
The Role of fadD19 and echA19 in Sterol Side Chain Degradation by Mycobacterium smegmatis

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. M. smegmatis fadD19 and echA19 Genes are not Essential for Viability
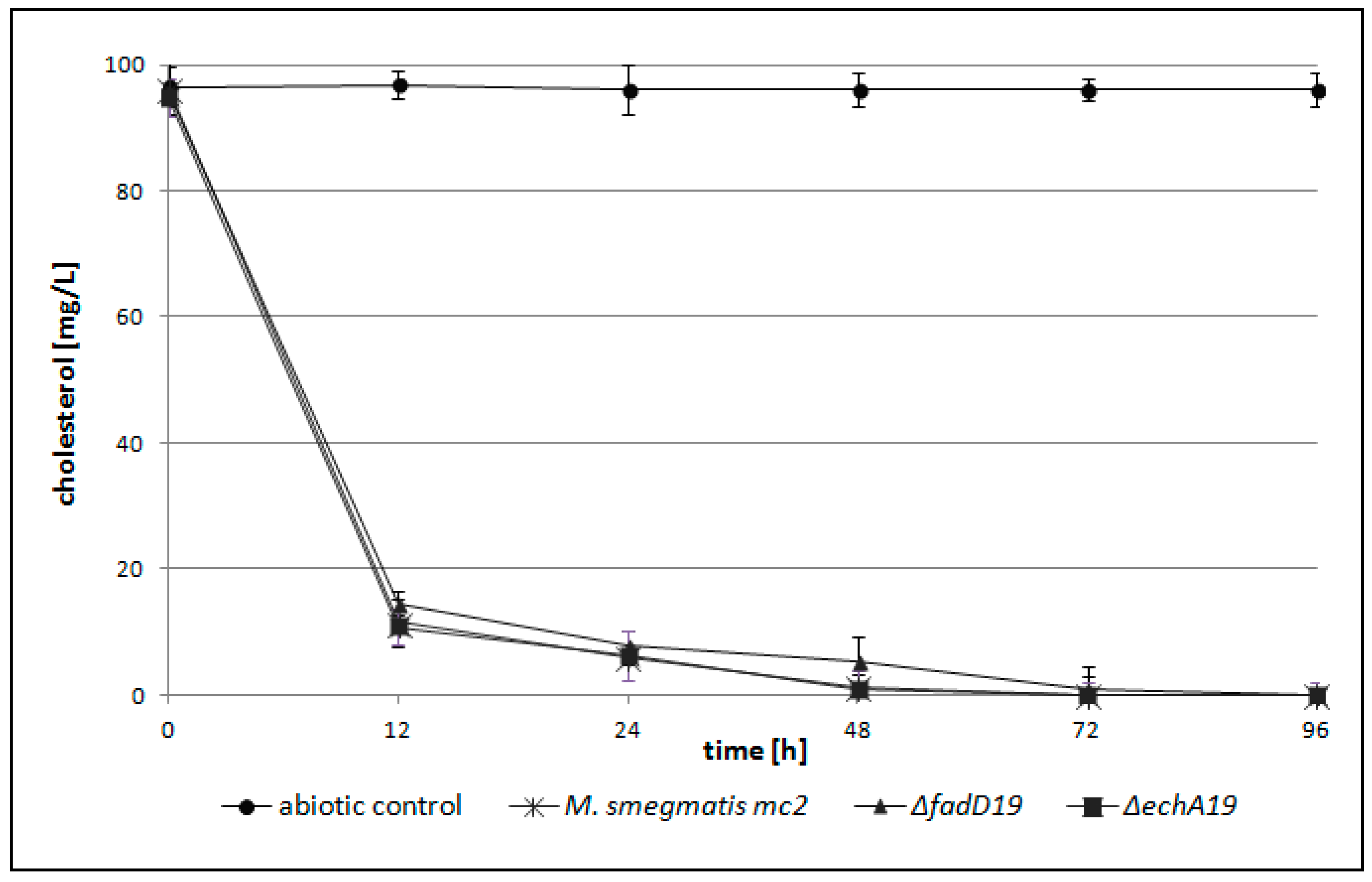
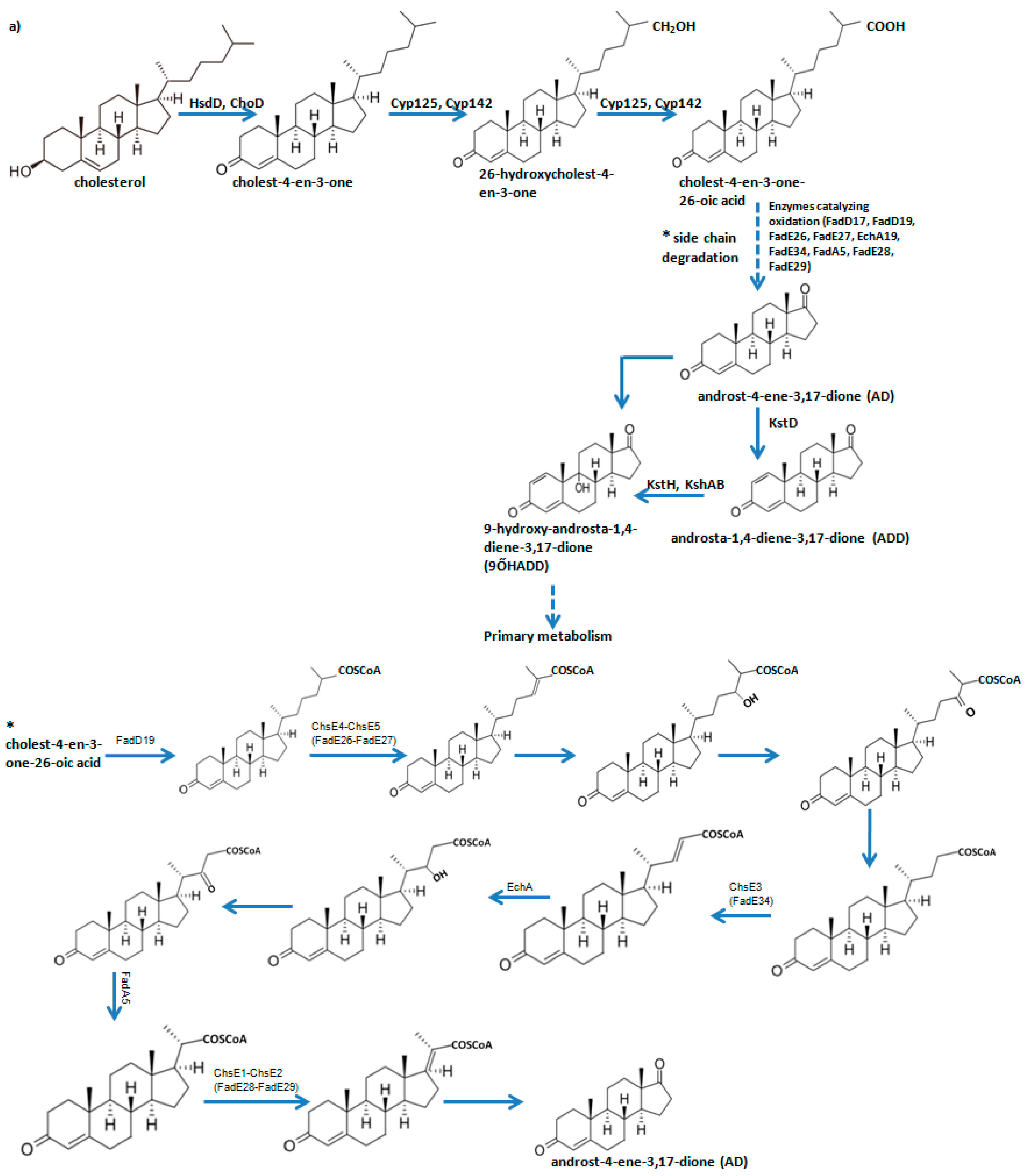
2.2. M. smegmatis fadD19 and echA19 Genes are not Essential for Cholesterol (Side Chain) Degradation
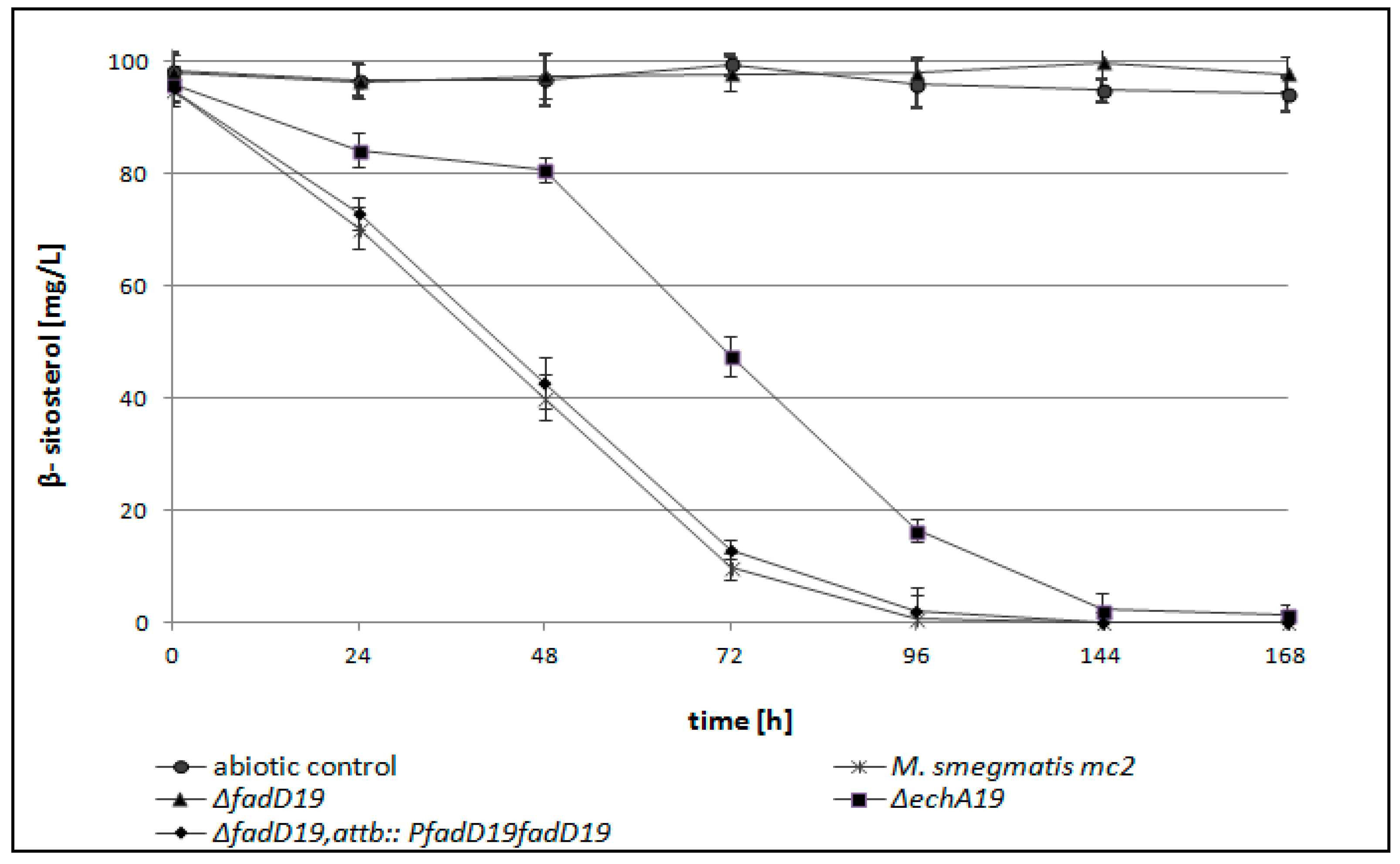
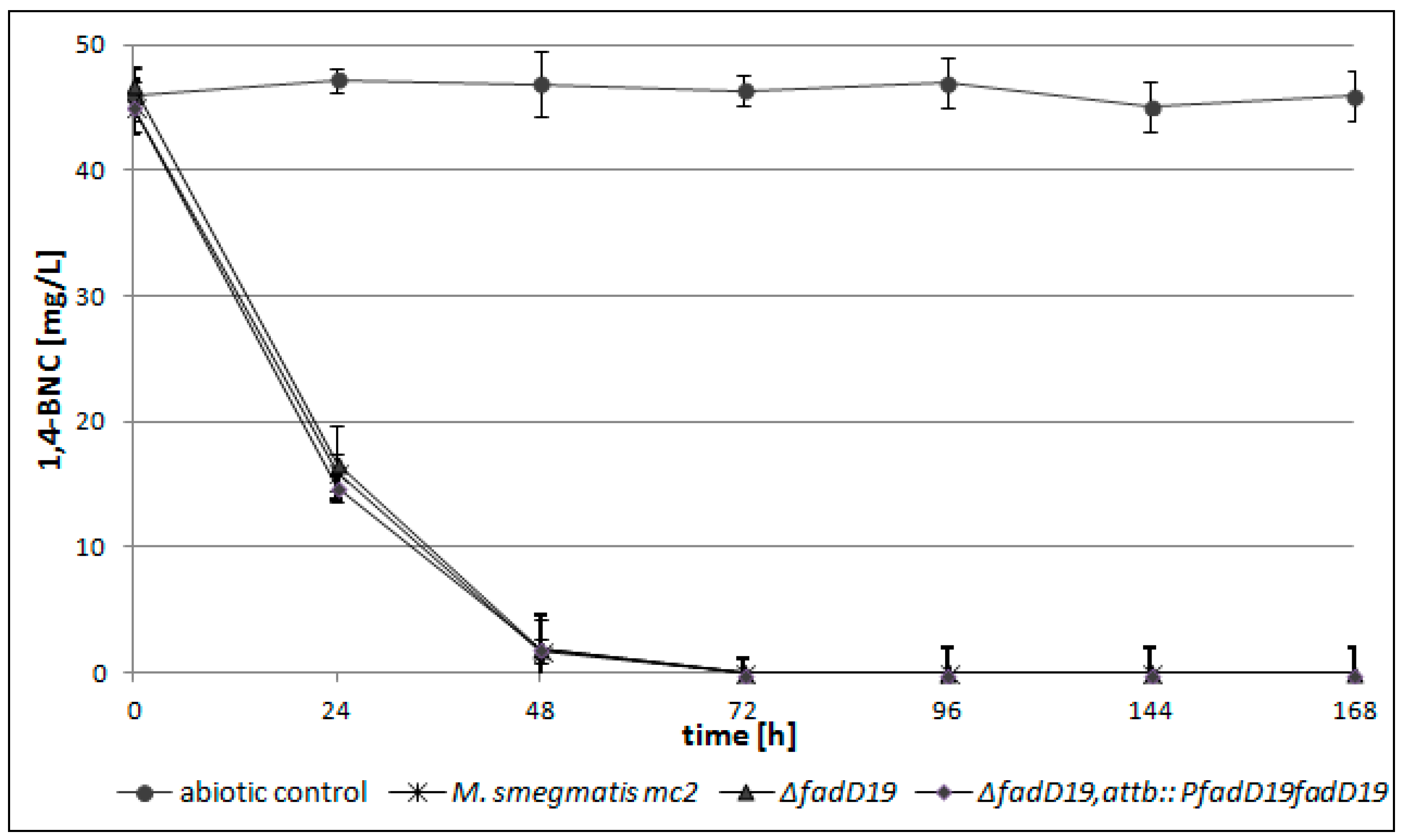
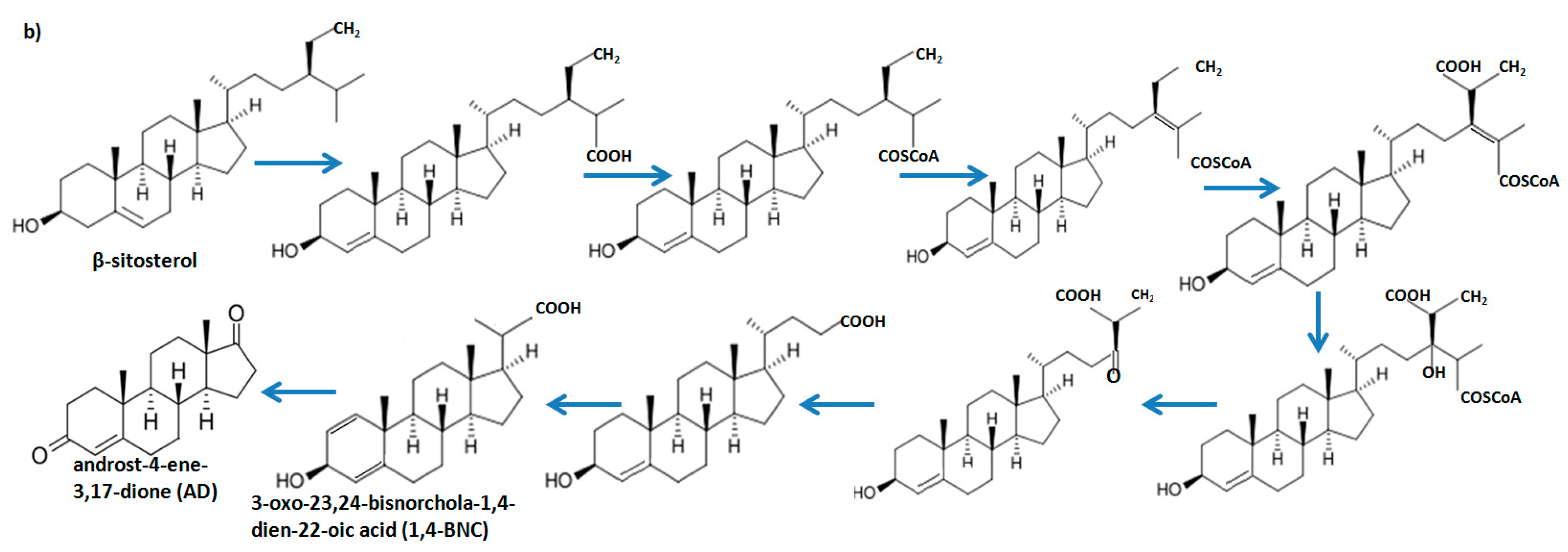
2.3. fadD19 but not echA19 is Required for β-Sitosterol (Side Chain) Degradation
3. Discussion
4. Experimental Section
4.1. Bacterial Strains and Culture Conditions
4.2. Plasmid Constructions
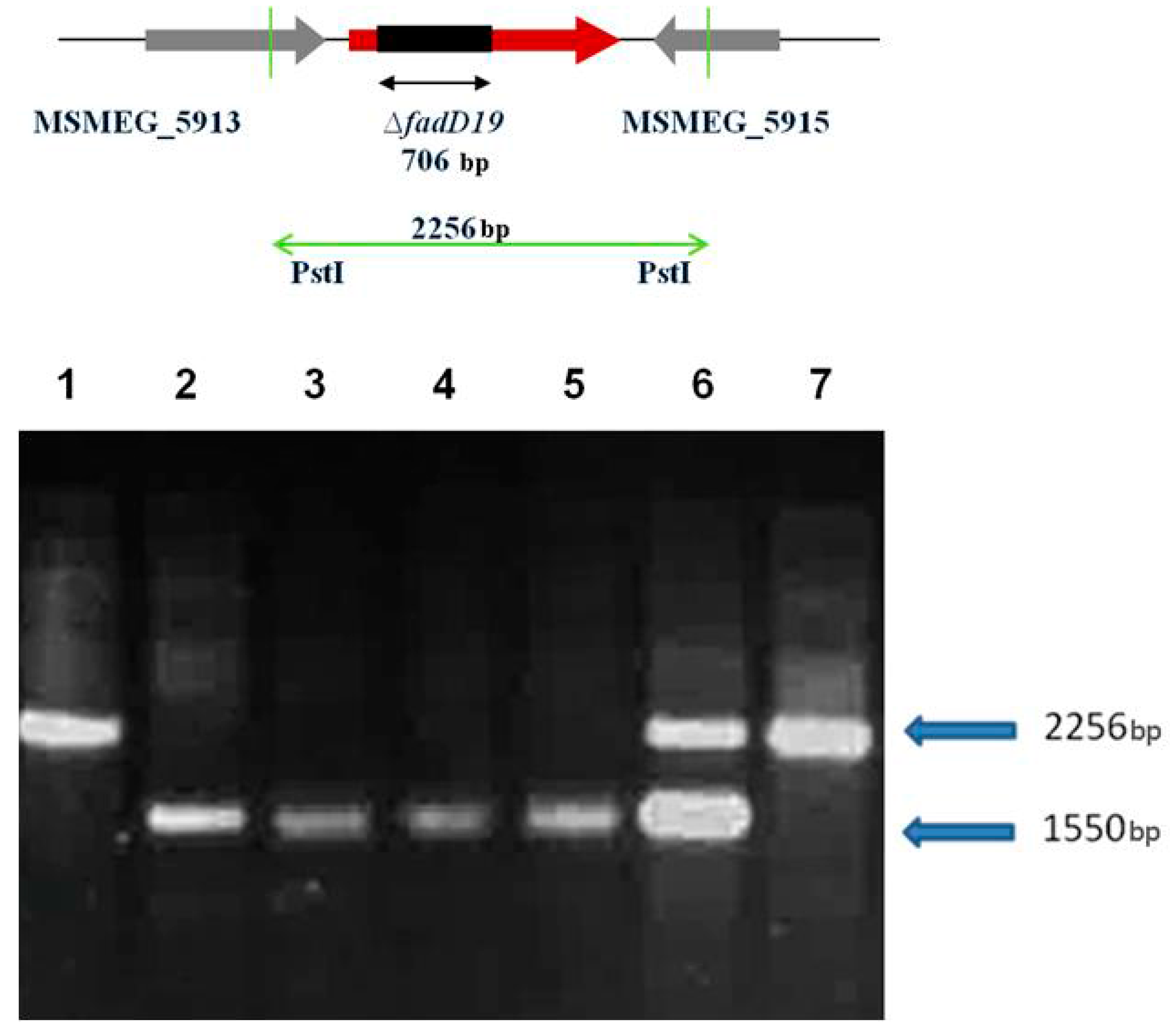
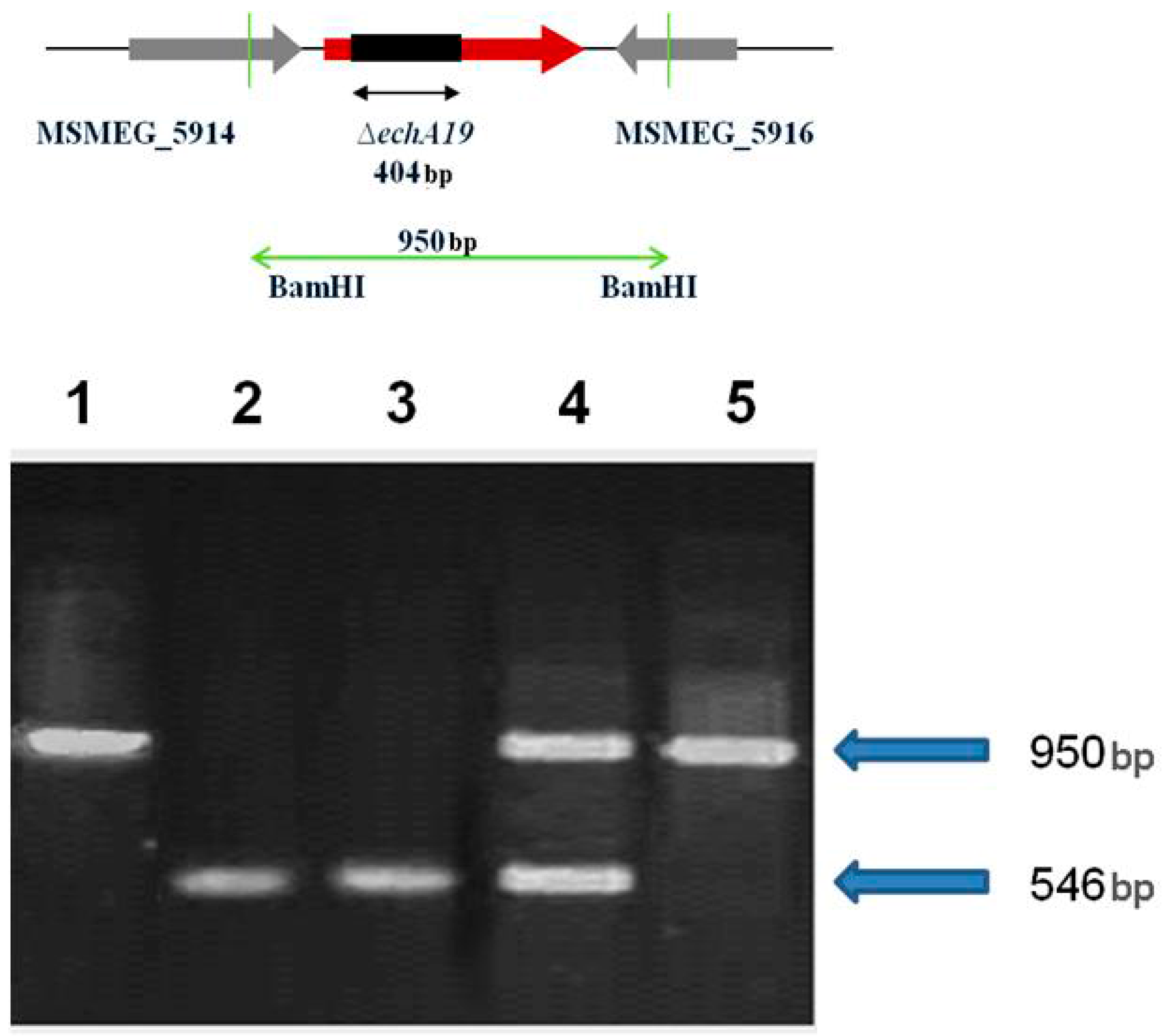
4.3. Disruption of fadD19 and echA19 Genes
4.4. Complementation Constructs
4.5. Sterols Degradation
4.5.1. Steroid Standards
4.5.2. Preparation of Cholesterol
4.5.3. Preparation of β-Sitosterol
4.5.4. HPLC MS/MS Analysis of Sterol Degradation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Naghibi, F.; Tabatabai Yazdi, M.; Sahebgharani, M.; Noori Daloii, M.R. Microbial transformation of cholesterol. J. Sci. Islam. Repub. Iran 2002, 13, 103–106. [Google Scholar]
- Donova, M.V.; Egorova, O.V. Microbial steroid transformations: Current state and prospects. Appl. Microbiol. Biotechnol. 2012, 94, 1423–1447. [Google Scholar] [CrossRef] [PubMed]
- Lad, N. Optimalization of parameters for hydrocortisone succinate bioconversion. Malays. J. Microbiol. 2011, 7, 7–13. [Google Scholar]
- Field, J.A.; Bolesma, F.; Baten, H.; Rulkens, W.H. Oxidation of anthracene in water solvent mixtures by the white-rot fungus Bjerkandera sp. strain BOS55. Appl. Microbiol. Biotechnol. 1995, 44, 234–240. [Google Scholar] [CrossRef]
- Cerniglia, C.E. Biodegradation of polycyclic aromatic hydrocarbons. Biodegradation 1992, 3, 351–368. [Google Scholar] [CrossRef]
