Stable Carbon Isotope Composition of the Lipids in Natural Ophiocordyceps sinensis from Major Habitats in China and Its Substitutes


Abstract
:1. Introduction
2. Results and Discussion
2.1. δ13C Values of the Bulk Samples of O. sinensis
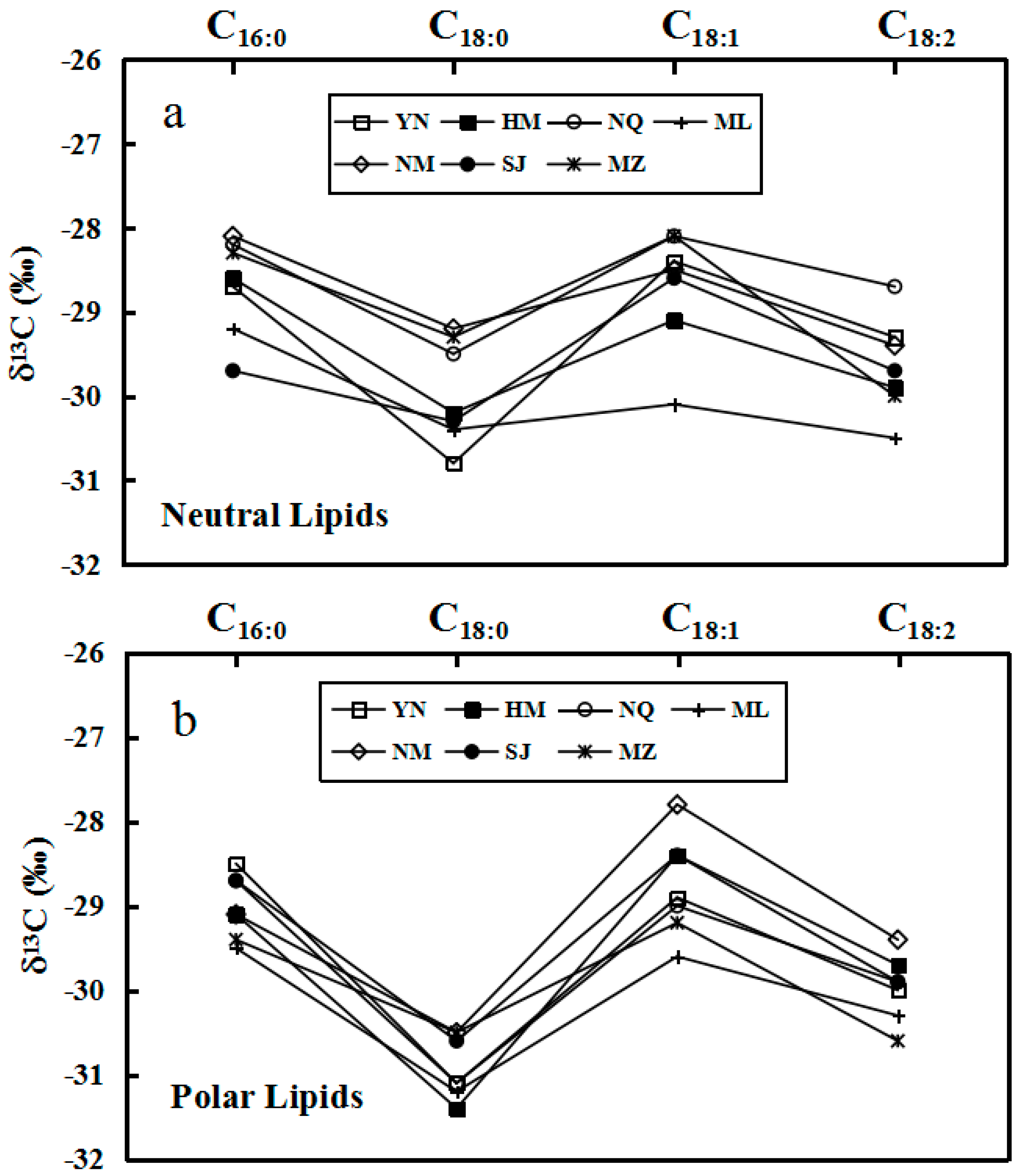
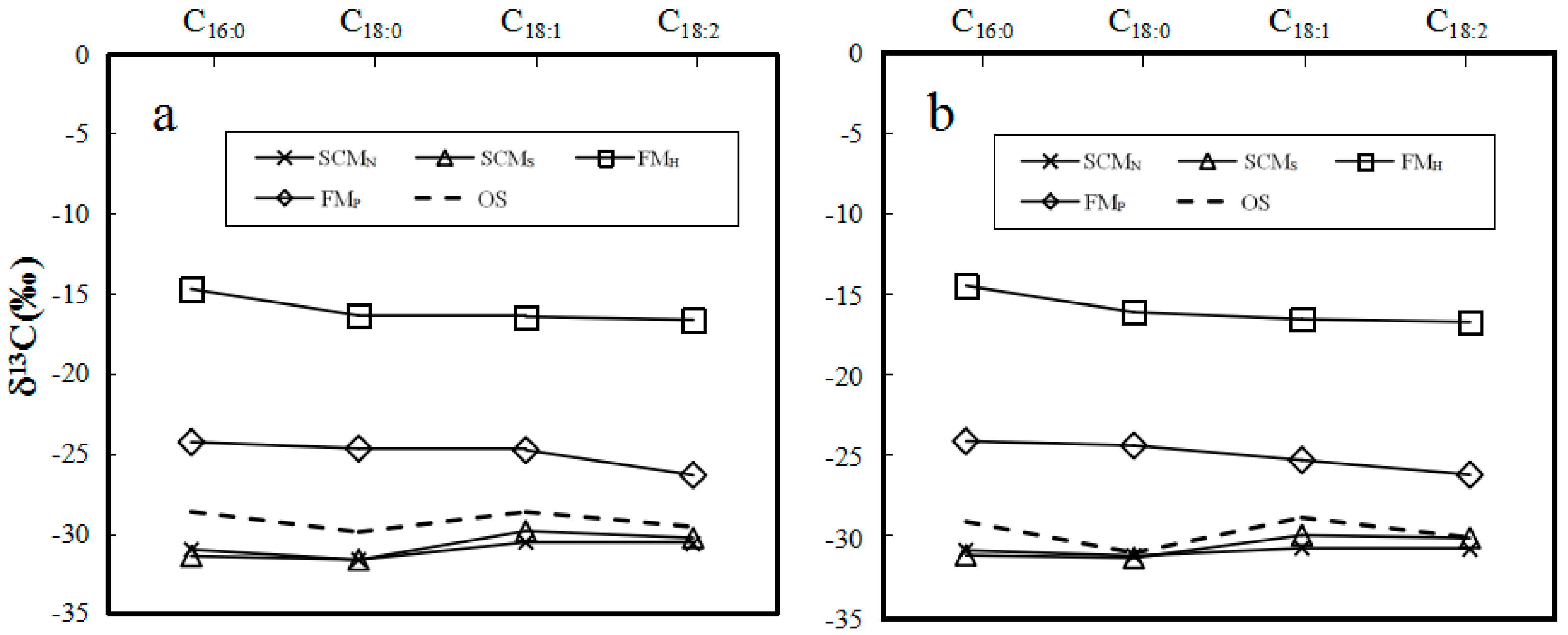
2.2. δ13C Values of Individual Major Fatty Acids of Neutral and Polar Lipids from O. sinensis and Its Substitutes
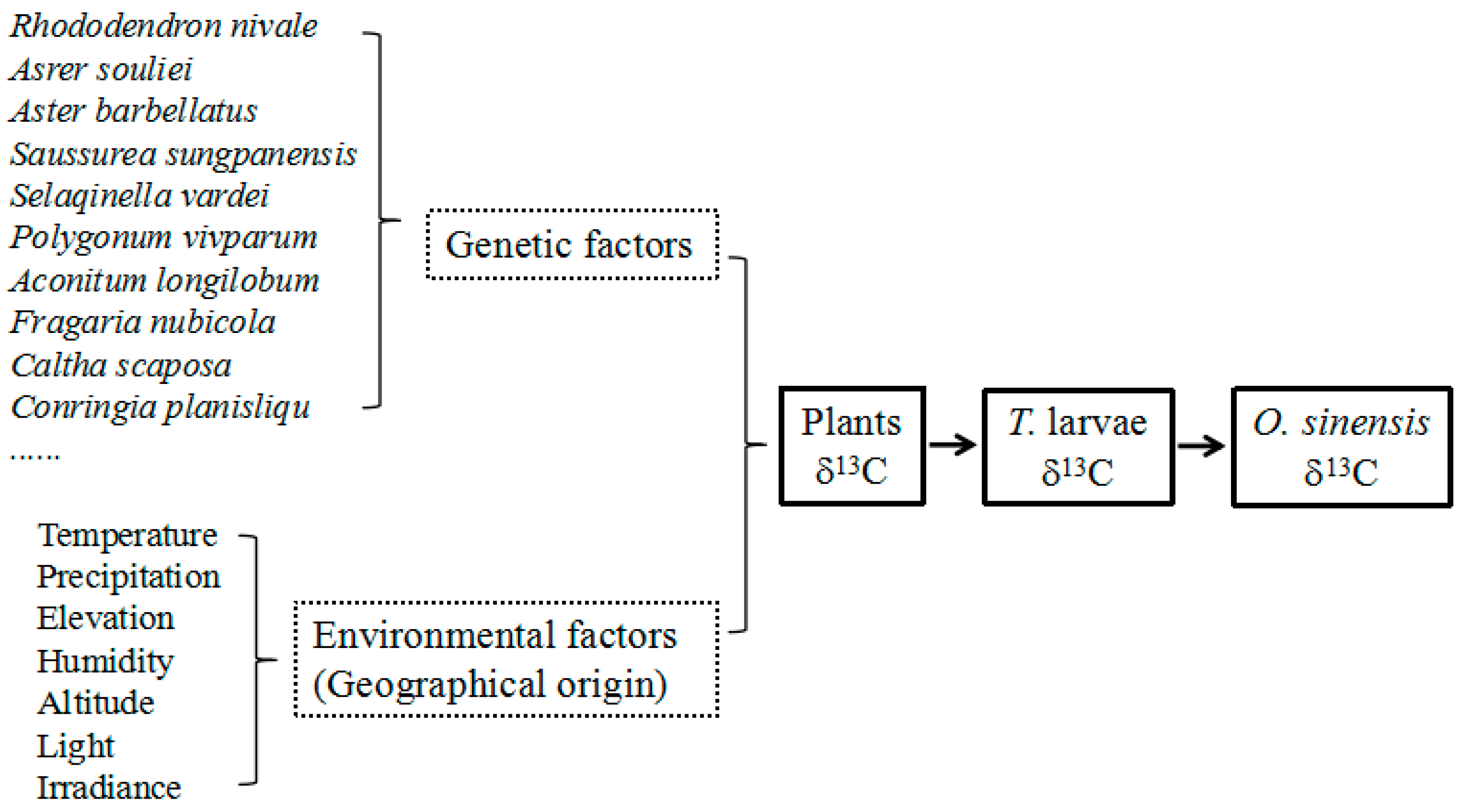
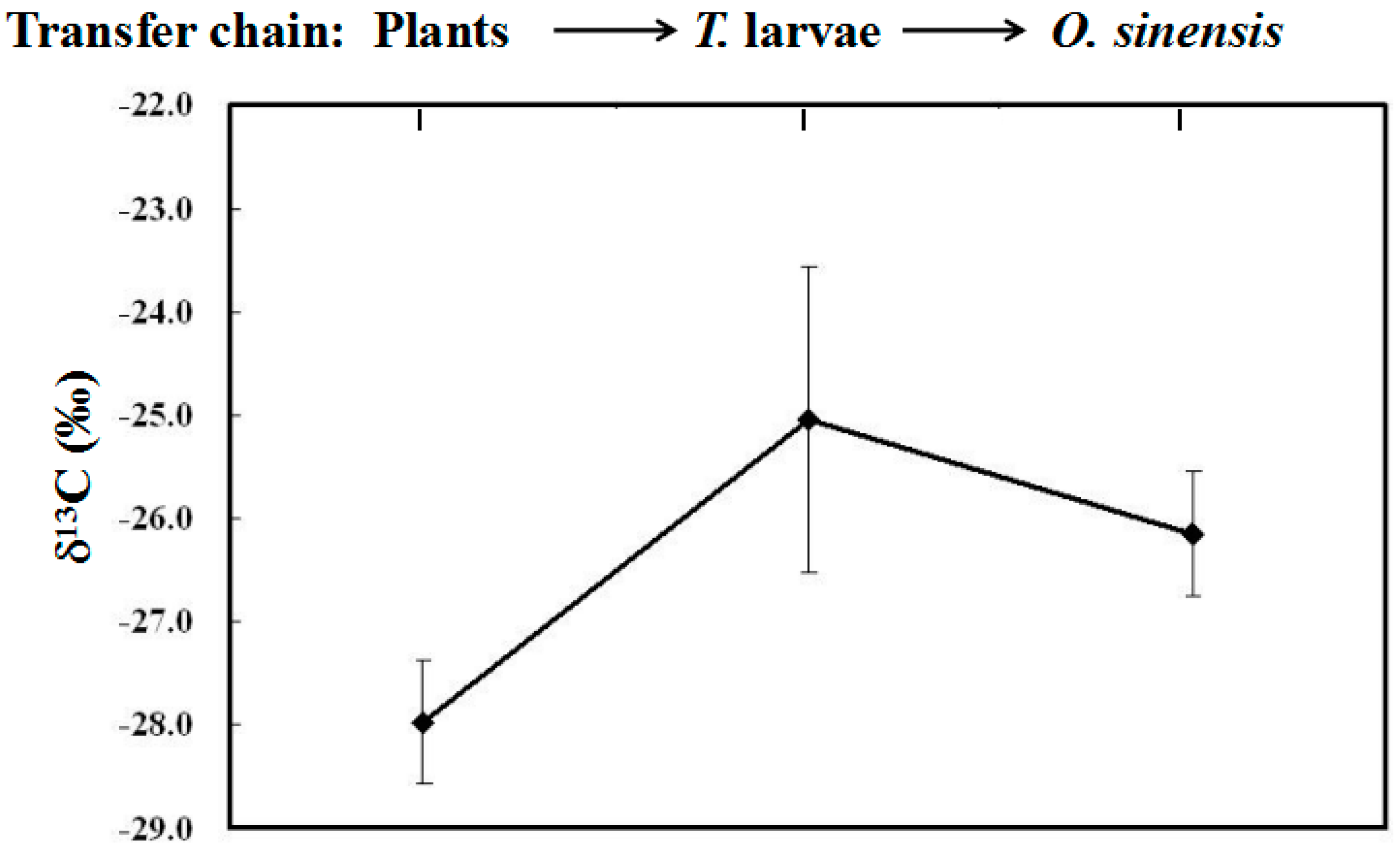
