
Phosphorylated Radix Cyathulae officinalis Polysaccharides Act as Adjuvant via Promoting Dendritic Cell Maturation

Abstract
:1. Introduction
2. Results and Discussions
2.1 Results
2.1.1. Chemical Properties of pRCPS
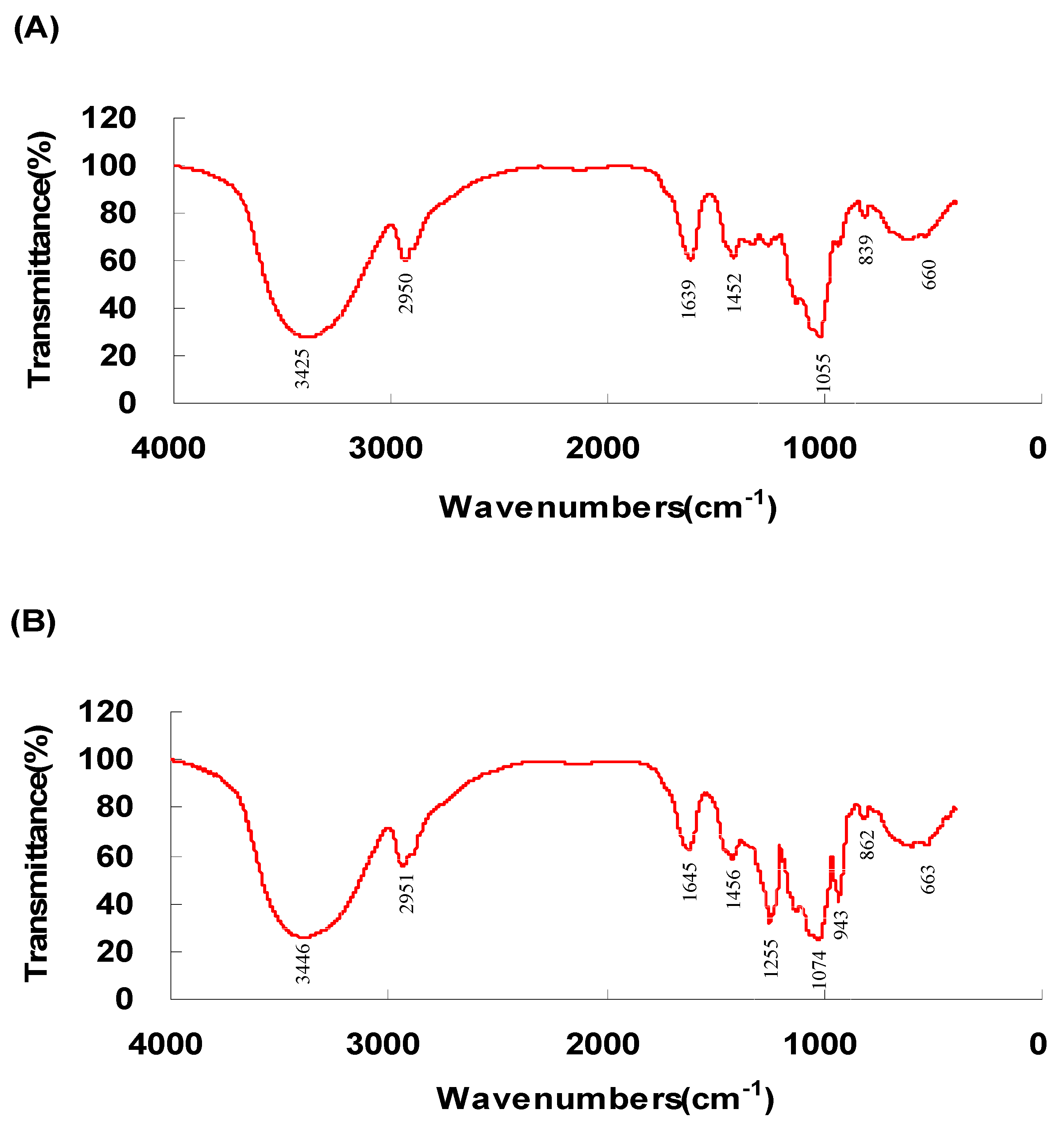
2.1.2. FR-IR Spectroscopy Analysis
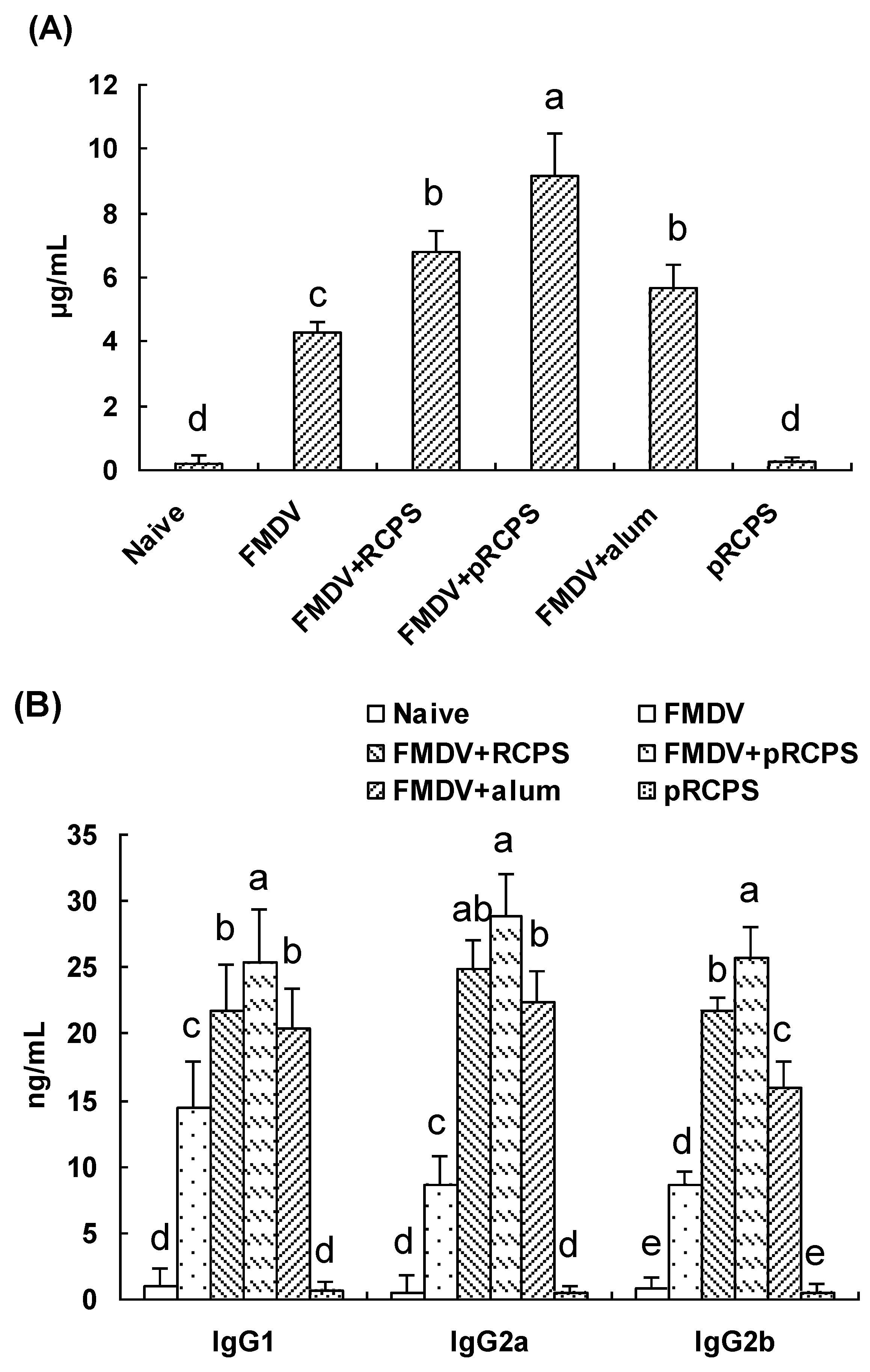
2.1.3. Effects of pRCPS on FMDV-Specific IgG and the IgG Subclasses
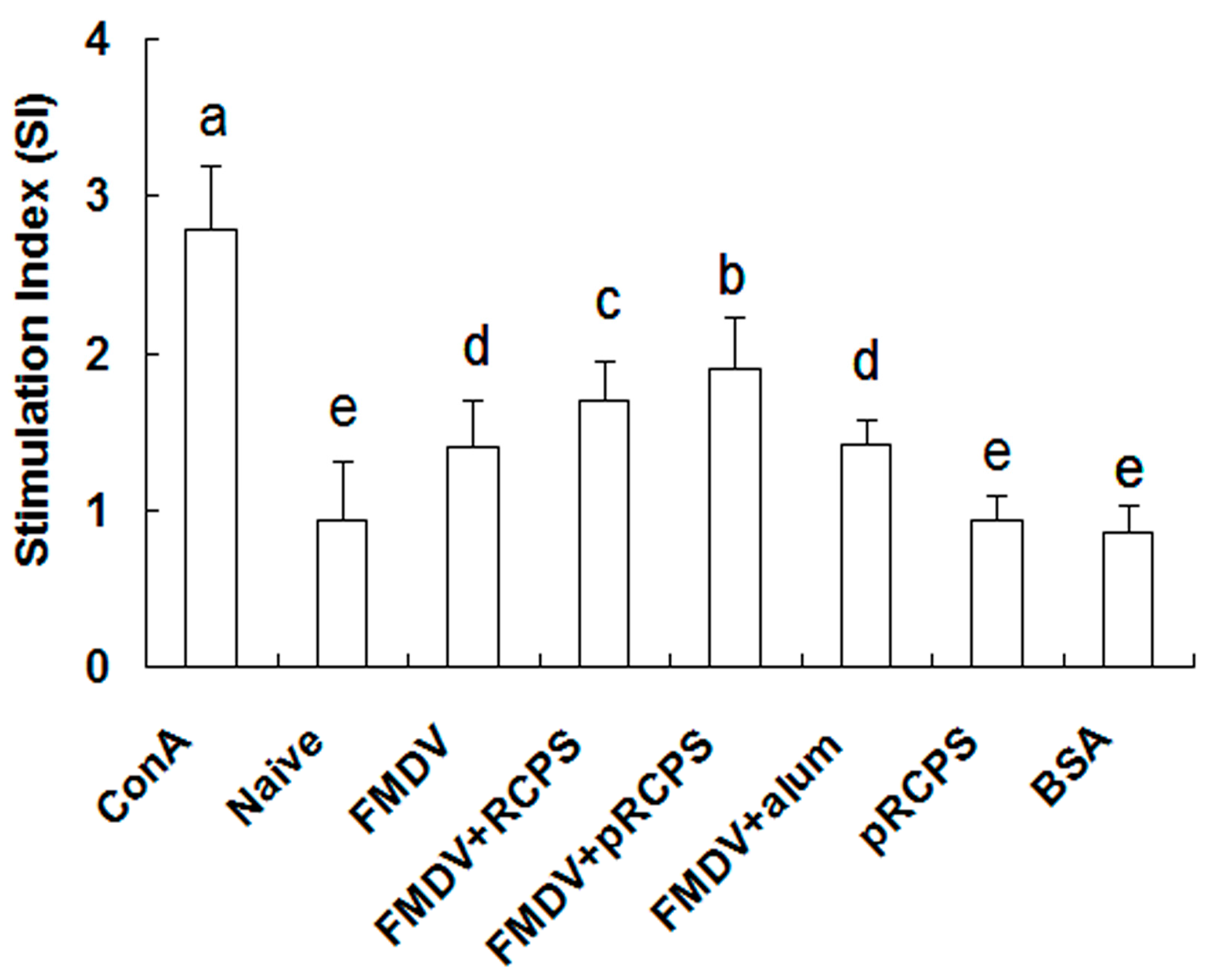
2.1.4. Effects of pRCPS on Splenocyte Proliferation
2.1.5. Effects of pRCPS on Helper T Cells
2.1.6. Effects of pRCPS on Cytotoxic T Lymphocyte (CTL)
2.1.7. Effects of pRCPS on Killing Activity of NK Cell
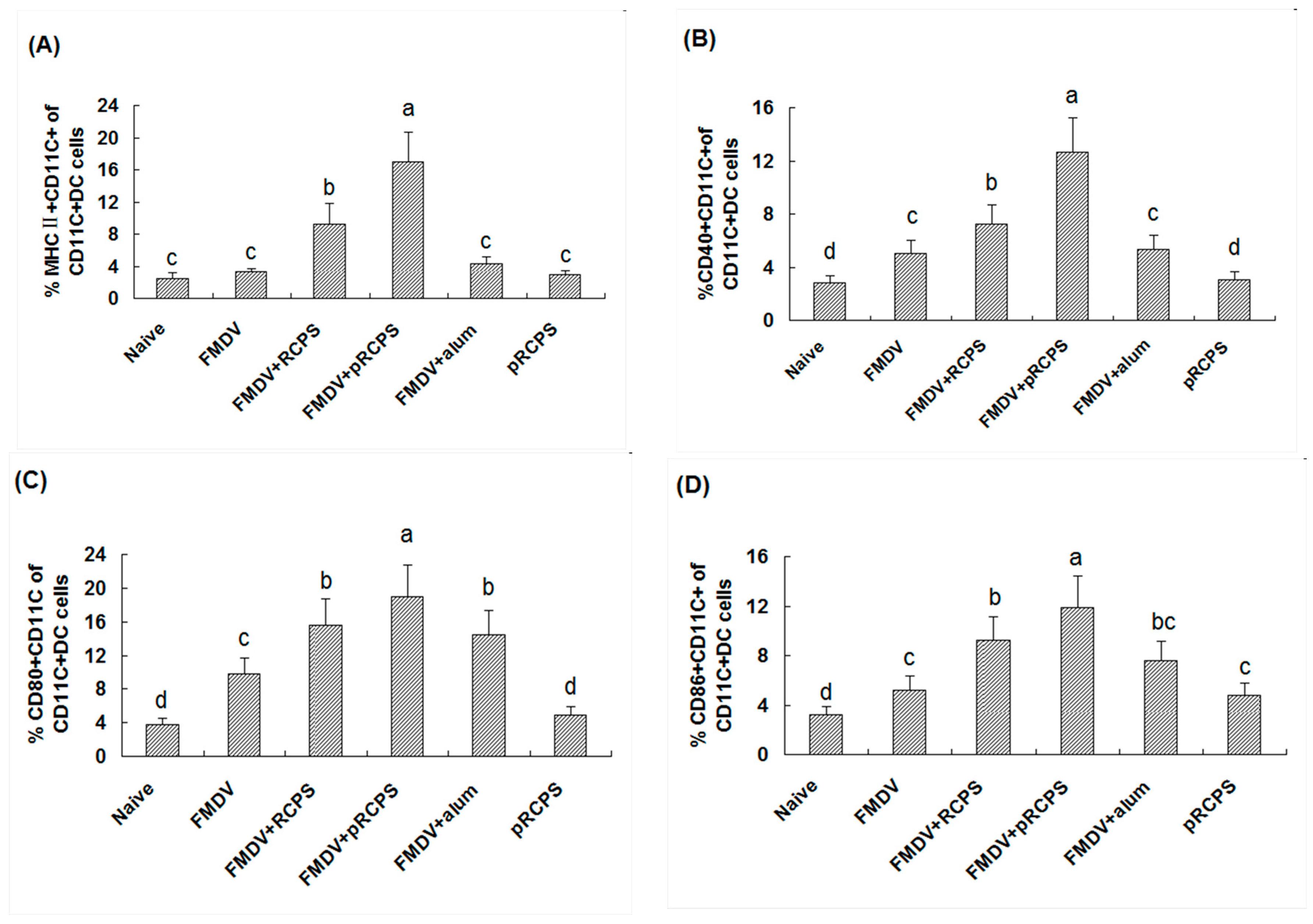
2.1.8. Effects of pRCPS on DCs
2.2. Discussion
3. Experimental Section
3.1. Reagents and Cell Line
3.2. Extraction and Phosphorylated Modification of Polysaccharide
3.3. Characterization of RCPS and pRCPS
3.3.1. Physicochemical Property Analysis
3.3.2. Analysis of Monosaccharide Composition, Uronic Acid, Content of Carbohydrate and Protein of pRCPS
3.3.3. Analysis the Phosphate Graft Quantity
3.3.4. Infrared Spectroscopy Analysis of pRCPS
3.4. Endotoxin Detection
3.5. Animal Vaccination
3.6. FMDV-Specific Antibody ELISA
3.7. Analysis Proliferation Activity of Splenocytes
3.8. Analysis Killing Activity of CTL Cell by Flow Cytometry (FCM)
3.9. Analysis Killing Activity of NK Cell by Flow Cytometry (FCM)
3.10. Analysis Intracellular Cytokine by Flow Cytometry (FCM)
3.11. DCs Surface Co-Stimulatory Molecules Staining
3.12. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| RCPS | Radix Cyathulae officinalis Kuan polysaccharides |
| CFSE | Carboxyfluorescein succinimidyl ester |
| DCs | dendritic cells |
| FACS | fluorescence activated cell sorting |
| FITC | Fluorescein isothiocyanate |
| PE | Phycoerythrin |
| PI | propidium iodide |
| IL-4 | interleukin-4 |
| IL-2 | interleukin-2 |
| IFN-γ | interferon-γ |
| LPS | lipopolysaccharide |
| ConA | Concanavalin A |
| IR | infrared |
| NK | natural killer cells |
| CTL | cytotoxic T lymphocyte |
References
- Engel, A.L.; Sun, A.C.; Gad, E.; Rastetter, L.R.; Strobe, K.; Yang, Y.; Dang, Y.S.; Disis, M.L.; Lu, H.L. Protein-bound polysaccharide activates dendritic cells and enhances OVA-specific T cell response as vaccine adjuvant. Immunobiology 2013, 218, 1468–1476. [Google Scholar] [CrossRef] [PubMed]