- Guerin, W.F.; Jones, G.E. Two-stage mineralization of phenanthrene by estuarine enrichment cultures. Appl. Environ. Microbiol. 1998, 54, 937–944. [Google Scholar]
- Mattos, K.A.; Oliveira, V.C.G.; Berrêdo-Pinho, M.; Amaral, J.J.; Antunes, L.C.M.; Melo, R.C.N.; Acosta, C.C.D.; Moura, D.F.; Olmo, R.; Han, J.; et al. Mycobacterium leprae intracellular survival relies on cholesterol accumulation in infected macrophages: A potential target for new drugs for leprosy treatment. Cell. Microbiol. 2014, 16, 797–815. [Google Scholar] [CrossRef] [PubMed]
- Fine-Coulson, K.; Reaves, B.J.; Karls, R.K.; Quinn, F.D. The role of lipid raft aggregation in the infection of type II pneumocytes by Mycobacterium tuberculosis. PLoS ONE 2012, 7, e45028. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Sassetti, C.M. Mycobacterial persistence requires the utilization of host cholesterol. Proc. Natl. Acad. Sci. USA 2008, 105, 4376–4380. [Google Scholar] [CrossRef] [PubMed]
- Brzostek, A.; Pawełczyk, J.; Rumijowska-Galewicz, A.; Dziadek, B.; Dziadek, J. Mycobacterium tuberculosis is able to accumulate and utilize cholesterol. J. Bacteriol. 2009, 191, 6584–6591. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.C.; Miner, M.D.; Pandey, A.K.; Gill, W.P.; Harik, N.S.; Sassetti, C.M.; Sherman, D.R. Igr. gene and Mycobacterium tuberculosis cholesterol metabolism. J. Bacteriol. 2009, 191, 5232–5239. [Google Scholar] [CrossRef] [PubMed]
- Yam, K.C.; D’Angelo, I.; Kalscheuer, R.; Zhu, H.; Wang, J.X. Studies of a ring-cleaving dioxygenase illuminate the role of cholesterol metabolism in the pathogenesis of Mycobacterium tuberculosis. PLoS Pathog. 2009, 5, e1000344. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; van der Geize, R.; Besra, G.S.; Gurcha, S.S.; Liu, A. 3-Ketosteroid 9alpha-hydroxylase is an essential factor in the pathogenesis of Mycobacterium tuberculosis. Mol. Microbiol. 