
Biosynthesis of Oligomeric Anthocyanins from Grape Skin Extracts

 ,
,
Abstract
:1. Introduction
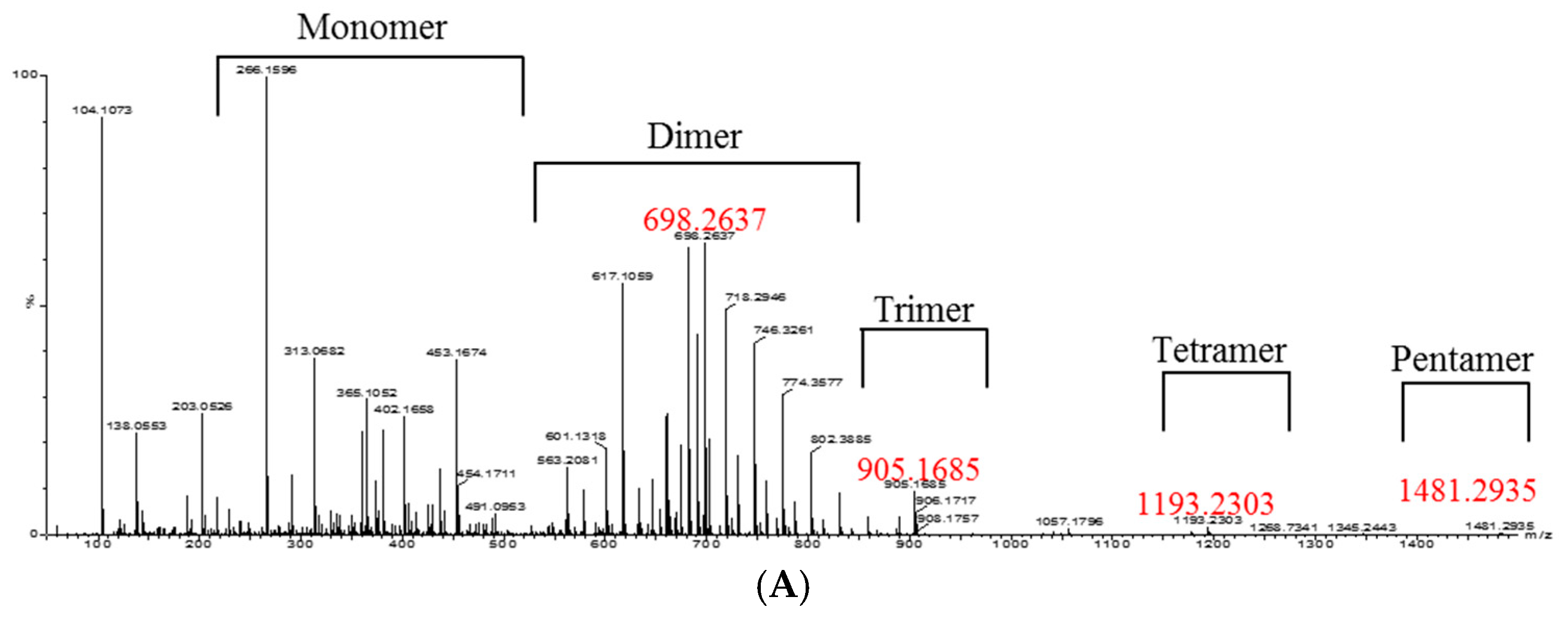
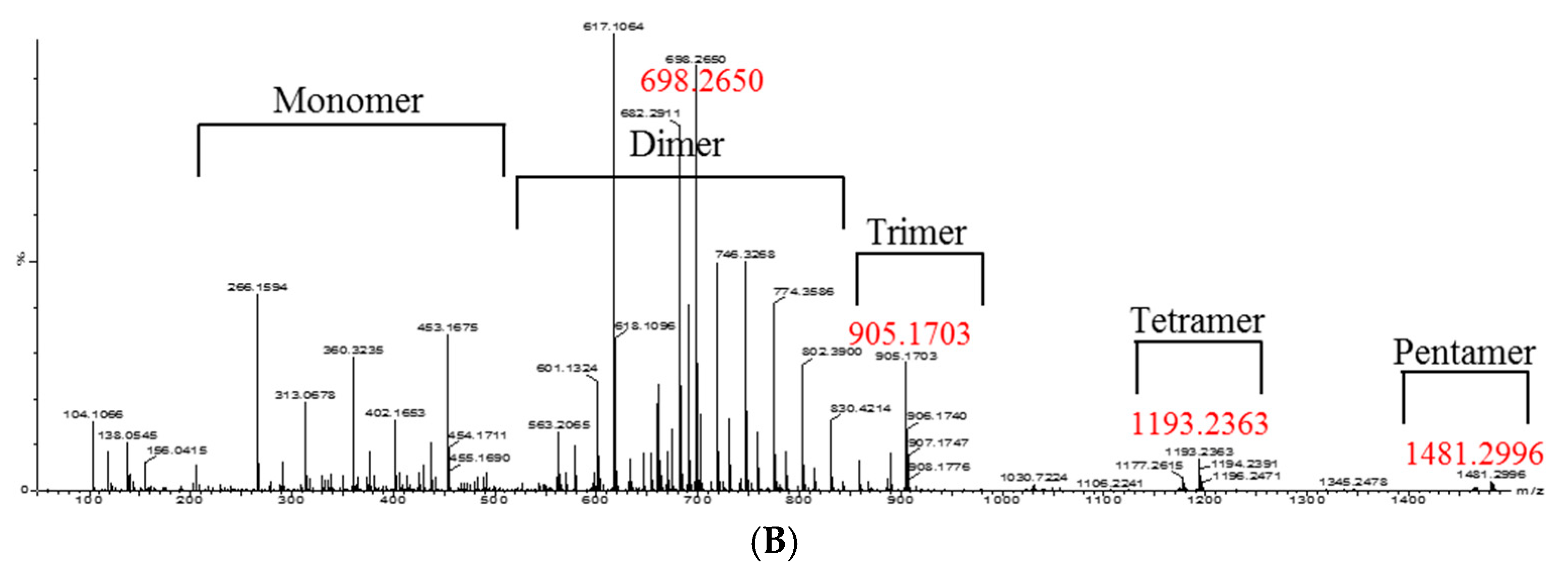
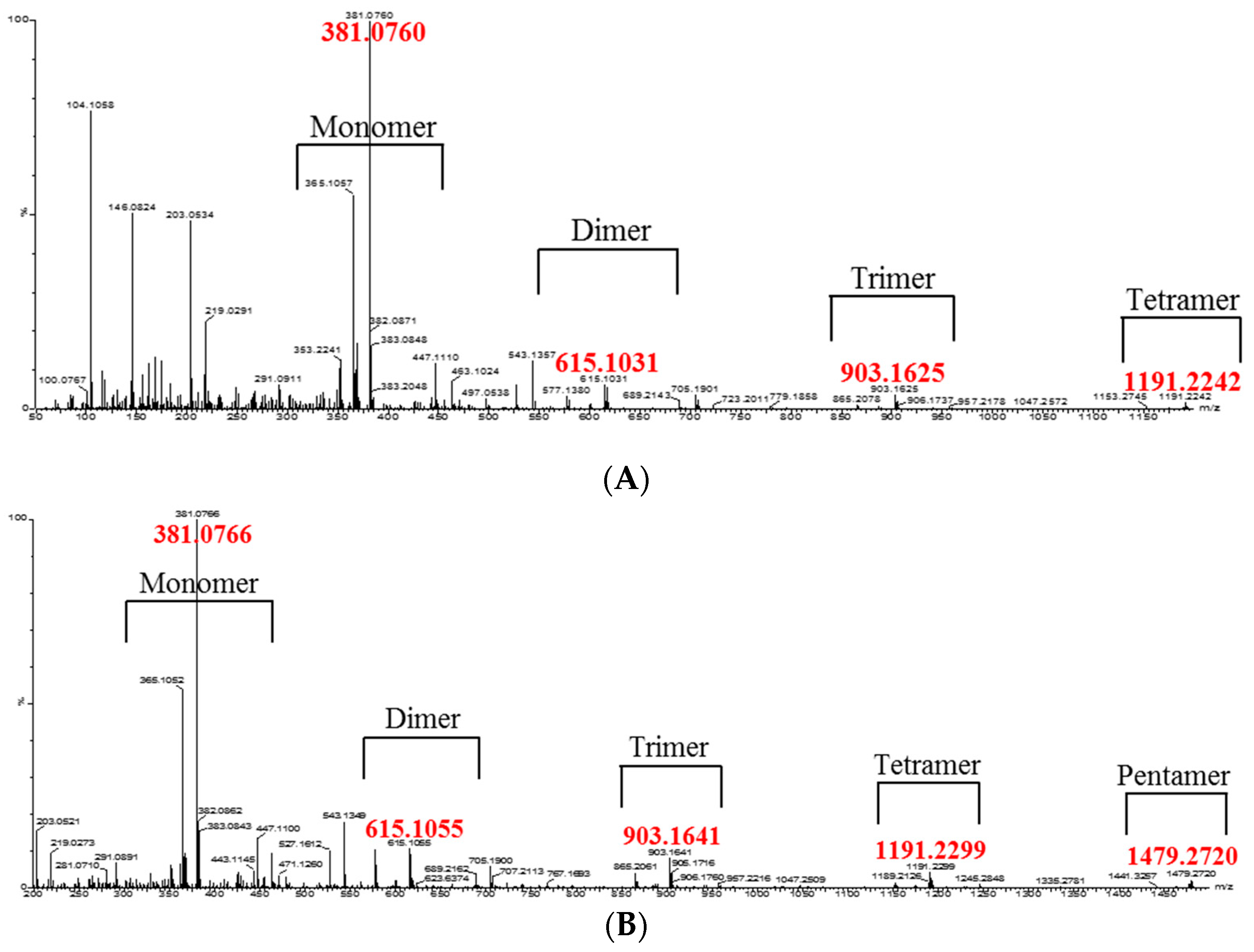
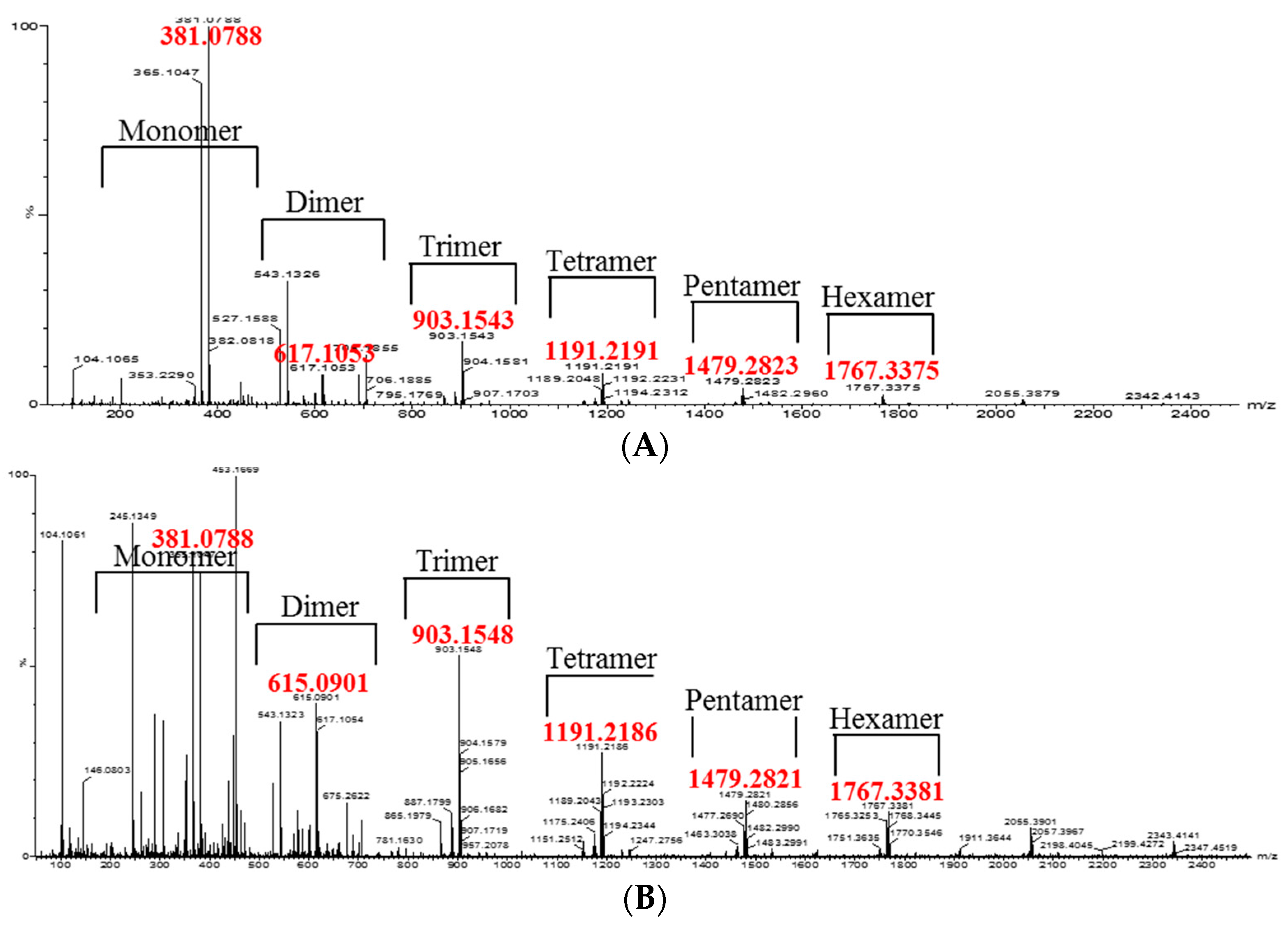
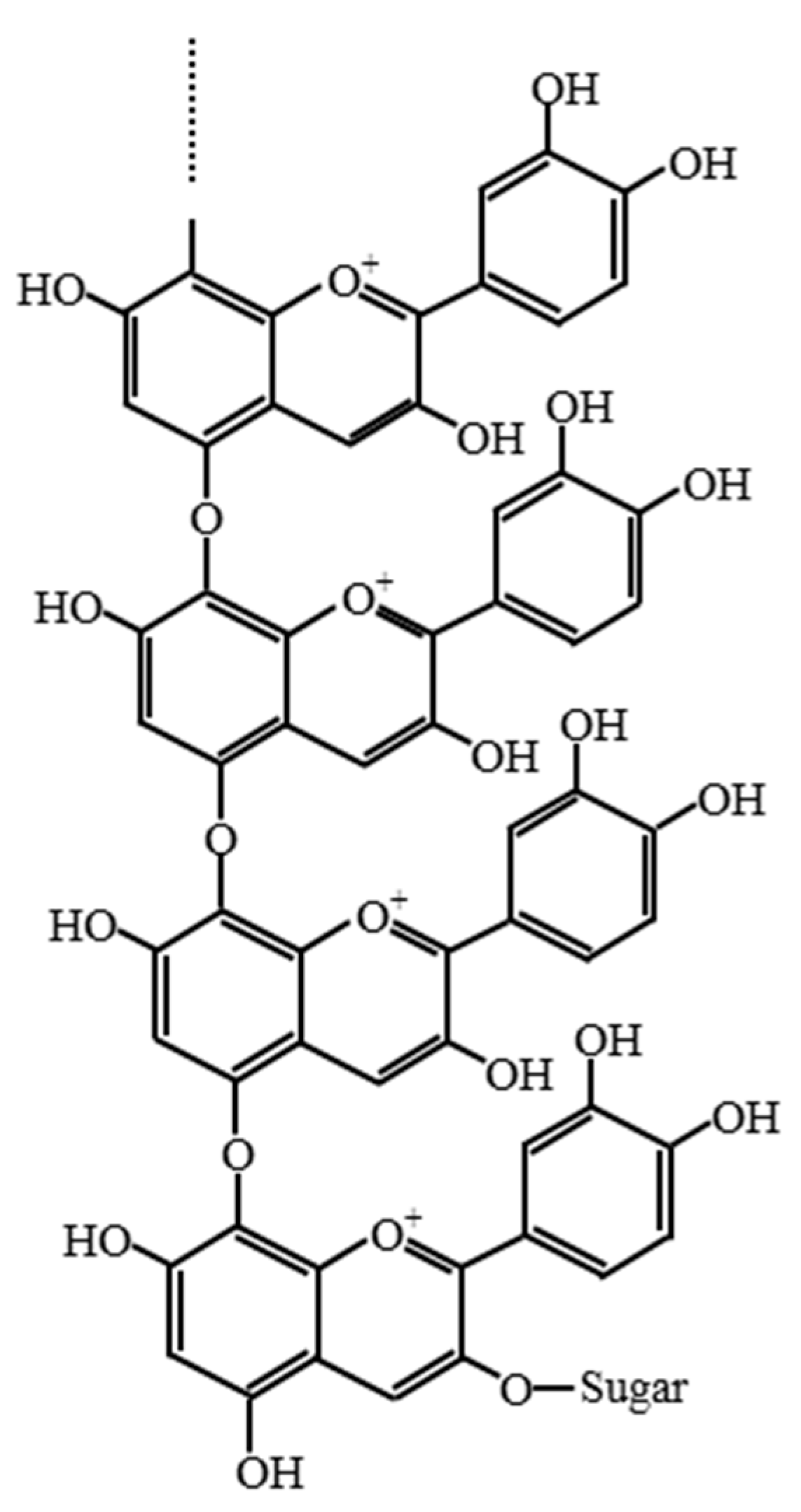
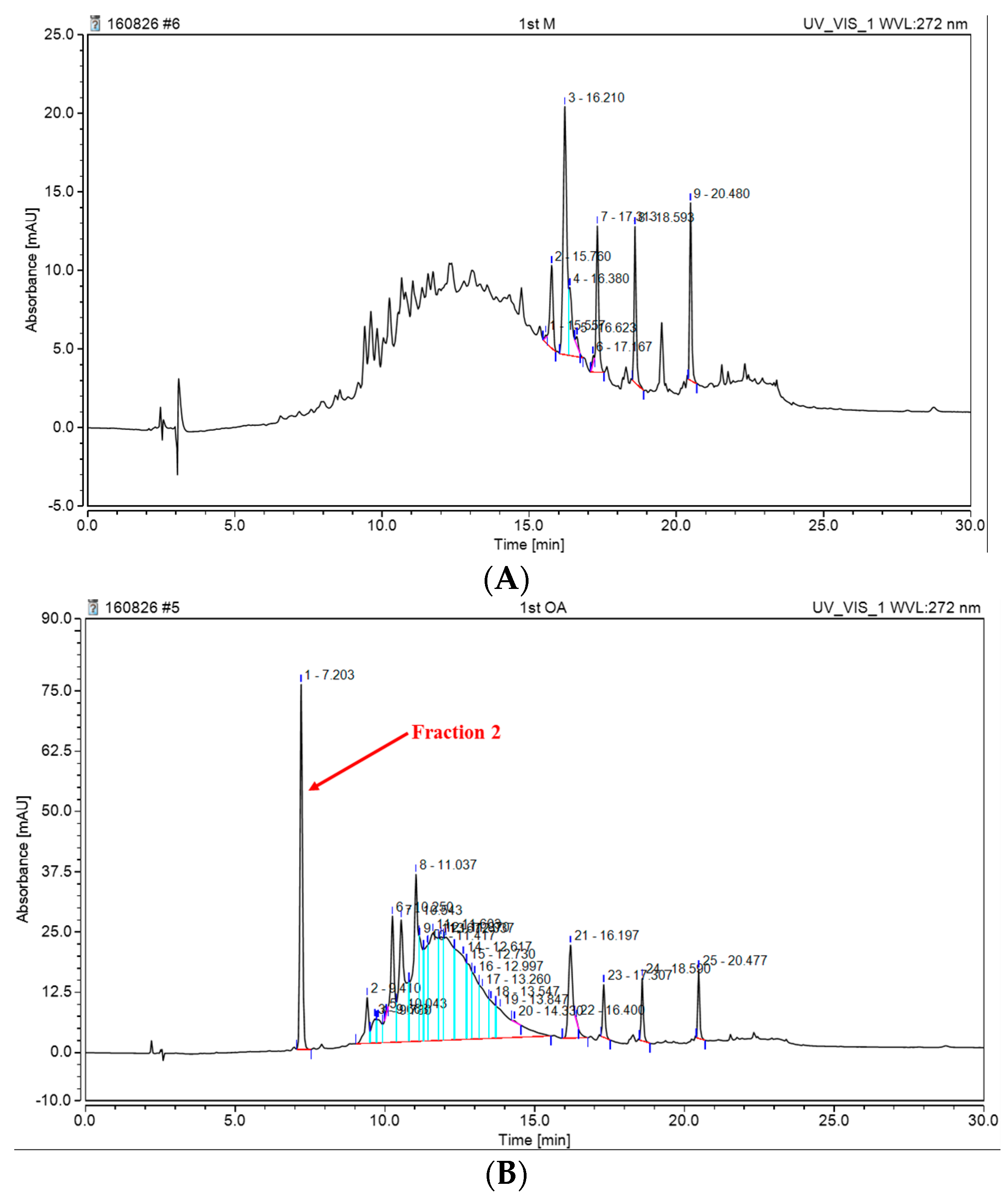
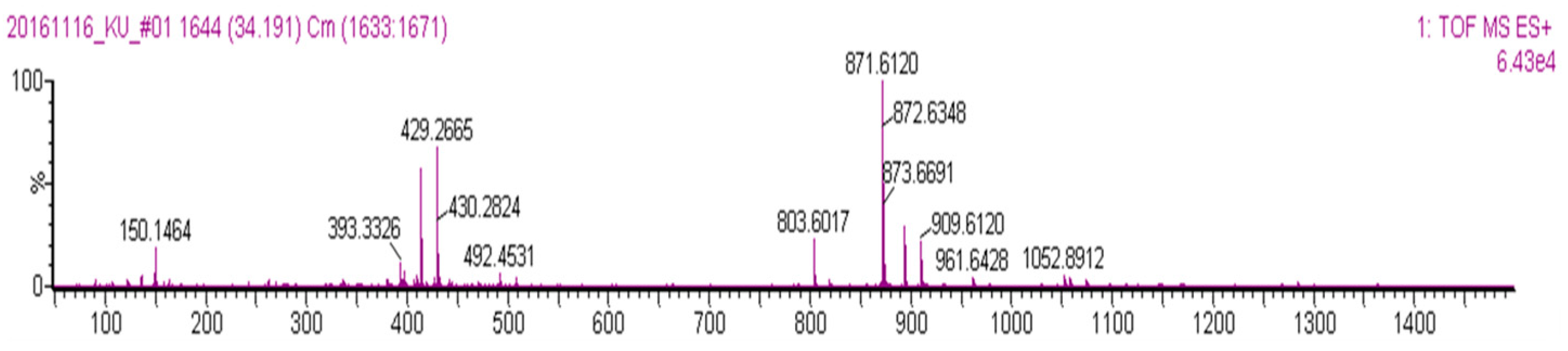
2. Results and Discussion
3. Materials and Methods
3.1. Materials
3.2. Culture Medium of Aspergillus niger
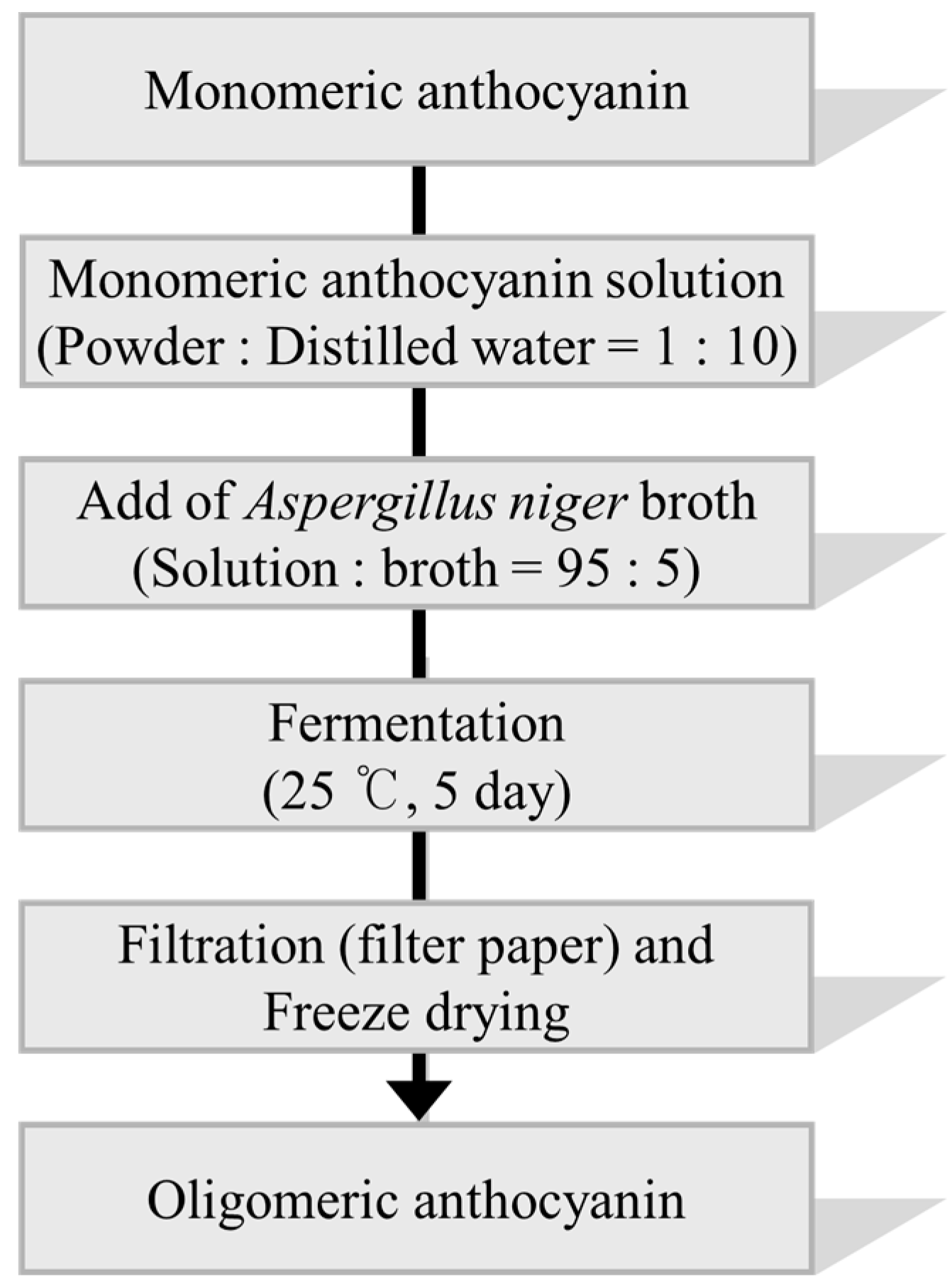
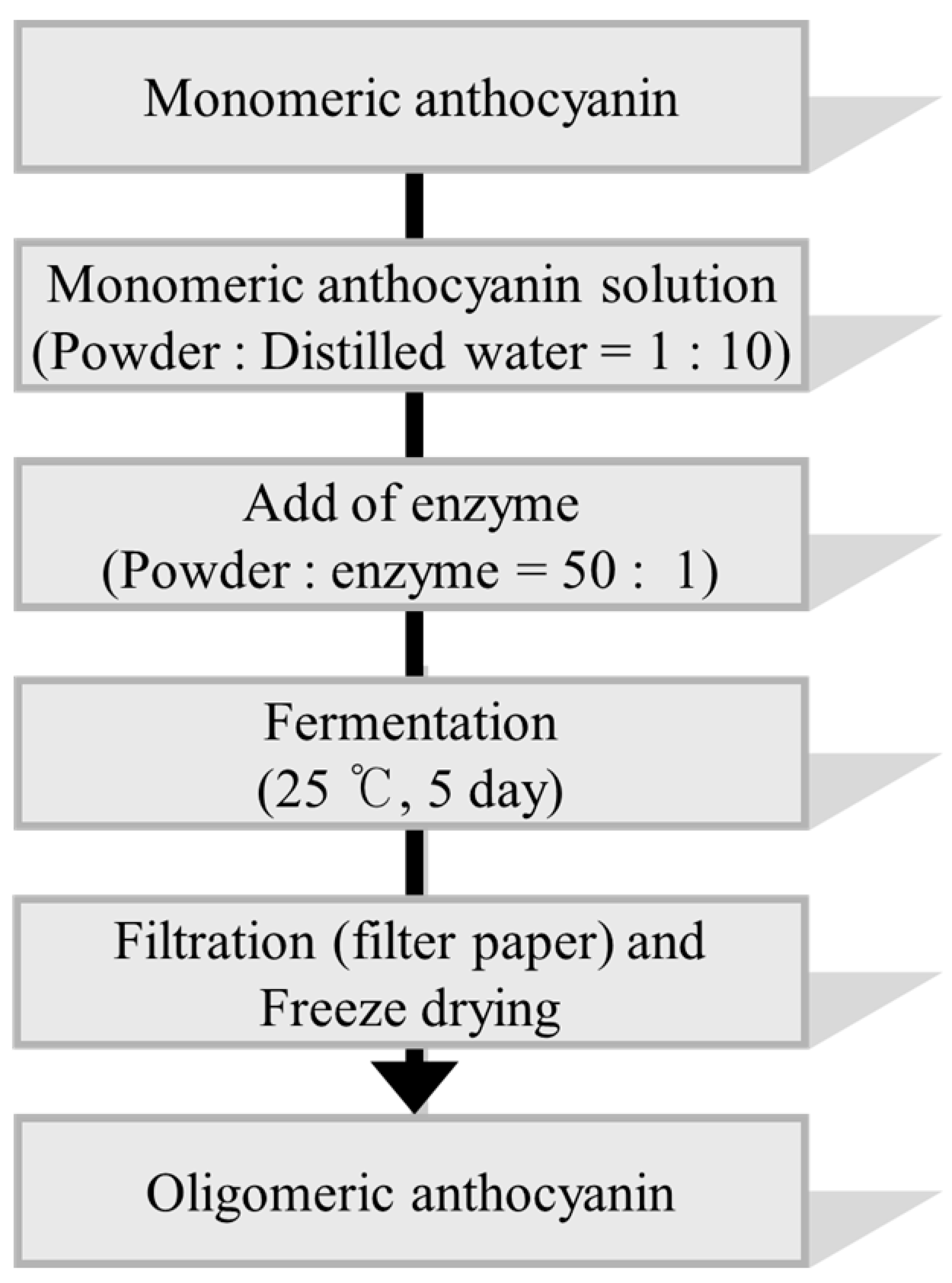
3.3. Synthesis of Oligomeric Anthocyanin by Fermentation Using Aspergillus niger
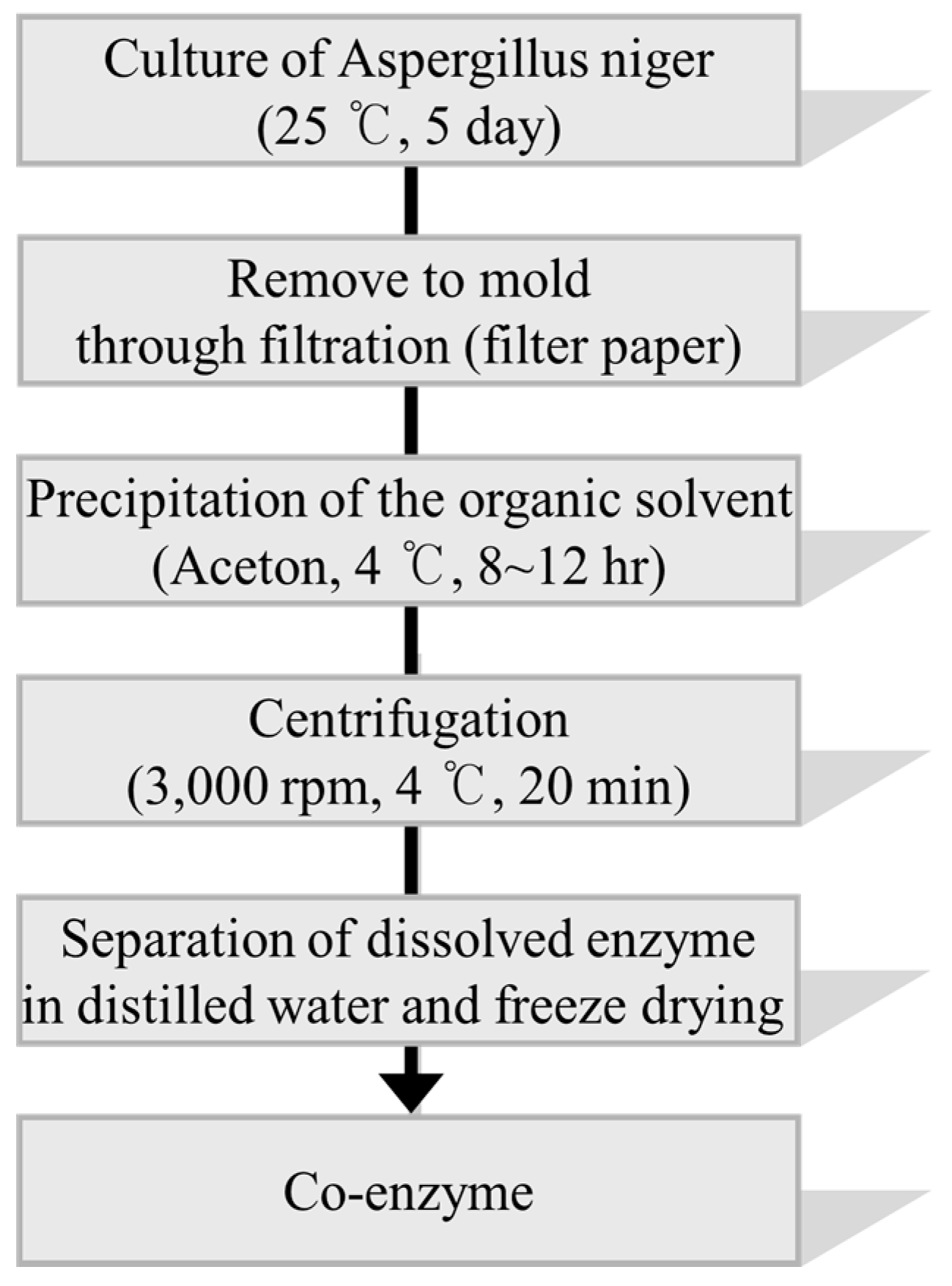
3.4. Separation of Fermented Crude Enzyme from the Culture of Aspergillus niger
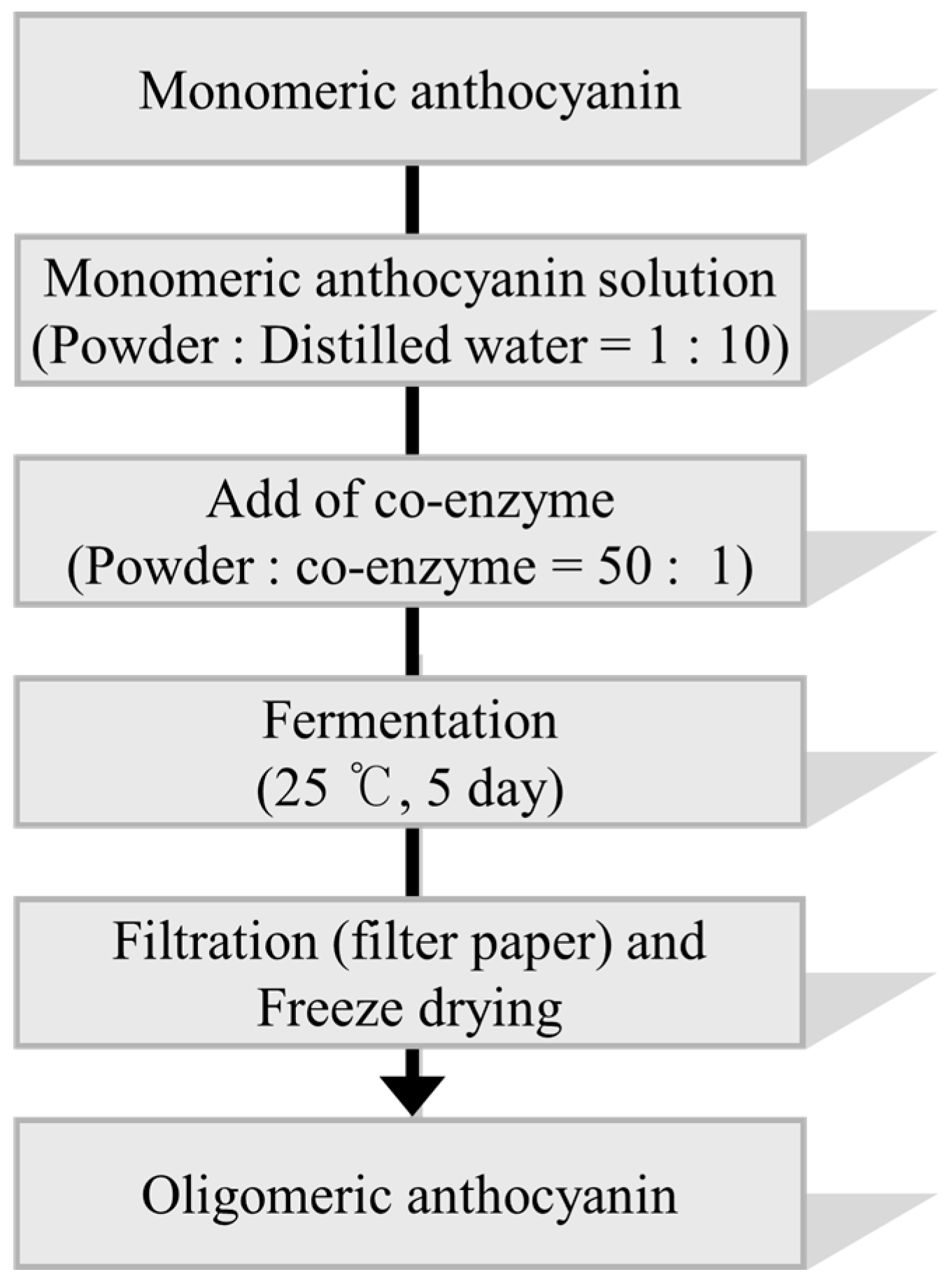
3.5. Synthesis of Oligomeric Anthocyanin Using Crude Enzyme
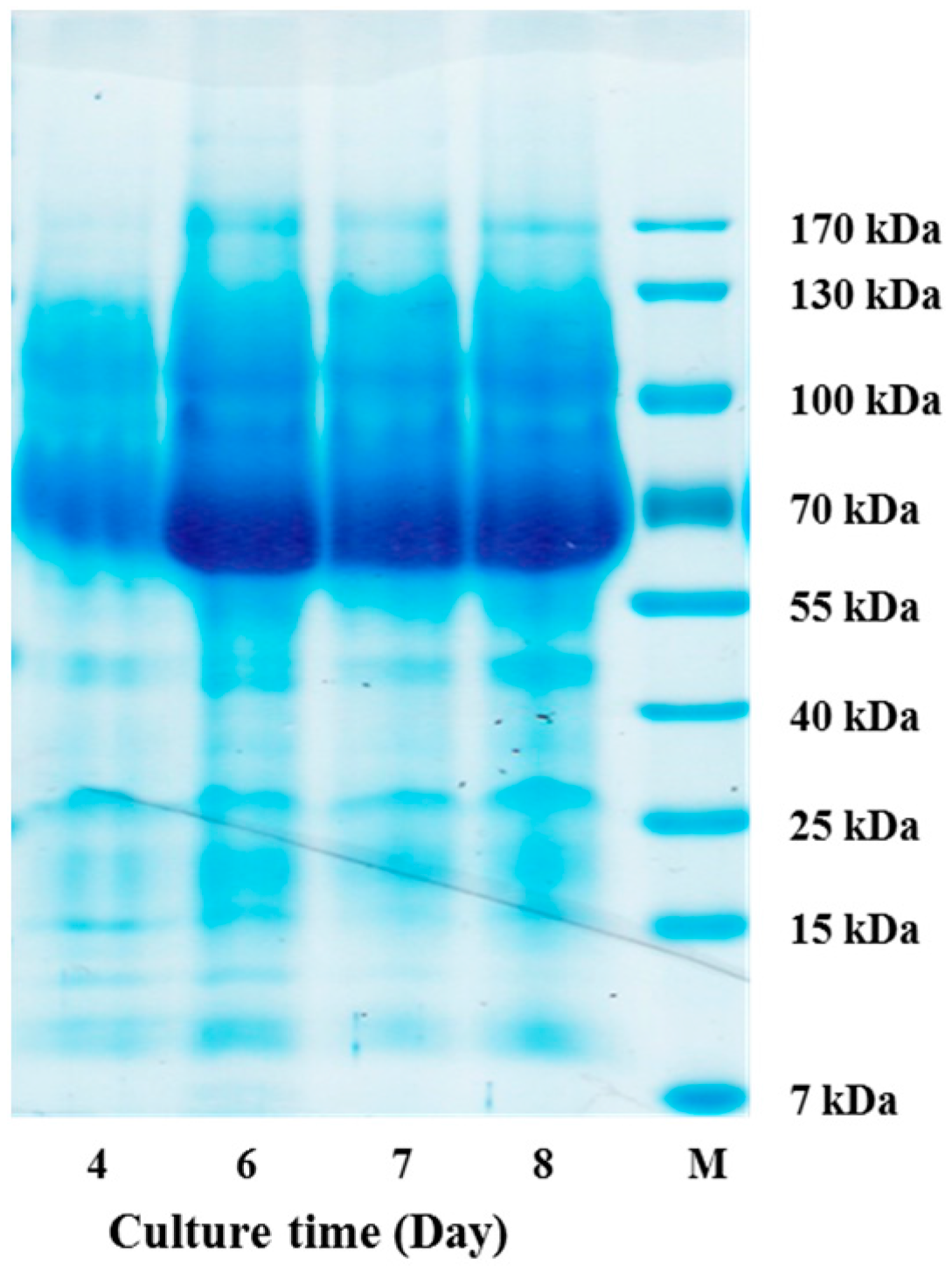
3.6. Analysis of Crude Enzyme from the Culture of Aspergillus niger
3.7. Synthesis of Oligomeric Anthocyanin Using Various Enzymes
3.8. Isolation of Oligomeric Anthocyanin and Analysis
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Peterson, J.; Dwyer, J. Flavonoids: Dietary occurrence and biochemical activity. Nutr. Res. 1998, 18, 1995–2018. [Google Scholar] [CrossRef]
- Delgado-Vargas, F.; Jiménez, A.R.; Paredes-López, O. Natural pigments: Carotenoids, anthocyanins, and betalains—Characteristics, biosynthesis, processing, and stability. Crit. Rev. Food Sci. Nutr. 2000, 40, 173–289. [Google Scholar] [CrossRef] [PubMed]
