
MRI Reporter Genes for Noninvasive Molecular Imaging

Abstract
:1. Introduction
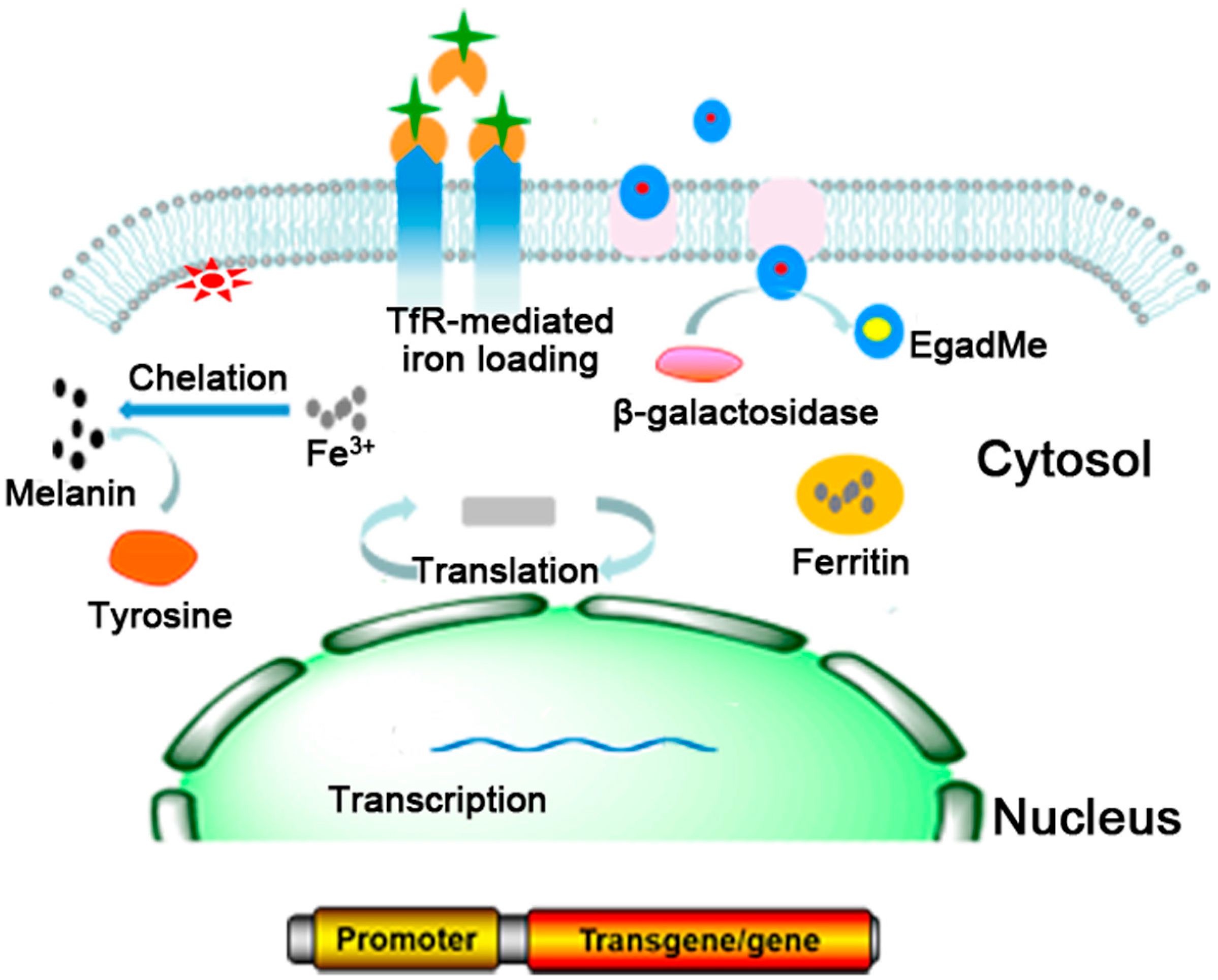
2. Commonly Used MRI Reporter Genes
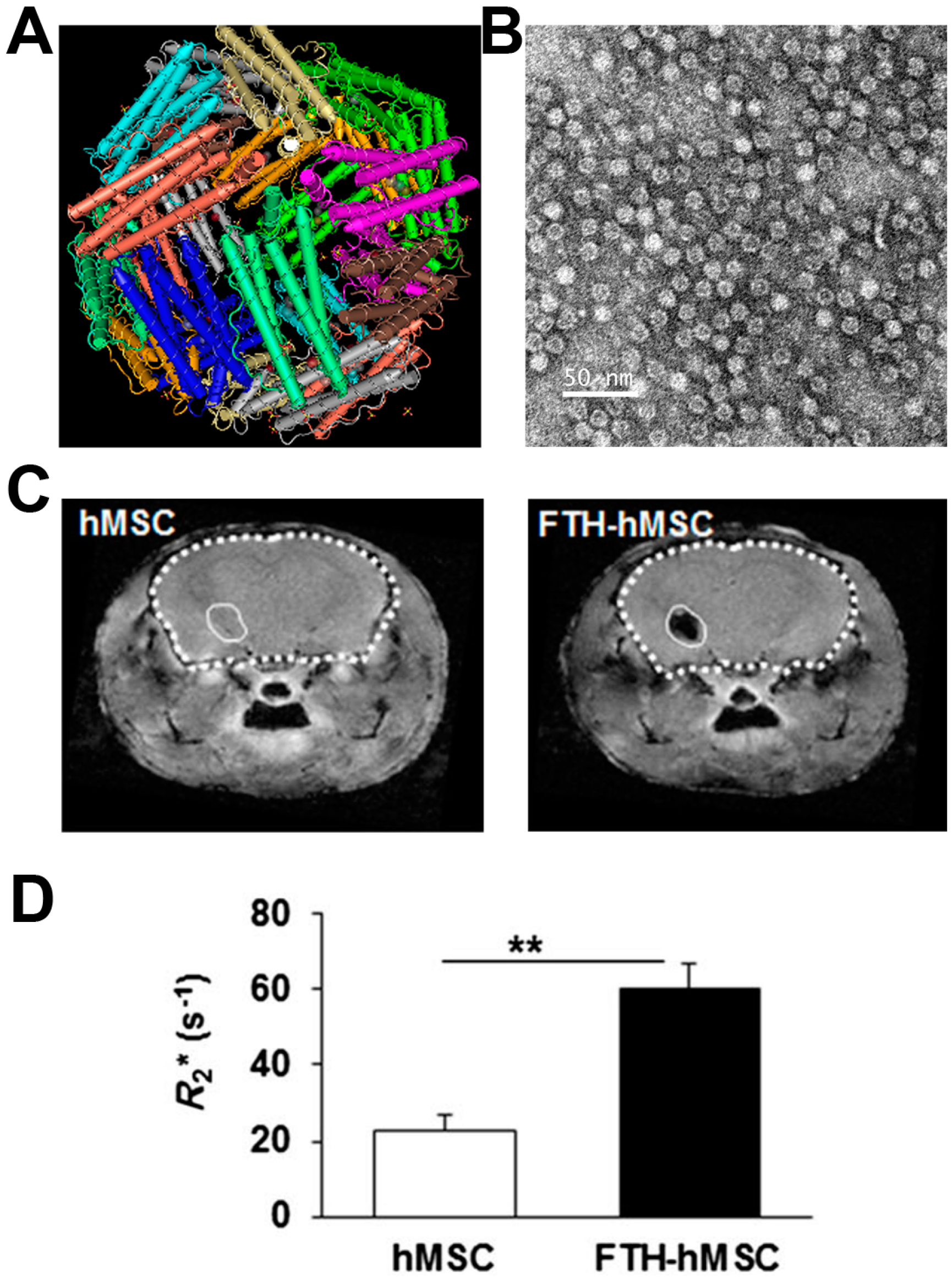
2.1. Ferritin
2.2. TfR
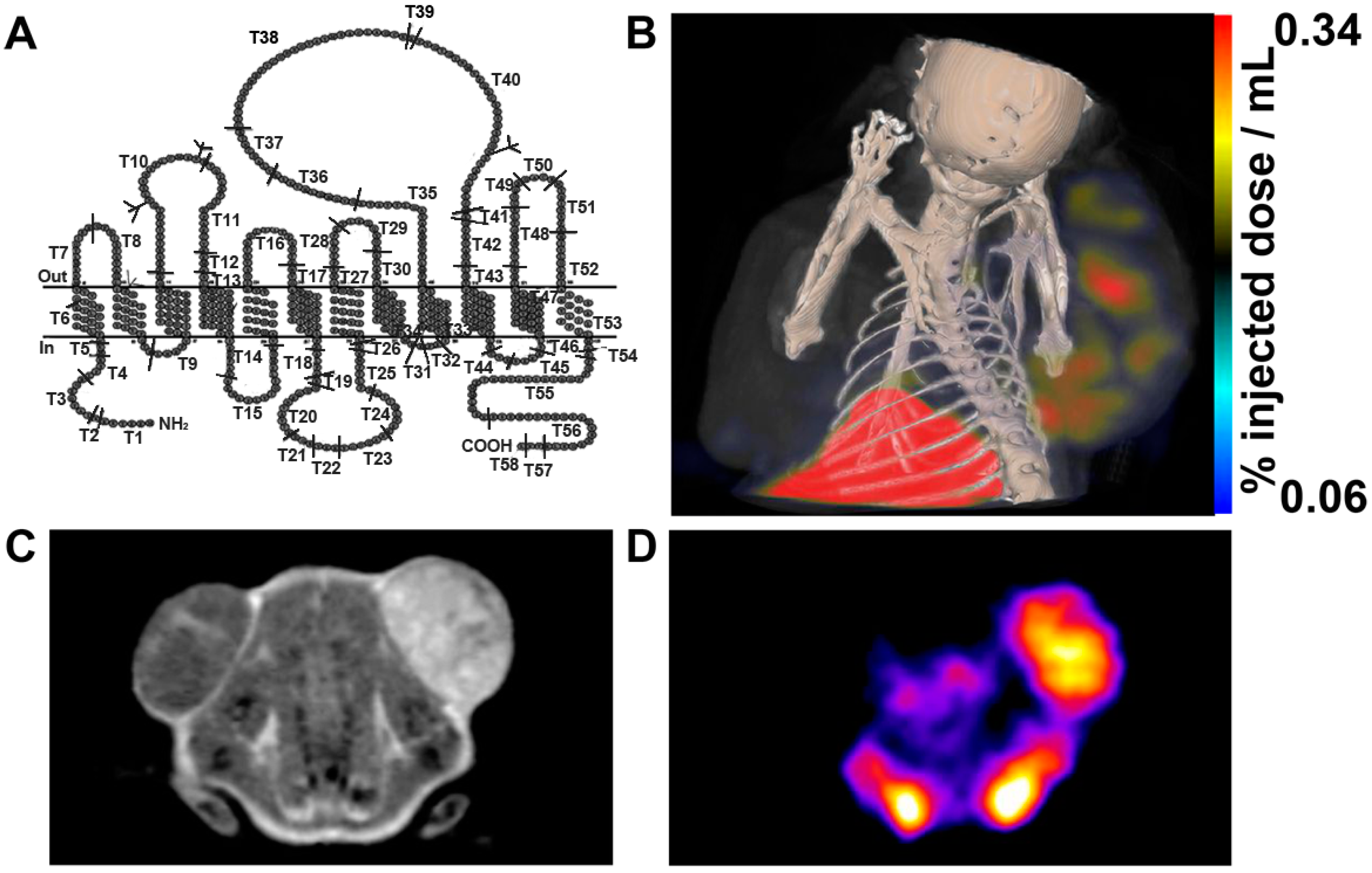
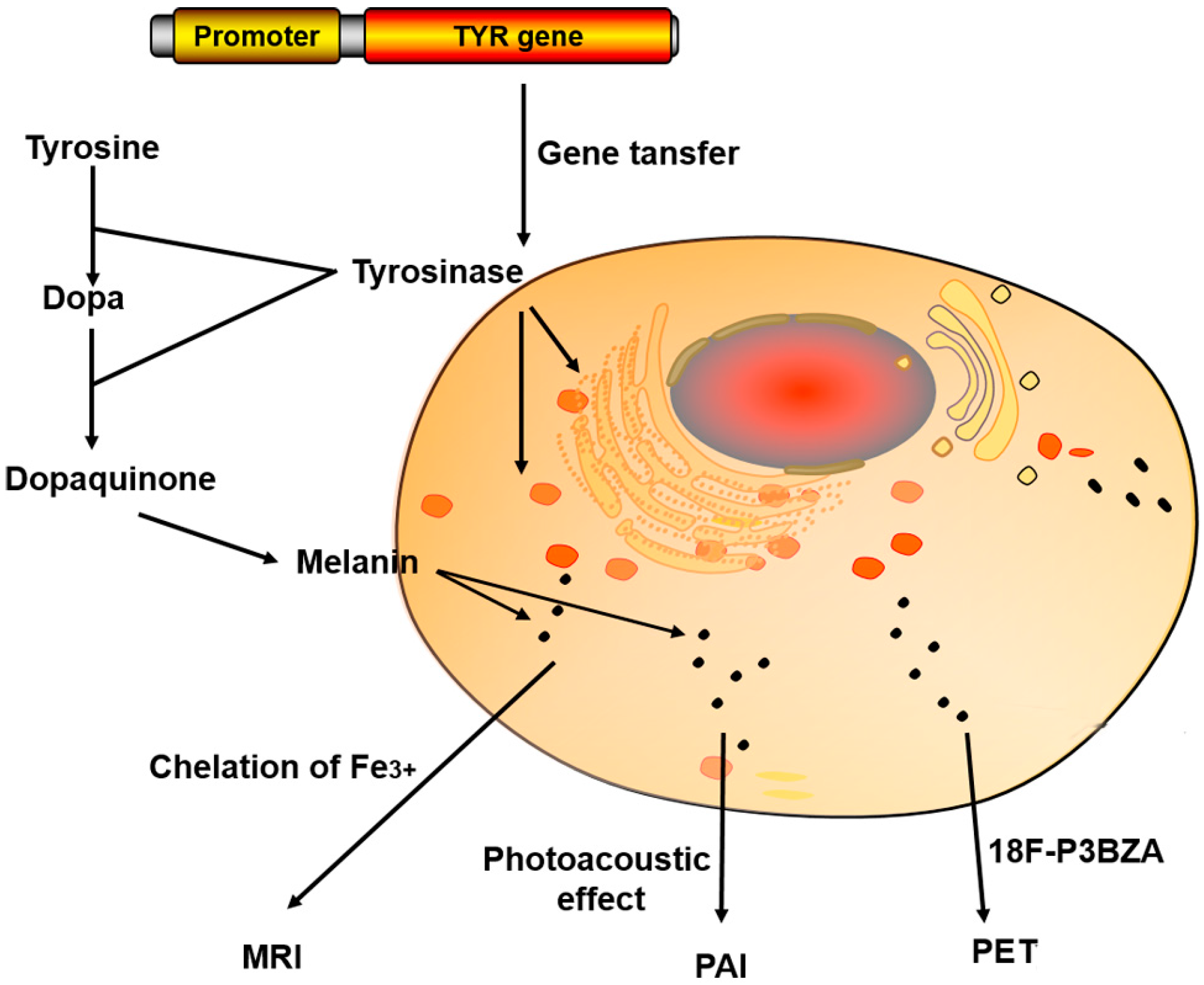
2.3. TYR
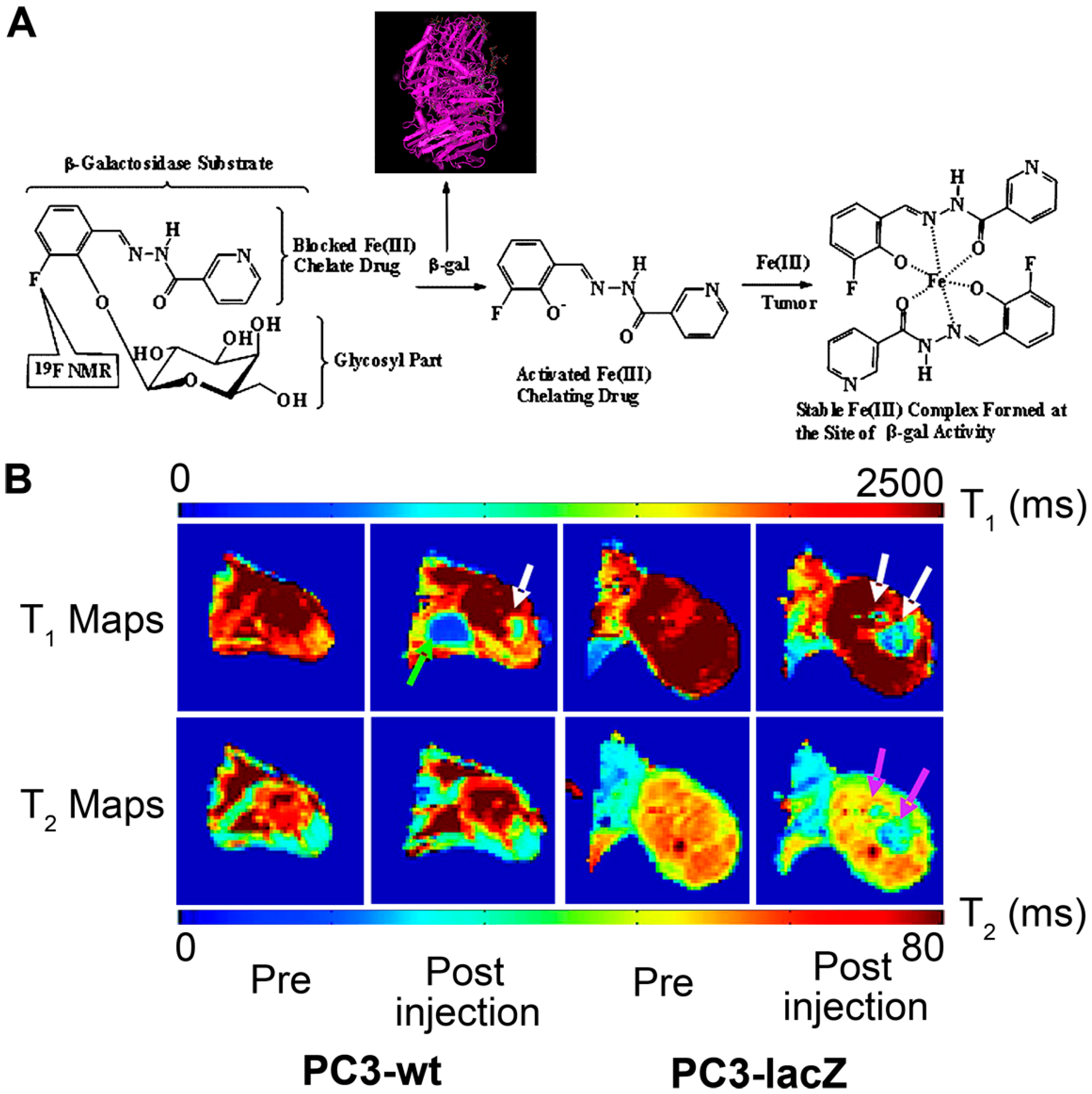
2.4. β-Galactosidase
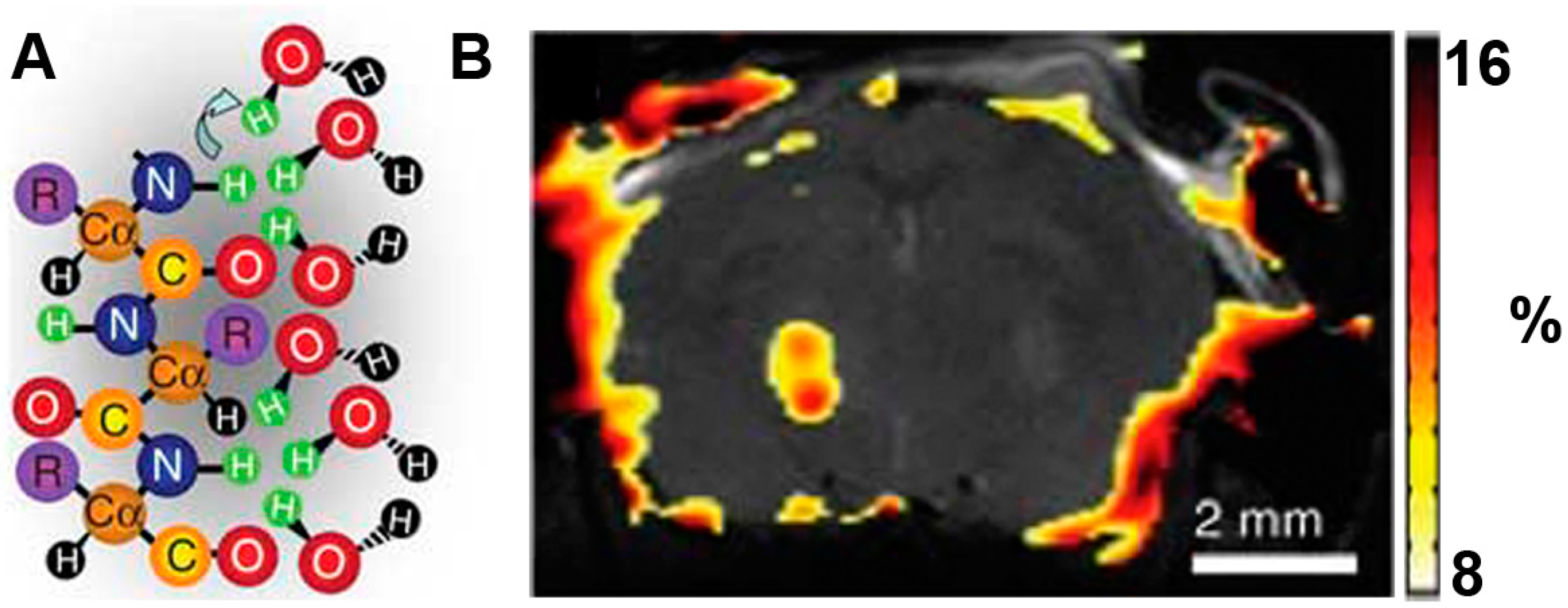
2.5. Lysine Rich-Protein (LRP)
3. Multimodality Imaging Reporter Genes
3.1. Single Reporter Genes for Multimodality Imaging
3.1.1. The Organic Anion Transporting Polypeptides (Oatp) 1
3.1.2. LacZ
3.1.3. TYR
3.1.4. Combination of Single Reporter Gene and Contrast Agents for Multimodality Imaging
3.2. Fusion Reporter Gene Used for Multimodality Imaging
4. Conclusions and Perspectives
Acknowledgments
Conflicts of Interest
References
- Oh, S.W.; Hwang do, W.; Lee, D.S. In vivo monitoring of microRNA biogenesis using reporter gene imaging. Theranostics 2013, 3, 1004–1011. [Google Scholar] [CrossRef] [PubMed]
- Deliolanis, N.C.; Ale, A.; Morscher, S.; Burton, N.C.; Schaefer, K.; Radrich, K.; Razansky, D.; Ntziachristos, V. Deep-tissue reporter-gene imaging with fluorescence and optoacoustic tomography: A performance overview. Mol. Imaging Biol. 2014, 16, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Vandsburger, M. Cardiac cell tracking with MRI reporter genes: Welcoming a new field. Curr. Cardiovasc. Imaging Rep. 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Chung, J.K. Molecular-genetic imaging based on reporter gene expression. J. Nucl. Med. 2008, 49 (Suppl. 2), 164S–179S. [Google Scholar] [CrossRef] [PubMed]
- Harney., A.S.; Meade., T.J. Molecular imaging of in vivo gene expression. Future Med. Chem. 2010, 2, 503–519. [Google Scholar] [CrossRef] [PubMed]
- Iordanova, B.; Ahrens, E.T. In vivo magnetic resonance imaging of ferritin-based reporter visualizes native neuroblast migration. NeuroImage 2012, 59, 1004–1012. [Google Scholar] [CrossRef] [PubMed]
- Xenopoulos, P.; Nowotschin, S.; Hadjantonakis, A.K. Live imaging fluorescent proteins in early mouse embryos. Methods Enzymol. 2012, 506, 361–389. [Google Scholar] [PubMed]
- Serganova, I.; Blasberg, R. Reporter gene imaging: Potential impact on therapy. Nucl. Med. Biol. 2005, 32, 763–780. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Cai, J.; Liu, B.; Zhong, Y.; Qin, Y. Cellular magnetic resonance imaging contrast generated by the ferritin heavy chain genetic reporter under the control of a Tet-On switch. Stem Cell Res. Ther. 2015, 6. [Google Scholar] [CrossRef]
- Cai, Y.; Cao, C.; He, X.; Yang, C.; Tian, L.; Zhu, R.; Pan, Y. Enhanced magnetic resonance imaging and staining of cancer cells using ferrimagnetic H-ferritin nanoparticles with increasing core size. Int. J. Nanomed. 2015, 10, 2619–2634. [Google Scholar]
- Lin, X.; Xie, J.; Niu, G.; Zhang, F.; Gao, H.; Yang, M.; Quan, Q.; Aronova, M.A.; Zhang, G.; Lee, S.; et al. Chimeric ferritin nanocages for multiple function loading and multimodal imaging. Nano Lett. 2011, 11, 814–819. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.; Basilion, J.P.; Chiocca, E.A.; Weissleder, R. Measuring transferrin receptor gene expression by NMR imaging. Biochim. Biophys. Acta 1998, 1402, 239–249. [Google Scholar] [CrossRef]
