
Identification of New Epididymal Luminal Fluid Proteins Involved in Sperm Maturation in Infertile Rats Treated by Dutasteride Using iTRAQ

Abstract
:1. Introduction
2. Results
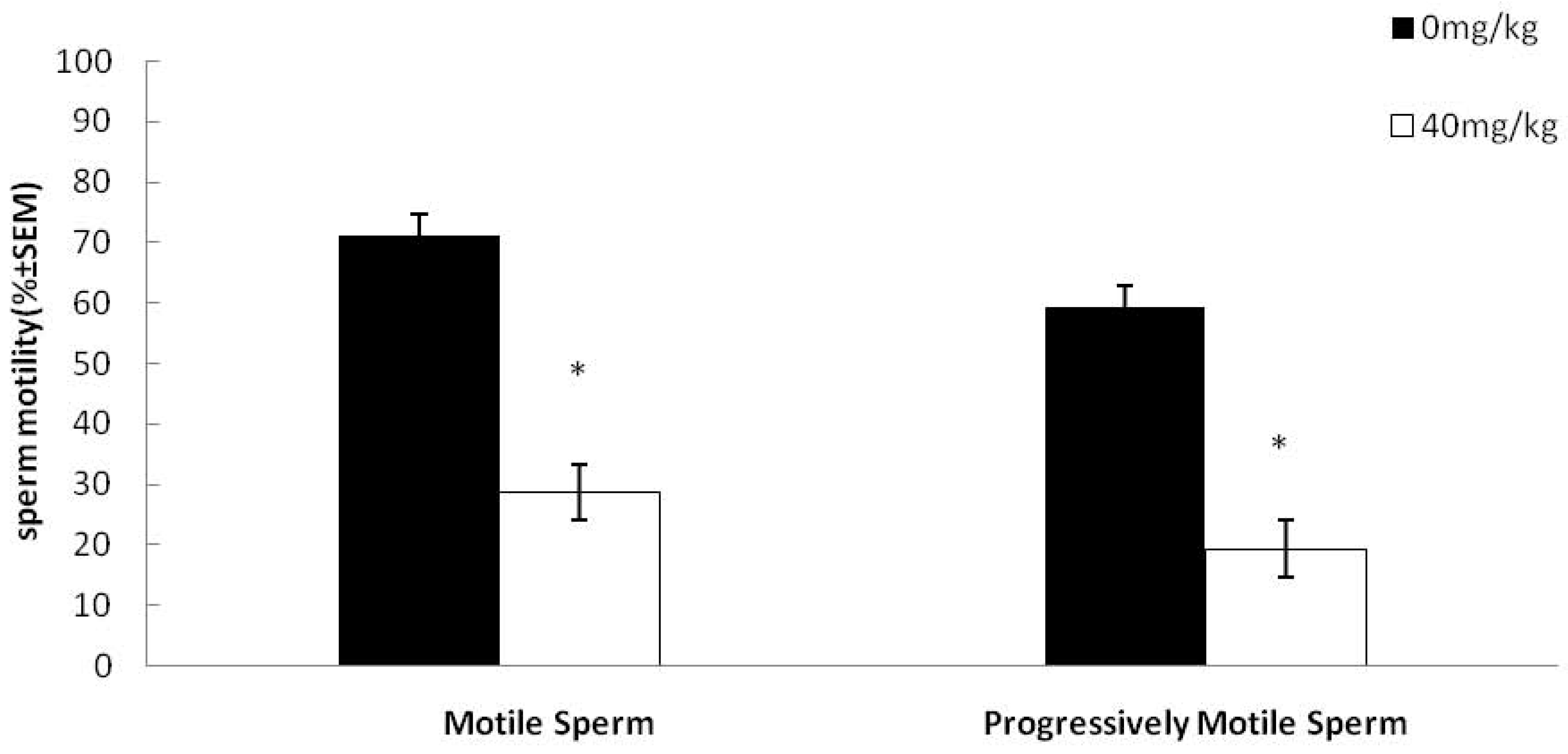
2.1. Effects of Dutasteride Treatment on Sperm Motility
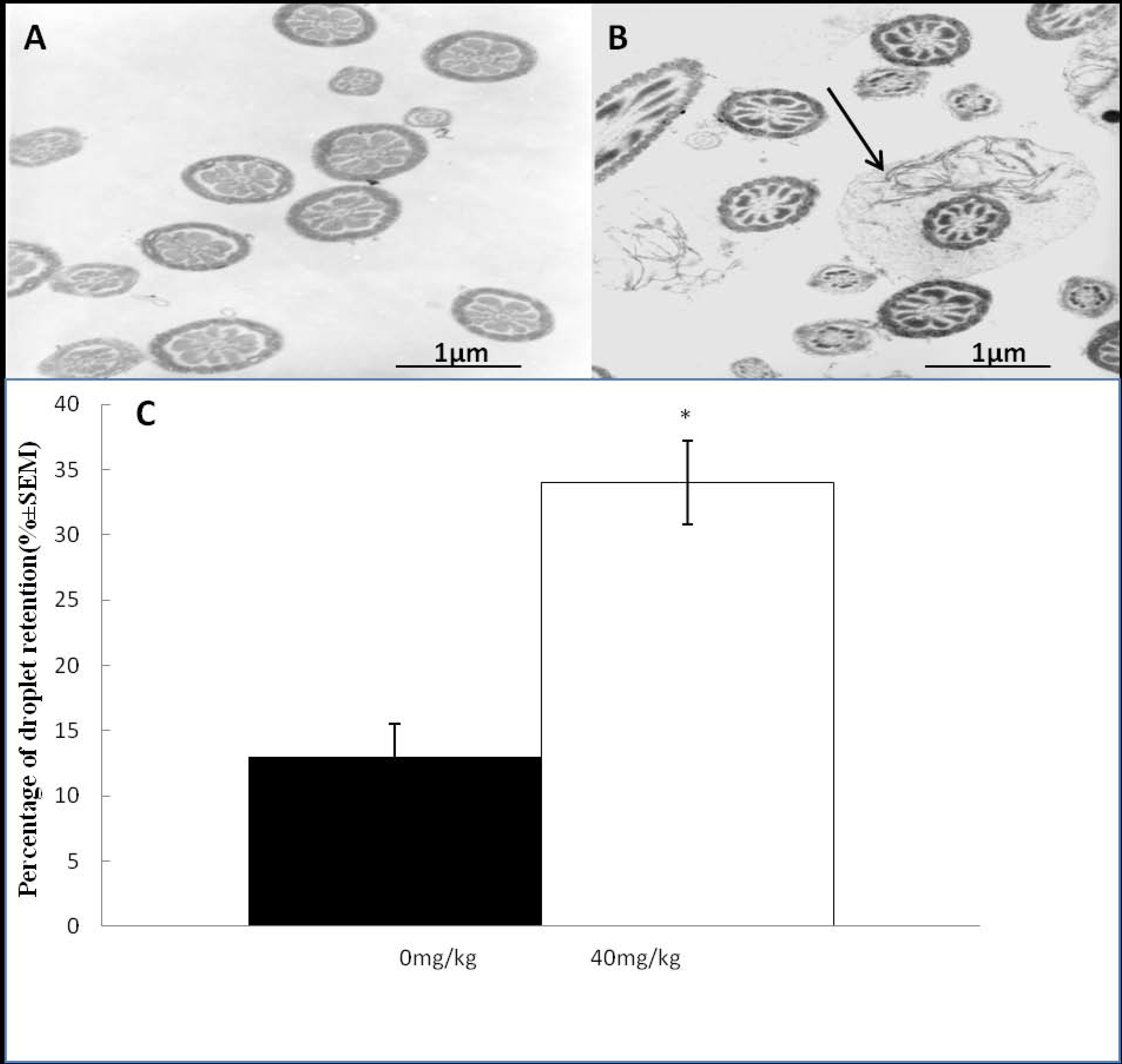
2.2. Effects of Dutasteride Treatment on Sperm Morphology
2.3. Effects of Dutasteride Treatment on Male Mating and Fertility
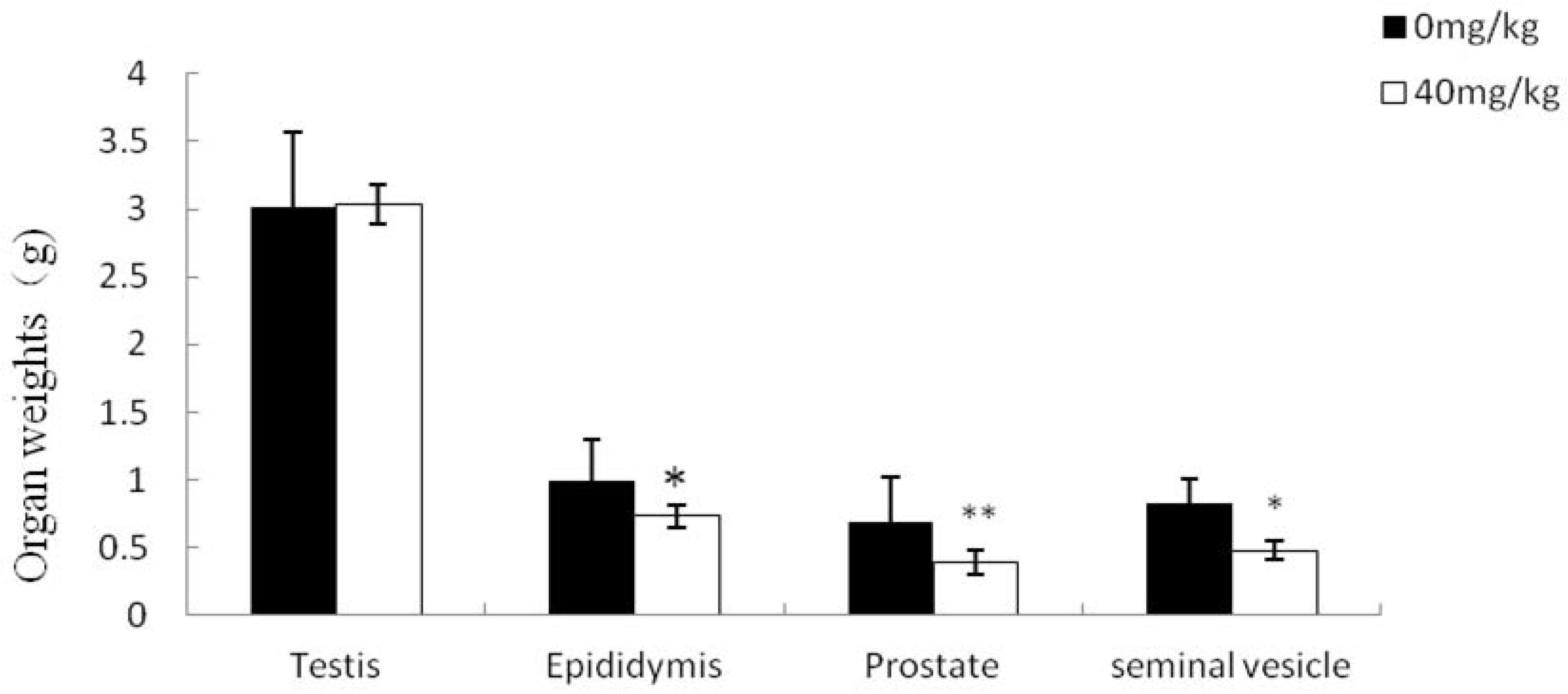
2.4. Effects of Dutasteride Treatment on Reproductive Organ Weights
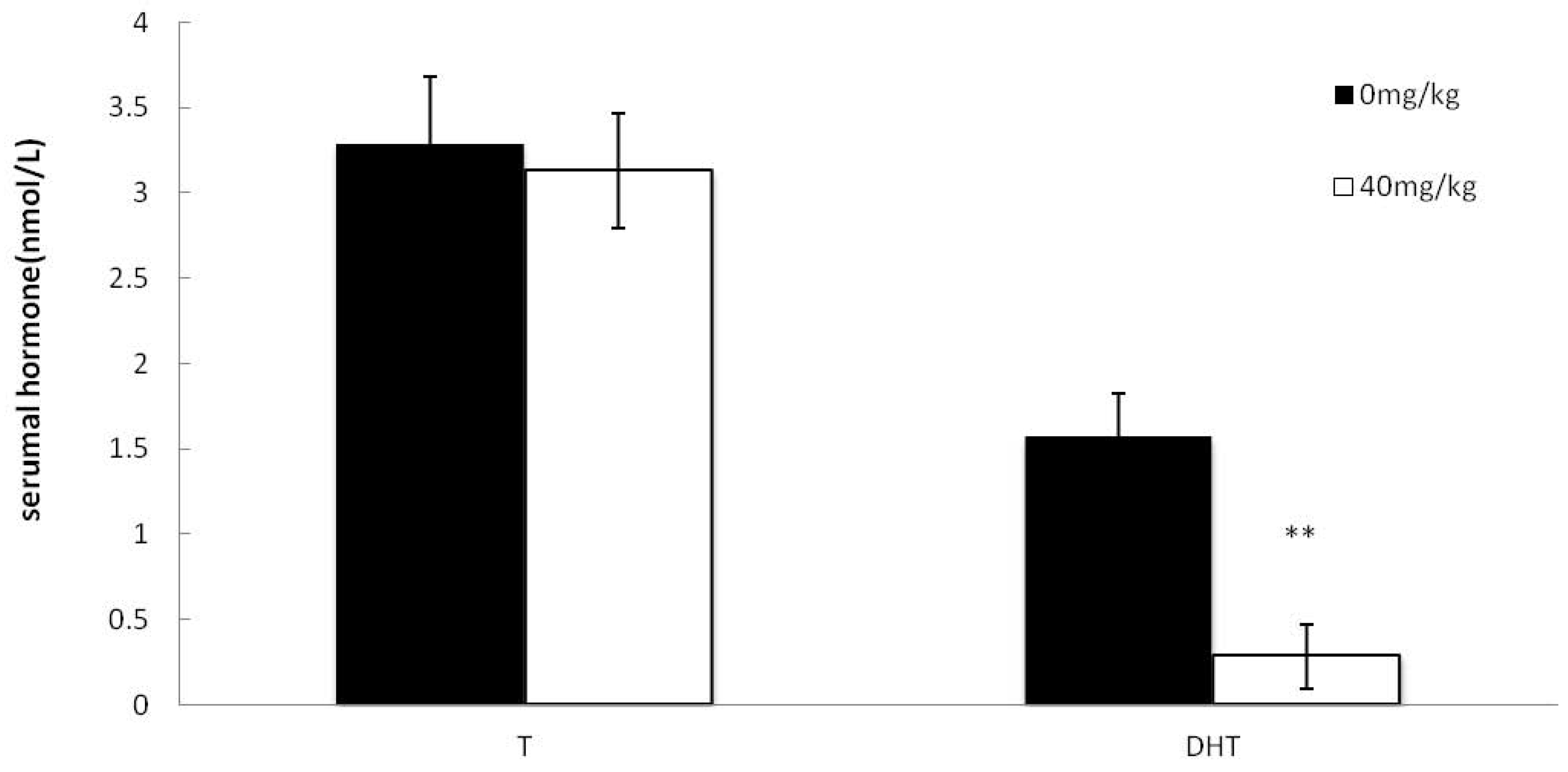
2.5. Effects of Dutasteride Treatment on Serumal T and DHT
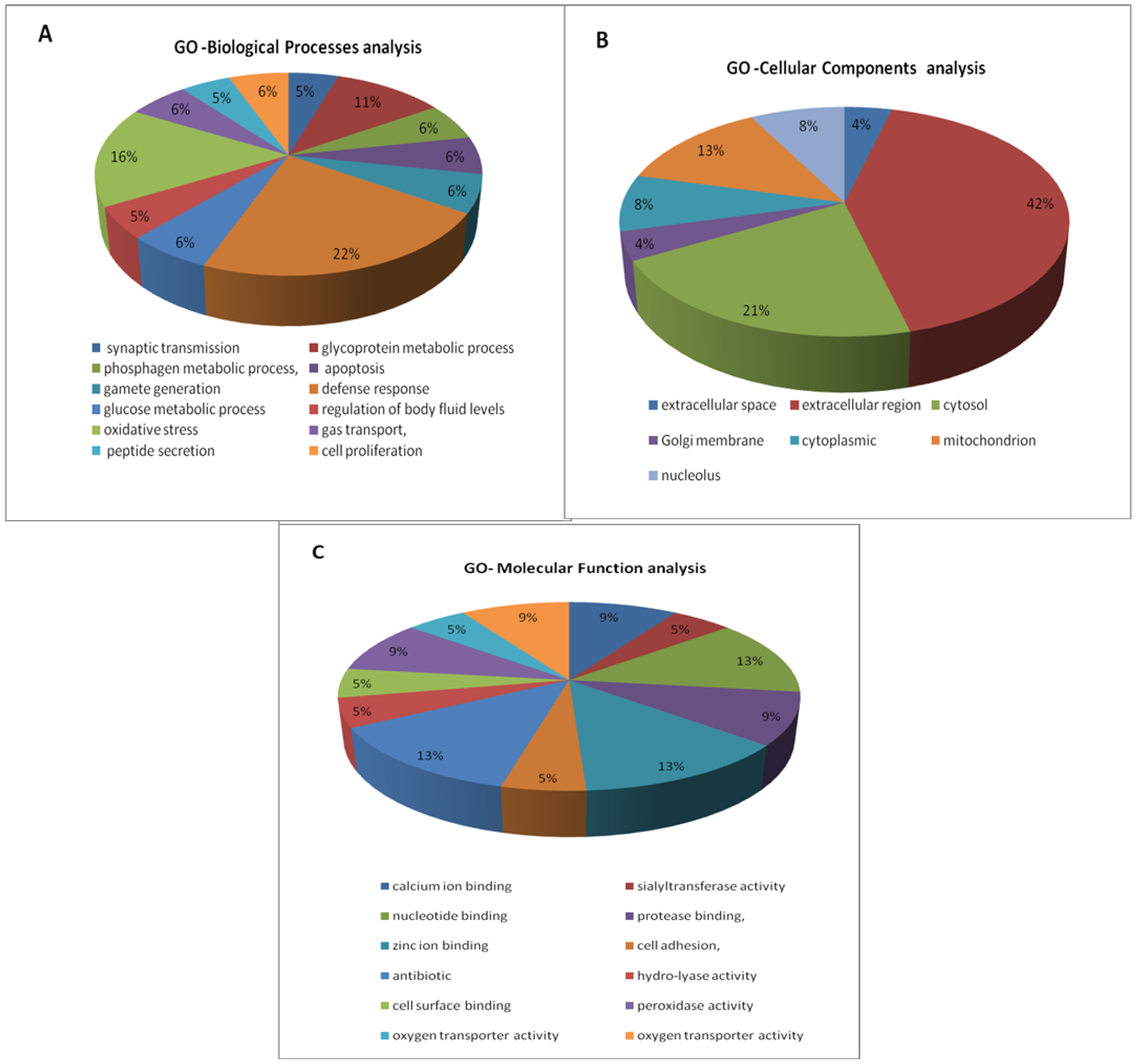
2.6. Differentially Expressed Proteins in Epididymal Luminal Fluids
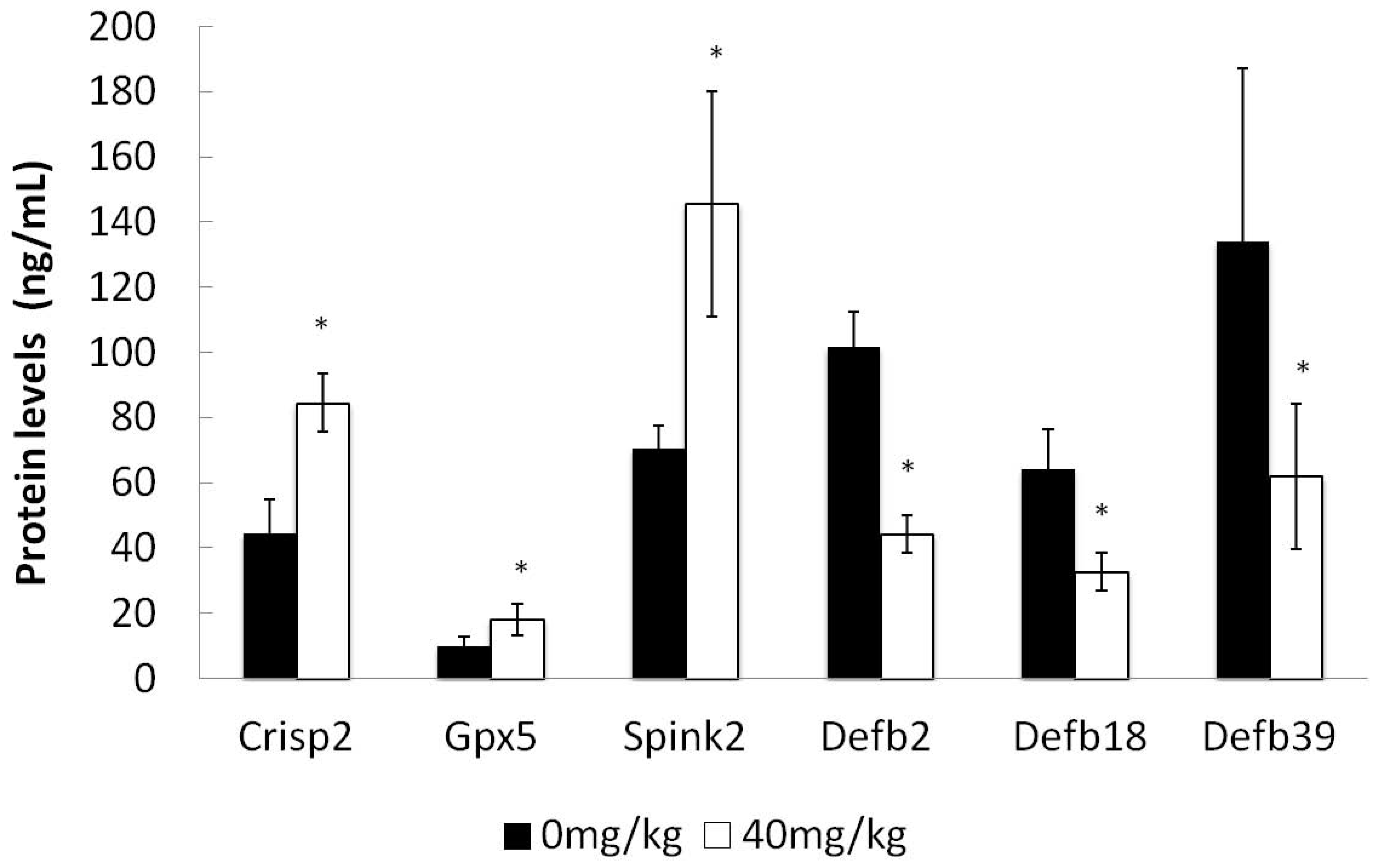
2.7. ELISA Detection of Selected Protein Concentrations
3. Discussion
4. Materials and Methods
4.1. Animals and Treatment Procedure
4.2. Epididymal Luminal Fluids Collection
4.3. Peptide Quantitation and iTRAQ Label
4.4. Peptide Fractionation with Strong Cation Exchange (SCX) Chromatography
4.5. LC-ESI -MS/MS Analysis by Q Exactive
4.6. Sequence Database Searching and Quantitative Analysis
4.7. Validation by Enzyme-Linked Immunosorbent Assay
4.8. Data Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| iTRAQ | isobaric tags for relative and absolute quantitation |
| DHT | dihydrotestosterone |
| T | testosterone |
| CASA | computer-assisted sperm analysis |
References
- Orgebin-Crist, M.C. Sperm maturation in rabbit epididymis. Nature 1967, 216, 816–818. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Han, C.; Jin, S.; Kwon, J.T.; Kim, J.; Jeong, J.; Kim, J.; Ham, S.; Jeon, S.; Yoo, Y.J.; et al. Reduced fertility and altered epididymal and sperm integrity in mice lacking ADAM7. Biol. Reprod. 2015, 93. [Google Scholar] [CrossRef] [PubMed]