- Leunga, M.Y.K.; Liu, C.; Koon, J.C.M.; Fung, K.P. Polysaccharide biological response modifiers. Immunol. Lett. 2006, 105, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.M.; Han, S.Y.; Zhao, D.Y.; Wang, G.Y. Adjuvant effect of polysaccharide from fruits of Physalis alkekengi L. in DNA vaccine against systemic candidiasis. Carbohydr. Polym. 2014, 109, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Su, X.Y.; Pei, Z.Y.; Hu, S.H. Ginsenoside Re as an adjuvant to enhance the immune response to the inactivated rabies virus vaccine in mice. Int. Immunopharmacol. 2014, 20, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Fadi, S.; Yoshikazu, H.O.; Samay, T.; Nikolai, P. A novel hepatitis B vaccine containing Advax™, a polysaccharide adjuvant derived from delta inulin, induces robust humoral and cellular immunity with minimal reactogenicity in preclinical testing. Vaccine 2013, 31, 1999–2007. [Google Scholar]
- Paulsen, B.S. Plant polysaccharides with immunostimulatory activities. Curr. Organ. Chem. 2001, 5, 939–950. [Google Scholar] [CrossRef]
- Ma, X.; Guo, Z.H.; Shen, Z.Q.; Wang, J.L.; Hu, Y.L.; Wang, D.Y. The immune enhancement of propolis adjuvant on inactivated porcine parvovirus vaccine in guinea pig. Cell. Immunol. 2011, 270, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Meng, X.Y.; Yang, R.L.; Qin, T.; Zhang, Y.; Fei, L.F.; Zhen, C.Z.; Zhang, W.L.; Wang, X.Y.; Hu, Y.L.; et al. Cordyceps militaris polysaccharides can improve the immune efficacy of Newcastle disease vaccine in chicken. Int. J. Biol. Macromol. 2013, 59, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Ma, X.; Wu, Y.; Chen, Y.; Zeng, L.; Liu, J.G.; Sun, W.D.; Wang, D.Y.; Hu, Y.L. Determine the structure of phosphorylated modification of icariin and its antiviral activity against duck hepatitis virus A. BMC Vet. Res. 2015, 11, 205. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Wang, D.Y.; Hu, Y.L.; Huang, X.Y.; Wang, J.M. Sulfated modification of epimedium polysaccharide and effects of the modifiers on cellular infectivity of IBDV. Carbohydr. Polym. 2008, 71, 180–186. [Google Scholar] [CrossRef]
- Nguyen, L.; Wang, D.Y.; Hu, Y.L.; Fan, Y.L.; Wang, J.M.; Abula, S.; Guo, L.W.; Zhang, J.; Khakame, S.K.; Zhang, B.K. Immuno-enhancing activity of sulfated Auricularia auricula polysaccharides. Carbohydr. Polym. 2012, 89, 1117–1122. [Google Scholar] [CrossRef] [PubMed]
