Senna singueana: Antioxidant, Hepatoprotective, Antiapoptotic Properties and Phytochemical Profiling of a Methanol Bark Extract

 , ,
, , 
Abstract
:1. Introduction
2. Results and Discussion
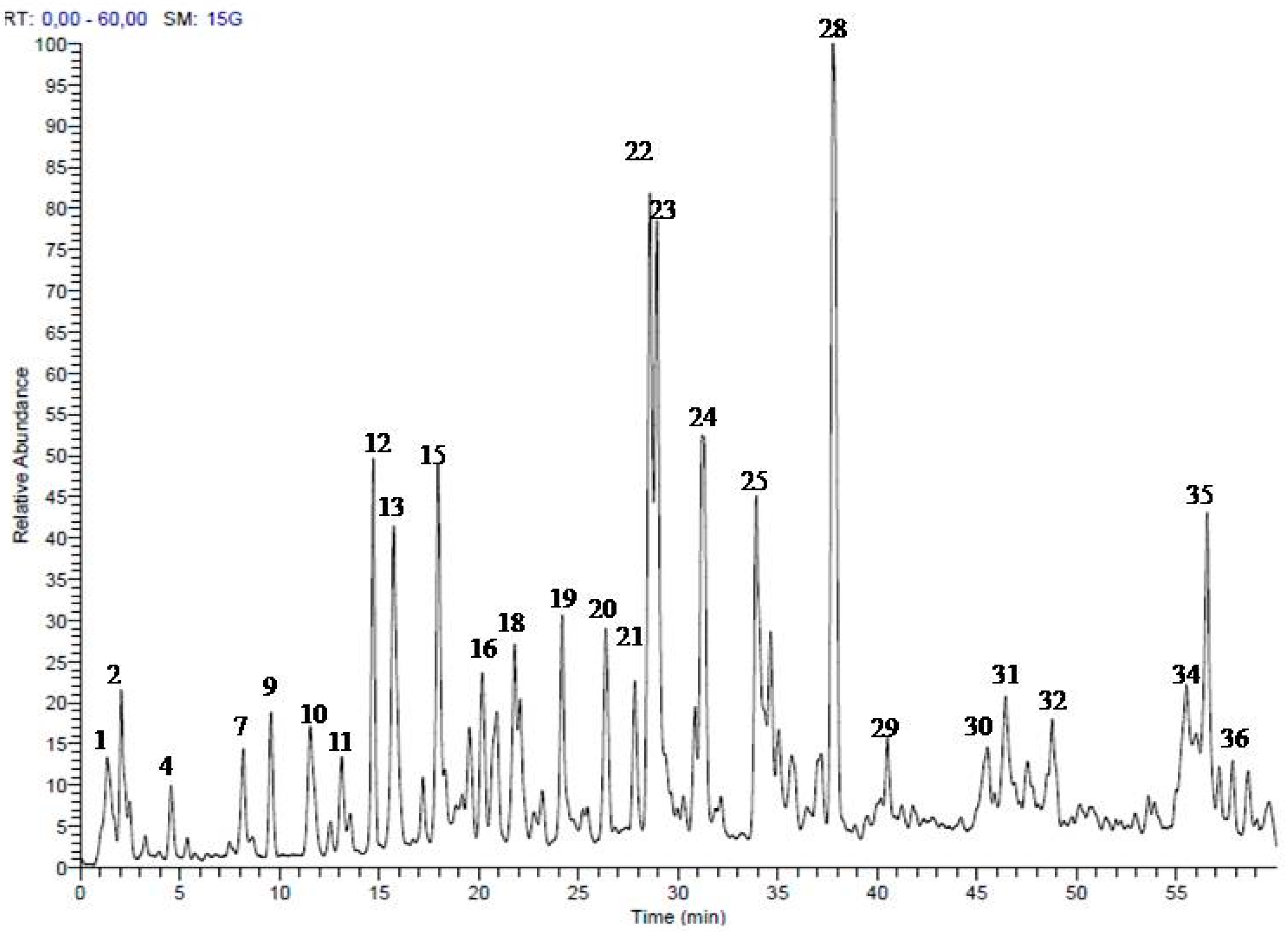
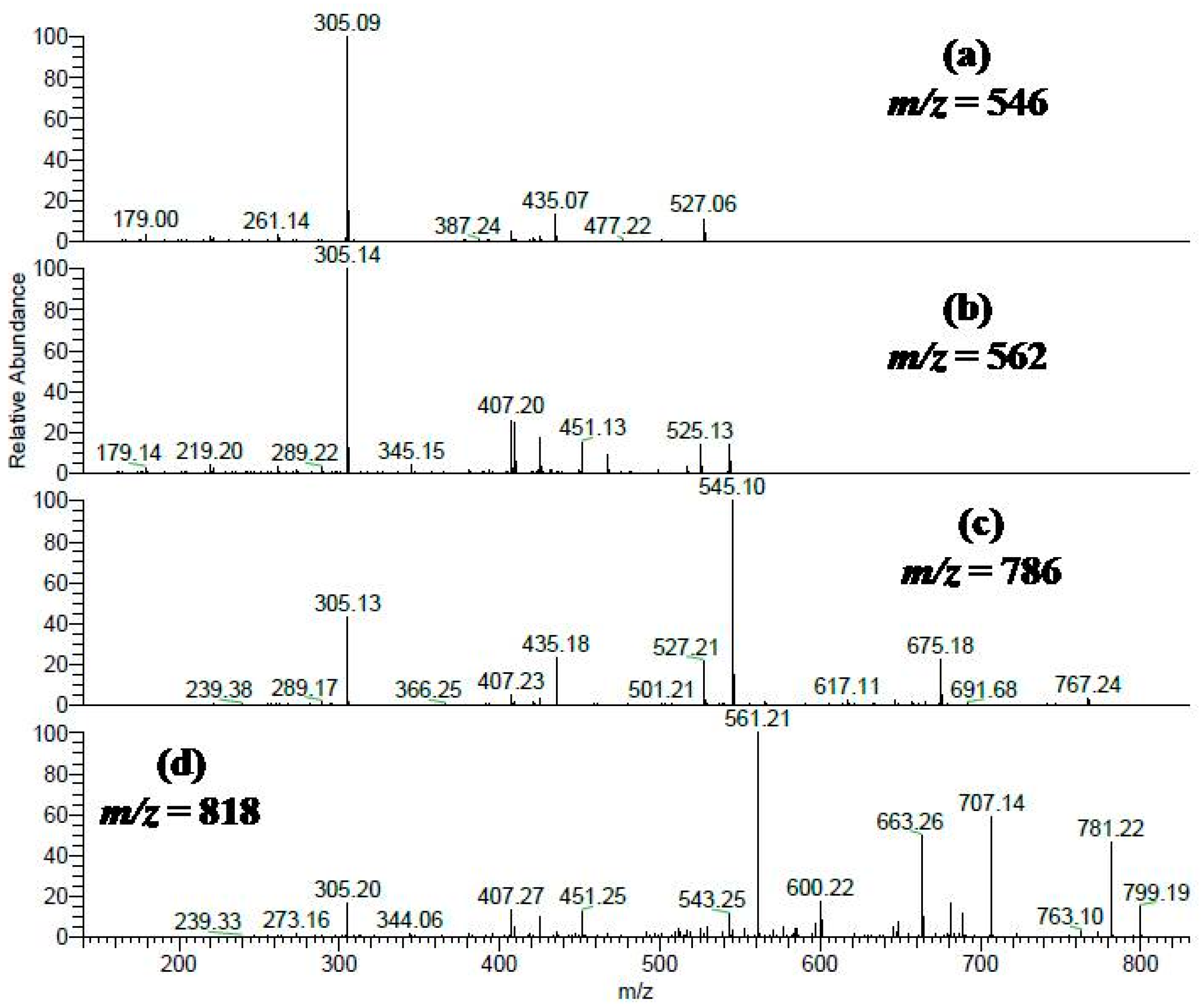
2.1. Secondary Metabolites in the Methanol Extract from S. singueana Bark
2.2. Biological Activities
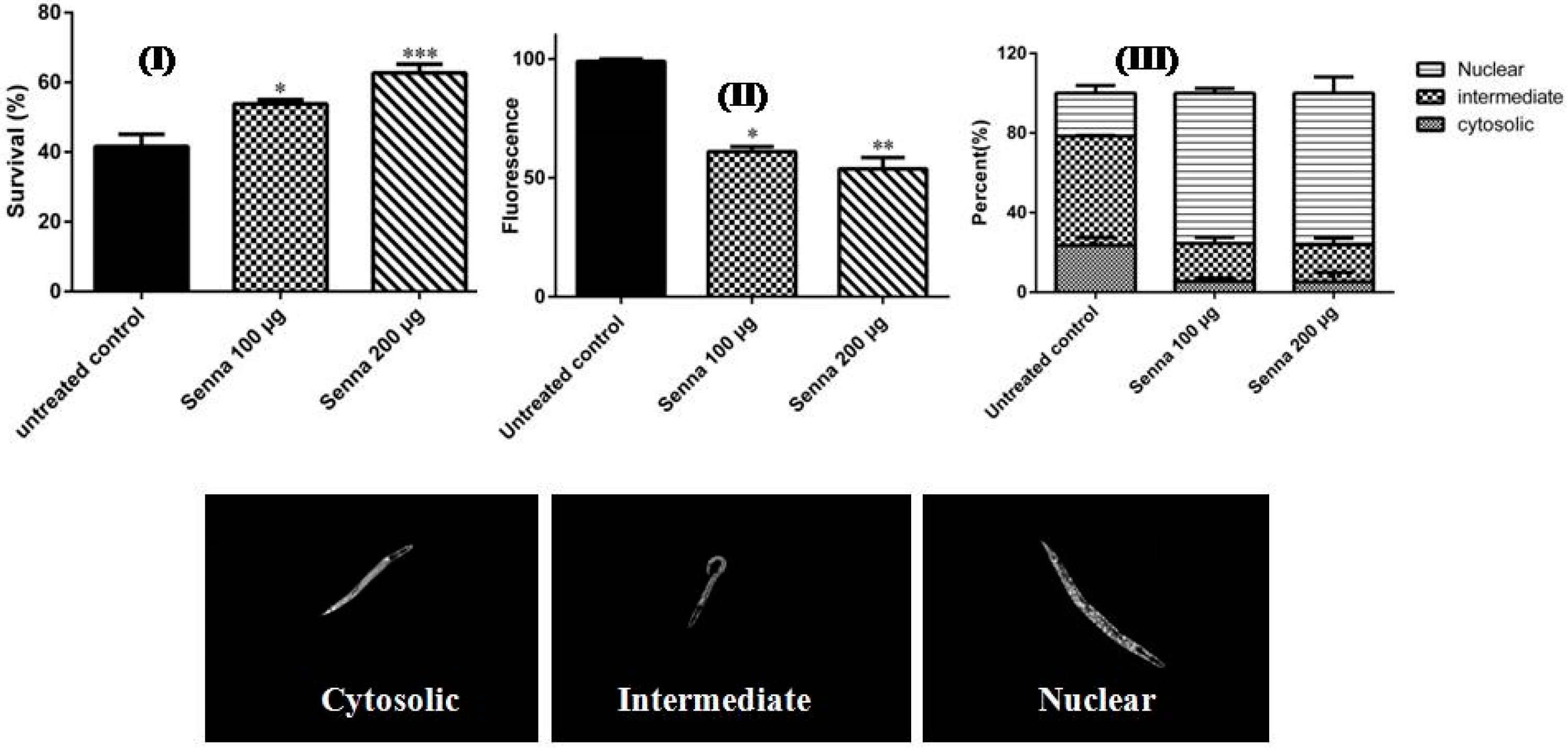
2.2.1. Antioxidant Activities
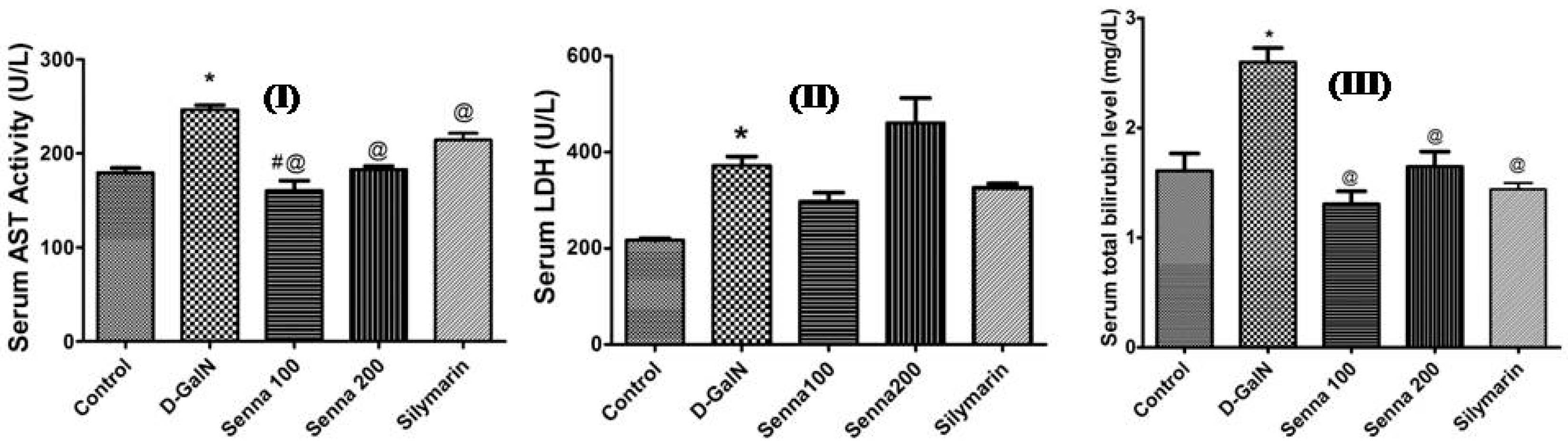
2.2.2. Hepatoprotective Activity
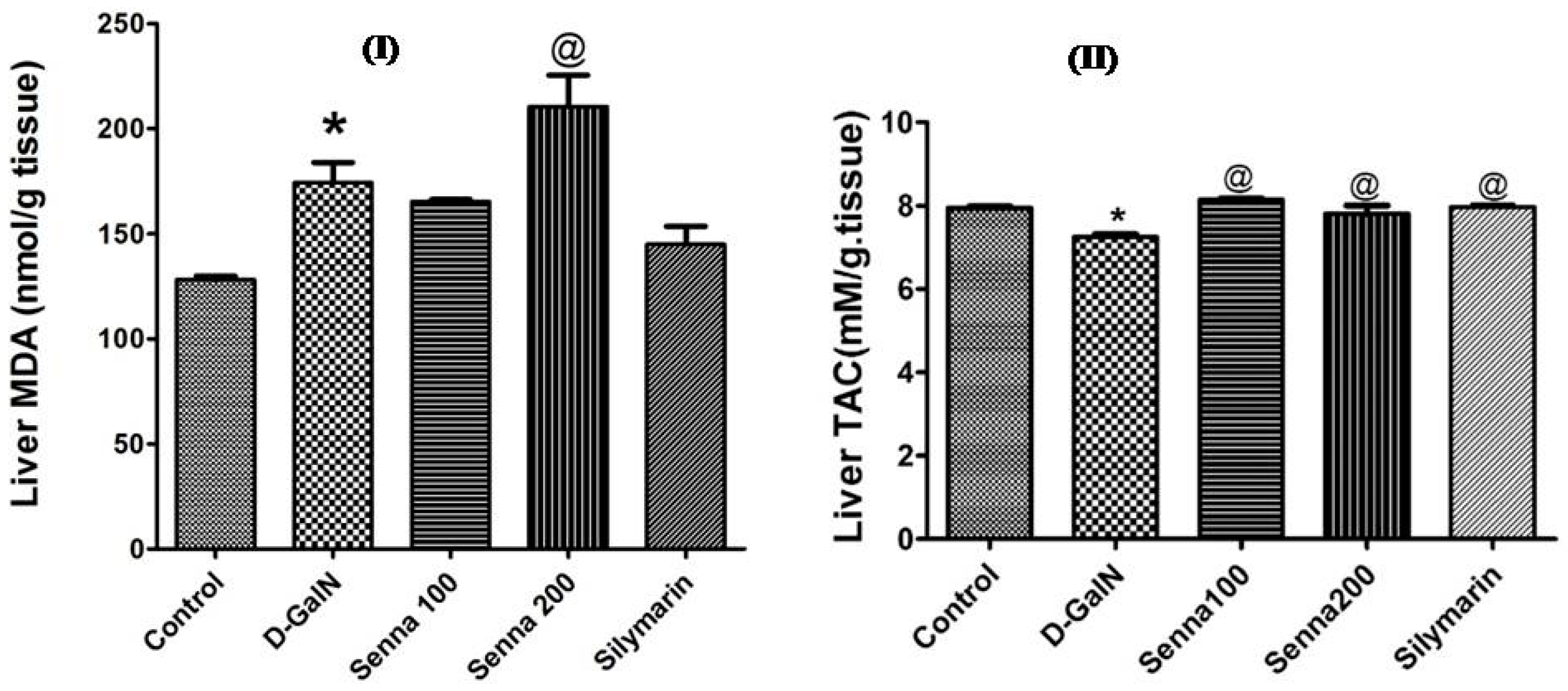
Effects on Liver Functions and Oxidative Stress Markers
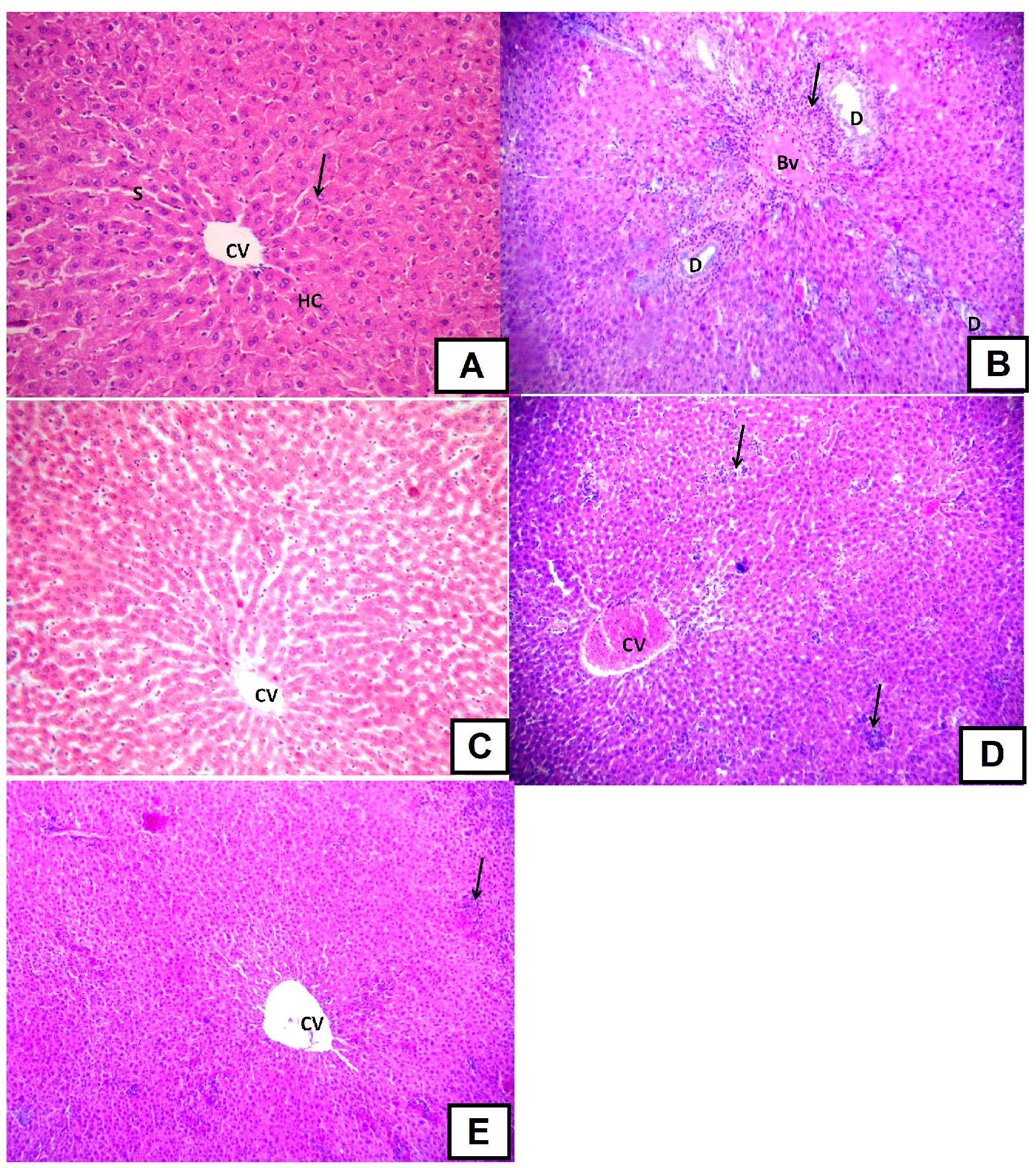
d-GalN-Induced Histopathological Changes
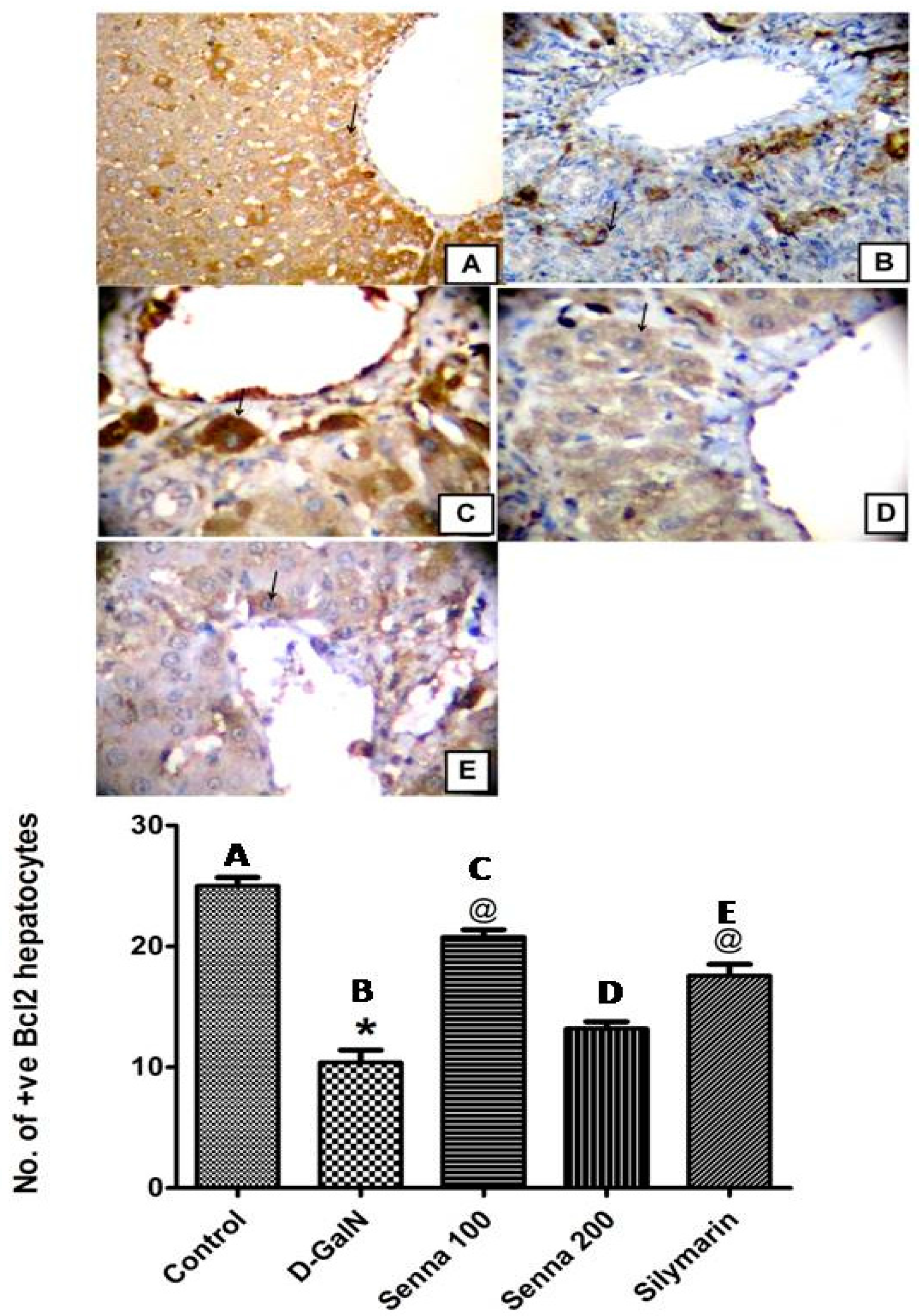
Effects on d-GalN-Induced Apoptosis
3. Materials and Methods
3.1. Plant Material and Extraction
3.2. HPLC-PDA-MS/MS
3.3. Biological Activity
Antioxidant Activities
3.4. Hepatoprotective Activity
3.4.1. Animals
3.4.2. Experimental Design
3.4.3. Blood and Tissue Sampling
3.4.4. Liver Function and Oxidative Stress Markers
3.4.5. Histological Techniques
3.4.6. Immunohistochemical Techniques
3.4.7. Morphometric Study
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Alfadda, A.A.; Sallam, R.M. Reactive oxygen species in health and disease. J. Biomed. Biotechnol. 2012, 14. [Google Scholar] [CrossRef] [PubMed]
- Youssef, F.S.; Ashour, M.L.; Ebada, S.S.; Sobeh, M.; El-Beshbishy, H.A.; Singab, A.N.; Wink, M. Antihyperglycaemic activity of the methanol extract from leaves of Eremophila maculata (Scrophulariaceae) in streptozotocin-induced diabetic rats. J. Pharm. Pharmacol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Sghaier, M.B.; Ismail, M.B.; Bouhlel, I.; Ghedira, K.; Chekir-Ghedira, L. Leaf extracts from Teucrium ramosissimum protect against DNA damage in human lymphoblast cell K562 and enhance antioxidant, antigenotoxic and antiproliferative activity. Environ. Toxicol. Pharmacol. 2016, 44, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Del Giudice, R.; Petruk, G.; Raiola, A.; Barone, A.; Monti, D.M.; Rigano, M.M. Carotenoids in fresh and processed tomato (Solanum lycopersicum) fruits protect cells from oxidative stress injury. J. Sci. Food Agric. 2017, 97, 1616–1623. [Google Scholar] [CrossRef] [PubMed]