- Gawecka, J.E.; Boaz, S.; Kasperson, K.; Nguyen, H.; Evenson, D.P.; Ward, W.S. Luminal fluid of epididymis and vas deferens contributes to sperm chromatin fragmentation. Hum. Reprod. 2015, 30, 2725–2736. [Google Scholar] [CrossRef] [PubMed]
- Koch, S.; Acebron, S.P.; Herbst, J.; Hatiboglu, G.; Niehrs, C. Post-transcriptional Wnt Signaling Governs Epididymal Sperm Maturation. Cell 2015, 163, 1225–1236. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.X.; Zhang, Y.L.; Xiao, L.; Zheng, M.; Leung, K.M.; Chan, M.Y.; Lo, P.S.; Tsang, L.L.; Wong, H.Y.; Ho, L.S.; et al. An epididymis-specific β-defensin is important for the initiation of sperm maturation. Nat. Cell Biol. 2004, 6, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Mandon, M.; Cyr, D.G. Tricellulin and its role in the epididymal epithelium of the rat. Biol. Reprod. 2015, 92. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.B.; Cortez-Retamozo, V.; Grigoryeva, L.S.; Hill, E.; Pittet, M.J.; Da Silva, N. Mononuclear phagocytes rapidly clear apoptotic epithelial cells in the proximal epididymis. Andrology 2014, 2, 755–762. [Google Scholar] [CrossRef] [PubMed]
- França, L.R.; Auharek, S.A.; Hess, R.A.; Dufour, J.M.; Hinton, B.T. Blood-tissue barriers: Morphofunctional and immunological aspects of the blood-testis and blood-epididymal barriers. Adv. Exp. Med. Biol. 2012, 763, 237–259. [Google Scholar] [PubMed]
- Hu, S.G.; Zou, M.; Yao, G.X.; Ma, W.B.; Zhu, Q.L.; Li, X.Q.; Chen, Z.J.; Sun, Y. Androgenic regulation of β-defensins in the mouse epididymis. Reprod. Biol. Endocrinol. 2014, 12. [Google Scholar] [CrossRef] [PubMed]