- Qin, T.; Chen, J.; Wang, D.Y.; Hu, Y.L.; Zhang, J.; Wang, M.; Qiu, S.L.; Gao, Z.Z.; Liu, R.R.; Yu, Y.Y.; et al. Selenylation modification can enhance immune-enhancing activity of Chinese angelica polysaccharide. Carbohydr. Polym. 2013, 95, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Kitazawa, H.; Yamaguchi, T.; Itoh, T. B-cell mitogenic activity of slime products produced from slime-forming, encapsulated Lactococcus lactis ssp. Cremoris. J. Dairy Sci. 1992, 75, 2946–2951. [Google Scholar] [CrossRef]
- Kitazawa, H.; Harata, T.; Uemura, J.; Saito, T.; Kaneko, T.; Itoh, T. Phosphate group requirement for mitogenic activation of lymphocytes by an extracellular phosphopolysaccharide from Lactobacillus delbrueckii ssp. Bulgaricus. Int. J. Food Microbiol. 1998, 40, 169–175. [Google Scholar] [CrossRef]
- Kitazawa, H.; Ishii, Y.; Uemura, J.; Kawai, Y.; Saito, T.; Kaneko, T.; Noda, K.; Itoh, T. Augmentation of macrophage functions by an extracellular phosphopolysaccharide from Lactobacillus delbrueckii ssp. Bulgaricus. Food Microbiol. 2000, 17, 109–118. [Google Scholar] [CrossRef]
- Sato, T.; Nishimura-Uemura, J.; Shimosato, T.; Kawai, Y.; Kitazawa, H.; Saito, T. Dextran from Leuconostoc mesenteroides Augments Immunostimulatory effects by the introduction of phosphate groups. J. Food Prot. 2004, 67, 1719–1724. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.N.; Hu, Y.L.; Wang, D.Y.; Liu, J.Z.; Guo, L.W. The comparison of immune-enhancing activity of sulfated polysaccharidses from Tremella and Condonpsis pilosula. Carbohydr. Polym. 2013, 98, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, C.; Nishimura-Uemura, J.; Tohno, M.; Shimosato, T.; Kawai, Y.; Ikegami, S.; Oda, M.; Saito, T.; Kitazawa, H. Oral administration of phosphorylated dextran regulates immune response in ovalbumin-immunized mice, Asian-Aust. J. Anim. Sci. 2010, 23, 106–115. [Google Scholar]
- Feng, H.B.; Du, X.G.; Tang, J.; Cao, X.H.; Han, X.F.; Chen, Z.Y.; Chen, Y.E.; Zeng, X.Y. Enhancement of the immune responses to foot-and-mouth disease vaccination in mice by oral administration of a Novel polysaccharide from the roots of Radix Cyathulae officinalis Kuan (RC). Cell. Immunol. 2013, 281, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.B.; Du, X.G.; Cao, X.H.; Wang, Z.H.; Tang, J. Enhancement of the immune responses to ovalbumin in mice by oral administration of the extract from Radix Cyathulae (RC). J. Med. Plant Res. 2013, 7, 1272–1279. [Google Scholar]
