
Seasonal Dynamics of Metabolites in Needles of Taxus wallichiana var. mairei


Abstract
:1. Introduction
2. Results and Discussion
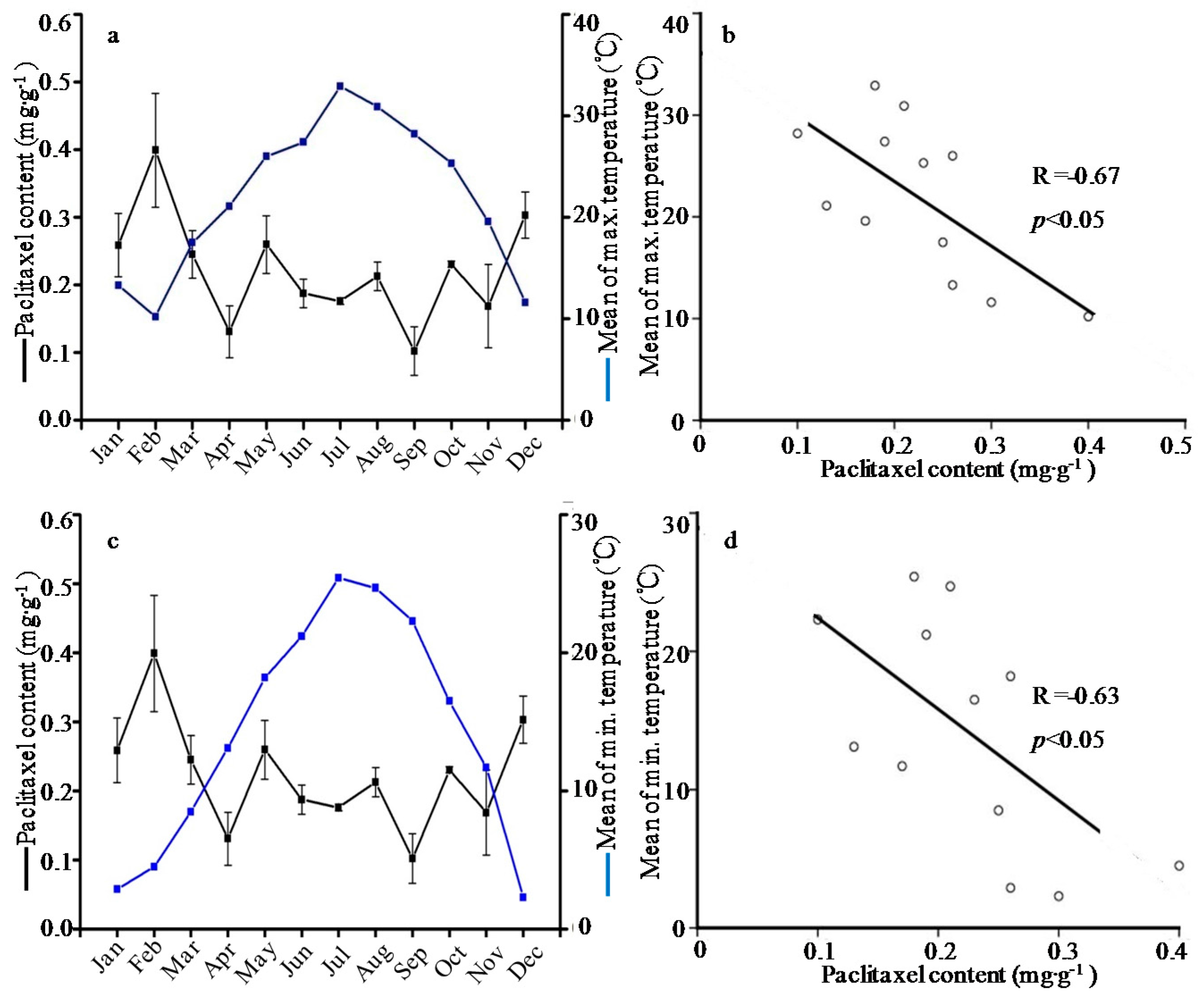
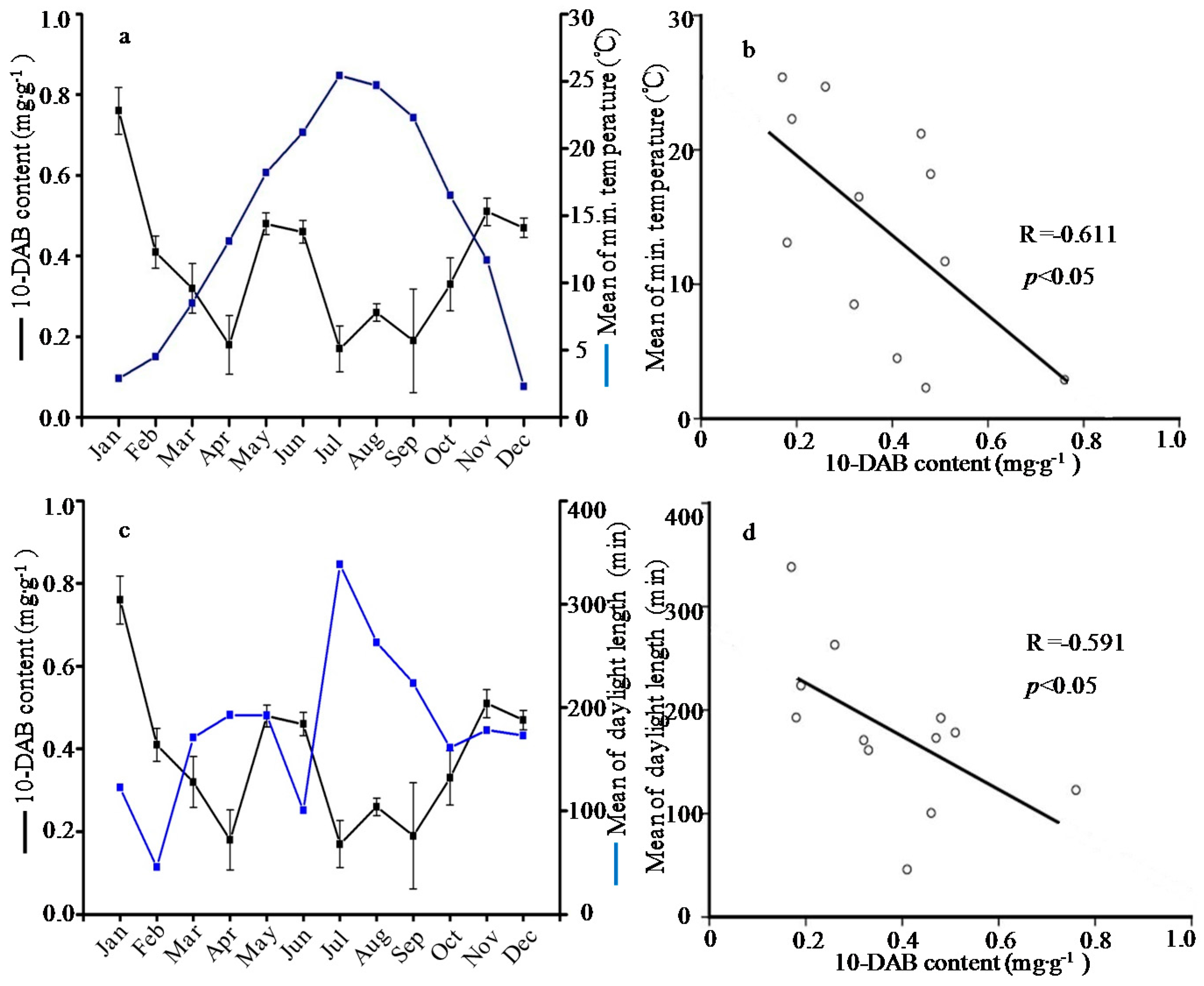
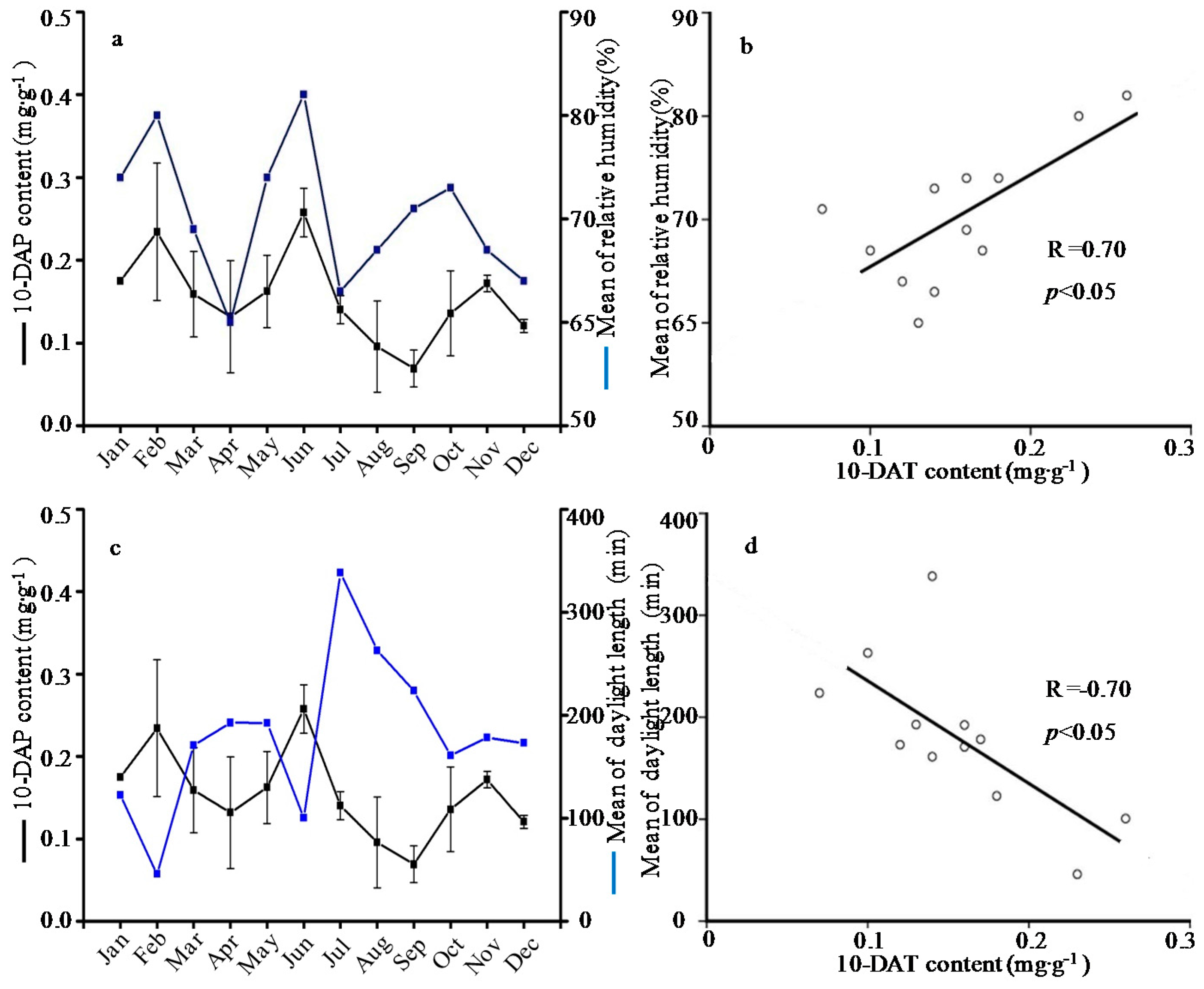
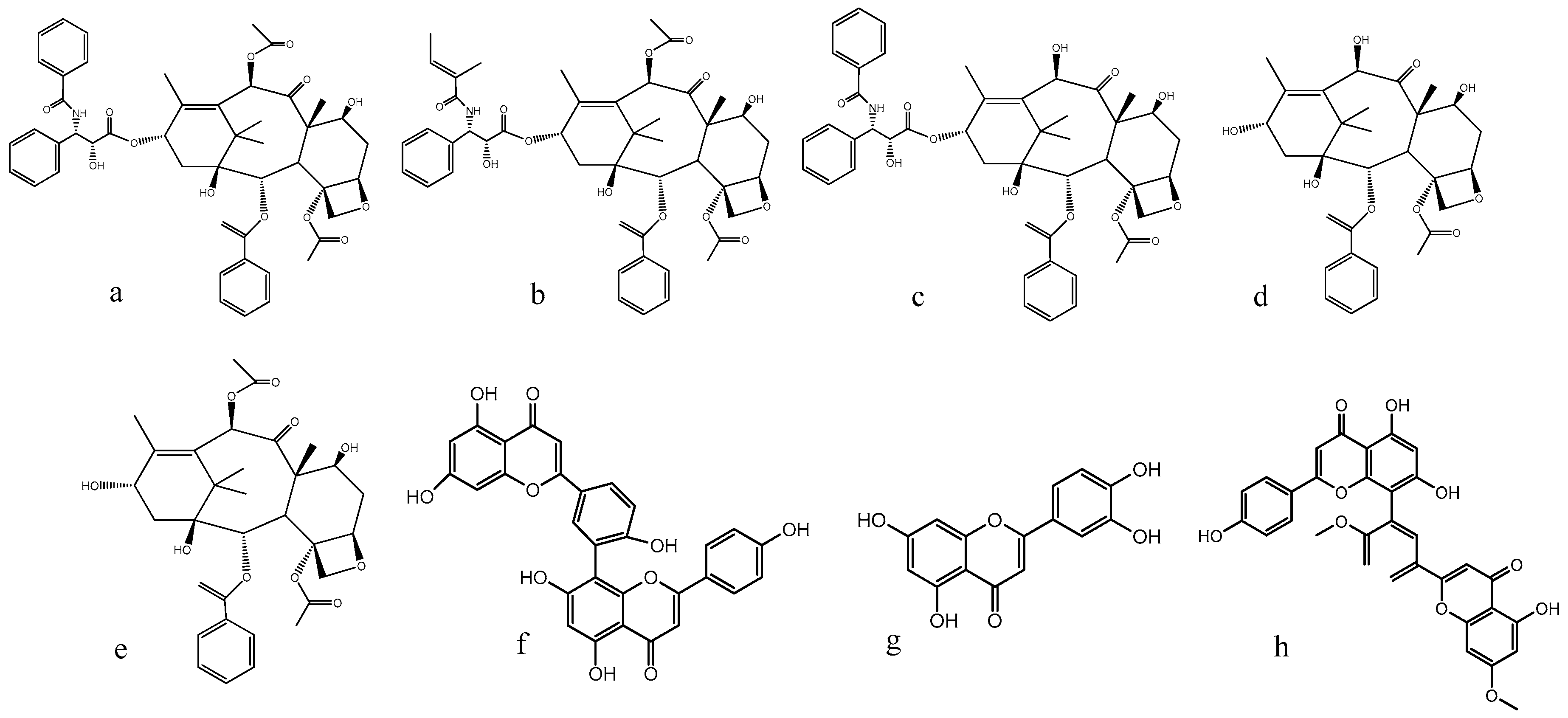
2.1. Seasonal Variation of the Taxoid Content
2.2. Seasonal Variations of the Flavonoid Content
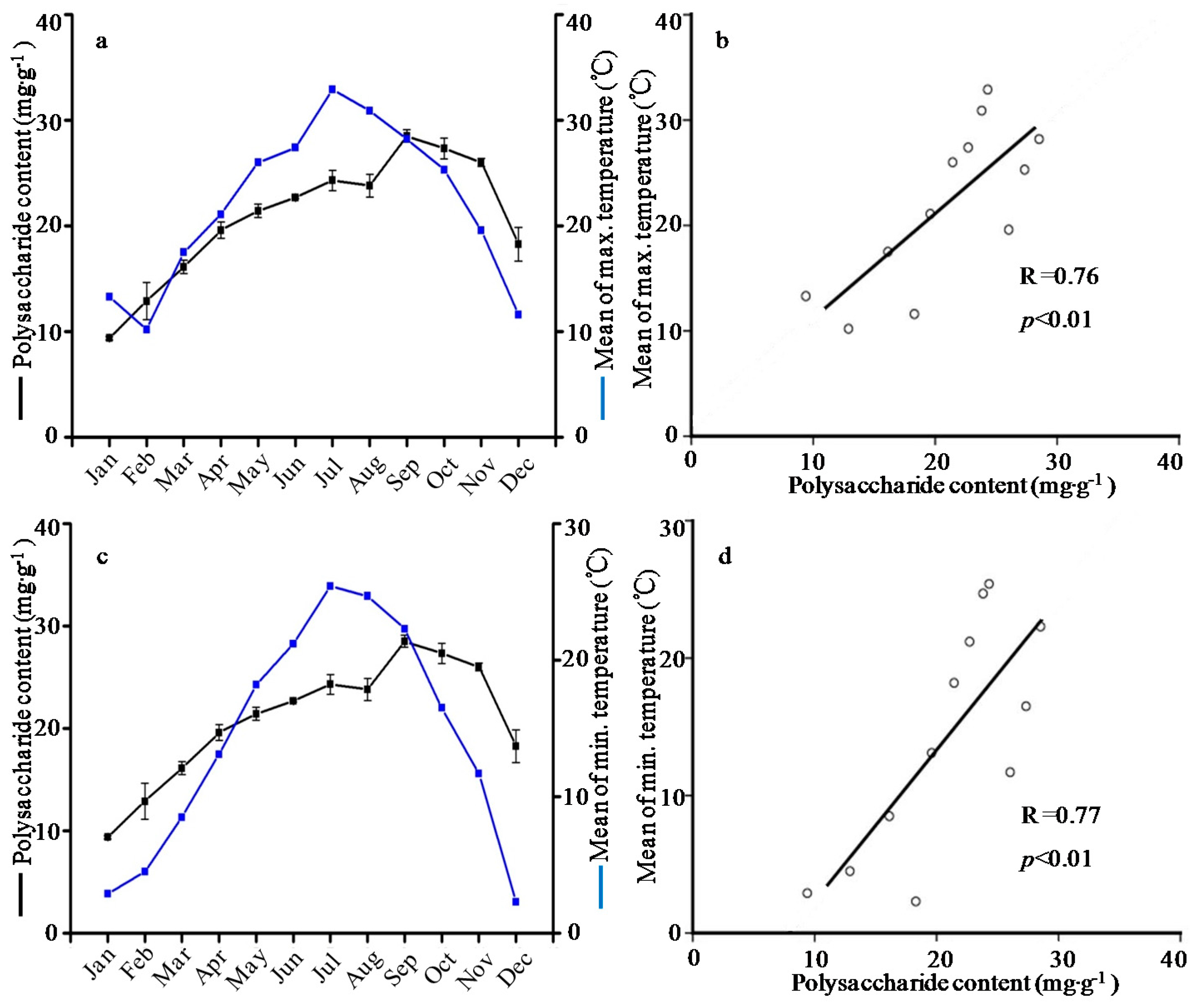
2.3. Seasonal Variation of Polysaccharide Content
3. Materials and Methods
3.1. Plant Materials and Chemicals
3.2. Meteorological Parameter Collection
3.3. Preparation of Samples
3.3.1. Preparation of Taxoid Sample
3.3.2. Preparation of Flavonoid Sample
3.3.3. Preparation of Polysaccharide Sample
3.4. Determination of Sample Component Contents
3.4.1. Determination of the Taxoid Content
3.4.2. Determination of the Flavonoid Content
3.4.3. Determination of the Polysaccharides
3.5. Statistical Analyses
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wu, Z.Y.; Raven, P.H.; Hong, D.Y. Flora of China; Science Press: Beijing, China, 2004; Volume 8. [Google Scholar]
- Shi, Q.W.; Kiyota, H. New natural taxane diterpenoids from Taxus species since 1999. Chem. Biodivers. 2005, 2, 1597–1623. [Google Scholar] [CrossRef] [PubMed]
- Qu, C.; Chen, Z. Antitumor effect of water decoctions of Taxus cuspidate on pancreatic cancer. Evid. Based Complement. Altern. Med. 2014, 2014, 291675. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.Q.; Fu, Y.Y.; Li, B.H.; Zhang, M.L.; Yang, X.L.; Xin, C.W.; Huang, P. PSY-1, a Taxus chinensis var. mairei extract, inhibits cancer cell metastasis by interfering with MMPs. Nat. Prod. Commun. 2014, 9, 241–245. [Google Scholar] [PubMed]
- Horwitz, S.B. Mechanism of action of taxol. Trends Pharmacol. Sci. 1992, 13, 134–136. [Google Scholar] [CrossRef]
- Markman, M.; Mekhail, T.M. Paclitaxel in cancer therapy. Expert Opin. Pharmacother. 2002, 3, 755–766. [Google Scholar] [CrossRef] [PubMed]