- Richardson, D.R.; Ponka, P. The molecular mechanisms of the metabolism and transport of iron in normal and neoplastic cells. Biochim. Biophys. Acta 1997, 1331. [Google Scholar] [CrossRef]
- Bengtsson, N.E.; Brown, G.; Scott, E.W.; Walter, G.A. lacZ as a genetic reporter for real-time MRI. Magn. Reson. Med. 2010, 63, 745–753. [Google Scholar] [PubMed]
- Feng, H.; Xia, X.; Li, C.; Song, Y.; Qin, C.; Zhang, Y.; Lan, X. TYR as a multifunctional reporter gene regulated by the Tet-on system for multimodality imaging: An in vitro study. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Genove, G.; DeMarco, U.; Xu, H.; Goins, W.F.; Ahrens, E.T. A new transgene reporter for in vivo magnetic resonance imaging. Nat. Med. 2005, 11, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Weissleder, R.; Moore, A.; Mahmood, U.; Bhorade, R.; Benveniste, H.; Chiocca, E.A.; Basilion, J.P. In vivo magnetic resonance imaging of transgene expression. Nat. Med. 2000, 6, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Weissleder, R.; Simonova, M.; Bogdanova, A.; Bredow, S.; Enochs, W.S.; Bogdanov, A., Jr. MR imaging and scintigraphy of gene expression through melanin induction. Radiology 1997, 204, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Louie, A.Y.; Hüber, M.M.; Ahrens, E.T.; Rothbächer, U.; Moats, R.; Jacobs, R.E.; Fraser, S.E.; Meade, T.J. In vivo visualization of gene expression using magnetic resonance imaging. Nat. Biotechnol. 2000, 18, 321–325. [Google Scholar] [PubMed]
- Gilad, A.A.; McMahon, M.T.; Walczak, P.; Winnard, P.T., Jr.; Raman, V.; van Laarhoven, H.W.; Skoglund, C.M.; Bulte, J.W.; van Zijl, P.C. Artificial reporter gene providing MRI contrast based on proton exchange. Nat. Biotechnol. 2007, 25, 217–219. [Google Scholar] [CrossRef] [PubMed]
- Gilad, A.A.; Winnard, P.T., Jr.; van Zijl, P.C.; Bulte, J.W. Developing MR reporter genes: Promises and pitfalls. NMR Biomed. 2007, 20, 275–290. [Google Scholar] [CrossRef] [PubMed]
- Hill, P.J.; Stritzker, J.; Scadeng, M.; Geissinger, U.; Haddad, D.; Basse-Lusebrink, T.C.; Gbureck, U.; Jakob, P.; Szalay, A.A. Magnetic resonance imaging of tumors colonized with bacterial ferritin-expressing Escherichia coli. PLoS ONE 2011, 6, e25409. [Google Scholar] [CrossRef] [PubMed]
- Iordanova, B.; Hitchens, T.K.; Robison, C.S.; Ahrens, E.T. Engineered mitochondrial ferritin as a magnetic resonance imaging reporter in mouse olfactory epithelium. PLoS ONE 2013, 8, e72720. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Joo, H.J.; Woo, J.S.; Choi, Y.S.; Choi, S.H.; Kim, H.; Moon, W.K. In vivo magnetic resonance imaging of transgenic mice expressing human ferritin. Mol. Imaging Biol. 2013, 15, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Lee, S.H.; Biswal, S. Magnetic resonance reporter gene imaging. Theranostics 2012, 2, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Huang, P.; Jacobson, O.; Wang, Z.; Liu, Y.; Lin, L.; Lin, J.; Lu, N.; Zhang, H.; Tian, R.; et al. Biomineralization-inspired synthesis of copper sulfide-ferritin nanocages as cancer theranostics. ACS Nano 2016, 10, 3453–3460. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Fuma, K.; Tabata, K.; Sawada, M. Ferritin reporter used for gene expression imaging by magnetic resonance. Biochem. Biophys. Res. Commun. 2009, 388, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Liu, Q.; Zhu, J.; Xie, F.; Li, L. Efficiency of ferritin as an MRI reporter gene in NPC cells is enhanced by iron supplementation. J. Biomed. Biotechnol. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Shan, L. Bimodal lentiviral vector encoding myc-tagged human ferritin heavy chain and green fluorescent protein (GFP). In Molecular Imaging and Contrast Agent Database (MICAD); National Center for Biotechnology Information (US): Bethesda, MD, USA, 2004. [Google Scholar]
- Vande Velde, G.; Raman Rangarajan, J.; Vreys, R.; Guglielmetti, C.; Dresselaers, T.; Verhoye, M.; Van der Linden, A.; Debyser, Z.; Baekelandt, V.; Maes, F.; et al. Quantitative evaluation of MRI-based tracking of ferritin-labeled endogenous neural stem cell progeny in rodent brain. NeuroImage 2012, 62, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Gong, M.F.; Yang, H.; Zhang, S.; Wang, G.X.; Su, T.S.; Wen, L.; Zhang, D. MR molecular imaging of tumours using ferritin heavy chain reporter gene expression mediated by the hTERT promoter. Eur. Radiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Vande Velde, G.; Rangarajan, J.R.; Toelen, J.; Dresselaers, T.; Ibrahimi, A.; Krylychkina, O.; Vreys, R.; van der Linden, A.; Maes, F.; Debyser, Z.; et al. Evaluation of the specificity and sensitivity of ferritin as an MRI reporter gene in the mouse brain using lentiviral and adeno-associated viral vectors. Gene Ther. 2011, 18, 594–605. [Google Scholar] [CrossRef] [PubMed]
- Cohen, B.; Ziv, K.; Plaks, V.; Harmelin, A.; Neeman, M. Ferritin nanoparticles as magnetic resonance reporter gene. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2009, 1, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Zurkiya, O.; Chan, A.W.S.; Hu, X. MagA is sufficient for producing magnetic nanoparticles in mammalian cells, making it an MRI reporter. Magn. Reson. Med. 2008, 59, 1225–1231. [Google Scholar] [CrossRef] [PubMed]
- Tennstaedt, A.; Aswendt, M.; Adamczak, J.; Collienne, U.; Selt, M.; Schneider, G.; Henn, N.; Schaefer, C.; Lagouge, M.; Wiedermann, D.; et al. Human neural stem cell intracerebral grafts show spontaneous early neuronal differentiation after several weeks. Biomaterials 2015, 44, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Woo, J.; Lee, J.H.; Joo, H.J.; Choi, Y.; Kim, H.; Moon, W.K.; Kim, S.J. In vivo tracking of dendritic cell using mri reporter gene, ferritin. PLoS ONE 2015, 10, e0125291. [Google Scholar] [CrossRef] [PubMed]