2010, 75, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Ouellet, H.; Johnston, J.B.; de Montellano, P.R. Cholesterol catabolism as a therapeutic target in Mycobacterium tuberculosis. Trends Microbiol. 2011, 19, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Brzezinska, M.; Szulc, I.; Brzostek, A.; Klink, M.; Kiełbik, M. The role of 3-ketosteroid 1(2)-dehydrogenase in the pathogenicity of Mycobacterium tuberculosis. BMC Microbiol. 2013, 13, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Van der Ven, B.C.; Fahey, R.J.; Lee, W.; Liu, Y.; Abramovitch, R.B.; Memmott, C.; Crowe, A.M.; Eltis, L.D.; Perola, E.; Deininger, D.D.; et al. Novel inhibitors of cholesterol degradation in Mycobacterium tuberculosis reveal how the bacterium’s metabolism is constrained by the intracellular environment. PLoS Pathog. 2015, 11, e1004679. [Google Scholar]
- Griffin, J.E.; Gawronski, J.D.; Dejesus, M.A.; Ioerger, T.R.; Akerley, B.J.; Sassetti, C.M. High-resolution phenotypic profiling defines genes essential for mycobacterial growth and cholesterol catabolism. PLoS Pathog. 2011, 7, e1002251. [Google Scholar] [CrossRef] [PubMed]
- Uhia, I.; Galan, B.; Morales, V.; Garcia, J.L. Initial step in the catabolism of cholesterol by Mycobacterium smegmatis mc2155. Environ. Microbiol. 2011, 13, 943–959. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Dubnau, E.; Smith, I.; Sampson, N.S. Rv1106c from Mycobacterium tuberculosis is a 3β-hydroxysteroid dehydrogenase. Biochemistry 2007, 46, 9058–9067. [Google Scholar] [CrossRef] [PubMed]
- Brzostek, A.; Rumijowska-Galewicz, A.; Dziadek, B.; Wójcik, E.A.; Dziadek, J. ChoD and HsdD can be dispensable for cholesterol degradation in mycobacteria. J. Steroid Biochem. Mol. Biol. 2013, 134, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Gao, J.; Smith, I.; Dubnau, E.; Sampson, N.S. Cholesterol is not essential source of nutrition for Mycobacterium tuberculosis during infection. J. Bacteriol. 2011, 193, 1473–1476. [Google Scholar] [CrossRef] [PubMed]