- Tolba, M.F.; Omar, H.A.; Azab, S.S.; Khalifa, A.E.; Abdel-Naim, A.B.; Abdel-Rahman, S.Z. Caffeic acid phenethyl ester: A review of its antioxidant activity, protective effects against ischemia-reperfusion injury and drug adverse reactions. Crit. Rev. Food Sci. Nutr. 2016, 56, 2183–2190. [Google Scholar] [CrossRef] [PubMed]
- Petruk, G.; Illiano, A.; Del Giudice, R.; Raiola, A.; Amoresano, A.; Rigano, M.M.; Piccoli, R.; Monti, D.M. Malvidin and cyanidin derivatives from açai fruit (Euterpe oleracea Mart.) counteract UV-A-induced oxidative stress in immortalized fibroblasts. J. Photochem. Photobiol. B 2017, 172, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Mabberley, D.J. Mabberley’s Plant-Book: A Portable Dictionary of Plants, Their Classifications and Uses, 3rd ed.; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Mondal, A. Phenolic constituents and traditional uses of Cassia (Fabaceae) plants: An update. Org. Biomol. Chem. 2014, 3, 93–141. [Google Scholar]
- Keter, L.K.; Mutiso, P.C. Ethnobotanical studies of medicinal plants used by traditional health practitioners in the management of diabetes in lower Eastern Province, Kenya. J. Ethnopharmacol. 2012, 139, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Moshi, M.J.; Mbwambo, Z.H. Experience of tanzanian traditional healers in the management of non-insulin dependent diabetes mellitus. Pharm. Biol. 2002, 40, 552–560. [Google Scholar] [CrossRef]
- Musa, M.S.; Abdelrasool, F.E.; Elsheikh, E.A.; Ahmed, L.A.; Mahmoud, A.L.E.; Yagi, S.M. Ethnobotanical study of medicinal plants in the Blue Nile State, south-eastern Sudan. J. Med. Plants Res. 2011, 5, 4287–4297. [Google Scholar]
- Gebrelibanos, M. In vitro erythrocyte haemolysis inhibition properties of Senna singueana extracts. Momona Ethiop. J. Sci. (MEJS) 2012, 4, 16–28. [Google Scholar]