- El-Khattouti, A.; Selimovic, D.; Haïkel, Y.; Megahed, M.; Gomez, C.R.; Hassan, M. Identification and analysis of CD133+ melanoma stem-like cells conferring resistance to taxol: An insight into the mechanisms of their resistance and response. Cancer Lett. 2014, 343, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.; Wang, Y.; Yu, X.; Guo, B.; Tang, K. A new endophytic taxane production fungus from Taxus chinensis. Appl. Biochem. Microbiol. 2009, 45, 81–86. [Google Scholar] [CrossRef]
- Kingston, D.G. The chemistry of taxol. Pharmacol. Ther. 1991, 52, 1–34. [Google Scholar] [CrossRef]
- Appendino, G. The phytochemistry of the yew tree. Nat. Prod. Rep. 1995, 12, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.W.; Oritani, T.; Sugiyama, T.; Murakami, R.; Wei, H.Q. Six new taxane diterpenoids from the seeds of Taxus chinensis var. mairei and Taxus yunnanensis. J. Nat. Prod. 1999, 62, 1114–1118. [Google Scholar] [CrossRef] [PubMed]
- Krauze-Baranowska, M. Flavonoids from the genus Taxus. Z. Naturforsch. C 2004, 59, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Li, Z.; Li, C.; Yang, L.; Yao, L.; Fu, Y.; He, X.; Shi, K.; Lu, Z. Optimized extraction of polysaccharides from Taxus chinensis var. mairei fruits and its antitumor activity. Int. J. Biol. Macromol. 2015, 75, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Zhang, F.; Yu, Z.; Lin, J.; Yang, L. Chemical characterization and in vitro antitumor activity of a single-component polysaccharide from Taxus chinensis var. mairei. Carbohydr. Polym. 2015, 133, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Yin, Y.; Zhang, D.; Yang, W.; Yu, R. Structural characterization and in vitro antitumor activity of a novel polysaccharide from Taxus yunnanensis. Carbohydr. Polym. 2013, 96, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Asker, M.M.S.; Shawky, B.T. Structural characterization and antioxidant activity of an extracellular polysaccharide isolated from Brevibacterium otitidis BTS 44. Food Chem. 2010, 123, 315–320. [Google Scholar] [CrossRef]
- Wang, Z.M.; Peng, X.; Lee, K.L.D.; Tang, J.C.O.; Cheung, P.C.K.; Wu, J.Y. Structural characterisation and immunomodulatory property of an acidic polysaccharide from mycelial culture of Cordyceps sinensis fungus Cs-HK1. Food Chem. 2011, 125, 637–643. [Google Scholar] [CrossRef]