- Glover, B.J.; Martin, C. Anthocyanins. Curr. Biol. 2012, 22, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Scotter, M.J. Methods for the determination of European Union-permitted added natural colours in foods: A review. Food Addit. Contam. Part A 2011, 28, 527–596. [Google Scholar] [CrossRef] [PubMed]
- Kovačević, D.B.; Putnik, P.; Dragović-Uzelac, V.; Vahčić, N.; Babojelić, M.S.; Levaj, B. Influences of organically and conventionally grown strawberry cultivars on anthocyanins content and color in purees and low-sugar jams. Food Chem. 2015, 181, 94–100. [Google Scholar]
- Kovačević, D.B.; Putnik, P.; Dragović-Uzelac, V.; Pedisić, S.; Jambrak, A.R.; Herceg, Z. Effects of cold atmospheric gas phase plasma on anthocyanins and color in pomegranate juice. Food Chem. 2016, 190, 317–323. [Google Scholar]
- Chou, P.H.; Matsui, S.; Misaki, K.; Matsuda, T. Isolation and identification of xenobiotic aryl hydrocarbon receptor ligands in dyeing wastewater. Environ. Sci. Technol. 2007, 41, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Bridle, P.; Timberlake, C. Timberlake, Anthocyanins as natural food coloursselected aspects. Food Chem. 1997, 58, 103–109. [Google Scholar] [CrossRef]
- Kong, J.M.; Chia, L.S.; Goh, N.K.; Chia, T.F.; Brouillard, R. Analysis and biological activities of anthocyanins. Phytochemistry 2003, 64, 923–933. [Google Scholar] [CrossRef]