2.3. Stable Carbon Isotope Fractionation among Plants, Larvae and O. sinensis
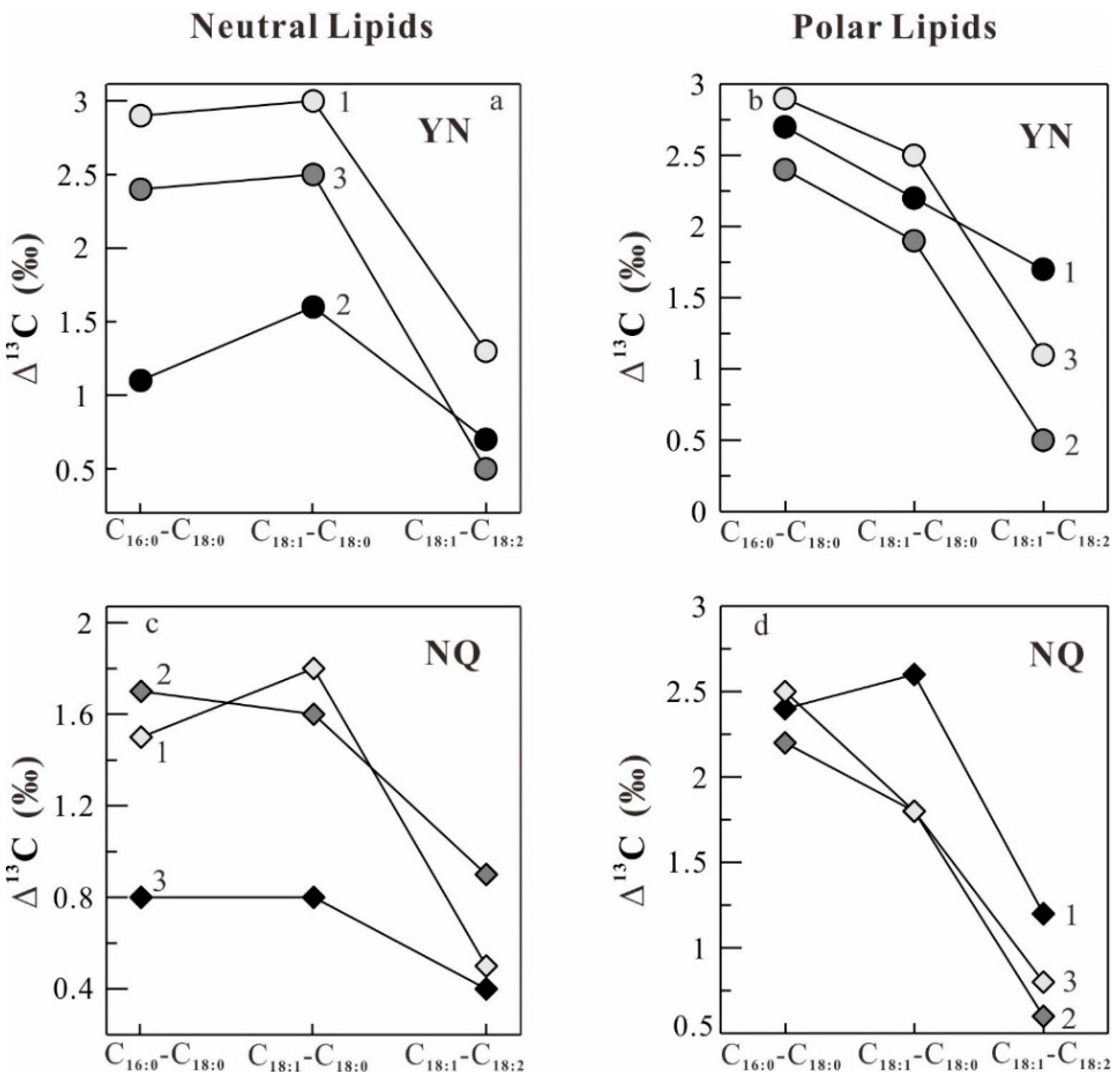
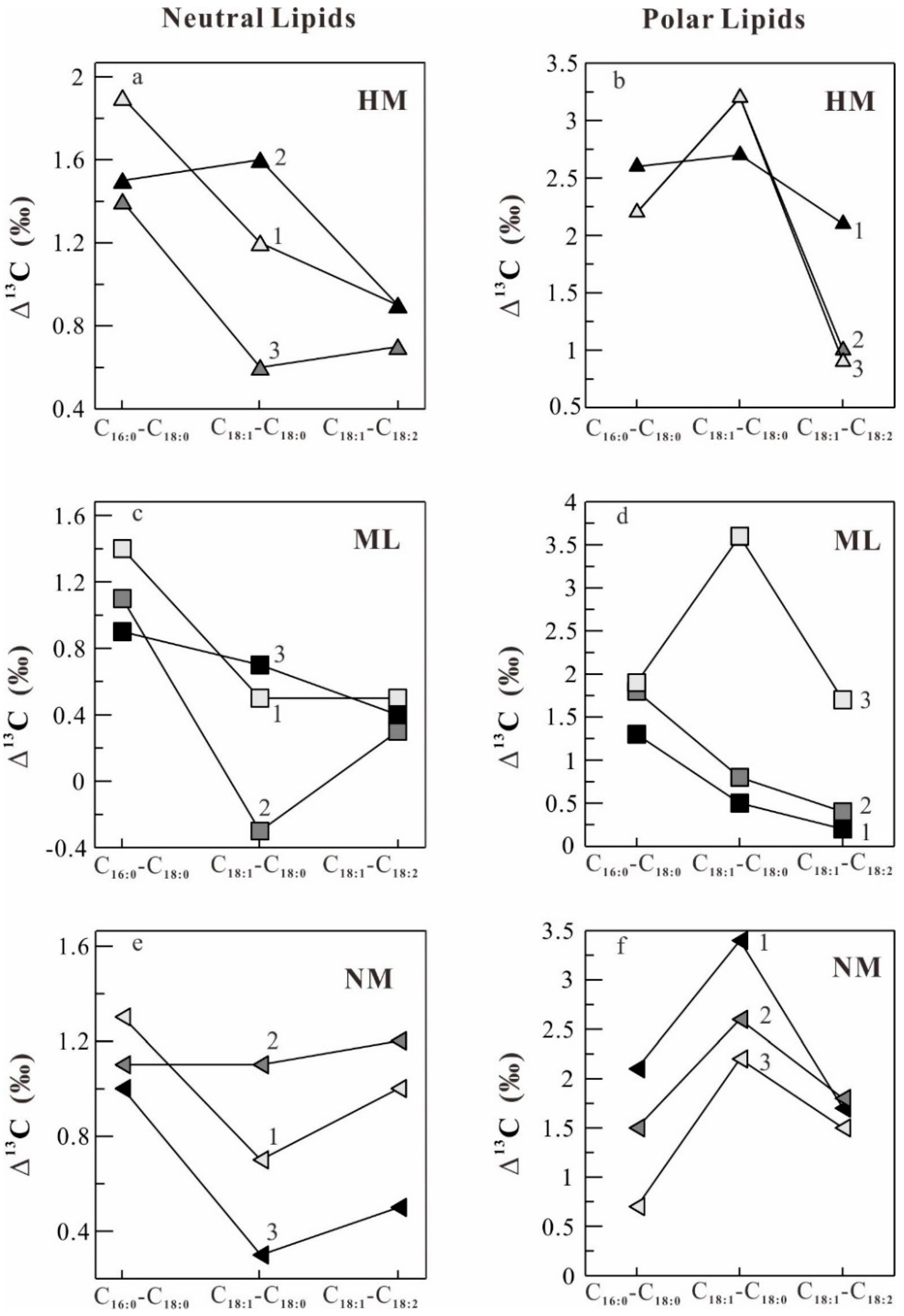
2.4. Stable Carbon Isotope Fractionation in the Lipids of O. sinensis
2.5. Relationship between the δ13C Values of O. sinensis and Its Habitats
2.6. Discrimination between O. sinensis and Its Substitutes
3. Materials and Methods
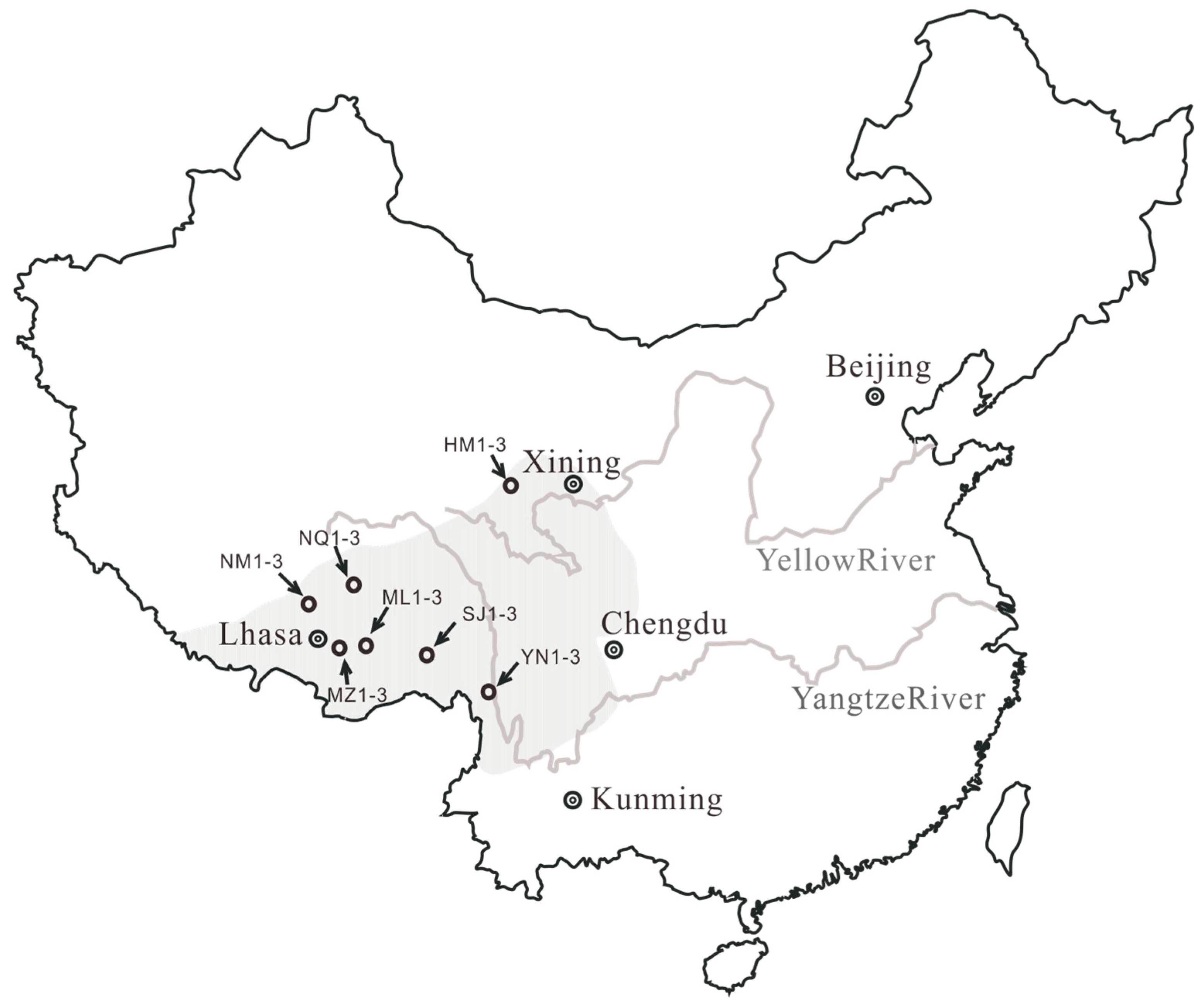
3.1. Samples
3.2. δ13C Analysis of the Bulk Samples
