
Multiple UDP-Glucuronosyltransferase and Sulfotransferase Enzymes are Responsible for the Metabolism of Verproside in Human Liver Preparations



Abstract
:1. Introduction
2. Results
2.1. In Vitro Metabolic Profile of Verproside in Human Hepatocytes
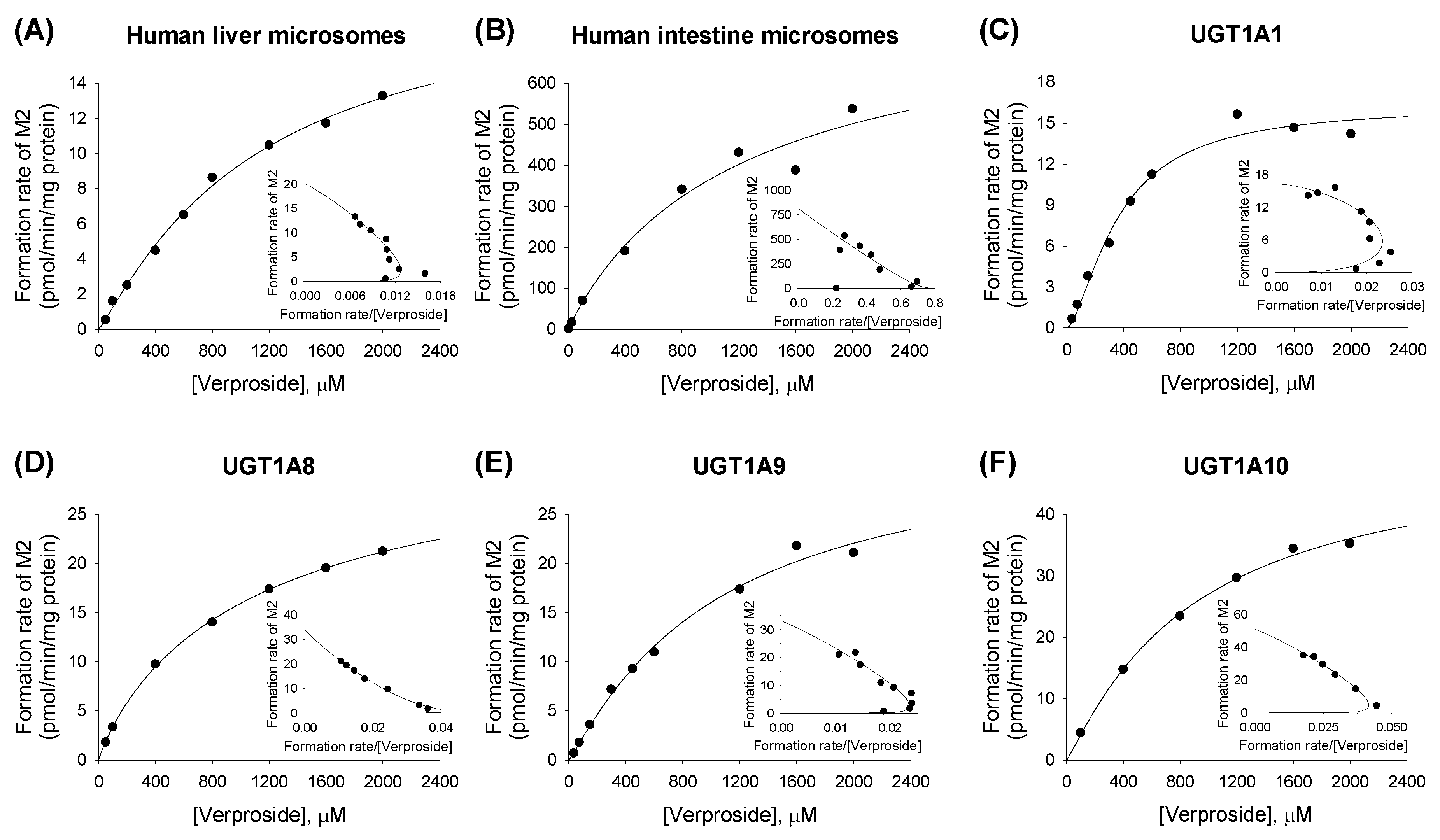
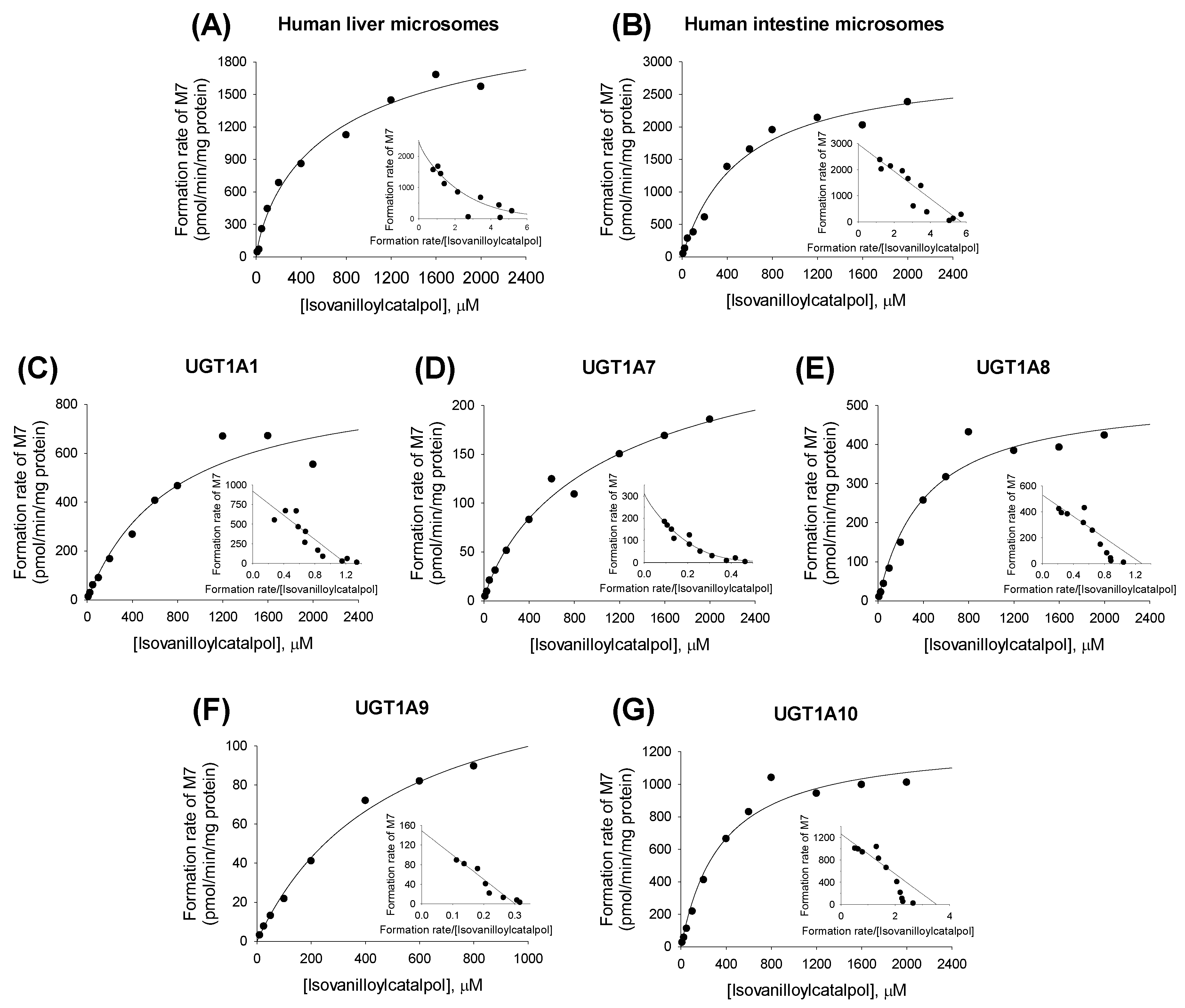
2.2. Characterization of Human UGT Enzymes Responsible for the Glucuronidation of Verproside, Isovanilloylcatalpol, and Picroside II
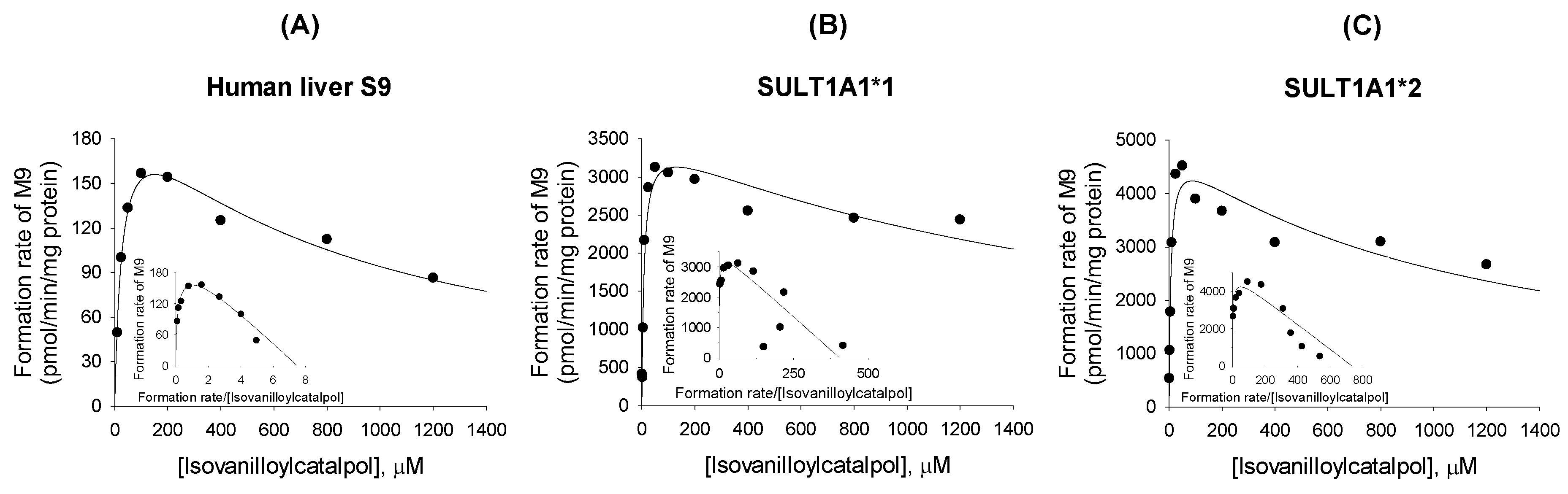
2.3. Characterization of Human SULT Enzymes Responsible for Sulfation of Verproside, Isovanilloylcatalpol, and Picroside II
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. In Vitro Metabolism of Verproside in Cryopreserved Human Hepatocytes
4.3. Characterization of Human UGTs Responsible for the Glucuronidation of Verproside and its Metabolites, Picroside II, and Isovanilloylcatalpol
4.4. Characterization of Human SULTs Responsible for the Sulfation of Verproside and its Metabolites, Picroside II, and Isovanilloylcatalpol
4.5. LC-HRMS Analysis of Verproside and its Metabolites
4.6. Data Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Harput, U.S.; Saracoglu, I.; Nagatsu, A.; Ogihara, Y. Iridoid glucosides from Veronica hederifolia. Chem. Pharm. Bull. 2002, 50, 1106–1108. [Google Scholar] [CrossRef] [PubMed]
- Saracoglu, I.; Oztunca, F.H.; Nagatsu, A.; Harput, U.S. Iridoid content and biological activities of Veronica cuneifolia subsp. cuneifolia and V. cymbalaria. Pharm. Biol. 2011, 49, 1150–1157. [Google Scholar] [CrossRef] [PubMed]
- Saracoglu, I.; Harput, U.S. In vitro cytotoxic activity and structure activity relationships of iridoid glucosides derived from Veronica species. Phytother. Res. 2012, 26, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Lu, Q.; Shu, Y.; Sun, Y.; Chen, F.; Tang, L. Iridoid glycosides fraction isolated from Veronica ciliata fisch. Protects against acetaminophen-induced liver injury in mice. Evid. Based Complement. Alternat. Med. 2017, 2017, 6106572. [Google Scholar] [CrossRef] [PubMed]