- Ziv, K.; Meir, G.; Harmelin, A.; Shimoni, E.; Klein, E.; Neeman, M. Ferritin as a reporter gene for MRI: Chronic liver over expression of H-ferritin during dietary iron supplementation and aging. NMR Biomed. 2010, 23, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Cho, H.R.; Kim, H.S.; Kim, Y.H.; Kang, K.W.; Kim, H.; Moon, W.K. Imaging and quantification of metastatic melanoma cells in lymph nodes with a ferritin MR reporter in living mice. NMR Biomed. 2012, 25, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Woo, J.; Choi, Y.; Hwang, E.H.; Choi, S.K.; Cho, K.W.; Moon, W.K. Noninvasive MRI and multilineage differentiation capability of ferritin-transduced human mesenchymal stem cells. NMR Biomed. 2015, 28, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Cohen, B.; Dafni, H.; Meir, G.; Harmelin, A.; Neeman, M. Ferritin as an endogenous MRI reporter for noninvasive imaging of gene expression in C6 glioma tumors. Neoplasia 2005, 7, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, T.; Hogemann, D.; Saeki, Y.; Tyminski, E.; Terada, K.; Weissleder, R.; Chiocca, E.A.; Basilion, J.P. MRI of transgene expression: Correlation to therapeutic gene expression. Neoplasia 2002, 4, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Feelders, R.A.; Kuiper-Kramer, E.P.; van Eijk, H.G. Structure, function and clinical significance of transferrin receptors. Clin. Chem. Lab. Med. 1999, 37. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Hamada, Y.; Li, J.; Cong, L.; Wang, N.; Li, Y.; Zheng, W.; Jiang, X. Targeted tumor delivery and controlled release of neuronal drugs with ferritin nanoparticles to regulate pancreatic cancer progression. J. Control. Release 2016. [Google Scholar] [CrossRef] [PubMed]
- Schaffert, D.H.; Okholm, A.H.; Sorensen, R.S.; Nielsen, J.S.; Torring, T.; Rosen, C.B.; Kodal, A.L.; Mortensen, M.R.; Gothelf, K.V.; Kjems, J. Intracellular delivery of a planar DNA origami structure by the transferrin-receptor internalization pathway. Small 2016. [Google Scholar] [CrossRef] [PubMed]
- Vandsburger, M.H.; Radoul, M.; Cohen, B.; Neeman, M. MRI reporter genes: Applications for imaging of cell survival, proliferation, migration and differentiation. NMR Biomed. 2013, 26, 872–884. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Feng, G.; Zhang, C.J.; Cai, X.; Cheng, X.; Liu, B. Real-time specific light-up sensing of transferrin receptor (TfR): Image-guided photodynamic ablation of cancer cells through controlled cytomembrane disintegration. Anal. Chem. 2016. [Google Scholar] [CrossRef]
- Patrick, P.S.; Rodrigues, T.B.; Kettunen, M.I.; Lyons, S.K.; Neves, A.A.; Brindle, K.M. Development of Timd2 as a reporter gene for MRI. Magn. Reson. Med. 2016, 75, 1697–1707. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.M.; Herrmann, A.; Moss, D.; Poptani, H.; Williams, S.R.; Murray, P.; Taylor, A. Evaluating the effectiveness of transferrin receptor-1 (TfR1) as a magnetic resonance reporter gene. Contrast Media Mol. Imaging 2016. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.M.; Moss, D.; Williams, S.R.; Murray, P.; Taylor, A. Overexpression of the MRI reporter genes ferritin and transferrin receptor affect iron homeostasis and produce limited contrast in mesenchymal stem cells. Int. J. Mol. Sci. 2015, 16, 15481–15496. [Google Scholar] [CrossRef] [PubMed]