- Fan, L.; Ding, S.; Ai, L.; Deng, K. Antitumor and immunomodulatory activity of water-soluble polysaccharide from Inonotus obliquus. Carbohydr. Polym. 2012, 90, 870–874. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Yang, B.; Tang, J.; Ma, Z.; Gao, Q.; Zhao, M. Structural analysis of water-soluble polysaccharides in the fruiting body of Dictyophora indusiata and their in vivo antioxidant activities. Carbohydr. Polym. 2012, 87, 343–347. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiao, J.; Liu, C.; Wu, X.; Zhang, Y. Isolation and purification of four flavone C-glycosides from antioxidant of bamboo leaves by macroporous resin column chromatography and preparative high-performance liquid chromatography. Food Chem. 2008, 107, 1326–1336. [Google Scholar] [CrossRef]
- Morimoto, Y.; Yasuhara, T.; Sugimoto, A.; Inoue, A.; Hide, I.; Akiyama, M.; Nakata, Y. Anti-allergic substances contained in the pollen of Cryptomeria japonica possess diverse effects on the degranulation of RBL-2H3 cells. J. Pharmacol. Sci. 2003, 92, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Park, Y.D.; Han, J.M.; Im, K.R.; Lee, B.W.; Jeong, I.Y.; Jeong, T.S.; Lee, W.S. Anti-atherosclerotic and anti-inflammatory activities of catecholic xanthones and flavonoids isolated from Cudrania tricuspidata. Bioorg. Med. Chem. Lett. 2006, 16, 5580–5583. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.M.; Aller, P. Quercetin decreases intracellular GSH content and potentiates the apoptotic action of the antileukemic drug arsenic trioxide in human leukemia cell lines. Biochem. Pharmacol. 2008, 75, 1912–1923. [Google Scholar] [CrossRef] [PubMed]
- Reddy, B.P.; Krupadanam, G.L.D. Chemical constituents of the leaves of Himalayan Taxus baccata: Use of DQF-COSY in the structure elucidation of biflavones. Indian J. Chem. 1996, 35, 283–285. [Google Scholar] [CrossRef]
- Parmar, V.S.; Jha, A. Chemical constituents of Taxus species. Stud. Nat. Prod. Chem. 1998, 20, 79–134. [Google Scholar]