- Lila, M.A. Anthocyanins and human health: An in vitro investigative approach. Biomed. Res. Int. 2004, 2004, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.N.; Chu, S.C.; Chiou, H.L.; Kuo, W.H.; Chiang, C.L.; Hsieh, Y.S. Mulberry anthocyanins, cyanidin 3-rutinoside and cyanidin 3-glucoside, exhibited an inhibitory effect on the migration and invasion of a human lung cancer cell line. Cancer Lett. 2006, 235, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.X. Potential mechanisms of cancer chemoprevention by anthocyanins. Curr. Mol. Med. 2003, 3, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Lee, H.K.; Kim, C.Y.; Hong, Y.J.; Choe, C.M.; You, T.W.; Seong, G.J. Purified high-dose anthocyanoside oligomer administration improves nocturnal vision and clinical symptoms in myopia subjects. Br. J. Nutr. 2005, 93, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Günes, O.; Yüceer, Y.K. Biosynthesis of eight-carbon volatiles from tomato and pepper pomaces by fungi: Trichoderma atroviride and Aspergillus sojae. J. Biosci. Bioeng. 2017, in press. [Google Scholar]
- Lee, S.M.; Lee, S.R.; Singh, D.; Oh, J.Y.; Jeon, E.J.; Ryu, H.S.; Lee, D.W.; Kim, B.S.; Lee, C.H. Comparative evaluation of microbial diversity and metabolite profiles in doenjang, a fermented soybean paste, during the two different industrial manufacturing processes. Food Chem. 2017, 221, 1578–1586. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Chen, J.; Li, H.; Sun, F.; Li, Y.; Shi, G. Pellet-dispersion strategy to simplify the seed cultivation of Aspergillus niger and optimize citric acid production. Bioprocess. Biosyst. Eng. 2017, 40, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Paulina, L.; José Manuel, S.; Armando, V.; José Manuel, D.; Isabel, B. Ultrasounds pretreatment of olive pomace to improve xylanase and cellulase production by solid-state fermentation. Bioresour. Technol. 2016, 214, 737–746. [Google Scholar] [Green Version]
- Colla, E.; Santos, L.O.; Deamici, K.; Magagnin, G.; Vendruscolo, M.; Costa, J.A.V. Simultaneous production of Amyloglucosidase and Exo-Polygalacturonase by Aspergillus niger in a rotating Drum reactor. Appl. Biochem. Biotechnol. 2017, 181, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Steaphane, V.; Emmanuelle, M.; Vea Ronique, C.; George, S.; Yoji, H. Mass spectrometric evidence for the existence of oligomeric anthocyanins in grape skins. J. Agric. Food Chem. 2004, 52, 7144–7151. [Google Scholar]
- Hwang, J.W.; Kim, E.K.; Lee, S.J.; Kim, Y.S.; Moon, S.H.; Jeon, B.T.; Sung, S.H.; Kim, E.T.; Park, P.J. Antioxidant activity and protective effect of anthocyanin oligomers on H2O2 triggered G2/M arrest in retinal cells. J. Agric. Food Chem. 2012, 60, 4282–4288. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.S.; Lam, C.W.K.; Chan, M.H.M.; Cheung, R.C.K.; Law, L.K.; Lit, L.C.W.; Ng, K.F.; Suen, M.W.M.; Tai, H.L. Electrospray Ionisation Mass Spectrometry: Principles and Clinical Applications. Clin. Biochem. Rev. 2003, 24, 3–12. [Google Scholar] [PubMed]