- Korkusuz, H.; Ulbrich, K.; Welzel, K.; Koeberle, V.; Watcharin, W.; Bahr, U.; Chernikov, V.; Knobloch, T.; Petersen, S.; Huebner, F. Transferrin-coated gadolinium nanoparticles as MRI contrast agent. Mol. Imaging Biol. 2013, 15, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, S.; Furukawa, T.; Saga, T. Molecular MR imaging of cancer gene therapy: Ferritin transgene reporter takes the stage. Magn. Reson. Med. Sci. 2010, 9, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Cheng, K.; Chen, K.; Hu, X.; Liu, Y.; Lan, X.; Zhang, Y.; Liu, H.; Xu, Y.; Bu, L.; et al. Tyrosinase as a multifunctional reporter gene for Photoacoustic/MRI/PET triple modality molecular imaging. Sci. Rep. 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Alfke, H.; Sto, H.; Nocken, F.; Heverhagen, J.T.; Kleb, B.; Czubayko, F.; Klose, K.J. In vitro MR imaging of regulated gene expression. Radiology 2003, 228, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Paproski, R.J.; Forbrich, A.E.; Wachowicz, K.; Hitt, M.M.; Zemp, R.J. Tyrosinase as a dual reporter gene for both photoacoustic and magnetic resonance imaging. Opt. Soc. Am. 2011, 2, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Enochs, W.S.; Petherick, P.; Bogdanova, A.; Mohr, U.; Weissleder, R. Paramagnetic metal scavenging by melanin: MR imaging. Radiology 1997, 204, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Tung, C.H.; Zeng, Q.; Shah, K.; Kim, D.E.; Schellingerhout, D.; Weissleder, R. In vivo imaging of beta-galactosidase activity using far red fluorescent switch. Cancer Res. 2004, 64, 1579–1583. [Google Scholar] [CrossRef] [PubMed]
- Josserand, V.; Texier-Nogues, I.; Huber, P.; Favrot, M.C.; Coll, J.L. Non-invasive in vivo optical imaging of the lacZ and luc gene expression in mice. Gene Ther. 2007, 14, 1587–1593. [Google Scholar] [CrossRef] [PubMed]
- Arena, F.; Singh, J.B.; Gianolio, E.; Stefania, R.; Aime, S. beta-Gal gene expression MRI reporter in melanoma tumor cells. Design, synthesis, and in vitro and in vivo testing of a Gd(III) containing probe forming a high relaxivity, melanin-like structure upon beta-Gal enzymatic activation. Bioconjug. Chem. 2011, 22, 2625–2635. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.E.; Jeong, S.W. Molecular imaging of atherosclerosis. J. Korean Med. Assoc. 2009, 52, 143–150. [Google Scholar] [CrossRef]
- Yu, J.X.; Kodibagkar, V.D.; Hallac, R.R.; Liu, L.; Mason, R.P. Dual 19F/1H MR gene reporter molecules for in vivo detection of beta-galactosidase. Bioconjug. Chem. 2012, 23, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Goffeney, N.; Bulte, J.W.; Duyn, J.; Bryant, L.H., Jr.; van Zijl, P.C. Sensitive NMR detection of cationic-polymer-based gene delivery systems using saturation transfer via proton exchange. J. Am. Chem. Soc. 2001, 123, 8628–8629. [Google Scholar] [CrossRef] [PubMed]
- Ward, K.M.; Aletras, A.H.; Balaban, R.S. A new class of contrast agents for MRI based on proton chemical exchange dependent saturation transfer (CEST). J. Magn. Reson. 2000, 143, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Bulte, J.W.; Gilad, A.A. CEST MRI reporter genes. Methods Mol. Biol. 2011, 711, 271–280. [Google Scholar] [PubMed]
