, J.F.; Gao, J.J.; Inagaki, Y.; Kokudo, N.; Nakata, M.; Tang, W. Flavonoids as potential anti-hepatocellular carcinoma agents: Recent approaches using HepG2 cell line. Drug Discov. Ther. 2013, 7. [Google Scholar] [CrossRef]
- Ahmed-Belkacem, A.; Guichou, J.F.; Brillet, R.; Ahnou, N.; Hernandez, E.; Pallier, C.; Pawlotsky, J.M. Inhibition of RNA binding to hepatitis C virus RNA-dependent RNA polymerase: A new mechanism for antiviral intervention. Nucleic Acids Res. 2014, 42, 9399–9409. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.C.; Chang, S.P.; Lin, L.C.; Li, C.L.; Richardson, C.D.; Lin, C.C.; Lin, L.T. Limonium sinense and gallic acid suppress hepatitis C virus infection by blocking early viral entry. Antivir. Res. 2015, 118, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Qiu, H.; Gong, J.; Liu, Q.; Xiao, H.; Chen, X.W.; Sun, B.L.; Yang, R.G. (−)-epigallocatechin-3-gallate inhibits the replication cycle of hepatitis C virus. Arch. Virol. 2012, 157, 1301–1312. [Google Scholar] [CrossRef] [PubMed]
- Fukazawa, H.; Suzuki, T.; Wakita, T.; Murakami, Y. A cell-based, microplate colorimetric screen identifies 7,8-benzoflavone and green tea gallate catechins as inhibitors of the hepatitis C virus. Biol. Pharm. Bull. 2012, 35, 1320–1327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Schuppan, D. Traditional Chinese Medicine (TCM) for fibrotic liver disease: Hope and hype. J. Hepatol. 2014, 61, 166–168. [Google Scholar] [CrossRef] [PubMed]
- Lam, W.; Bussom, S.; Guan, F.; Jiang, Z.; Zhang, W.; Gullen, E.A.; Liu, S.H.; Cheng, Y.C. The four-herb Chinese medicine PHY906 reduces chemotherapy-induced gastrointestinal toxicity. Sci. Transl. Med. 2010, 2. [Google Scholar] [CrossRef] [PubMed]
- Galani Tietcheu, B.R.; Sass, G.; Njayou, N.F.; Mkounga, P.; Tiegs, G.; Moundipa, P.F. Anti-hepatitis C virus activity of crude extract and fractions of Entada africana in genotype 1b replicon systems. Am. J. Chin. Med. 2014, 42, 853–868. [Google Scholar] [CrossRef] [PubMed]
- State Pharmacopoeia Committee. Pharmacopoeia of the People’s Republic of China, 2010 ed.; China Medical Science and Technology Press: Beijing, China, 2010. [Google Scholar]
- Lohmann, V.; Korner, F.; Koch, J.; Herian, U.; Theilmann, L.; Bartenschlager, R. Replication of subgenomic hepatitis C virus RNAs in a hepatoma cell line. Science 1999, 285, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Tsou, L.K.; Cai, J.; Aya, T.; Dutschman, G.E.; Gullen, E.A.; Grill, S.P.; Chen, A.P.; Lindenbach, B.D.; Hamilton, A.D.; et al. A novel class of meso-tetrakis-porphyrin derivatives exhibits potent activities against hepatitis C virus genotype 1B replicons in vitro. Antimicrob. Agents Chemother. 2010, 54, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C.; Motzer, R.J.; Tong, Y.; Bosl, G.J. Computerized quantitation of synergism and antagonism of taxol, topotecan, and cisplatin against human teratocarcinoma cell growth: A rational approach to clinical protocol design. J. Natl. Cancer Inst. 1994, 86, 1517–1524. [Google Scholar] [CrossRef] [PubMed]
- Greco, W.R.; Park, H.S.; Rustum, Y.M. Application of a new approach for the quantitation of drug synergism to the combination of cis-diamminedichloroplatinum and 1-β-d-arabinofuranosylcytosine. Cancer Res. 1990, 50, 5318–5327. [Google Scholar] [PubMed]
- Sample Availability: All samples of the compounds in this research including ECG, EGCG, EC, EGC, Gallic acid, (+)-Catechin as well as fractions from S. suberectus are available from the authors.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Latin Name | Organ/Channel | Huh-Luc/Neo-ET | |
|---|---|---|---|---|