- Madubunyi, I.I.; Ode, O.J. In vitro and in vivo antioxidant potential of the methanolic extract of Cassia singueana Delile (Fabaceae) Lock leaves. Comp. Clin. Pathol. 2012, 21, 1565–1569. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Islam, M.S. Anti-diabetic effects of the acetone fraction of Senna singueana stem bark in a type 2 diabetes rat model. J. Ethnopharmacol. 2014, 153, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Adzu, B.; Abbah, J.; Vongtau, H.; Gamaniel, K. Studies on the use of Cassia singueana in malaria ethnopharmacy. J. Ethnopharmacol. 2003, 88, 261–267. [Google Scholar] [CrossRef]
- Hiben, M.G.; Sibhat, G.G.; Fanta, B.S.; Gebrezgi, H.D.; Tesema, S.B. Evaluation of Senna singueana leaf extract as an alternative or adjuvant therapy for malaria. J. Tradit. Complement. Med. 2016, 6, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Kariuki, H.N.; Kanui, T.I.; Yenesew, A.; Mbugua, P.M.; Patel, N.B. Antinocieptive activity of the root extracts of Rhus natalensis Kraus and Senna singueana. Phytopharmacol. 2012, 2, 312–317. [Google Scholar]
- Shawa, I.T.; Mponda, J.; Msefula, C.; Manda, H.; Gondwe, M.; Maliwichi-Nyirenda, C. Brine shrimp lethality and phytochemical determination of aqueous extracts of Senna singueana, musa paradisiaca and ziziphus mucronata in Malawi. J. Basic Appl. Res. 2015, 1, 82–88. [Google Scholar]
- Nibret, E.; Ashour, M.L.; Rubanza, C.D.; Wink, M. Screening of some tanzanian medicinal plants for their trypanocidal and cytotoxic activities. Phytother. Res. 2010, 24, 945–947. [Google Scholar] [CrossRef] [PubMed]
- Ojinnaka, C.M. African Herbal Medicine: The Chemical Constituents and Phytotherapeutic Application of Some West African Senna (syn. Cassia species), 4th ed.; Jeson Books: Port Harcourt, Nigeria, 2012. [Google Scholar]
- Babiaka, S.B.; Ntie-Kang, F.; Ndingkokhar, B.; Mbah, J.A.; Sippl, W.; Yong, J.N. The chemistry and bioactivity of Southern African flora II: Flavonoids, quinones and minor compound classes. RSC Adv. 2015, 5, 57704–57720. [Google Scholar] [CrossRef]
- Farag, M.A.; Porzel, A.; Mahrous, E.A.; El-Massry, M.M.; M. Wessjohann, L.A. Integrated comparative metabolite profiling via MS and NMR techniques for Senna drug quality control analysis. Anal. Bioanal. Chem. 2015, 407, 1937–1949. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, H.; Roxo, M.; Krstin, S.; Röhrig, T.; Richling, E.; Wink, M. An anthocyanin-rich extract of acai (euterpe precatoria Mart.) increases stress resistance and retards aging-related markers in Caenorhabditis elegans. J. Agric. Food Chem. 2016, 64, 1283–1290. [Google Scholar] [CrossRef] [PubMed]