- Liu, Y.; Feng, S.; Song, L.; He, G.; Chen, M.; Huang, D. Secondary Metabolites in Durian Seeds: Oligomeric Proanthocyanidins. Molecules 2013, 18, 14172–14185. [Google Scholar] [CrossRef] [PubMed]
- Seiya, C. A historical perspective for the Catalytic reaction mechanism of Glycosidase; So as to bring about breakthrough in Confusing Situation. Biosci. Biotechnol. Biochem. 2012, 76, 215–231. [Google Scholar]
- Sample Availability: Samples of the compounds are not available.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermentation | Monomeric Anthocyanin (g) | Oligomeric Anthocyanin (g) | Yield (%) |
|---|---|---|---|
| Aspergillus niger | 10.1254 | 7.9835 | 78.27 ± 1.99 a |
| 5.0354 | 4.0235 | ||
| 1.0024 | 0.7624 | ||
| Crude enzyme | 10.0038 | 8.7685 | 87.93 ± 0.36 b |
| 2.1539 | 1.9025 | ||
| 1.0657 | 0.9357 | ||
| Glucosidase | 10.0387 | 8.5168 | 85.11 ± 0.45 b |
| 5.1348 | 4.3578 | ||
| 1.0222 | 0.8753 |
| Gene I.D. | Protein Name | Probability | Molecular Weight |
|---|---|---|---|
| gi|224027 | Glucoamylase G1 | 627 | 65,448 |
| gi|134081727 | Unnamed protein product [Aspergillus niger] | 274 | 75,190 |
| gi|765328 | Acid phosphatase, orthophosphoric monoester phosphohydrolase, APase (EC 3.1.3.2) [Aspergillus ficuum, NRRL 3135, Peptide, 583 aa] | 265 | 64,211 |
| gi|257187 | α-Glucosidase P2 subunit, ANP P2 subunit {EC 3.2.1.20} [Aspergillus niger, Peptide, 719 aa] | 181 | 79,656 |
| gi|2344 | Preproglucoamylase G2 [Aspergillus niger] | 531 | 56,695 |
| gi|145242978 | Hypothetical protein ANI_1_1546094 [Aspergillus niger CBS 513.88] | 351 | 59,208 |