- Robaire, B.; Hamzeh, M. Androgen action in the epididymis. J. Androl. 2011, 32, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Hamzeh, M.; Robaire, B. Identification of early response genes and pathway activated by androgens in the initial segment and caput regions of the regressed rat epididymis. Endocrinology 2010, 151, 4504–4514. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, X.; Zhu, G.; Zhou, Z.; Wang, Y.; Chen, D.; Meng, Z. Effect of surgical castration on expression of TRPM8 in urogenital tract of male rats. Mol. Biol. Rep. 2012, 39, 4797–4802. [Google Scholar] [CrossRef] [PubMed]
- Di Salle, E.; Giudici, D.; Radice, A.; Zaccheo, T.; Ornati, G.; Nesi, M.; Panzeri, A.; Délos, S.; Martin, P.M. PNU 157706, a novel dual type I and II5α-reductase inhibitor. J. Steroid. Biochem. Mol. Biol. 1998, 64, 179–186. [Google Scholar] [CrossRef]
- Henderson, N.A.; Robaire, B. Effects of PNU157706, a dual 5α-reductase inhibitor, on rat epididymal sperm maturation and fertility. Biol. Reprod. 2005, 72, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Pujianto, D.A.; Loanda, E.; Sari, P.; Midoen, Y.H.; Soeharso, P. Sperm-associated antigen11A is expressed exclusively in the principal cells of the mouse caput epididymis in an androgen-dependent manner. Reprod. Biol. Endocrinol 2013, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.W.; Qu, L.J.; Zhou, X.Y.; Zhou, J.Y.; Li, G.T.; Bi, J.H.; Guo, X.J.; Li, Z.; Cao, L.; Zhu, Y. Inhibitory effect of dutasteride on the expressions of epididymal Claudin1 and β-catenin in male rats. Zhonghua Nan Ke Xue 2015, 21, 17–22. [Google Scholar] [PubMed]