- Bergantin, C.; Maietti, A.; Cavazzini, A.; Pasti, L.; Tedeschi, P.; Brandolini, V.; Marchetti, N. Bioaccessibility and HPLC-MS/MS chemical characterization of phenolic antioxidants in red chicory (Cichorium intybus). J. Funct. Foods 2017, 33, 94–102. [Google Scholar] [CrossRef]
- Sobeh, M.; Mahmoud, M.F.; Abdelfattah, M.A.O.; El-Beshbishy, H.A.; El-Shazly, A.M.; Wink, M. Albizia harveyi: Phytochemical profiling, antioxidant, antidiabetic and hepatoprotective activities of the bark extract. Med. Chem. Res. 2017. [Google Scholar] [CrossRef]
- Sobeh, M.; Mahmoud, M.F.; Abdelfattah, M.A.O.; El-Beshbishy, H.A.; El-Shazly, A.M.; Wink, M. Hepatoprotective and hypoglycemic effects of a tannin rich extract from Ximenia americana var. caffra root. Phytomedicine 2017, 33, 36–42. [Google Scholar] [CrossRef]
- Malan, E.; Swinny, E.; Ferreira, D.; Steynberg, P. The structure and synthesis of proguibourtinidins from Cassia abbreviata. Phytochemistry 1996, 41, 1209–1213. [Google Scholar] [CrossRef]
- Coetzee, J.; McIteka, L.; Malan, E.; Ferreira, D. Structure and synthesis of the first procassinidin dimers based on epicatechin, and gallo- and epigallo-catechin. Phytochemistry 2000, 53, 795–804. [Google Scholar] [CrossRef]
- Mokgotho, M.P.; Gololo, S.S.; Masoko, P.; Mdee, L.K.; Mbazima, V.; Shai, L.J.; Bagla, V.P.; Eloff, J.N.; Mampuru, L. Isolation and chemical structural characterisation of a compound with antioxidant activity from the roots of Senna italica. J. Evid. Based Complemen. Alter. Med. 2013. [Google Scholar] [CrossRef] [PubMed]
- Sagnia, B.; Fedeli, D.; Casetti, R.; Montesano, C.; Falcioni, G.; Colizzi, V. Antioxidant and anti-inflammatory activities of extracts from Cassia alata, Eleusine indica, Eremomastax speciosa, Carica papaya and Polyscias fulva medicinal plants collected in Cameroon. PLoS ONE 2014, 9, e103999. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Wink, M. Chlorophyll enhances oxidative stress tolerance in Caenorhabditis elegans and extends its lifespan. Peer J. 2016, 4, e1879. [Google Scholar] [CrossRef] [PubMed]