- Feng, H.B.; Du, X.G.; Liu, J.; Han, X.F.; Cao, X.H.; Zeng, X.Y. Novel polysaccharide from Radix Cyathulae officinalis Kuan can improve immune response to ovalbumin in mice. Int. J. Biol. Macromol. 2014, 65, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Ni, Q.S.; Wang, B.; Li, X.Y. Effects of achyranthes bidentata polysaccharides on sera antibody and blood biochemical parameters in chicken. Sichuan Anim. Vet. Sci. 2011, 38, 27–31. [Google Scholar]
- Jia, R.Y.; Zhou, M.J.; Ge, Y. Effect of polysaccharide from the Radix Cyathulae officinalis Kuan (RC) on immune function of peripheral blood lymphocyte. Chin. J. Vet. Med. 2009, 45, 35–38. [Google Scholar]
- Zhang, J.; Chen, J.; Wang, D.Y.; Hu, Y.L.; Zhang, Y.S.; Qin, T.; Liu, C.; Sheng, X.; Nguyen, L. Immune-enhancing activity comparison of sulfated ophiopogonpolysaccharide and sulfated jujube polysaccharide. Int. J. Biol. Macromol. 2013, 52, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Suflet, D.M.; Chitanu, G.C.; Desbrières, J. Phosphorylated polysaccharides. 2. Synthesis and properties of phosphorylated dextran. Carbohydr. Polym. 2010, 82, 1271–1277. [Google Scholar] [CrossRef]
- Montomoli, E.; Piccirella, S.; Khadang, B.; Mennitto, E.; Camerini, R.; de Rosa, A. Current adjuvants and new perspectives in vaccine formulation. Expert Rev. Vaccines 2011, 10, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Soren, K.; Ricardo, S.M.; Alfredo, D.; Hans, G.; Kristian, D. Preparation and characterisation of quillaja saponin with less heterogeneity than Quil-A. Vaccine 2000, 18, 2244–2249. [Google Scholar]
- Sun, W.J.; Meng, K.; Qi, C.H.; Yang, X.Y.; Wang, Y.G.; Fan, W.T.; Yan, Z.G.; Zhao, X.N.; Liu, A.S. Immune-enhancing activity of polysaccharides isolated from Atractylodis macrocephalae Koidz. Carbohydr. Polym. 2015, 126, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Chen, J.; Qin, T.; Hu, Y.; Wang, D.; Fan, Q.; Zhang, C.; Chen, X.; Chen, X.; Liu, C.; et al. Effects of Selenylation Modification on Immune-Enhancing Activity of Garlic Polysaccharide. PLoS ONE 2014, 9, e86377. [Google Scholar] [CrossRef] [PubMed]