- Van Rozendaal, E.L.; Lelyveld, G.P.; van Beek, T.A. Screening of the needles of different yew species and cultivars for paclitaxel and related taxoids. Phytochemistry 2000, 53, 383–389. [Google Scholar] [CrossRef]
- Afreen, F.; Zobayed, S.M.A.; Kozai, T. Spectral quality and UV-B stress stimulate glycyrrhizin concentration of Glycyrrhiza uralensis in hydroponic and pot system. Plant Physiol. Biochem. 2005, 43, 1074–1081. [Google Scholar] [CrossRef] [PubMed]
- Puttarak, P.; Panichayupakaranant, P. Factors affecting the content of pentacyclic triterpenes in Centella asiatica raw materials. Pharm. Biol. 2012, 50, 1508–1512. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, S.; Galasso, S.; Piccolella, S.; Kretschmer, N.; Pan, S.P.; Marciano, S.; Bauer, R.; Monaco, P. Seasonal variation in phenolic composition and antioxidant and anti-inflammatory activities of Calamintha nepeta (L.) Savi. Food Res. Int. 2015, 69, 121–132. [Google Scholar] [CrossRef]
- Yang, F.J.; Pang, H.H.; Zhang, X.K.; Sun, J.Y.; Zu, Y.G. Quantitative changes of anti-cancer active components in Taxus chinensis var. mairei. Chin. J. Appl. Ecol. 2008, 19, 911–914. [Google Scholar]
- Wang, C.W.; Peng, S.L.; Li, M.G.; Li, W.J.; Lin, F.L. Review of factors affecting the taxoid content of Taxus spp. Acta Ecol. Sin. 2006, 26, 1583–1590. [Google Scholar]
- Yang, F.J.; Pang, H.H.; Zu, Y.G.; Zhang, X.K.; Gao, Y.X. Relationships between the growth and taxol content of Taxus chinensis var. mairei and environment factors. Bull. Bot. Res. 2010, 30, 742–746. [Google Scholar]
- Kelsey, R.G.; Vance, N.C. Taxol and cephalomannine concentrations in the foliage and bark of shade-grown and sun-exposed Taxus brevifolia trees. J. Nat. Prod. 1992, 55, 912–917. [Google Scholar] [CrossRef]
- Griffin, J.; Hook, I. Taxol content of Irish yews. Planta Med. 1996, 62, 370–372. [Google Scholar] [CrossRef] [PubMed]
- Vance, N.C.; Kelsey, R.G.; Sabin, T.E. Seasonal and tissue variation in taxane concentrations of Taxus brevifolia. Phytochemistry 1994, 36, 1241–1244. [Google Scholar] [CrossRef]