- Liu, X.; Jin, S.H.; Liu, X.X.; Wang, W.J.; Liu, F.J. Proteome profiling of the sperm maturation milieu in the rhesus monkey (Macaca mulatta) epididymis. Reprod. Fertil. Dev. 2014. [Google Scholar] [CrossRef] [PubMed]
- Dacheux, J.L.; Dacheux, F.; Labas, V.; Ecroyd, H.; Nixon, B.; Jones, R.C. New proteins identified in epididymal fluid from the platypus (Ornithorhynchus anatinus). Reprod. Fertil. Dev. 2009, 21, 1002–1007. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.S.; Webb, S.; Lettice, L.; Tardif, S.; Kilanowski, F.; Tyrrell, C.; Macpherson, H.; Semple, F.; Tennant, P.; Baker, T.; et al. Partial deletion of chromosome 8 β-defensin cluster confers sperm dysfunction and infertility in male mice. PLoS Genet. 2013, 9, e1003826. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Dong, J.; Gu, Y.; Liu, H.; Xin, A.; Shi, H.; Sun, F.; Zhang, Y.; Lin, D.; Diao, H. The novel human β-defensin 114 regulates lipopolysaccharide (LPS)-mediated inflammation and protects sperm from motility loss. J. Biol. Chem. 2013, 288, 12270–12282. [Google Scholar] [CrossRef] [PubMed]
- Seachrist, D.D.; Johnson, E.; Magee, C.; Clay, C.M.; Graham, J.K.; Veeramachaneni, D.N. Keri RA. Overexpression of follistatin in the mouse epididymis disrupts fluid resorption and sperm transit in testicular excurrent ducts. Biol. Reprod. 2012, 87. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, C.M.; Queiróz, D.B.; Patrão, M.T.; Denadai-Souza, A.; Romano, R.M.; Silva, E.J.; Avellar, M.C. Dynamic changes in the spatio-temporal expression of the β-defensin SPAG11C in the developing rat epididymis and its regulation by androgens. Mol. Cell Endocrinol. 