- Van der Geize, R.; Yam, K.; Heuser, T.; Wilbrink, M.H.; Hara, H.; Anderton, M.C.; Sim, E.; Dijkhuizen, L.; Davies, J.E.; Mohn, W.W.; et al. A gene cluster encoding cholesterol catabolism in a soil actinomycete provides insight into Mycobacterium tuberculosis survival in macrophages. Proc. Natl. Acad. Sci. USA 2007, 104, 1947–1952. [Google Scholar] [CrossRef] [PubMed]
- Nesbitt, N.M.; Yang, X.; Fontan, P.; Kolesnikova, I.; Smith, I.; Sampson, N.S.; Dubnau, E. A thiolase of Mycobacterium tuberculosis is required for virulence and production of androstenedione and adrostadienedione from cholesterol. Infect. Immun. 2010, 78, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Wilbrink, M.H.; van der Geize, R.; Dijkhuizen, L. Molecular characterization of ltp3 and ltp4, essential for C24-branched chain sterol-side-chain degradation in Rhodoccocus rhodochrous DSM 43269. Microbiology 2012, 158, 3054–3062. [Google Scholar] [CrossRef] [PubMed]
- Uhia, I.; Galan, B.; Kendall, S.L.; Stoker, N.G.; Garcia, J. Cholesterol metabolism in Mycobacterium smegmatis. Environ. Microbiol. Rep. 2012, 4, 168–182. [Google Scholar] [CrossRef] [PubMed]
- Wilbrink, M.H.; Petrusma, M.; Dijkhuizen, L.; van der Geize, R. FadD19 of Rhodococcus rhodochorus DSM43269, a s-coenzyme A ligase essential for degradation of C24 branched sterol side chains. Appl. Environ. Microbiol. 2011, 77, 4455–4464. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.J.; Madrona, Y.; Ortiz de Montellano, P.R. Cholesterol ester oxidation by Mycobacterial Cytochrome P450. J. Biol. Chem. 2014, 289, 30417–30425. [Google Scholar] [CrossRef] [PubMed]