- Ballero, M.; Loi, M.C.; van Rozendaal, E.L.M.; van Beek, T.A.; van de Haar, C.; Poli, F.; Appendino, G. Analysis of pharmaceutically relevant taxoids in wild yew trees from Sardinia. Fitoterapia 2003, 74, 34–39. [Google Scholar] [CrossRef]
- Wheeler, N.C.; Jech, K.; Masters, S.; Brobst, S.W.; Alvarado, A.B.; Hoover, A.J.; Snader, K.M. Effects of genetic, epigenetic, and environmental factors on taxol content in Taxus brevifolia and related species. J. Nat. Prod. 1992, 55, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Li, J.C.; Ma, Z.H.; Yuan, Y.J.; Sun, A.C.; Hu, C.X. Dynamic effects of cerium on syntheses of soluble protein and taxol in suspension culture of Taxus chinensis var. mairei cells. J. Rare Earths 2001, 19, 223–228. [Google Scholar]
- Zhang, H.; Yang, M.H. Several affecting factors of taxol contents of yew bark. Chin. Trad. Herb. Drugs 2002, 33, 39–41. [Google Scholar]
- Martin, D.; Tholl, D.; Gershenzon, J.; Bohlmann, J. Methyl jasmonate induces traumatic resin ducts, terpenoid resin biosynthesis, and terpenoid accumulation in developing xylem of Norway spruce stems. Plant Physiol. 2002, 129, 1003–1018. [Google Scholar] [CrossRef] [PubMed]
- Castelo, A.V.M.; DelMenezzi, C.H.S.; Resck, I.S. Seasonal variation in the yield and the chemical composition of essential oils from two Brazilian native arbustive species. J. Appl. Sci. 2012, 12, 753. [Google Scholar] [CrossRef]
- Chaves, N.; Escudero, J.C.; Gutiérrez-Merino, C. Seasonal variation of exudate of Cistus ladanifer. J. Chem. Ecol. 1993, 19, 2577–2591. [Google Scholar] [CrossRef] [PubMed]
- Lajis, M.; Abas, F.; Neto, V.; Khatib, A.; Shaari, K.; Shitan, M.; Kneer, R. Discrimination of three Pegaga (Centella) varieties and determination of growth-lighting effects on metabolites content based on chemometry of 1H nuclear magnetic resonance spectroscopy. J. Agric. Food Chem. 2012, 60, 410–417. [Google Scholar]