- Farrar, C.T.; Buhrman, J.S.; Liu, G.; Kleijn, A.; Lamfers, M.L.; McMahon, M.T.; Gilad, A.A.; Fulci, G. Establishing the lysine-rich protein CEST reporter gene as a CEST MR imaging detector for oncolytic virotherapy. Radiology 2015, 275, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Longo, D.L.; Sun, P.Z.; Consolino, L.; Michelotti, F.C.; Uggeri, F.; Aime, S. A general MRI-CEST ratiometric approach for pH imaging: Demonstration of in vivo pH mapping with iobitridol. J. Am. Chem. Soc. 2014, 136, 14333–14336. [Google Scholar] [CrossRef] [PubMed]
- Sheth, V.R.; Li, Y.; Chen, L.Q.; Howison, C.M.; Flask, C.A.; Pagel, M.D. Measuring in vivo tumor pHe with CEST-FISP MRI. Magn. Reson. Med. 2012, 67, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.K.; Schlosser, M.J.; van Zijl, P.C.; Pomper, M.G.; Golay, X.; Zhou, J. Amide proton transfer imaging of human brain tumors at 3T. Magn. Reson. Med. 2006, 56, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.Z.; Zhou, J.; Sun, W.; Huang, J.; van Zijl, P.C. Detection of the ischemic penumbra using pH-weighted MRI. J. Cereb. Blood Flow Metab. 2007, 27, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Bar-Shir, A.; Liang, Y.; Chan, K.W.; Gilad, A.A.; Bulte, J.W. Supercharged green fluorescent proteins as bimodal reporter genes for CEST MRI and optical imaging. Chem. Commun. 2015, 51, 4869–4871. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.H.; Wang, Y.; Zhang, F.; Zhang, X.; Lu, L.J.; Shuai, X.T.; Shen, J. In vivo monitoring of neural stem cells after transplantation in acute cerebral infarction with dual-modal MR imaging and optical imaging. Biomaterials 2014, 35, 4627–4635. [Google Scholar] [CrossRef] [PubMed]
- Riegler, J.; Liew, A.; Hynes, S.O.; Ortega, D.; O’Brien, T.; Day, R.M.; Richards, T.; Sharif, F.; Pankhurst, Q.A.; Lythgoe, M.F. Superparamagnetic iron oxide nanoparticle targeting of MSCs in vascular injury. Biomaterials 2013, 34, 1987–1994. [Google Scholar] [CrossRef] [PubMed]
- De Jong, M.; Essers, J.; van Weerden, W.M. Imaging preclinical tumour models: Improving translational power. Nat. Rev. Cancer 2014, 14, 481–493. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-F.; Liu, T.; Guo, Y.-L.; Gao, F.-B. Construction and identification of the adenoviral vector with dual reporter gene for multimodality molecular imaging. J. Huazhong Univ. Sci. Technol. 2013, 33, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Cho, H.R.; Choi, S.H.; Woo, J.S.; Moon, W.K. In vivo imaging of tumor transduced with bimodal lentiviral vector encoding human ferritin and green fluorescent protein on a 1.5T clinical magnetic resonance scanner. Cancer Res. 2010, 70, 7315–7324. [Google Scholar] [CrossRef] [PubMed]
- Bhirde, A.; Xie, J.; Swierczewska, M.; Chen, X. Nanoparticles for cell labeling. Nanoscale 2011, 3, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Shichinohe, H.; Yamauchi, T.; Saito, H.; Houkin, K.; Kuroda, S. Bone marrow stromal cell transplantation enhances recovery of motor function after lacunar stroke in rats. Acta Neurobiol. Exp. 2013, 73, 354–363. [Google Scholar]
- Louie, A. Multimodality imaging probes: Design and challenges. Chem. Rev. 2010, 110, 3146–3195. [Google Scholar] [CrossRef] [PubMed]
