Evaluation of the Nutritional Quality of Chinese Kale (Brassica alboglabra Bailey) Using UHPLC-Quadrupole-Orbitrap MS/MS-Based Metabolomics


Abstract
:1. Introduction
2. Results and Discussion
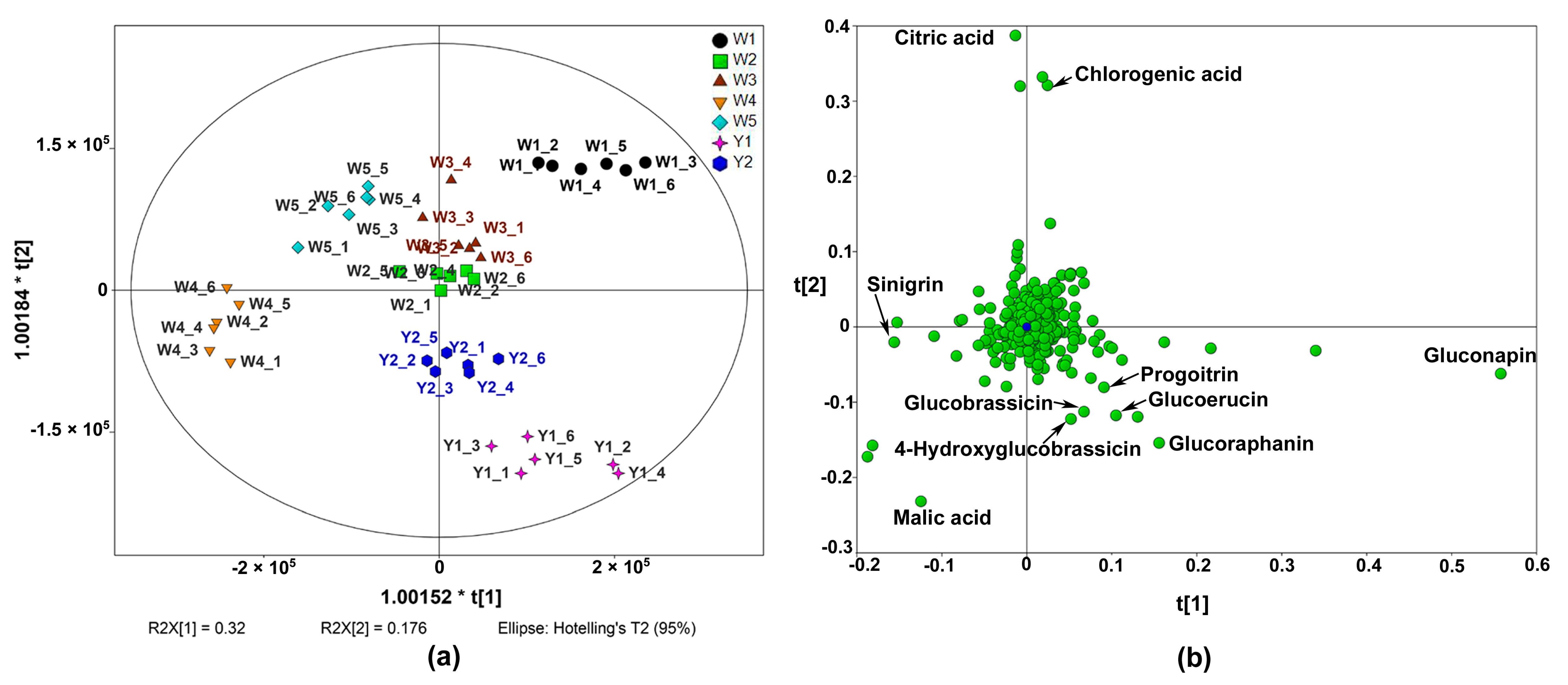
2.1. Metabolic Profiles of Different Chinese Kale Cultivars
2.2. Nutritive Compounds in Different Chinese Kale Cultivars
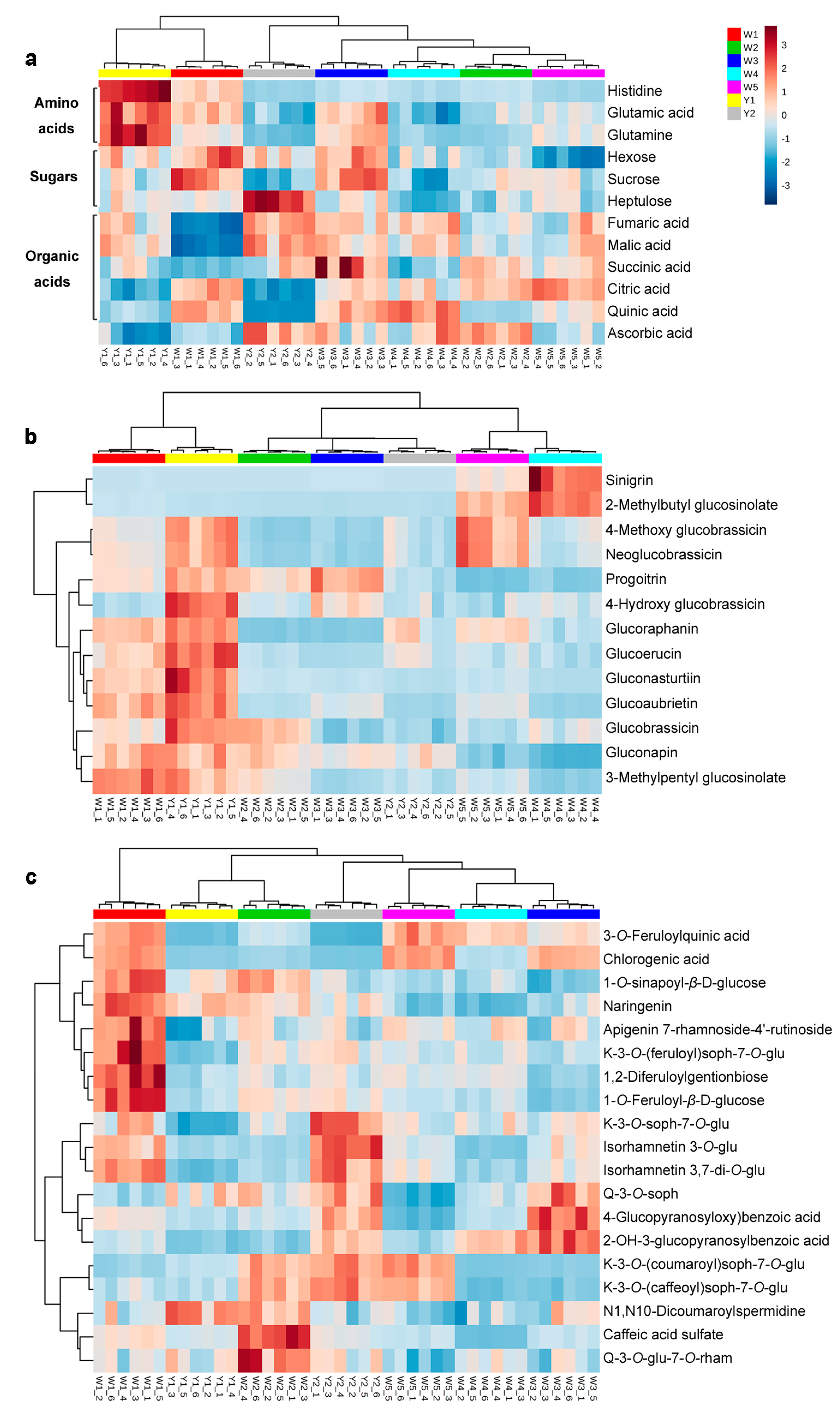
2.2.1. Primary Metabolites in Different Chinese Kale Cultivars
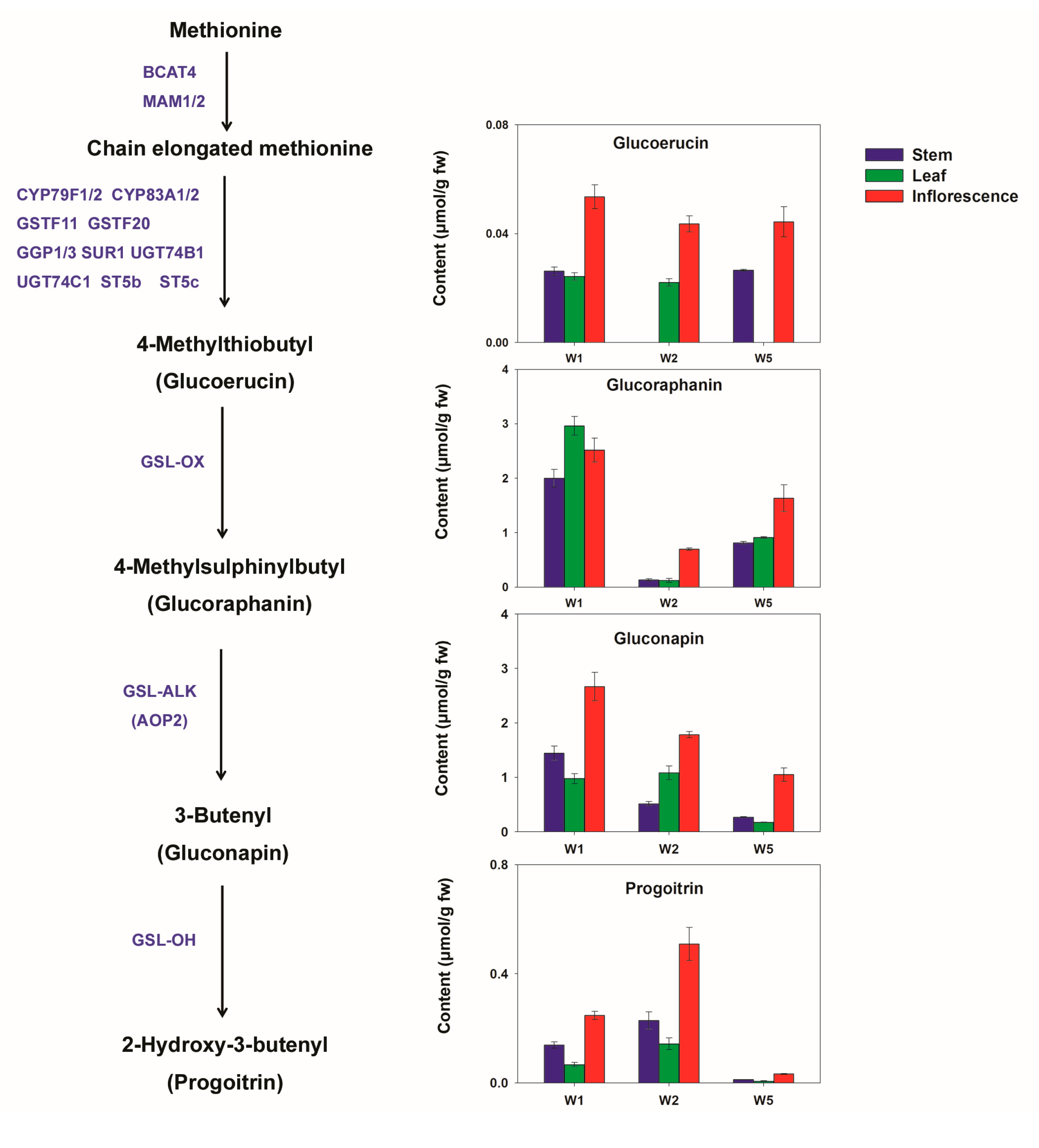
2.2.2. Glucosinolates in Different Chinese Kale Cultivars
2.2.3. Phenolic Acids and Flavonoids in Different Chinese Kale Cultivars
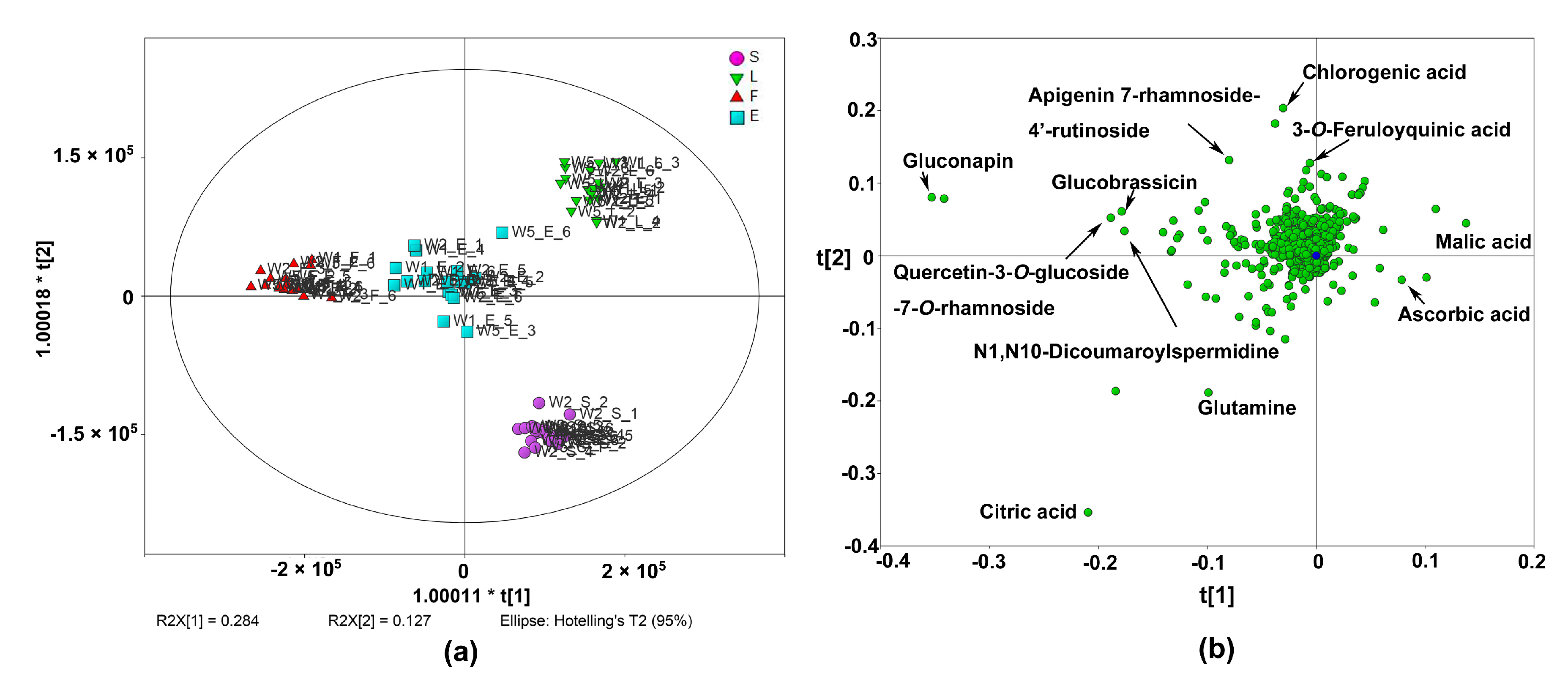
2.3. Metabolic Profiles of Different Edible Parts of Chinese Kale
2.4. Nutritive Compounds in Different Edible Parts of Chinese Kale
2.4.1. Primary Metabolites in Different Edible Parts
2.4.2. Glucosinolates in Different Edible Parts
2.4.3. Phenolic Acids and Flavonoids in Different Edible Parts
3. Materials and Methods
3.1. Plant Materials and Reagents
3.2. Metabolites Extraction
3.3. Chromatographic and Mass Spectrometric Conditions
3.4. Data Processing and Multivariate Analysis
3.5. Quantitative Analysis of Desulfo-Glucosinolates
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jahangir, M.; Kim, H.K.; Choi, Y.H.; Verpoorte, R. Health-affecting compounds in Brassicaceae. Compr. Rev. Food Sci. Food Saf. 2009, 8, 31–43. [Google Scholar] [CrossRef]
- Sun, B.; Liu, N.; Zhao, Y.; Yan, H.; Wang, Q. Variation of glucosinolates in three edible parts of chinese kale (Brassica alboglabra Bailey) varieties. Food Chem. 2011, 124, 941–947. [Google Scholar] [CrossRef]
- Sun, B.; Yan, H.; Zhang, F.; Wang, Q. Effects of plant hormones on main health-promoting compounds and antioxidant capacity of Chinese kale. Food Res. Int. 2012, 48, 359–366. [Google Scholar] [CrossRef]
- Wolucka, B.A.; Goossens, A.; Inzé, D. Methyl jasmonate stimulates the de novo biosynthesis of vitamin C in plant cell suspensions. J. Exp. Bot. 2005, 56, 2527–2538. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Traka, M.; Mithen, R. Glucosinolates, isothiocyanates and human health. Phytochem. Rev. 2009, 8, 269–282. [Google Scholar] [CrossRef]
- Keck, A.S.; Finley, J.W. Cruciferous vegetables: Cancer protective mechanisms of glucosinolate hydrolysis products and selenium. Integr. Cancer Ther. 2004, 3, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2010, 16, 251–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aires, A.; Rosa, E.; Carvalho, R. Effect of nitrogen and sulfur fertilization on glucosinolates in the leaves and roots of broccoli sprouts (Brassica oleracea var Italica). J. Sci. Food Agric. 2006, 86, 1512–1516. [Google Scholar] [CrossRef]