3.3. δ13C Analysis of Individual Major Fatty Acids in Neutral and Polar Lipids
3.4. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, Y.; Wang, X.L.; Jiao, L.; Jiang, Y.; Li, H.; Jiang, S.P.; Lhosumtseiring, N.; Fu, S.Z.; Dong, C.H.; Zhan, Y.; et al. A survey of the geographic distribution of Ophiocordyceps sinensis. J. Microbiol. 2011, 49, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, U.B.; Bawa, K.S. Impact of climate change on potential distribution of Chinese caterpillar fungus (Ophiocordyceps sinensis) in Nepal Himalaya. PLoS ONE 2014, 9, e106405. [Google Scholar] [CrossRef] [PubMed]
- Sung, G.H.; Hyweljones, N.L.; Sung, J.M.; Luangsaard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the Clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Yang, J.L.; Zhu, P.; Cheng, K.D.; He, H.X.; Zhu, H.X. Non-support of species complex hypothesis of Cordyceps sinensis by targeted rDNA-ITS sequence analysis. Mycosystema 2009, 28, 724–730. [Google Scholar]
- Hawksworth, D.L. A new dawn for the naming of fungi: Impacts of decisions made in Melbourne in July 2011 on the future publication and regulation of fungal names. IMA Fungus 2011, 2, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.C.; Hsieh, C.; Lin, F.Y.; Hsu, T.H. A systematic review of the mysterious caterpillar fungus Ophiocordyceps sinensis in dongchongxiacao (dōng chóng xià cǎo) and related bioactive ingredients. J. Tradit. Complement. Med. 2013, 3, 16–32. [Google Scholar] [CrossRef]