- Ray, P. Multimodality molecular imaging of disease progression in living subjects. J. Biosci. 2011, 36, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Patrick, P.S.; Hammersley, J.; Loizou, L.; Kettunen, M.I.; Rodrigues, T.B.; Hu, D.E.; Tee, S.S.; Hesketh, R.; Lyons, S.K.; Soloviev, D.; et al. Dual-modality gene reporter for in vivo imaging. Proc. Natl. Acad. Sci. USA 2014, 111, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Patrick, P.S.; Lyons, S.K.; Rodrigues, T.B.; Brindle, K.M. Oatp1 enhances bioluminescence by acting as a plasma membrane transporter for d-luciferin. Mol. Imaging Biol. 2014, 16, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Wang, L.V. Photoacoustic imaging in biomedicine. Rev. Sci. Instrum. 2006, 77. [Google Scholar] [CrossRef]
- Krumholz, A.; Vanvickle-Chavez, S.J.; Yao, J.; Fleming, T.P.; Gillanders, W.E.; Wang, L.V. Photoacoustic microscopy of tyrosinase reporter gene in vivo. J. Biomed. Opt. 2011, 16. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, S.; Miao, Z.; Jiang, H.; Deng, Z.; Hong, X.; Cheng, Z. A novel aliphatic 18F-labeled probe for PET imaging of melanoma. Mol. Pharm. 2013, 10, 3384–3391. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, T.; Anton, M.; Dumler, K.; Seidl, S.; Pelisek, J.; Saraste, A.; Welling, A.; Hofmann, F.; Oostendorp, R.A.; Gansbacher, B.; et al. Combined reporter gene PET and iron oxide MRI for monitoring survival and localization of transplanted cells in the rat heart. J. Nucl. Med. 2009, 50, 1088–1094. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Nieves, E.; Angeletti, R.H.; Orr, G.A.; Wolkoff, A.W. Rat organic anion transporting protein 1A1 (Oatp1a1): Purification and phosphopeptide assignment. Biochemistry 2006, 45, 3357–3369. [Google Scholar] [CrossRef] [PubMed]
- De la Zerda, A.; Zavaleta, C.; Keren, S.; Vaithilingam, S.; Bodapati, S.; Liu, Z.; Levi, J.; Smith, B.R.; Ma, T.J.; Oralkan, O.; et al. Carbon nanotubes as photoacoustic molecular imaging agents in living mice. Nat. Nanotechnol. 2008, 3, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Jarrett, B.R.; Kauzlarich, S.M.; Louie, A.Y. Core/shell quantum dots with high relaxivity and photoluminescence for multimodality imaging. J. Am. Chem. Soc. 2007, 129, 3848–3856. [Google Scholar] [CrossRef] [PubMed]
- Ashwal, S.; Ghosh, N.; Turenius, C.I.; Dulcich, M.; Denham, C.M.; Tone, B.; Hartman, R.; Snyder, E.Y.; Obenaus, A. Reparative effects of neural stem cells in neonatal rats with hypoxic-ischemic injury are not influenced by host sex. Pediatr. Res. 2014, 75, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Jeon, Y.H.; Hwang, M.H.; Kim, J.E.; Park, T.I.; Ha, J.H.; Lee, S.W.; Ahn, B.C.; Lee, J. Dual reporter gene imaging for tracking macrophage migration using the human sodium iodide symporter and an enhanced firefly luciferase in a murine inflammation model. Mol. Imaging Biol. 2013, 15, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Dosa, E.; Tuladhar, S.; Muldoon, L.L.; Hamilton, B.E.; Rooney, W.D.; Neuwelt, E.A. MRI using ferumoxytol improves the visualization of central nervous system vascular malformations. Stroke 2011, 42, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Dubey, P. Reporter gene imaging of immune responses to cancer: Progress and challenges. Theranostics 2012, 2, 355–362. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene/Protein | Contrast Mechanism | Observed Change | Test System | Ref. |