- Recio, M.C.; Giner, R.M.; Manez, S.; Rios, J.L. Structural considerations on the iridoids as anti-inflammatory agents. Planta Med. 1994, 60, 232–234. [Google Scholar] [CrossRef] [PubMed]
- Kupeli, E.; Harput, U.S.; Varel, M.; Yesilada, E.; Saracoglu, I. Bioassay-guided isolation of iridoid glucosides with antinociceptive and anti-inflammatory activities from Veronica anagallis-aquatica L. J. Ethnopharmacol. 2005, 102, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Dinda, B.; Debnath, S.; Harigaya, Y. Naturally occurring iridoids. A review, part 1. Chem. Pharm. Bull. 2007, 55, 159–222. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.H.; Kim, H.J.; Lee, K.H.; Kang, S.C.; Zee, O.P. Antioxidative iridoid glycosides and phenolic compounds from Veronica peregrina. Arch. Pharm. Res. 2009, 32, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Sun, Y.; Shu, Y.; Tan, S.; Yin, L.; Guo, Y.; Tang, L. HSCCC separation of the two iridoid glycosides and three phenolic compounds from Veronica ciliata and their in vitro antioxidant and anti-hepatocarcinoma activities. Molecules 2016, 21, E1234. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-R.; Young Lee, M.; Ahn, K.; Park, B.-Y.; Kwon, O.-K.; Joung, H.; Lee, J.; Kim, D.-Y.; Lee, S.; Kim, J.H.; et al. Suppressive effect of verproside isolated from Pseudolysimachion longifolium on airway inflammation in a mouse model of allergic asthma. Int. Immunopharmacol. 2006, 6, 978–986. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.U.; Sung, M.H.; Ryu, H.W.; Lee, J.; Kim, H.-S.; In, H.J.; Ahn, K.-S.; Lee, H.-J.; Lee, H.-K.; Shin, D.-H.; et al. Verproside inhibits TNF-α-induced MUC5AC expression through suppression of the TNF-α/NF-κB pathway in human airway epithelial cells. Cytokine 2016, 77, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Lee, H.S.; Oh, S.-R.; Lee, H.