- Brown, P.D.; Tokuhisa, J.G.; Reichelt, M.; Gershenzon, J. Variation of glucosinolate accumulation among different organs and developmental stages of Arabidopsis thaliana. Phytochemistry 2003, 62, 471–481. [Google Scholar] [CrossRef]
- Ciska, E.; Martyniak-Przybyszewska, B.; Kozlowska, H. Content of glucosinolates in cruciferous vegetables grown at the same site for two years under different climatic conditions. J. Agric. Food Chem. 2000, 48, 2862–2867. [Google Scholar] [CrossRef] [PubMed]
- Farnham, M.W.; Wilson, P.E.; Stephenson, K.K.; Fahey, J.W. Genetic and environmental effects on glucosinolate content and chemoprotective potency of broccoli. Plant Breed. 2004, 123, 60–65. [Google Scholar] [CrossRef]
- Song, L.; Thornalley, P.J. Effect of storage, processing and cooking on glucosinolate content of Brassica vegetables. Food Chem. Toxicol. 2007, 45, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Sun, B.; Miao, H.; Cai, C.; Xu, C.; Wang, Q. Variation of glucosinolates and quinone reductase activity among different varieties of Chinese kale and improvement of glucoraphanin by metabolic engineering. Food Chem. 2015, 168, 321–326. [Google Scholar] [CrossRef] [PubMed]
- De Vos, R.C.; Moco, S.; Lommen, A.; Keurentjes, J.J.; Bino, R.J.; Hall, R.D. Untargeted large-scale plant metabolomics using liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2007, 2, 778–791. [Google Scholar] [CrossRef] [PubMed]
- Maldini, M.; Natella, F.; Baima, S.; Morelli, G.; Scaccini, C.; Langridge, J.; Astarita, G. Untargeted metabolomics reveals predominant alterations in lipid metabolism following light exposure in broccoli sprouts. Int. J. Mol. Sci. 2015, 16, 13678–13691. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.J.; García-Villalba, R.; Garrido, Y.; Gil, M.I.; Tomás-Barberán, F.A. Untargeted metabolomics approach using UPLC-ESI-QTOF-MS to explore the metabolome of fresh-cut iceberg lettuce. Metabolomics 2016, 12, 138. [Google Scholar] [CrossRef]