2015, 404, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Biswas, B.; Yenugu, S. Transcriptional regulation of the rat sperm-associated antigen 11e (Spag 11e) gene during endotoxin challenge. Mol. Genet. Genomics 2014, 289, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Dorin, J.R.; Barratt, C.L. Importance of β-defensins in sperm function. Mol. Hum. Reprod. 2014, 20, 821–826. [Google Scholar] [CrossRef] [PubMed]
- Narciandi, F.; Lloyd, A.; Meade, K.G.; O’Farrelly, C.A. Novel subclass of bovine β-defensins links reproduction and immunology. Reprod. Fertil. Dev. 2014, 26, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Robson, A.; Houghton, B.C.; Jepson, C.A.; Ford, W.C.; Frayne, J. Epididymal specific, selenium-independent GPX5 protects cells from oxidative stress-induced lipid peroxidation and DNA mutation. Hum. Reprod. 2013, 28, 2332–2342. [Google Scholar]
- Saltzman, A.G.; Hiipakka, R.A.; Chang, C.; Liao, S. Androgen repression of the production of a 29-kilodalton protein and its mRNA in the rat ventral prostate. J. Biol. Chem. 1987, 262, 432–437. [Google Scholar] [PubMed]
- Gravina, G.L.; Festuccia, C.; Millimaggi, D.; Dolo, V.; Tombolini, V.; de Vito, M.; Vicentini, C.; Bologna, M. Chronic azacitidine treatment results in differentiating effects, sensitizes against bicalutamide in androgen-independent prostate cancer cells. Prostate 2008, 68, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Ohsako, S.; Miyabara, Y.; Nishimura, N.; Kurosawa, S.; Sakaue, M.; Ishimura, R.; Sato, M.; Takeda, K.; Aoki, Y.; Sone, H.; Tohyama, C.; et al. Maternal exposure to a low dose of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) suppressed the development of reproductive organs of male rats: Dose-dependent increase of mRNA levels of 5α-reductase type 2 in contrast to decrease of androgen receptor in the pubertal ventral prostate. Toxicol. Sci. 2001, 60, 132–143. [Google Scholar] [PubMed]