- Cribbs, D.H.; Ghochikyan, A.; Vasilevko, V.; Tran, M.; Petrushina, I.; Sadzikava, N. Adjuvant-dependent modulation of Th1 and Th2 responses to immunization with beta-amyloid. Int. Immunol. 2003, 15, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Mazumdar, T.; Anam, K.; Ali, N. A mixed Th1/Th2 response elicited by a liposomal formulation of Leishmania vaccine instructs Th1 responses and resistance to Leishmania donovaniin susceptible BALB/c mice. Vaccine 2004, 22, 1162–1171. [Google Scholar] [CrossRef] [PubMed]
- Shahabi, S.; Azizi, H.; Mazloomi, E.; Tappeh, K.H.; Seyedi, S.; Mohammadzadeh, H. A novel adjuvant, the mixture of alum and naltrexone, augments vaccine-induced immunity against Plasmodium berghei. Immunol. Investig. 2014, 43, 653–666. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, S. T-cell subsets (Th1 versus Th2). Annu. Allergy Asthma Immunol. 2000, 85, 9–18. [Google Scholar] [CrossRef]
- Odbileg, R.; Lee, S.I.; Ohashi, K.; Onuma, M. Cloning and sequence analysis of llama (lama glama), Th2 (IL-4, IL-10 and IL-13), cytokines. Vet. Immunol. Immunopathol. 2005, 104, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, T.; Murakami, K.; Nishimura, I.; Nakano, T.; Obata, A. A sulfated polysaccharide, fucoidan, enhances the immunomodulatory effects of lactic acid bacteria. Int. J. Mol. Med. 2012, 29, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Han, Y.P.; Jin, K.; Wan, Y.F. OL-048 Dynamic changes of CTLs, NK cells, and NKT cells in patients with acute hepatitis B infection. Int. J. Infect. Dis. 2011, 15, 31. [Google Scholar] [CrossRef]
- Shimada, M.; Yoshizaki, S.; Ichino, M.; Klinman, D.M.; Okuda, K. Apoptosis of antigen-specific CTLs contributes to low immune response in gut-associated lymphoid tissue post vaccination. Vaccine 2014, 32, 5198–5205. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.S.; Wu, Y.W.; Xu, S.F.; Sun, H.X.; Chen, F.Y.; Yao, L. Antitumor and immunomodulatory activity of polysaccharides from the roots of Actinidia eriantha. J. Ethnopharmacol. 2009, 125, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, Z.; Sheng, W.; Fu, R.; Li, L.; Zhang, T.; Wu, Y.; Xing, L.; Song, J.; Wang, H.; et al. Abnormalities of quantities and functions of natural killer cells in severe aplastic anemia. Immunol. Investig. 2014, 43, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Moretta, L.; Bottino, C.; Cantoni, C.; Mingari, M.C.; Moretta, A. Human natural killer cell function and receptors. Curr. Opin. Pharmacol. 2001, 1, 387–391. [Google Scholar] [CrossRef]