- Witzel, K.; Neugart, S.; Ruppel, S.; Schreiner, M.; Wiesner, M.; Baldermann, S. Recent progress in the use of ߢomics technologies in brassicaceous vegetables. Front. Plant Sci. 2015, 6, 244. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Sharaf Eldin, M.G.; Kassem, H.; Abou el Fetouh, M. Metabolome classification of Brassica napus L. organs via via UPLC-QTOF-PDA-MS and their anti-oxidant potential. Phytochem. Anal. 2013, 24, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Arasu, M.V.; Jiang, N.; Choi, S.H.; Yong, P.L.; Park, J.T.; Al-Dhabi, N.A.; Kim, S.J. Metabolite profiling of phenolics, anthocyanins and flavonols in cabbage (Brassica oleracea var. Capitata). Ind. Crops Prod. 2014, 60, 8–14. [Google Scholar] [CrossRef]
- Park, S.Y.; Lim, S.H.; Ha, S.H.; Yeo, Y.; Park, W.T.; Kwon, D.Y.; Park, S.U.; Kim, J.K. Metabolite profiling approach reveals the interface of primary and secondary metabolism in colored cauliflowers (Brassica oleracea L. ssp. Botrytis). J. Agric. Food Chem. 2013, 61, 6999–7007. [Google Scholar] [CrossRef] [PubMed]
- Mie, A.; Laursen, K.H.; Aberg, K.M.; Forshed, J.; Lindahl, A.; Thorup-Kristensen, K.; Olsson, M.; Knuthsen, P.; Larsen, E.H.; Husted, S. Discrimination of conventional and organic white cabbage from a long-term field trial study using untargeted LC-MS-based metabolomics. Anal. Bioanal. Chem. 2014, 406, 2885–2897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennig, K.; de Vos, R.C.; Maliepaard, C.; Dekker, M.; Verkerk, R.; Bonnema, G. A metabolomics approach to identify factors influencing glucosinolate thermal degradation rates in Brassica vegetables. Food Chem. 2014, 155, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Millán, L.; Sampedro, M.C.; Sánchez, A.; Delporte, C.; Antwerpen, P.V.; Goicolea, M.A.; Barrio, R.J. Liquid chromatography-quadrupole time of flight tandem mass spectrometry-based targeted metabolomic study for varietal discrimination of grapes according to plant sterols content. J. Chromatogr. A 2016, 1454, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Glauser, G.; Veyrat, N.; Rochat, B.; Wolfender, J.L.; Turlings, T.C. Ultra-high pressure liquid chromatography-mass spectrometry for plant metabolomics: A systematic comparison of high-resolution quadrupole-time-of-flight and single stage Orbitrap mass spectrometers. J. Chromatogr. A 2013, 1292, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, A. Combining UHPLC and high-resolution MS: A viable approach for the analysis of complex samples? TrAC Trends Anal. Chem. 2014, 63, 113–128. [Google Scholar] [CrossRef]
- Sun, J.; Xiao, Z.; Lin, L.; Lester, G.E.; Wang, Q.; Harnly, J.M.; Chen, P. Profiling polyphenols in five Brassica species microgreens UHPLC-PDA-ESI/HRMSn. J. Agric. Food Chem. 2013, 61, 10960–10970. [Google Scholar] [CrossRef] [PubMed]