- Abbas, S.; Wink, M. Epigallocatechin gallate from green tea (Camellia sinensis) increases lifespan and stress resistance in Caenorhabditis elegans. Planta Med. 2009, 75, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Abbas, S.; Wink, M. Green tea extract induces the resistance of Caenorhabditis elegans against oxidative stress. Antioxidants 2014, 3, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Sudji, I.R.; Wang, E.; Joubert, E.; van Wyk, B.-E.; Wink, M. Ameliorative effect of aspalathin from rooibos (Aspalathus linearis) on acute oxidative stress in Caenorhabditis elegans. Phytomedicine 2013, 20, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Wink, M. Natural lignans from Arctium lappa as antiaging agents in Caenorhabditis elegans. Phytochemistry 2015, 117, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.M. Acute liver failure in the United States. In Seminars in Liver Disease; Thieme Medical Publishers Inc.: New York, NY, USA, 2003. [Google Scholar]
- Weng, H.-L.; Cai, X.; Yuan, X.; Liebe, R.; Dooley, S.; Li, H.; Wang, T.-L. Two sides of one coin: Massive hepatic necrosis and progenitor cell-mediated regeneration in acute liver failure. Front. Physiol. 2015, 6, 178. [Google Scholar] [CrossRef] [PubMed]
- Keppler, D.; Lesch, R.; Reutter, W.; Decker, K. Experimental hepatitis induced by d-galactosamine. Exp. Mol. Pathol. 1968, 9, 279–290. [Google Scholar] [CrossRef]
- Youssef, F.S.; Ashour, M.L.; Sobeh, M.; El-Beshbishy, H.A.; Singab, A.N.; Wink, M. Eremophila maculata-isolation of a rare naturally-occurring lignan glycoside and the hepatoprotective activity of the leaf extract. Phytomedicine 2016, 23, 1484–1493. [Google Scholar] [CrossRef] [PubMed]
- Youle, R.J.; Strasser, A. The BCL-2 protein family: Opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Ganai, A.A.; Jahan, S.; Ahad, A.; Abdin, M.; Farooqi, H. Glycine propionyl l-carnitine attenuates d-galactosamine induced fulminant hepatic failure in wistar rats. Chem. Biol. Interact. 2014, 214, 33–40. [Google Scholar] [CrossRef] [PubMed]