- Wang, H.; Zheng, X.; Wei, H.; Tian, Z.; Sun, R. Important role for NKp30 in synapse formation and activation of NK cells. Immunol. Investig. 2012, 41, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Raïch-Reguéa, D.; Glancy, M.; Thomson, A.W. Regulatory dendritic cell therapy: From rodents to clinical application. Immunol. Lett. 2014, 161, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Park, M.J.; Ryu, H.S.; Kim, J.S.; Lee, H.K.; Kang, J.S.; Yun, J.; Kim, S.Y.; Lee, M.K.; Hong, J.T.; Kim, Y.; et al. Platycodon grandiflorum polysaccharide induces dendritic cell maturation via TLR4 signaling. Food Chem. Toxicol. 2014, 72, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.G.; Xu, W.; Sun, L.; You, Y.; Li, F.; Li, Q.B.; Zou, P. Immunomodulatory function of regulatory dendritic cells induced by mesenchymal stem cells. Immunol. Investig. 2012, 41, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Miga, A.; Masters, S.; Gonzalez, M.; Noelle, R.J. The role of CD40-CD154 interactions in the regulation of cell mediated immunity. Immunol. Investig. 2000, 29, 111–114. [Google Scholar] [CrossRef]
- Shin, B.R.; Kim, H.S.; Yun, M.J.; Lee, H.K.; Kim, Y.J.; Kim, S.Y.; Lee, M.K.; Hong, J.T.; Kim, Y.; Han, S.B. Promoting effect of polysaccharide isolated from Morifructus on dendritic cell maturation. Food Chem. Toxicol. 2013, 51, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Zhao, B.; Li, J.; Cao, X.; Diao, M.; Feng, H.; Chen, X.; Chen, Z.; Zeng, X. Astragaluspolysaccharides enhance immune responses of HBV DNA vaccination via moting the dendritic cell maturation and suppressing Treg frequency in mice. Int. Immunopharmacol. 2012, 14, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.W.; Li, J.F.; Hu, Y.X.; Cheng, G.L.; Zhu, X.Y.; Liu, F.Q.; Zhang, Y.J.; Liu, Z.J.; Xu, J.Q. Effects of astragalus polysaccharide on the immune response to foot-and-mouth disease vaccine in mice. Carbohydr. Polym. 2010, 82, 680–686. [Google Scholar] [CrossRef]
- Sun, X.; Pan, D.D.; Zeng, X.Q.; Cao, J.X. Phosphorylation Modification of Polysaccharides from Entermorpha. Food Sci. 2013, 24, 73–77. [Google Scholar]