- Kumaraswamy, K.G.; Kushalappa, A.C.; Choo, T.M.; Dion, Y.; Rioux, S. Mass spectrometry based metabolomics to identify potential biomarkers for resistance in barley against fusarium head blight (Fusarium graminearum). J. Chem. Ecol. 2011, 37, 846–856. [Google Scholar] [CrossRef] [PubMed]
- Bijttebier, S.; Zhani, K.; D'Hondt, E.; Noten, B.; Hermans, N.; Apers, S.; Voorspoels, F. Generic characterization of apolar metabolites in red chili peppers (Capsicum frutescens L.) by Orbitrap mass spectrometry. J. Agric. Food Chem. 2014, 62, 4812–4831. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Li, Q.; Cai, H.; Tu, S.; Cai, B. Investigation of the chemical changes from crude and processed paeoniae radix alba-atractylodis macrocephalae rhizoma herbal pair extracts by using Q Exactive high-performance benchtop quadrupole-Orbitrap LC-MS/MS. Evid. Based Complement. Altern. Med. 2014, 2014, 170959. [Google Scholar] [CrossRef] [PubMed]
- Forcisi, S.; Moritz, F.; Kanawati, B.; Tziotis, D.; Lehmann, R.; Schmittkopplin, P. Liquid chromatography-mass spectrometry in metabolomics research: Mass analyzers in ultra high pressure liquid chromatography coupling. J. Chromatogr. A 2013, 1292, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Halkier, B.A.; Du, L. The biosynthesis of glucosinolates. Trends Plant Sci. 1997, 2, 425–431. [Google Scholar] [CrossRef]
- Padilla, G.; Cartea, M.E.; Velasco, P.; de Haro, A.; Ordás, A. Variation of glucosinolates in vegetable crops of Brassica rapa. Phytochemistry 2007, 68, 536–545. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Milner, J.A. Targets for indole-3-carbinol in cancer prevention. J. Nutr. Biochem. 2005, 16, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Verkerk, R.; Schreiner, M.; Krumbein, A.; Ciska, E.; Holst, B.; Rowland, I.; De Schrijver, R.; Hansen, M.; Gerhäuse, C.; Mithen, R.; et al. Glucosinolates in Brassica vegetables: The influence of the food supply chain on intake, bioavailability and human health. Mol. Nutr. Food Res. 2009, 53, S219–S265. [Google Scholar] [CrossRef] [PubMed]
- Halkier, B.A.; Gershenzon, J. Biology and biochemistry of glucosinolates. Annu. Rev. Plant Biol. 2006, 57, 303–333. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.Z.; Harnly, J.M. Identification of the phenolic components of collard greens, kale, and Chinese broccoli. J. Agric. Food Chem. 2009, 57, 7401–7408. [Google Scholar] [CrossRef] [PubMed]