- Alqahtani, A.; Tongkao-On, W.; Li, K.M.; Razmovski-Naumovski, V.; Chan, K.; Li, G.Q. Seasonal variation of triterpenes and phenolic compounds in Australian Centella asiatica (L.) Urb. Phytochem. Anal. 2015, 26, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Tomás-Lorente, F.; Garcia-Grau, M.M.; Tomás-Barberán, F.A. Flavonoids from Vicia faba seed exudates. Z. Naturforsch. C 1990, 45, 1070–1072. [Google Scholar]
- Chaves, N.; Escudero, J.C.; Gutierrez-Merino, C. Role of ecological variables in the seasonal variation of flavonoid content of Cistus ladanifer exudate. J. Chem. Ecol. 1997, 23, 579–603. [Google Scholar] [CrossRef]
- Fu, X.; Zhou, X.; Deng, B.; Shang, X.; Fang, S. Seasonal and genotypic variation of water-soluble polysaccharide content in leaves of Cyclocarya paliurus. South. For. 2015, 77, 231–236. [Google Scholar]
- Cui, G.; Zhang, W.; Zhang, A.; Mu, H.; Bai, H.; Duan, J.; Wu, C. Variation in antioxidant activities of polysaccharides from Fructus Jujubae in South Xinjiang area. Int. J. Biol. Macromol. 2013, 57, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Ge, G.B.; Luan, H.W.; Zhang, Y.Y.; He, Y.Q.; Liu, X.B.; Yang, L.; Wang, Z.T.; Yang, L. Profiling of yew hair roots from various species using ultra-performance liquid chromatography/electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2008, 22, 2315–2323. [Google Scholar] [CrossRef] [PubMed]
- Ruan, X.; Zhan, L.M.; Gao, X.X.; Yan, L.Y.; Zhang, H.; Zhu, Z.Y.; Wang, Q.; Jiang, D.A. Separation and purification of flavonoid from Taxus remainder extracts free of taxoids using polystyrene and polyamide resin. J. Sep. Sci. 2013, 36, 1925–1934. [Google Scholar] [PubMed]
- Wei, X.; Xi, X.; Wu, M.; Wang, Y. A novel method for quantitative determination of tea polysaccharide by resonance light scattering. Spectrochim. Acta Mol. Biomol. Spectrosc. 2011, 79, 928–933. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds paclitaxel, 10-deacetylbaccatin III, baccatin III, cephalomannine, 10-deacetyltaxol, ginkgetin, amentoflavone and quercetin are available from the authors.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Collection Date | Content of Investigated Taxoids (mg·g−1) | Content of Investigated Flavonoids (mg·g−1) | Content of Investigated Polysaccharides (mg·g−1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 10-DAB | Baccatin | 10-DAT | Cephalomannine | Paclitaxel | Sum | Quercetin | Amentoflavone | Ginkgetin | Total Flavonoids | Polysaccharides | |