| Anti-HCV Activity (100 μg/mL) | Cytotoxicity (100 μg/mL) or IC50 | |||
| Drug tonify and nourish the yin | Fructus lycii (fruit) | Liver, Kidney, heart | N.D. a | N.D. |
| Tonic drug | Radix astragali (root) | Spleen, stomach, middle burner, Lung | N.D. | N.D. |
| Drug tonify yang | Rubus chingii Hu (fruit) | Liver, Kidney | N.D. | N.D. |
| Drug dispel hot heat | Flos chrysanthemi Indici (flower) | Lung, Liver | N.D. | N.D. |
| Drug dispel hot heat | Cassia obtusifolia (seed) | Liver and Large Intestine | N.D. | JMZ-E25: 102.6 ± 2.5 μg/mL b; JMZ-E75: 121.5 ± 7.9 μg/mL |
| Drug dispel hot heat | Lonicera japonica (L. japonica) Thunb. (flower) | Lung, heart, stomach, large intestine, liver | N.D. | N.D. |
| Drug dispel hot heat | Forsythia suspense (Thunb.) Vahl (fruit) | Heart, liver, gallbladder, small intestine | N.D. | N.D. |
| Drug promote blood circulation | Poria cocos (Schw.) Wolf) (sclerotia) | Heart, Kidney, Spleen, Lung, stomach | N.D. | N.D. |
| Drug promote blood circulation | Carthamus tinctorius L. (flower) | Heart, Liver, Kidney; | N.D. | N.D. |
| Drug promote blood circulation | Spatholobus suberectus (stem) | Liver and Kidney | JXT JXT-E0 JXT-E25 JXT-E50 JXT-E75 | JXT-E0: 89.1 ± 9.4 μg/mL |
| Drug promote blood circulation | Leonurus japonicas Houtt. (leaf) | Liver, heart kidney and Pericardium | N.D. | N.D. |
| Drug regulate and tonify the blood, alleviate food stagnation | Crataegus pinnatifida Bge. (fruit) | Spleen, stomach and liver | N.D. | N.D. |
| Fractions | Anti-HCV Activity (EC50) a | Cytotoxicity (IC50) b |
|---|---|---|
| JXT | 1.68 ± 0.39 μg/mL | >100 μg/mL |
| JXT-E0 | 74.1 ± 1.0 μg/mL | 89.1 ± 9.4 μg/mL |
| JXT-E25 | 0.45 ± 0.08 μg/mL | >100 μg/mL |
| JXT-E50 | 1.03 ± 0.06 μg/mL | >100 μg/mL |
| JXT-E75 | 1.13 ± 0.15 μg/mL | >100 μg/mL |
| Telaprevir | 185.0 ± 35.0 nM (125.77 ± 23.76 ng/mL) | |
| Sofosbuvir | 26.3 ± 5.5 nM (13.94 ± 2.92 ng/mL) | |
| Daclatasvir | 0.008 ± 0.001 nM (5.911 ± 0.522 pg/mL) | |
| Simeprevir | 1.27 ± 0.25 nM (42.50 ± 3.90 ng/mL) | |
| IFNα-2a | 0.760 ± 0.115 fg/mL |
| Fraction | ECG (%) | EGCG (%) | EC (%) | EGC (%) | Gallic Acid (%) | (+)-Catechin (%) | Tannin (Mean ± SD, %) (n = 3) |
|---|---|---|---|---|---|---|---|
| JXT | N.D. * | 0.01 | 1.08 | 0.59 | 0.01 | 0.34 | 20.2 ± 1.1 |
| JXT-E0 | N.D. * | N.D. * | 0.09 | 0.03 | N.D. * | 0.01 | 0.2 ± 1.9 |
| JXT-E25 | N.D. * | 0.01 | N.D. * | 0.12 | 0.04 | 0.04 | 0.3 ± 1.3 |
| JXT-E50 | N.D. * | 0.01 | 2.10 | 1.17 | N.D. * | 0.69 | 27.0 ± 1.6 |
| JXT-E75 | 0.04 | 0.10 | 0.39 | 0.03 | 0.01 | 0.01 | 20.2 ± 1.2 |
| Compound | CID | Anti-HCV Activity (EC50, 72 h) | Cytotoxicity (IC50, 72 h) |
|---|---|---|---|
| ECG | 107905 | 69.833 ± 5.620 μM | >100 μM |
| EGCG | 65064 | 68.667 ± 4.509 μM | >100 μM |
| EC | 72276 | >100 μM | >100 μM |
| EGC | 72277 | >100 μM | >100 μM |
| Gallic acid | 370 | >100 μM | >100 μM |
| (+)-Catechin | 9064 | >100 μM | >100 μM |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.-R.; Wang, A.-Q.; Lin, L.-G.; Qiu, H.-C.; Wang, Y.-T.; Wang, Y. In Vitro Study on Anti-Hepatitis C Virus Activity of Spatholobus suberectus Dunn. Molecules 2016, 21, 1367. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21101367
Chen S-R, Wang A-Q, Lin L-G, Qiu H-C, Wang Y-T, Wang Y. In Vitro Study on Anti-Hepatitis C Virus Activity of Spatholobus suberectus Dunn. Molecules. 2016; 21(10):1367. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21101367
Chicago/Turabian StyleChen, Shao-Ru, An-Qi Wang, Li-Gen Lin, Hong-Cong Qiu, Yi-Tao Wang, and Ying Wang. 2016. "In Vitro Study on Anti-Hepatitis C Virus Activity of Spatholobus suberectus Dunn" Molecules 21, no. 10: 1367. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21101367