- Piazzon, A.; Vrhovsek, U.; Masuero, D.; Mattivi, F.; Mandoj, F.; Nardini, M. Antioxidant activity of phenolic acids and their metabolites: synthesis and antioxidant properties of the sulfate derivatives of ferulic and caffeic acids and of the acyl glucuronide of ferulic acid. J. Agric. Food Chem. 2012, 60, 12312–12323. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Lei, J.; Chen, G.; Chen, H.; Cao, B.; Chen, C. De novo transcriptome assembly of Chinese kale and global expression analysis of genes involved in glucosinolate metabolism in multiple tissues. Front. Plant Sci. 2017, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. Metaboanalyst 3.0-making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, L.; Wang, S.; Tao, X.; Wang, Y.; Sun, A.; He, H. Assessment of glucosinolates in Chinese kale by near-infrared spectroscopy. Int. J. Food Prop. 2014, 17, 1668–1679. [Google Scholar] [CrossRef]
Sample Availability: Samples of Chinese kale are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Rt (min) | Compound | Molecular Formula | Experimetal m/z [M − H]− | Theoretical m/z [M − H]− | Accuracy (ppm) | Fragment Ion(s) |
|---|---|---|---|---|---|---|---|
| Amino acids | |||||||
| 1 | 0.71 | Aspartic acid | C4H7NO4 | 132.0302 | 132.0302 | 0.07 | 71.0138, 88.0403, 114.0196, 115.0036 |
| 2 | 0.73 | Asparagine | C4H8N2O3 | 131.0458 | 131.0462 | 2.98 | 113.0351, 95.0254, 70.0292 |
| 3 | 0.80 | Proline | C5H9NO2 | 114.0555 | 114.0560 | 4.60 | 71.0131, 87.0081 |
| 4 | 0.73 | Glutamine | C5H10N2O3 | 145.0614 | 145.0618 | 3.03 | 109.0406, 127.0526, 128.0352, 84.0454 |
| 5 | 0.76 | Glutamic acid | C5H9NO4 | 146.0456 | 146.0459 | 1.76 | 102.0560, 128.0353 |
| 6 | 0.73 | Histidine | C6H9N3O2 | 154.0621 | 154.0622 | 0.50 | 93.0457, 110.0723, 137.0356 |
| Sugars and derivatives | |||||||
| 7 | 0.76 | Glucose/Fructose/Fucose/Mannose/Galactose | C6H12O6 | 179.0558 | 179.0561 | 1.58 | 59.0138, 71.0138, 89.0244, 113.0240 |
| 8 | 0.76 | Sucrose | C12H22O11 | 341.1089 | 341.1089 | 0.03 | 71.0138, 89.0243, 113.0243, 161.0455, 179.0560 |
| 9 | 0.76 | Sedoheptulose/d-Mannoheptulose/d-Glucoheptose | C7H14O7 | 209.0662 | 209.0667 | 2.18 | 85.0288, 87.0081, 129.0188, 159.0296 |
| 10 | 0.68 | d-Glucose-6-phosphate/d-Fructose-6-phosphate/d-Mannose-6-phosphate | C6H13O9P | 259.0229 | 259.0224 | 1.83 | 78.9590, 96.9696, 138.9801, 199.0013 |