| gi|145231236 | Phospholipase C PLC-C [Aspergillus niger CBS 513.88] | 410 | 49,652 |
| gi|145235505 | Serine carboxypeptidase [Aspergillus niger CBS 513.88] | 297 | 62,560 |
| gi|145252338 | Phosphatidylglycerol specific phospholipase [Aspergillus niger CBS 513.88] | 261 | 53,895 |
| gi|4185610 | Phytase [Aspergillus niger] | 218 | 50,997 |
| gi|145241119 | 3-Phytase B [Aspergillus niger CBS 513.88] | 256 | 52,453 |
| gi|145241490 | 1,3-β-Glucanosyltransferase gel3 [Aspergillus niger CBS 513.88] | 161 | 56,721 |
| gi|83655609 | Acid phosphatase [Aspergillus niger] | 142 | 52,725 |
| gi|145242970 | Hypothetical protein ANI_1_1540094 [Aspergillus niger CBS 513.88] | 128 | 45,753 |
| gi|145256696 | Protein ecm33 [Aspergillus niger CBS 513.88] | 125 | 41,026 |
| gi|317026828 | Serine-type carboxypeptidase F [Aspergillus niger CBS 513.88] | 118 | 57,756 |
| gi|145248273 | Polyamine oxidase [Aspergillus niger CBS 513.88] | 110 | 58,728 |
| gi|145248205 | Aspartic-type endopeptidase opsB [Aspergillus niger CBS 513.88] | 104 | 50,958 |
| gi|145234270 | Glutaminase GtaA [Aspergillus niger CBS 513.88] | 99 | 75,470 |
| gi|350633205 | Hypothetical protein ASPNIDRAFT_55058 [Aspergillus niger ATCC 1015] | 87 | 22,487 |
| gi|350631594 | Hypothetical protein ASPNIDRAFT_53033 [Aspergillus niger ATCC 1015] | 63 | 57,162 |
| gi|145235707 | FAD binding domain protein [Aspergillus niger CBS 513.88] | 59 | 61,292 |
| gi|145233743 | α-Galactosidase B [Aspergillus niger CBS 513.88] | 392 | 48,796 |