- Xie, S.; Zhu, Y.; Ma, L.; Lu, Y.; Zhou, J.; Gui, Y.; Cao, L. Genome-wide profiling of gene expression in the epididymis of α-chlorohydrin-induced infertile rats using an oligonucleotide microarray. Reprod. Biol. Endocrinol. 2010, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- BioBam company. Functional Annotation and Genomics. Available online: http://www.blast2go.org (accessed on 8 July 2015).
- National Institute of Allergy and Infectious Diseases (NIAID), NIH. DAVID Bioinformatics Resources 6.7. Available online: http://david.abcc.ncifcrf.gov (accessed on 15 July 2015).
- Sample Availability: Samples of the compound dual 5α-reductase inhibitor (dutasteride) are available from the authors.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dutasteride | n | VAP (μm/s) | VSL (μm/s) | VCL (μm/s) | ALH (μm) | BCF (Hz) | STR (%) | LIN (%) |
|---|---|---|---|---|---|---|---|---|
| 0 mg/kg | 8 | 148.5 ± 23.0 | 108.3 ± 15.2 | 234.2 ± 33.0 | 14.8 ± 1.3 | 26.5 ± 2.6 | 72.3 ± 4.3 | 48.2 ± 3.6 |
| 40 mg/kg | 8 | 135.8 ± 10.2 | 80.663 ± 7.2 ** | 233.5 ± 32.3 | 12.1 ± 1.6 * | 21.3 ± 1.5 | 59.7 ± 5.2 ** | 36.1 ± 3.3 * |
| Dutasteride | n | Headless (%) | Tailless (%) | Angulated (%) | Broken (%) | Other (%) | Total (%) |
|---|---|---|---|---|---|---|---|
| 0 mg/kg | 8 | 0.51 ± 0.12 | 0.83 ± 0.15 | 3.23 ± 0.29 | 0.25 ± 0.04 | 0.26 ± 0.12 | 5.07 ± 0.83 |
| 40 mg/kg | 8 | 4.12 ± 0.26 * | 4.23 ± 0.26 * | 5.26 ± 0.86 * | 1.29 ± 0.26 * | 0.72 ± 0.03 * | 15.6 ± 3.25 * |
| Dutasteride | Copulation Index | Pregnancy Index | Fertility Index |
|---|---|---|---|
| 0mg/kg | 100 (16/16) | 100 (16/16) | 100 (16/16) |