- El Swefy, S.; Hasan, R.A.; Ibrahim, A.; Mahmoud, M.F. Curcumin and hemopressin treatment attenuates cholestasis-induced liver fibrosis in rats: Role of CB1 receptors. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2016, 389, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Sobeh, M.; Mamadalieva, N.Z.; Mohamed, T.; Krstin, S.; Youssef, F.S.; Ashour, M.L.; Azimova, S.S.; Wink, M. Chemical profiling of Phlomis thapsoides (Lamiaceae) and in vitro testing of its biological activities. Med. Chem. Res. 2016, 25, 2304–2315. [Google Scholar] [CrossRef]
- Sobeh, M.; ElHawary, E.A.; Peixoto, H.; Labib, R.M.; Handoussa, H.; Swilam, N.; El-Khatib, A.H.; Sharapov, F.; Mahmoud, T.; Krstin, S. Identification of phenolic secondary metabolites from Schotia brachypetala sond. (Fabaceae) and demonstration of their antioxidant activities in Caenorhabditis elegans. Peer J. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Link, P.; Roth, K.; Sporer, F.; Wink, M. Carlina acaulis exhibits antioxidant activity and counteracts Aβ toxicity in Caenorhabditis elegans. Molecules 2016, 21, 871. [Google Scholar] [CrossRef] [PubMed]
- Stiernagle, T. Maintenance of C. elegans; Wormbook, Ed.; The C. elegans Research Community, Wormbook: Pasadena, CA, USA, 2006; pp. 51–67. [Google Scholar]
- Young, D.S. Effects of Drugs on Clinical Laboratory Tests, 4th ed.; The American Association for Clinical Chemistry: Washington, DC, USA, 1995. [Google Scholar]
- Balistreri, W.; Shaw, L. Liver function. In Fundamentals of Clinical Chemistry, 3rd ed.; WB Saunders Co.: Philadelphia, PA, USA, 1987; pp. 729–761. [Google Scholar]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Koracevic, D.; Koracevic, G.; Djordjevic, V.; Andrejevic, S.; Cosic, V. Method for the measurement of antioxidant activity in human fluids. J. Clin. Pathol. 2001, 54, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, J.; Stevens, A.; Turner, D. Theoryand Practice of Histological Techniques, 4th ed.; Churchill Living Stone: Edinburgh, UK, 1996; pp. 47–67. [Google Scholar]
- Ozen, O.A.; Kus, M.A.; Kus, I.; Alkoc, O.A.; Songur, A. Protective effects of melatonin against formaldehyde-induced oxidative damage and apoptosis in rat testes: An immunohistochemical and biochemical study. Syst. Biol. Reprod. Med. 2008, 54, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Han, Y.; Yang, H.; Li, S. Effects of SSTF on the expression of apoptosis-associated gene Bcl-2 and Bax by cardiomyocytes induced by H2O2. Guangdong Med. J. 2005, 28, 1590–1591. [Google Scholar]