| gi|317031802 | Histidine acid phosphatase [Aspergillus niger CBS 513.88] | 153 | 53,047 |
| gi|317025164 | Aspartic endopeptidase (AP1) [Aspergillus niger CBS 513.88] | 483 | 46,701 |
| gi|145242664 | Sulphydryl oxidase [Aspergillus niger CBS 513.88] | 264 | 43,471 |
| gi|74626383 | RecName: Probable α-galactosidase B; AltName: Melibiase B; Flags: Precursor | 175 | 48,753 |
| gi|134083538 | Unnamed protein product [Aspergillus niger] | 173 | 45,226 |
| gi|400801 | RecName: Pectin lyase A; Short=PLA; AltName: Full-Pectinlyase II; Short=PLII; Flags: Precursor | 135 | 39,830 |
| gi|145235303 | Hypothetical protein ANI_1_496034 [Aspergillus niger CBS 513.88] | 103 | 52,301 |
| gi|134055991 | Unnamed protein product [Aspergillus niger] | 85 | 41,620 |
| gi|134076313 | Unnamed protein product [Aspergillus niger] | 85 | 45,581 |
| gi|145251519 | Phosphoglycerate mutase family protein [Aspergillus niger CBS 513.88] | 79 | 19,282 |
| gi|350633205 | Hypothetical protein ASPNIDRAFT_55058 [Aspergillus niger ATCC 1015] | 73 | 22,487 |
| gi|145232359 | Endopolygalacturonase C [Aspergillus niger CBS 513.88] | 241 | 37,796 |
| gi|145235523 | Glucan endo-1,3-β-glucosidase eglC [Aspergillus niger CBS 513.88] | 129 | 46,778 |
| gi|145230419 | Glycosidase crf1 [Aspergillus niger CBS 513.88] | 107 | 39,862 |
| gi|129935 | RecName: Full=Endopolygalacturonase II; Short=EPG-II; AltName: Full=Pectinase 2; AltName: Full=PolygalacturonaseII; Short=PG-II; AltName: Full=Polygalacturonase X2; Flags: Precursor | 89 | 37,489 |