- Feng, H.B.; Fan, J.; Song, Z.H.; Du, X.G.; Chen, Y.; Wang, J.S.; Song, G.D. Characterization and immunoenhancement activities of Eucommia ulmoides polysaccharides. Carbohydr. Polym. 2016, 136, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Han, X.F.; Shen, S.; Liu, T.; Du, X.G.; Cao, X.H.; Feng, H.B.; Zeng, X.Y. Characterization and antioxidant activities of the polysaccharides from Radix Cyathulae officinalis Kuan. Int. J. Biol. Macromol. 2015, 72, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Filisetti-Cozzi, T.M.; Carpita, N.C. Measurement of uronic acids without interference from neutral sugars. Anal. Biochem. 1991, 197, 157–162. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Biochem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Z.Y.; Yang, L.; Yang, X.; Wang, X.; Zhang, Z. In vitro antioxidant activities of sulfated derivatives of polysaccharides extracted from Auricularia Auricular. Int. J. Mol. Sci. 2011, 12, 3288–3302. [Google Scholar] [CrossRef] [PubMed]
- Veterinary Pharmacopoeia Commission of the People’s Republic of China. Veterinary. In Pharmacopoeia of the People’s Republic of China, Part I; Chemical Industrial Press: Beijing, China, 2010; pp. 88–89. [Google Scholar]
- Du, X.G.; Chen, X.B.; Zhao, B.; Lv, Y.; Zhang, H.Y.; Liu, H.M.; Chen, Z.Y.; Chen, Y.E.; Zeng, X.Y. Astragaluspolysaccharides enhance the humoral and cellular immune responses of hepatitis B surface antigen vaccination through inhibiting the expression of transforming growth factorband the frequency of regulatory T cells. FEMS Immunol. Med. Microbiol. 2011, 63, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.B.; Fan, J.; Du, X.G.; Song, Z.H.; Wang, Z.H.; Han, X.F. Sulfated Radix Cyathulae officinalis polysaccharides act as adjuvant via promoting the dendritic cell maturation and suppressing treg frequency. Immunol. Investig. 2015, 44, 288–308. [Google Scholar] [CrossRef] [PubMed]
- Oehen, S.; Brduscha-Riem, K. Differentiation of naive CTL to effector and memory CTL, correlation of effector function with phenotype and cell division. J. Immunol. 1998, 161, 5338–5546. [Google Scholar] [PubMed]
- Zou, Q.; Zhong, Y.W.; Su, H.R.; Kang, Y.M.; Jin, J.; Liu, Q.; Geng, S.; Zhao, G.; Wang, B. Enhancement of humoral and cellular responses to HBsAg DNA vaccination by immunization with praziquantel through inhibition TGF-β/Smad 2,3 signaling. Vaccine 2010, 28, 2032–2038. [Google Scholar] [CrossRef] [PubMed]