-K.; Lee, H.S. Pharmacokinetics of verproside after intravenous and oral administration in rats. Arch. Pharm. Res. 2009, 32, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Hwang, D.-K.; Jeong, H.-U.; Ji, H.; Oh, S.-R.; Lee, Y.; Yoo, J.; Shin, D.; Lee, H. In vitro and in vivo metabolism of verproside in rats. Molecules 2012, 17, 11990–12002. [Google Scholar] [CrossRef] [PubMed]
- Cerny, M.A. Prevalence of non–cytochrome P450–mediated metabolism in Food and Drug Administration–approved oral and intravenous drugs: 2006–2015. Drug Metab. Dispos. 2016, 44, 1246–1252. [Google Scholar] [CrossRef] [PubMed]
- Foti, R.S.; Dalvie, D.K. Cytochrome P450 and non–cytochrome P450 oxidative metabolism: Contributions to the pharmacokinetics, safety, and efficacy of xenobiotics. Drug Metab. Dispos. 2016, 44, 1229–1245. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.U.; Kim, J.H.; Kong, T.Y.; Choi, W.G.; Lee, H.S. Comparative metabolism of honokiol in mouse, rat, dog, monkey, and human hepatocytes. Arch. Pharm. Res. 2016, 39, 516–530. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zheng, Y.; Fu, F.; Ji, H.; Chen, X.; Zhao, Y.; Zhao, D.; Li, N.; Zhang, L. Assessment of UDP-glucuronosyltransferase catalyzed formation of picroside II glucuronide in microsomes of different species and recombinant UGTs. Xenobiotica 2011, 41, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Strassburg, C.P.; Manns, M.P.; Tukey, R.H. Expression of the UDP-glucuronosyltransferase 1A locus in human colon. Identification and characterization of the novel extrahepatic UGT1A8. J. Biol. Chem. 1998, 273, 8719–8726. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Radominska-Pandya, A.; Tephly, T.R. Cloning and expression of human UDP-glucuronosyltransferase (UGT) 1A8. Arch. Biochem. Biophys. 1998, 356, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Miners, J.O.; Knights, K.M.; Houston, J.B.; Mackenzie, P.I. In vitro-in vivo correlation for drugs and other compounds eliminated by glucuronidation in humans: Pitfalls and promises. Biochem. Pharmacol. 2006, 71, 1531–1539. [Google Scholar] [CrossRef] [PubMed]