| 11 | 0.73 | Glucoheptonic acid | C7H14O8 | 225.0614 | 225.0616 | 0.72 | 179.0561, 59.0132, 89.0246, 119.0346 |
| 12 | 0.98 | 2-O-Sulfo-l-idopyranuronic acid | C6H10O10S | 272.9923 | 272.9922 | 0.43 | 96.9596, 115.0036, 158.0161 |
| Organic acids | |||||||
| 13 | 0.76 | Quinic acid | C7H12O6 | 191.0555 | 191.0561 | 3.05 | 85.0295, 127.0396, 173.0451 |
| 14 | 0.79 | Fumaric acid | C4H4O4 | 115.0031 | 115.0037 | 4.83 | 71.0138 |
| 15 | 0.80 | Malic acid | C4H6O5 | 133.0141 | 133.0142 | 0.97 | 71.0138, 115.0036 |
| 16 | 0.81 | Citric acid | C6H8O7 | 191.0194 | 191.0197 | 1.60 | 87.0087, 111.0088, 129.0193, 173.0091 |
| 17 | 1.29 | Succinic acid | C4H6O4 | 117.0192 | 117.0193 | 0.85 | 73.0295, 99.0087, 117.0193 |
| Vitamins | |||||||
| 18 | 0.8 | Iso/Ascorbic acid | C6H8O6 | 175.0246 | 175.0248 | 1.14 | 59.0138, 71.0138, 87.0082, 115.0036 |
| 19 | 4.0 | Pantothenic acid | C9H17NO5 | 218.1028 | 218.1034 | 2.61 | 88.0397, 146.0821 |
| Glucosinolates | |||||||
| 20 | 0.80 | Glucoraphanin | C12H23NO10S3 | 436.0414 | 436.0411 | 0.68 | 96.9596, 178.0169, 259.0129, 274.9895, 372.0429 |
| 21 | 0.81 | Sinigrin | C10H17NO9S2 | 358.0277 | 358.0272 | 1.43 | 96.9596, 195.0327, 241.0018, 259.0124, 274.9895 |
| 22 | 0.81 | Progoitrin | C11H19NO10S2 | 388.0380 | 388.0377 | 0.66 | 96.9596, 195.0327, 241.0018, 259.0124, 274.9895 |
| 23 | 1.69 | Gluconapin | C11H19NO9S2 | 372.0431 | 372.0428 | 0.78 | 96.9596, 195.0327, 241.0018, 259.0124, 274.9895 |
| 24 | 2.21 | 4-Hydroxyglucobrassicin | C16H20N2O10S2 | 463.0497 | 463.0486 | 2.29 | 96.9596, 195.0327, 241.0018, 259.0124 |
| 25 | 4.31 | Glucoerucin | C12H23NO9S3 | 420.0468 | 420.0462 | 1.43 | 96.9596, 195.0327, 259.0124, 274.9895 |
| 26 | 4.51 | 2-Methylbutyl glucosinolate/3-Methylbutyl glucosinolate | C12H23NO9S2 | 388.0747 | 388.0741 | 1.49 | 96.9596, 195.0327, 301.0601, 259.0124, 274.9895, 343.0698 |
| 27 | 4.56 | Glucobrassicin | C16H20N2O9S2 | 447.0539 | 447.0537 | 0.37 | 96.9596, 259.0124, 274.9895, 241.0018, 205.0433 |
| 28 | 4.57 | Glucoaubrietin | C15H21NO10S2 | 438.0544 | 438.0534 | 2.28 | 96.9596, 195.0327, 241.0018, 259.0124, 274.9901 |
| 29 | 4.87 | 4-Methoxyglucobrassicin | C17H22N2O10S2 | 477.0648 | 477.0643 | 1.06 | 96.9596, 195.0327, 259.0124, 314.0433 |
| 30 | 4.76 | Gluconasturtiin | C15H21NO9S2 | 422.0592 | 422.0585 | 1.74 | 96.9584, 195.0323, 259.0131, 274.9904 |
| 31 | 4.98 | 3-Methylpentyl glucosinolate/4-Methylpentyl glucosinolate | C13H25NO9S2 | 402.0898 | 402.0898 | 0.06 | 96.9596, 195.0327, 241.0018, 259.0124, 274.9895, 315.0759 |