- Dumitriu, I.; Mohr, W.; Kolowos, W.; Kern, P.; Kalden, J.; Herrmann, M. 5,6-Carboxyfluorescein diacetate succnimidyl ester-labeled apoptotic and necrotic as well as detergent treated cells can be traced in composite cell samples. Anal. Biochem. 2001, 299, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Sample of the compound pRCPS is available from the authors.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | CD4+ IL-4 | CD4+ IL-2 | CD4+ IFN-γ | CD8+ IFN-γ |
|---|---|---|---|---|
| Naive | 1.32 ± 0.21 d | 1.55 ± 0.16 d | 1.41 ± 0.18 d | 0.98 ± 0.07 d |
| FMDV | 3.35 ± 0.37 c | 2.98 ± 0.28 c | 2.17 ± 0.28 c,d | 2.13 ± 0.09 c |
| FMDV + RCPS | 6.31 ± 0.28 b | 4.02 ± 0.36 b | 7.19 ± 1.04 b | 4.19 ± 0.07 b |
| FMDV + pRCPS | 9.91 ± 0.56 a | 6.18 ± 0.23 a | 8.67 ± 1.31 a | 6.48 ± 0.19 a |
| FMDV + alum | 4.95 ± 0.38 c | 3.11 ± 0.14 c | 3.73 ± 0.48 c | 2.93 ± 0.07 c |
| pRCPS | 1.89 ± 0.24 d | 1.67 ± 0.17 d | 1.93 ± 0.39 d | 1.56 ± 0.31 d |
| Group | Vaccine | Adjuvant |
|---|---|---|
| Naive | Naive | |
| FMDV | 200 μL FMD vaccine | |
| FMDV + RCPS | 200 μL FMD vaccine | 0.5 mg pRCPS |
| FMDV + pRCPS | 200 μL FMD vaccine | 0.5 mg pRCPS |
| FMDV + alum | 200 μL FMD vaccine | 200 μg alum |
| pRCPS | 0.5 mg pRCPS |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, H.; McDonough, S.P.; Fan, J.; Yang, S.; Zhao, X.; Lu, Y.; Gan, Y.; Yi, X.; Chang, Y.-F. Phosphorylated Radix Cyathulae officinalis Polysaccharides Act as Adjuvant via Promoting Dendritic Cell Maturation. Molecules 2017, 22, 106. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22010106
Feng H, McDonough SP, Fan J, Yang S, Zhao X, Lu Y, Gan Y, Yi X, Chang Y-F. Phosphorylated Radix Cyathulae officinalis Polysaccharides Act as Adjuvant via Promoting Dendritic Cell Maturation. Molecules. 2017; 22(1):106. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22010106
Chicago/Turabian StyleFeng, Haibo, Sean P. McDonough, Jing Fan, Shiping Yang, Xuelian Zhao, Yong Lu, Yun Gan, Xiao Yi, and Yung-Fu Chang. 2017. "Phosphorylated Radix Cyathulae officinalis Polysaccharides Act as Adjuvant via Promoting Dendritic Cell Maturation" Molecules 22, no. 1: 106. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22010106