- Salama, S.M.; Abdulla, M.A.; AlRashdi, A.S.; Ismail, S.; Alkiyumi, S.S.; Golbabapour, S. Hepatoprotective effect of ethanolic extract of Curcuma longa on thioacetamide induced liver cirrhosis in rats. BMC Complement. Altern. Med. 2013, 13, 56. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Sample of the plant materials available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Rt (min) | [M − H]− | MS/MS | Proposed Compounds | Ref. |
|---|---|---|---|---|---|
| 1 | 1.33 | 133 | 115 | Malic acid | [26] |
| 2 | 1.68 | 447 | 163, 315 | p-Coumaric acid galloyl-pentoside | |
| 3 | 2.48 | 153 | Protocatechuic acid | ||
| 4 | 4.52 | 305 | 179, 221, 287 | (epi)Gallocatechin | [25] |
| 5 | 5.33 | 417 | 161, 205, 270 | Unknown | |
| 6 | 5.90 | 593 | 289, 441 | (epi)Catechin digallate | |
| 7 | 8.18 | 305 | 125, 179, 287 | (epi)Gallocatechin | [25] |
| 8 | 8.48 | 593 | 305, 425, 575 | (epi)Gallocatechin-(epi)catechin | [25] |
| 9 | 9.57 | 289 | 179, 205, 245 | (epi)Catechin | [26] |
| 10 | 11.52 | 577 | 289, 425, 559 | (epi)Catechin-(epi)catechin | [26] |
| 11 | 13.10 | 577 | 289, 425, 559 | (epi)Catechin-(epi)catechin | [26] |
| 12 | 14.65 | 289 | 179, 205, 245 | Catechin | [26] |
| 13 | 15.76 | 561 | 179, 305, 543 | (epi)Guibourtinidol-(epi)gallocatechin | |
| 14 | 17.14 | 561 | 289, 451, 543 | (epi)Catechin-(epi)afzelechin | [26] |
| 15 | 17.90 | 561 | 179, 305, 543 | (epi)Guibourtinidol-(epi)gallocatechin | |
| 16 | 20.16 | 561 | 179, 305, 543 | (epi)Guibourtinidol-(epi)gallocatechin | |
| 17 | 20.87 | 545 | 289, 409, 527 | (epi)Guibourtinidol-(epi)catechin | [27] |
| 18 | 21.82 | 561 | 179, 305, 543 | (epi)Guibourtinidol-(epi)gallocatechin | |
| 19 | 24.20 | 545 | 179, 289, 527 | (epi)Guibourtinidol-(epi)catechin | [27] |
| 20 | 26.37 | 545 | 179, 289, 527 | (epi)Guibourtinidol-(epi)catechin | [27] |
| 21 | 27.82 | 545 | 179, 289, 527 | (epi)Guibourtinidol-(epi)catechin | [27] |
| 22 | 28.64 | 545 | 179, 305, 527 | (ent)Cassiaflavan-(epi)gallocatechin | [28] |
| 23 | 29.30 | 545 | 179, 305, 527 | (ent)Cassiaflavan-(epi)gallocatechin | [28] |