- Capyk, J.K.; Kalscheuer, R.; Stewart, G.R.; Liu, J.; Kwon, H.; Zhao, R.; Okamoto, S.; Jacobs, W.R.; Eltis, L.D.; Mohn, W.W. Enzyme Catalysis and Regulation: Mycobacterial Cytochrome P450 125 (Cyp125) Catalyzes the Terminal Hydroxylation of C27 Steroids. J. Biol. Chem. 2009, 284, 35534–35542. [Google Scholar] [CrossRef] [PubMed]
- McLean, K.J.; Lafite, P.; Levy, C.; Cheesman, R.E.; Mast, N.; Pikuleva, I.A.; Leys, D.; Munro, A.W. The structure of Mycobacterium tuberculosis CYP125: Molecular basis for cholesterol binding in a P450 needed for host infection. J. Biol. Chem. 2009, 284, 35524–35533. [Google Scholar] [CrossRef] [PubMed]
- Johnston, J.B.; Ouellet, H.; Ortiz de Montellano, P.R. Functional redundancy of steroid C26-monooxygenase activity in Mycobacterium tuberculosis revealed by biochemical and genetic analyses. J. Biol. Chem. 2010, 285, 36352–36360. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, M.D.; McLean, K.J.; Levy, C.; Mast, N.; Pikuleva, I.A.; Lafite, P.; Rigby, S.E.J.; Leys, D.; Munro, A.W. Structural and biochemical characterization of Mycobacterium tuberculosis CYP142: Evidence for multiple cholesterol 27-hydroxylase activities in a human pathogen. J. Biol. Chem. 2010, 285, 38270–38282. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.T.; Sampson, N.S. Mycobacterium tuberculosis utilizes a unique heterotetrameric structure for dehydrogenation of the cholesterol side chain. Biochemistry 2013, 52, 2895–2904. [Google Scholar] [CrossRef] [PubMed]
- Ruprecht, A.; Maddox, J.; Stirling, A.J.; Visaggio, N.; Seah, S.Y.K. Characterization of novel acyl Coenzyme A dehydrogenases involved in bacterial steroid degradation. J. Bacteriol. 2015, 197, 1360–1367. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Lu, R.; Guja, K.E.; Wipperman, M.F.; Clair, J.R.St.; Bonds, A.C.; Garcia-Diaz, M.; Sampson, N. Unraveling cholesterol catabolism in Mycobacterium tuberculosis: ChsE4-ChsE5 α2β2 acyl-CoA dehydrogenase initiates β-oxidation of 3-oxo-cholest-4-en-26-oyl CoA. ACS Infect. Dis. 2015, 1, 110–125. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Schmitz, W.; Sampson, N.S. α-methyl acyl CoA racemase provides Mycobacterium tuberculosis catabolic access to cholesterol esters. Biochemistry 2015, 54, 5669–5672. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.T.; VanderVen, B.C.; Sherman, D.R.; Russell, D.G.; Sampson, N.S. Pathway profiling in Mycobacterium tuberculosis: Elucidation of cholesterol-derived catabolite and enzymes that catalyze its metabolism. J. Biol. Chem. 2011, 286, 43668–43678. [Google Scholar] [CrossRef] [PubMed]
- Parish, T.; Stocker, N.G. Use of a flexible cassette method to generate a double unmarked Mycobacterium tuberculosis tlyA plcABC mutant by gene replacement. Microbiology 2000, 146, 1969–1975. [Google Scholar] [CrossRef] [PubMed]
- Wipperman, M.F.; Sampson, N.S.; Thomas, S.T. Pathogen roid rage: Cholesterol utilization by Mycobacterium tuberculosis. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 269–293. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.J.; Boshoff, H.I.; Krishnan, N.; Gonzales, J.; Schnappinger, D.; Robertson, B.D. The Mycobacterium tuberculosis β-oxidation genes echA5 and fadB3 are dispensable for growth in vitro and in vivo. Tuberculosis 2011, 91, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Guja, K.E.; Thomas, S.T.; Garcia-Diaz, M.; Sampson, N. A distinct MaoC-like enoyl-CoA hydratase architecture mediates cholesterol catabolism in Mycobacterium tuberculosis. ACS Chem. Biol. 2014, 9, 2632–2645. [Google Scholar] [CrossRef] [PubMed]