| January | 0.76 ± 0.06a | 0.47 ± 0.12b | 0.18 ± 0.00abc | 0.10 ± 0.02bc | 0.26 ± 0.05abc | 1.77 ± 0.38a | 0.12 ± 0.01ab | 0.66 ± 0.01ab | 0.15 ± 0.05bcd | 3.91 ± 0.69de | 9.39 ± 0.17j |
| February | 0.41 ± 0.04bcd | 0.54 ± 0.13ab | 0.23 ± 0.08ab | 0.12 ± 0.00ab | 0.40 ± 0.08a | 1.71 ± 0.51ab | 0.10 ± 0.01abc | 0.80 ± 0.02a | 0.20 ± 0.09bc | 5.97 ± 0.87cde | 12.89 ± 1.76i |
| March | 0.32 ± 0.06bcd | 0.35 ± 0.01bc | 0.16 ± 0.05abc | 0.12 ± 0.01ab | 0.25 ± 0.04bcd | 1.19 ± 0.14bcd | 0.10 ± 0.02abc | 0.45 ± 0.02abc | 0.12 ± 0.03bcd | 3.27 ± 0.73e | 16.12 ± 0.63h |
| April | 0.18 ± 0.07d | 0.32 ± 0.01bc | 0.13 ± 0.07abc | 0.08 ± 0.04bc | 0.13 ± 0.04cd | 0.84 ± 0.19de | 0.06 ± 0.01bc | 0.30 ± 0.00bc | 0.11 ± 0.01bcd | 4.30 ± 0.18de | 19.59 ± 0.77fg |
| May | 0.48 ± 0.03b | 0.43 ± 0.08bc | 0.16 ± 0.04abc | 0.10 ± 0.00bc | 0.26 ± 0.04abc | 1.44 ± 0.06abc | 0.14 ± 0.05a | 0.49 ± 0.02abc | 0.08 ± 0.00cd | 13.75 ± 1.64a | 21.43 ± 0.62ef |
| June | 0.46 ± 0.03bcd | 0.47 ± 0.13b | 0.26 ± 0.03a | 0.12 ± 0.04ab | 0.19 ± 0.02bcd | 1.50 ± 0.51abc | 0.16 ± 0.00a | 0.31 ± 0.00bc | 0.11 ± 0.05bcd | 13.06 ± 0.73a | 22.70 ± 0.13de |
| July | 0.17 ± 0.06d | 0.45 ± 0.05bc | 0.14 ± 0.02abc | 0.09 ± 0.00bc | 0.18 ± 0.00bcd | 1.03 ± 0.11cde | 0.14 ± 0.02a | 0.38 ± 0.00bc | 0.13 ± 0.00bcd | 11.26 ± 1.22b | 24.30 ± 0.96cd |
| August | 0.26 ± 0.02bcd | 0.39 ± 0.08bc | 0.10 ± 0.06bc | 0.12 ± 0.01ab | 0.21 ± 0.02bcd | 1.09 ± 0.13cde | 0.10 ± 0.00abc | 0.45 ± 0.01abc | 0.42 ± 0.15a | 15.82 ± 1.45a | 23.81 ± 1.08d |
| September | 0.19 ± 0.13cd | 0.20 ± 0.02c | 0.07 ± 0.02c | 0.04 ± 0.00c | 0.10 ± 0.04d | 0.61 ± 0.08d | 0.04 ± 0.00c | 0.46 ± 0.03abc | 0.16 ± 0.03bcd | 9.72 ± 0.42bc | 28.52 ± 0.57a |
| October | 0.33 ± 0.07bcd | 0.56 ± 0.12ab | 0.14 ± 0.05abc | 0.13 ± 0.00ab | 0.23 ± 0.00bcd | 1.39 ± 0.04abc | 0.12 ± 0.01ab | 0.30 ± 0.00bc | 0.23 ± 0.10bc | 8.66 ± 1.40bc | 27.33 ± 0.98ab |