|---|---|---|---|---|
| Ferritin | Sequesters iron from labile intracellular iron pool and acts as an intracellular SPIO analog | T2 changes from 45 to 20 ms with 30 µg Ferritin expressed per mg of total protein (14 T) | Cell culture; viral-mediated transfection in mouse brain | [16] |
| Transferrin receptor (TfR) | Transferrin-conjugated SPIO particles are internalized by ectopically expressed TfR on transfected cells | 50% change in T2-weighted MRI signal (3 mg iron injected per mouse, 7.1 T) | Mice implanted with TfR-expressing gliosarcoma cells | [17] |
| Tyrosinase (TYR) | TYR produces melanin to chelate metal ions (Fe3+) | 37% increase in T1-weighted MRI signal (1.5 T) | Transfected mouse fibroblasts and HEK cells | [18] |
| β-galactosidase | Expressed β-gal cleaves a caged synthetic Gd3+ compounds | 60% T1-weighted signal increase (3.2 nmol per frog embryo, 12 T) | Xenopuslaevisembryos transfected withLacZ | [19] |
| Lysine rich-protein (LRP) | Chemical exchange saturation transfer (CEST) | 134% signal increase (6 days after 5 × 104 cells transplanted into the striatum of NOD-SCID male mice, 11.7 T) | Mice implanted with LRP and EGFP expressing xenografts respectively in opposite hemispheres | [20] |
| Molecular Imaging Methods | Resolution | Depth | Sensitivity | Cost | Potential Clinical Uses |
|---|---|---|---|---|---|
| OI | 1–5 mm | up to < 5 cm | 10−9–10−12 mol/L | low cost | very low |
| MRI | 10–100 µm | no limit | 10−3–10−5 mol/L | high cost | yes |
| PET | 1–2 mm | no limit | 10−10–10−12 mol/L | high cost | yes |
| SPECT | 0.3–1 mm | no limit | 10−10–10−12 mol/L | high cost | yes |
| Single/Fusion Reporter Gene | Gene/Protein | Contrast Mechanism | Imaging Method | Ref. |
|---|---|---|---|---|
| Single reporter gene | Oatp1 | (1) Transfer MRI contrast agents (2) Mediate uptake of Gd3+ or 111In based hepatotrophic contrast agents | MRI (T1WI) SPECT | [79,80] |
| LacZ | (1) Express β-gal to cleave Gd3+ compounds (2) Visualize lacZ gene expression with activated fluorescent contrast agents | MRI (T1WI) NIR | [4,5,56] | |
| TYR | (1) Broad optical absorption for photoacoustic effect (2) Chelate metal ions (Fe3+) providing contrast for MRI (3) Melanin-avid PET probes, 18F-P3BZA | PAI MRI (T2WI) PET | [52,81,82,83] | |
| Multimodality reporter gene | Single reporter gene and contrast agent | Sodium iodide symporter (NIS) gene and iron oxide | PET MRI | [84] |
| Fusion reporter gene | Ferritin gene fused with green fluorescent protein gene | MRI NIR | [74] |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Tian, R.; Liu, T.; Liu, G. MRI Reporter Genes for Noninvasive Molecular Imaging. Molecules 2016, 21, 580. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21050580
Yang C, Tian R, Liu T, Liu G. MRI Reporter Genes for Noninvasive Molecular Imaging. Molecules. 2016; 21(5):580. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21050580
Chicago/Turabian StyleYang, Caixia, Rui Tian, Ting Liu, and Gang Liu. 2016. "MRI Reporter Genes for Noninvasive Molecular Imaging" Molecules 21, no. 5: 580. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21050580