- Rowland, A.; Miners, J.O.; Mackenzie, P.I. The UDP-glucuronosyltransferases: Their role in drug metabolism and detoxification. Int. J. Biochem. Cell Biol. 2013, 45, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Riches, Z.; Stanley, E.L.; Bloomer, J.C.; Coughtrie, M.W.H. Quantitative evaluation of the expression and activity of five major sulfotransferases (SULTs) in human tissues: The SULT “pie”. Drug Metab. Dispos. 2009, 37, 2255–2261. [Google Scholar] [CrossRef] [PubMed]
- Runge-Morris, M.; Kocarek, T.A. Expression of the sulfotransferase 1C family: Implications for xenobiotic toxicity. Drug Metab. Rev. 2013, 45, 450–459. [Google Scholar] [CrossRef] [PubMed]
- O'Mara, T.A.; Ferguson, K.; Fahey, P.; Marquart, L.; Yang, H.P.; Lissowska, J.; Chanock, S.; Garcia-Closas, M.; Thompson, D.; Dunning, A.M.; et al. CHEK2, MGMT, SULT1E1 and SULT1A1 polymorphisms and endometrial cancer risk. Twin Res. Hum. Genet. 2011, 14, 328–332. [Google Scholar] [CrossRef] [PubMed]
- James, M.O.; Ambadapadi, S. Interactions of cytosolic sulfotransferases with xenobiotics. Drug Metab. Rev. 2013, 45, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Coughtrie, M.W.; Johnston, L.E. Interactions between dietary chemicals and human sulfotransferases-molecular mechanisms and clinical significance. Drug Metab. Dispos. 2001, 29, 522–528. [Google Scholar] [PubMed]
- Zhou, T.; Chen, Y.; Huang, C.; Chen, G. Caffeine induction of sulfotransferases in rat liver and intestine. J. Appl. Toxicol. 2012, 32, 804–809. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Compound Name | Retention Time (min) | Detected ([M − H]−) (m/z) | Error (ppm) | Product Ions (m/z) |
|---|---|---|---|---|---|
| Verproside | 7.25 | 497.12939 | −1.5 | 355.0772, 221.0450, 153.0184 | |
| M1 | Verproside glucuronide | 2.99 | 673.16138 | −0.9 | 497.1301, 335.0773, 221.0450, 175.0241 |
| M2 | Verproside glucuronide | 6.11 | 673.16138 | −0.9 | 497.1301, 335.0773, 221.0450, 175.0241 |