| November | 0.51 ± 0.03b | 0.78 ± 0.22a | 0.17 ± 0.01abc | 0.08 ± 0.07bc | 0.17 ± 0.06bcd | 1.71 ± 0.31ab | 0.06 ± 0.01bc | 0.20 ± 0.00c | 0.27 ± 0.10b | 7.02 ± 0.93bcd | 26.03 ± 0.37bc |
| December | 0.47 ± 0.02bc | 0.57 ± 0.04ab | 0.12 ± 0.01abc | 0.18 ± 0.03a | 0.30 ± 0.03ab | 1.64 ± 0.01ab | 0.06 ± 0.00bc | 0.11 ± 0.00c | 0.03 ± 0.00d | 7.18 ± 1.62bcd | 18.29 ± 1.59g |
| Month | Related Season | Mean of Maximum Temperature (°C) | Mean of Minimum Temperature (°C) | Daylight (min) | Relative Humidity (%) | Total Rainfall (mm) |
|---|---|---|---|---|---|---|
| December | Winter | 11.64 | 2.31 | 173.1 | 64.20 | 114.32 |
| January | 13.26 | 2.94 | 122.76 | 74.77 | 34.07 | |
| February | 10.25 | 4.46 | 46.02 | 80.18 | 89.16 | |
| March | Spring | 17.47 | 8.53 | 171.04 | 69.80 | 114.50 |
| April | 21.13 | 13.06 | 192.83 | 60.03 | 112.52 | |
| May | 26.02 | 18.18 | 192.38 | 74.53 | 73.59 | |
| June | Summer | 27.39 | 21.23 | 100.60 | 82.36 | 324.20 |
| July | 32.88 | 25.40 | 338.29 | 63.32 | 69.28 | |
| August | 30.87 | 24.73 | 263.14 | 67.23 | 251.64 | |
| September | Autumn | 28.20 | 22.29 | 223.75 | 71.02 | 32.89 |
| October | 25.29 | 16.55 | 161.32 | 73.37 | 424.00 | |
| November | 19.57 | 11.68 | 178.30 | 67.40 | 42.64 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Zheng, Z.-S.; Cheng, F.; Ruan, X.; Jiang, D.-A.; Pan, C.-D.; Wang, Q. Seasonal Dynamics of Metabolites in Needles of Taxus wallichiana var. mairei. Molecules 2016, 21, 1403. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21101403
Yang L, Zheng Z-S, Cheng F, Ruan X, Jiang D-A, Pan C-D, Wang Q. Seasonal Dynamics of Metabolites in Needles of Taxus wallichiana var. mairei. Molecules. 2016; 21(10):1403. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21101403
Chicago/Turabian StyleYang, Li, Zan-Sheng Zheng, Fang Cheng, Xiao Ruan, De-An Jiang, Cun-De Pan, and Qiang Wang. 2016. "Seasonal Dynamics of Metabolites in Needles of Taxus wallichiana var. mairei" Molecules 21, no. 10: 1403. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21101403