| 32 | 5.22 | Neoglucobrassicin | C17H22N2O10S2 | 477.0644 | 477.0643 | 0.22 | 96.9596, 259.0124, 446.0465 |
| Phenolic acids | |||||||
| 33 | 4.49 | Chlorogenic acid | C16H18O9 | 353.0881 | 353.0878 | 0.87 | 191.0561, 173.0455, 135.0452 |
| 34 | 4.63 | Caffeic acid 3-β-d-glucoside | C15H18O9 | 341.0887 | 341.0878 | 2.65 | 135.0447, 161.0236, 179.0349 |
| 35 | 4.84 | 3-O-Feruloylquinic acid | C17H20O9 | 367.1036 | 367.1034 | 0.49 | 173.0455, 191.0561, 93.0339 |
| 36 | 6.92 | 1,2-Diferuloylgentionbiose | C32H38O17 | 693.2051 | 693.2036 | 2.16 | 175.0389, 193.0497, 217.0500 |
| 37 | 3.23 | 2-Hydroxy-3- β-d-glucopyranosylbenzoic acid | C13H16O9 | 315.0724 | 315.0716 | 2.59 | 152.0113, 153.0191, 108.0202 |
| 38 | 4.35 | 4-(β-d-Glucopyranosyloxy)benzoic acid | C13H16O8 | 299.0774 | 299.0772 | 0.66 | 137.0250, 179.0349, 239.0559, 89.0237 |
| 39 | 4.90 | 1-O-Feruloyl-β-d-glucose | C16H20O9 | 355.1039 | 355.1034 | 1.35 | 175.0399, 193.0506 |
| 40 | 4.95 | Caffeic acid 3-sulfate/Caffeic acid 4-sulfate | C9H8O7S | 258.9921 | 258.9918 | 1.15 | 135.0447, 179.0347 |
| 41 | 4.98 | 1-O-Sinapoyl-β-d-glucose | C17H22O10 | 385.1142 | 385.1140 | 0.50 | 205.0506, 223.0623, 164.0477 |
| 42 | 5.92 | N1,N10-Dicoumaroylspermidine | C25H31N3O4 | 436.2247 | 436.2241 | 1.27 | 119.0502, 273.1610, 316.1667 |
| 43 | 4.50 | 1-O-Caffeoyl-(β-d-glucose-6-O-sulfate) | C15H18O12S | 421.0452 | 421.0446 | 1.42 | 96.9584, 161.0233, 179.0347 |
| 44 | 4.86 | p-Coumaric acid glucoside | C15H18O8 | 325.0933 | 325.0929 | 1.35 | 117.0331, 145.0282, 163.0400 |
| Flavonoids | |||||||
| 45 | 6.30 | Isorhamnetin 3-O-glucoside | C22H22O12 | 477.1046 | 477.1038 | 1.64 | 314.0432, 315.0510, 285.0405 |
| 46 | 4.87 | Isorhamnetin 3,7-di-O-glucoside | C28H32O17 | 639.1590 | 639.1566 | 3.67 | 314.0435, 315.0510, 476.0963, 477.1035 |
| 47 | 4.60 | Isorhamnetin 3-O-sophoroside-7-O-glucoside | C34H42O22 | 801.2120 | 801.2095 | 3.15 | 314.0434, 315.0506, 476.0964, 477.1034, 639.1575 |
| 48 | 5.12 | Quercetin 3-O-sophoroside | C27H30O17 | 625.1420 | 625.1410 | 1.57 | 151.0037, 300.0277, 301.0354, 463.0892 |
| 49 | 5.19 | Quercetin-3-O- glucoside-7-O- rhamnoside | C27H30O16 | 609.1473 | 609.1461 | 2.00 | 283.0250, 284.0328, 285.0406, 446.0860, 447.0930 |
| 50 | 4.72 | Quercetin 3-O-(feruloyl)sophoroside-7-O-glucoside | C43H48O25 | 963.2431 | 963.2412 | 2.00 | 191.03395, 284.0326, 285.0400, 609.1469, 801.1893 |