- Casabon, I.; Swain, K.; Crowe, A.M.; Eltis, L.D.; Mohn, W.W. Actinobacterial acyl-CoA synthetases involved in steroid side chain catabolism. J. Bacteriol. 2014, 196, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Rosłoniec, K.Z.; Wilbrink, M.H.; Capyk, J.K.; Mohn, W.W.; Ostendorf, M.; van der Geize, R.; Dijkhuizen, L.; Eltis, L.D. Cytochrome P450 125 (CYP125) catalyses C26-hydroxylation to initiate sterol side-chain degradation in Rhodococcus jostii RHA1. Mol. Microbiol. 2009, 74, 1031–1043. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.L.; Uhia, I.; Galan, B. Catabolism and biotechnological applications of cholesterol degrading bacteria. Microb. Biotechnol. 2012, 5, 679–699. [Google Scholar] [CrossRef] [PubMed]
- Snapper, S.B.; Melton, R.E.; Mustafa, S.; Kieser, T.; Jacobs, W.R., Jr. Isolation and characterization of efficient plasmid transformation mutants of Mycobacterium smegmatis. Mol. Microbiol. 1990, 4, 1911–1919. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (h) | Cholestenone | AD | ADD | Cholestenone | AD | ADD | Cholestenone | AD | ADD |
|---|---|---|---|---|---|---|---|---|---|
| Mycobacterium smegmatis mc2155 | ΔfadD19 | ΔechA19 | |||||||
| 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 12 | 11.19 | 0 | 0 | 11.02 | 0 | 0 | 10.01 | 0 | 0 |
| 24 | 10.31 | 1.95 | 0.83 | 14.13 | 2.25 | 1.05 | 11.05 | 0.75 | 0.9 |
| 48 | 0 | 4.51 | 6.2 | 0.74 | 3.94 | 6.98 | 1.05 | 3.02 | 6.14 |
| 72 | 0 | 4.06 | 9.4 | 0 | 5.11 | 7.5 | 0 | 4.97 | 7.5 |
| 96 | 0 | 4.04 | 8.5 | 0 | 6 | 8.71 | 0 | 7.34 | 9 |
| 144 | 0 | 2.1 | 6.1 | 0 | 2.07 | 5.09 | 0 | 1.79 | 6.01 |
| Time (h) | 1,4-BNC | ADD | 1,4-BNC | ADD | 1,4-BNC | ADD |
|---|---|---|---|---|---|---|
| Mycobacterium smegmatis mc2155 | ΔfadD19 | ΔechA19 | ||||
| 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 24 | 10.24 | 0.51 | 0 | 0 | 13.05 | 0 |
| 72 | 17.81 | 3.57 | 0 | 0 | 29.1 | 2.1 |
| 144 | 2.34 | 8.75 | 0 | 0 | 4.75 | 7.05 |
| 168 | 0 | 3.01 | 0 | 0 | 0.21 | 8.95 |
| PLASMID | DESCRIPTION | SOURCE |
|---|---|---|
| CLONING VECTORS | ||
| p2NIL | Recombination vector, non-replicating in mycobacteria, KmR | Parish and Stocker [ 29] |