| gi|133176 | RecName: Full=Ribonuclease M; Short=RNase M | 89 | 26,590 |
| gi|134055750 | Unnamed protein product [Aspergillus niger] | 84 | 27,072 |
| gi|145229151 | Endo-1,3(4)-β-glucanase [Aspergillus niger CBS 513.88] | 83 | 46,311 |
| gi|134075575 | Hypothetical protein An07g00170 [Aspergillus niger] | 69 | 90,993 |
| gi|134083538 | Unnamed protein product [Aspergillus niger] | 67 | 45,226 |
| gi|145252266 | GPI anchored cell wall protein [Aspergillus niger CBS 513.88] | 64 | 19,022 |
| gi|83638302 | Xylanase [Aspergillus phoenicis] | 117 | 10,944 |
| gi|350633205 | Hypothetical protein ASPNIDRAFT_55058 [Aspergillus niger ATCC 1015] | 92 | 22,487 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, J.-W.; Natarajan, S.B.; Kim, Y.-S.; Kim, E.-K.; Lee, J.W.; Moon, S.-H.; Jeon, B.-T.; Park, P.-J. Biosynthesis of Oligomeric Anthocyanins from Grape Skin Extracts. Molecules 2017, 22, 497. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22030497
Hwang J-W, Natarajan SB, Kim Y-S, Kim E-K, Lee JW, Moon S-H, Jeon B-T, Park P-J. Biosynthesis of Oligomeric Anthocyanins from Grape Skin Extracts. Molecules. 2017; 22(3):497. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22030497
Chicago/Turabian StyleHwang, Jin-Woo, Sithranga Boopathy Natarajan, Yon-Suk Kim, Eun-Kyung Kim, Jae Woong Lee, Sang-Ho Moon, Byong-Tae Jeon, and Pyo-Jam Park. 2017. "Biosynthesis of Oligomeric Anthocyanins from Grape Skin Extracts" Molecules 22, no. 3: 497. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22030497