| 51 | 4.59 | Kaempferol 3-O-sophoroside-7-O-glucoside | C33H40O21 | 771.1998 | 771.1989 | 3.74 | 284.0327, 285.0401, 446.0861, 447.0912, 609.1476 |
| 52 | 4.82 | Kaempferol 3-O-(sinapyl) sophoroside-7-O-glucoside | C44H50O25 | 977.2593 | 977.2568 | 2.54 | 284.0326, 285.0399, 446.0860, 609.1473, 815.2058 |
| 53 | 4.74 | Kaempferol 3-O-(caffeoyl)sophoroside-7-O-glucoside | C42H46O24 | 933.2328 | 933.2306 | 2.35 | 284.0327, 285.0403, 609.1472, 771.1789 |
| 54 | 4.94 | Kaempferol 3-O-(p-coumaroyl)sophoroside-7-O-glucoside | C42H46O23 | 917.2375 | 917.2357 | 1.98 | 284.0327, 285.0399, 446.0858, 591.1365, 609.1474, 755.1841 |
| 55 | 4.89 | Kaempferol 3-O-(feruloyl)sophoroside-7-O-glucoside | C43H48O24 | 947.2485 | 947.2463 | 2.37 | 284.0326, 285.0398, 446.0857, 591.1361, 609.1478, 785.1940 |
| 56 | 6.73 | Apigenin 7-rhamnoside-4′-rutinoside | C33H40O18 | 723.2150 | 723.2142 | 1.10 | 175.0390, 193.0498, 205.0499, 223.0608 |
| 57 | 9.21 | Naringenin | C15H12O5 | 271.0612 | 271.0612 | 0.07 | 151.0037, 177.0193, 119.0502 |
| Other compounds | |||||||
| 58 | 4.76 | 3-Acetoxy-4-methoxybenzenesulfonic acid/4-Ethoxy-2-sulfobenzoic acid | C9H10O6S | 245.0124 | 245.0125 | 0.43 | 165.0553, 79.9566 |
| 59 | 4.75 | Methyl 3,4-O-isopropylidene-2-O-(methylsulfonyl)pentopyranoside | C10H18O7S | 281.0702 | 281.0700 | 0.60 | 96.9594, 201.1127, 281.0702 |
| 60 | 0.80 | Glutathione | C10H17N3O6S | 306.0764 | 306.0765 | 0.33 | 128.0353, 160.0071, 254.0781, 272.0890, 288.0660 |
| 61 | 6.44 | N-Ethylmaleimide-S-glutathione | C16H22N4O8S | 429.1082 | 429.1085 | 0.80 | 385.1181, 343.1071, 241.0024, 96.9594 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-Q.; Hu, L.-P.; Liu, G.-M.; Zhang, D.-S.; He, H.-J. Evaluation of the Nutritional Quality of Chinese Kale (Brassica alboglabra Bailey) Using UHPLC-Quadrupole-Orbitrap MS/MS-Based Metabolomics. Molecules 2017, 22, 1262. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22081262
Wang Y-Q, Hu L-P, Liu G-M, Zhang D-S, He H-J. Evaluation of the Nutritional Quality of Chinese Kale (Brassica alboglabra Bailey) Using UHPLC-Quadrupole-Orbitrap MS/MS-Based Metabolomics. Molecules. 2017; 22(8):1262. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22081262
Chicago/Turabian StyleWang, Ya-Qin, Li-Ping Hu, Guang-Min Liu, De-Shuang Zhang, and Hong-Ju He. 2017. "Evaluation of the Nutritional Quality of Chinese Kale (Brassica alboglabra Bailey) Using UHPLC-Quadrupole-Orbitrap MS/MS-Based Metabolomics" Molecules 22, no. 8: 1262. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22081262