| 40mg/kg | 81.3 (13/16) | 38.4 (5/13) ** | 31.3 (5/16) * |
| Uniprot Accession | Gene ID | Protein Name | Gene Name | Coverage (%) | Unique Peptides | MW (kDa) | Ratio | p-Value |
|---|---|---|---|---|---|---|---|---|
| Up-regulated proteins | ||||||||
| O88752 | 293267 | hemoglobin, epsilon 1 | Hbe1 | 12.24 | 1 | 16.09 | 3.010 | 0.000002 |
| P04631 | 25742 | S100 calcium binding protein B | S100b | 7.61 | 1 | 10.73 | 2.349 | 0.0002 |
| O88205 | 360445 | cysteine-rich secretory protein 2 | Crisp2 | 4.12 | 1 | 27.33 | 1.975 | 0.003 |
| Q6ZYN8 | 303692 | α-2,6-sialyltransferase 2 | St6galnac2 | 2.41 | 1 | 41.87 | 1.77 | 0.014 |
| O35244 | 94167 | peroxiredoxin 6 | Prdx6 | 40.63 | 9 | 24.80 | 1.746 | 0.017 |
| Q6B345 | 445415 | S100 calcium binding protein A11 (calizzarin) | S100a11 | 9.18 | 1 | 11.06 | 1.744 | 0.017 |
| Q6IE49 | 408234 | serine peptidase inhibitor, Kazal type 2 | Spink2 | 27.91 | 3 | 9.69 | 1.744 | 0.017 |
| P30710 | 113919 | glutathione peroxidase 5 | Gpx5 | 34.84 | 6 | 25.37 | 1.730 | 0.019 |
| P15429 | 25438 | enolase 3, beta, muscle | Eno3 | 13.82 | 1 | 46.98 | 1.718 | 0.020 |
| P14480 | 24366 | fibrinogen beta chain | Fgb | 23.38 | 11 | 54.20 | 1.717 | 0.020 |
| P25809 | 29593 | creatine kinase, mitochondrial 1, ubiquitous | Ckmt1b | 3.83 | 1 | 47.00 | 1.700 | 0.023 |
| P01041 | 25308 | cystatin B (stefin B) | Cstb | 7.14 | 1 | 11.19 | 1.634 | 0.036 |
| P63255 | 691657 | cysteine-rich intestinal protein | Crip1 | 23.38 | 3 | 8.54 | 1.631 | 0.037 |
| P81827 | 619560 | urinary protein 2 | Rup2 | 10.89 | 1 | 10.95 | 1.608 | 0.043 |
| Q9WUH4 | 25177 | four and a half LIM domains 1 | Fhl1 | 5.71 | 1 | 31.88 | 1.593 | 0.047 |
| P37361 | 117038 | metallothionein 3 | Mt3 | 18.18 | 1 | 6.80 | 1.585 | 0.049 |