| M3 | Verproside sulfate | 7.36 | 577.08594 | −1.6 | 497.1301, 335.0773, 221.0451, 153.0184 |
| M4 | Picroside II | 11.70 | 511.14484 | −1.7 | 349.0928, 235.0608, 167.0341 |
| M5 | Isovanilloylcatalpol | 12.32 | 511.14484 | −1.7 | 349.0928, 235.0608, 167.0341 |
| M6 | Picroside II glucuronide | 4.87 | 687.17657 | −1.7 | 511.1458, 349.0930, 235.0609, 167.0342 |
| M7 | Isovanilloylcatalpol glucuronide | 7.98 | 687.17657 | −1.7 | 511.1458, 349.0930, 235.0609, 167.0342 |
| M8 | Picroside II sulfate | 8.16 | 591.10150 | −1.5 | 511.1457, 349.0927, 235.0609, 167.0341 |
| M9 | Isovanilloylcatalpol sulfate | 9.44 | 591.10150 | −1.5 | 511.1457, 349.0927, 235.0609, 167.0341 |
| Enzymes | M1 Formation | M2 Formation | ||||||
|---|---|---|---|---|---|---|---|---|
| Km (μM) | Vmax | n | Clint | Km (μM) | Vmax | n | Clint | |
| Human Liver Microsomes | 791.3 (174.9) | 22.6 (2.5) | 1.2 | 0.029 | 1109.9 (265.9) | 20.0 (2.5) | 1.1 | 0.018 |
| Human Intestinal Microsomes | 1251.8 (1608.4) | 159.4 (100.3) | 1.1 | 0.126 | 1215.2 (1460.9) | 810.2 (449.0) | 1.0 | 0.667 |
| UGT1A1 | 316.5 (54.2) | 32.4 (2.6) | 1.4 | 0.102 | 363.7 (53.1) | 16.3 (1.2) | 1.5 | 0.045 |
| UGT1A8 | 492.3 (30.1) | 77.4 (1.9) | 1.1 | 0.157 | 1158.7 (159.4) | 34.2 (2.0) | 0.9 | 0.030 |
| UGT1A9 | 2272.1 (2873.8) | 43.2 (31.4) | 1.1 | 0.019 | 1052.5 (440.9) | 33.0 (6.8) | 1.1 | 0.031 |
| UGT1A10 | 802.3 (266.4) | 280.5 (54.6) | 1.4 | 0.350 | 894.1 (229.5) | 50.9 (6.4) | 1.1 | 0.057 |
| Enzymes | Km (μM) | Vmax | n | Clint |
|---|---|---|---|---|
| Human liver microsomes | 818.1 (591.4) | 2470.7 (633.1) | 0.7847 | 3.02 |
| Human intestinal microsomes | 520.9 (96.6) | 2974.3 (201.4) | - | 5.71 |
| UGT1A1 | 768.8 (250.7) | 919.6 (128.0) | - | 1.20 |
| UGT1A7 | 1312.6 (939.6) | 312.9 (92.3) | 0.8375 | 0.238 |
| UGT1A8 | 412.4 (98.6) | 529.3 (42.2) | - | 1.28 |
| UGT1A9 | 495.7 (77.9) | 149.2 (11.7) | - | 0.301 |
| UGT1A10 | 358.5 (74.0) | 1265.6 (82.5) | - | 3.53 |
| Enzymes | M3 Formation | M9 Formation | ||||||