| 24 | 31.31 | 833 | 305, 561, 577 | (epi)Gallocatechin-(epi)catechin-(ent)cassiaflavan | |
| 25 | 33.96 | 529 | 289, 419, 511 | (ent)Cassiaflavan-(epi)catechin | [28] |
| 26 | 34.72 | 817 | 305, 561, 663 | (epi)Guibourtinidol-(epi)guibourtinidol-(epi)gallocatechin | |
| 27 | 37.58 | 833 | 305, 593, 723 | (epi)Gallocatechin-(epi)catechin-(ent)cassiaflavan | |
| 28 | 37.73 | 529 | 179, 289, 511 | (ent)Cassiaflavan-(epi)catechin | [28] |
| 29 | 40.56 | 801 | 289, 545, 691 | (epi)Guibourtinidol-(epi)guibourtinidol-(epi)catechin | |
| 30 | 45.35 | 801 | 305, 561, 691 | (ent)Cassiaflavan-(epi)guibourtinidol-(epi)gallocatechin | |
| 31 | 46.41 | 785 | 305, 545, 767 | (ent)Cassiaflavan-(ent)cassiaflavan-(epi)gallocatechin | |
| 32 | 48.75 | 801 | 305, 425, 561 | (ent)Cassiaflavan-(epi)guibourtinidol-(epi)gallocatechin | |
| 33 | 53.87 | 785 | 289, 545, 675 | (ent)Cassiaflavan-(epi)guibourtinidol-(epi)catechin | |
| 34 | 55.50 | 785 | 305, 545, 675 | (ent)Cassiaflavan-(ent)cassiaflavan-(epi)gallocatechin | |
| 35 | 56.50 | 785 | 305, 545, 675 | (ent)Cassiaflavan-(ent)cassiaflavan-(epi)gallocatechin | |
| 36 | 58.62 | 769 | 289, 529 | (ent)Cassiaflavan-(ent)cassiaflavan-(epi)catechin |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobeh, M.; Mahmoud, M.F.; Hasan, R.A.; Cheng, H.; El-Shazly, A.M.; Wink, M. Senna singueana: Antioxidant, Hepatoprotective, Antiapoptotic Properties and Phytochemical Profiling of a Methanol Bark Extract. Molecules 2017, 22, 1502. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22091502
Sobeh M, Mahmoud MF, Hasan RA, Cheng H, El-Shazly AM, Wink M. Senna singueana: Antioxidant, Hepatoprotective, Antiapoptotic Properties and Phytochemical Profiling of a Methanol Bark Extract. Molecules. 2017; 22(9):1502. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22091502
Chicago/Turabian StyleSobeh, Mansour, Mona F. Mahmoud, Rehab A. Hasan, Haroan Cheng, Assem M. El-Shazly, and Michael Wink. 2017. "Senna singueana: Antioxidant, Hepatoprotective, Antiapoptotic Properties and Phytochemical Profiling of a Methanol Bark Extract" Molecules 22, no. 9: 1502. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22091502