| Down-regulated proteins | ||||||||
| P10860 | 24399 | glutamate dehydrogenase 1 | Glud1 | 1.43 | 1 | 61.38 | 0.667 | 0.014 |
| Q32ZF7 | 641646 | defensin beta 39 | Defb39 | 20.55 | 1 | 8.30 | 0.660 | 0.011 |
| Q5PQU1 | 24903 | kininogen 1 | Kng1 | 24.65 | 5 | 47.73 | 0.655 | 0.011 |
| P04797 | 24383 | glyceraldehyde-3-phosphate dehydrogenase | Gapdh | 31.23 | 4 | 35.81 | 0.557 | 0.0003 |
| Q32ZH4 | 641655 | defensin beta 18 | Defb18 | 10.59 | 1 | 9.92 | 0.532 | 0.0002 |
| Q32ZI5 | 641631 | defensin beta 2 | Defb2 | 9.86 | 1 | 8.11 | 0.517 | 0.00007 |
| P08932 | 288001 | kininogen 1-like 1 | Kng1l1 | 27.21 | 5 | 47.67 | 0.487 | 0.00001 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, S.-W.; Li, G.-T.; Qu, L.-J.; Cao, Y.; Wang, Q.; Zhou, J.-Y.; Zhong, R.-H.; Guo, X.-J.; Zhu, Y. Identification of New Epididymal Luminal Fluid Proteins Involved in Sperm Maturation in Infertile Rats Treated by Dutasteride Using iTRAQ. Molecules 2016, 21, 602. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21050602
Xie S-W, Li G-T, Qu L-J, Cao Y, Wang Q, Zhou J-Y, Zhong R-H, Guo X-J, Zhu Y. Identification of New Epididymal Luminal Fluid Proteins Involved in Sperm Maturation in Infertile Rats Treated by Dutasteride Using iTRAQ. Molecules. 2016; 21(5):602. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21050602
Chicago/Turabian StyleXie, Shu-Wu, Guo-Ting Li, Li-Juan Qu, Yang Cao, Qian Wang, Jie-Yun Zhou, Rui-Hua Zhong, Xiang-Jie Guo, and Yan Zhu. 2016. "Identification of New Epididymal Luminal Fluid Proteins Involved in Sperm Maturation in Infertile Rats Treated by Dutasteride Using iTRAQ" Molecules 21, no. 5: 602. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21050602