|---|---|---|---|---|---|---|---|---|
| Km (μM) | Vmax | n | Clint | Km (μM) | Vmax | Ki (μM) | Clint | |
| Human liver S9 | 1.21 (0.19) | 203.3 (19.3) | 1.3 | 168.0 | 28.8(5.3) | 215.6 (15.4) | 792.4 (147.5) | 27.5 |
| SULT1A1*1 | 0.69 (0.23) | 9019.9 (1457.6) | 1.2 | 13,131 | 8.8 (2.3) | 3550.7 (268.7) | 1938.6(726.7) | 380.8 |
| SULT1A1*2 | 1.10 (0.66) | 27,610.0 (7798.5) | 1.0 | 25,039 | 6.7 (1.9) | 4889.2 (395.7) | 1127.5 (365.0) | 729.7 |
| Enzymes | Km (μM) | Vmax | Ki (μM) | Clint |
|---|---|---|---|---|
| Human liver S9 | 45.1 (22.0) | 472.2 (101.9) | 799.8 (417.1) | 10.5 |
| SULT1A1*1 | 9.0 (2.6) | 10,100 (921) | 844.4 (263.0) | 1122 |
| SULT1A1*2 | 10.8 (3.8) | 13,490 (1710) | 468.9 (163.9) | 1249 |
| SULT1A2 | 54.0 (13.4) | 6134.5 (688.0) | 2859.3 (1854.2) | 113.6 |
| SULT1A3 | 1271.7 (423.6) | 82,370 (21746) | 1265.9 (551.3) | 64.8 |
| SULT1C4 | 553.1 (33.4) | 61,140 (2036) | - | 110.5 |
| SULT1E1 | 12.9 (3.4) | 36,250 (3274) | 995.3 (356.9) | 2810 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-H.; Hwang, D.-K.; Moon, J.-Y.; Lee, Y.; Yoo, J.S.; Shin, D.H.; Lee, H.S. Multiple UDP-Glucuronosyltransferase and Sulfotransferase Enzymes are Responsible for the Metabolism of Verproside in Human Liver Preparations. Molecules 2017, 22, 670. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22040670
Kim J-H, Hwang D-K, Moon J-Y, Lee Y, Yoo JS, Shin DH, Lee HS. Multiple UDP-Glucuronosyltransferase and Sulfotransferase Enzymes are Responsible for the Metabolism of Verproside in Human Liver Preparations. Molecules. 2017; 22(4):670. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22040670
Chicago/Turabian StyleKim, Ju-Hyun, Deok-Kyu Hwang, Ju-Yeon Moon, Yongnam Lee, Ji Seok Yoo, Dae Hee Shin, and Hye Suk Lee. 2017. "Multiple UDP-Glucuronosyltransferase and Sulfotransferase Enzymes are Responsible for the Metabolism of Verproside in Human Liver Preparations" Molecules 22, no. 4: 670. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22040670