| pGOAL17 | Source of PacI cassette, AmpR | Parish and Stocker [ 29] |
| pMV306 | Mycobacterial integrating vector, KmR | Med-Imune Inc. |
| VECTORS USED FOR THE GENE REPLACEMENT | ||
| pNW03 | p2NIL carrying a fragment of the 5 ′fadD19 gene and the DNA located upstream of the gene (fadD19Gr1-2) restricted recognition sequences for Kpn I and BamH1 | This study |
| pNW04 | pNW03 carrying a fragment of the 3′fadD19 gene and the DNA located below of the gene (fadD19Gr3-4), bounded by recognition sequences BamH1 and HindIII | This study |
| pNW05 | pNW04 with PacI cassette from pGOAL17 | This study |
| pNW08 | p2NIL carrying a fragment of the 5 ′echA19 gene and the DNA located upstream of the gene (echA19Gr1-2) restricted recognition sequences for Pst I and HindIII | This study |
| pNW09 | pNW08 carrying a fragment of the 3′echA19 gene and the DNA located below of the gene (echA19Gr3-4), bounded by recognition sequences KpnI and HindIII | This study |
| pNW10 | pNW09 with PacI cassette from pGOAL17 | This study |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wrońska, N.; Brzostek, A.; Szewczyk, R.; Soboń, A.; Dziadek, J.; Lisowska, K. The Role of fadD19 and echA19 in Sterol Side Chain Degradation by Mycobacterium smegmatis. Molecules 2016, 21, 598. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21050598
Wrońska N, Brzostek A, Szewczyk R, Soboń A, Dziadek J, Lisowska K. The Role of fadD19 and echA19 in Sterol Side Chain Degradation by Mycobacterium smegmatis. Molecules. 2016; 21(5):598. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21050598
Chicago/Turabian StyleWrońska, Natalia, Anna Brzostek, Rafał Szewczyk, Adrian Soboń, Jarosław Dziadek, and Katarzyna Lisowska. 2016. "The Role of fadD19 and echA19 in Sterol Side Chain Degradation by Mycobacterium smegmatis" Molecules 21, no. 5: 598. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21050598