- Chen, P.X.; Wang, S.A.; Nie, S.P.; Marcone, M. Properties of Cordyceps sinensis: A review. J. Funct. Foods 2013, 5, 550–569. [Google Scholar] [CrossRef]
- Qiu, X.H.; Cao, L.; Han, R.C. The progress, issues and perspectives in the research of Ophiocordyceps sinensis. J. Environ. Entomol. 2016, 38, 1–23. [Google Scholar]
- Zhao, J.; Xie, J.; Wang, L.Y.; Li, S.P. Advanced development in chemical analysis of Cordyceps. J. Pharm. Biomed. Anal. 2014, 87, 271–289. [Google Scholar] [CrossRef] [PubMed]
- Yue, K.; Ye, M.; Zhou, Z.; Sun, W.; Lin, X. The genus Cordyceps: A chemical and pharmacological review. J. Pharm. Pharmacol. 2013, 65, 474–493. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.H.; Cheng, Z.; Yang, X.L.; Li, S.; Ding, Z.Q.; Zhou, T.S.; Zhang, W.J.; Chen, J.K. Genetic diversity and structure of Cordyceps sinensis populations from extensive geographical regions in China as revealed by inter-simple sequence repeat markers. J. Microbiol. 2008, 46, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Quan, Q.M.; Chen, L.L.; Wang, X.; Li, S.; Yang, X.L.; Zhu, Y.G.; Wang, M.; Cheng, Z. Genetic diversity and distribution patterns of host insects of caterpillar fungus Ophiocordyceps sinensis in the Qinghai-Tibet Plateau. PLoS ONE 2014, 9, e92293. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.X.; Xu, X.M.; Wu, C.F.; Lin, L.; Zou, S.C.; Luan, T.G.; Yuan, J.P.; Wang, J.H. Fatty acid composition of lipids in wild Cordyceps sinensis, from major habitats in China. Biomed. Prev. Nutr. 2012, 2, 42–50. [Google Scholar] [CrossRef]
- Winkler, D. Yartsa gunbu (Cordyceps sinensis) and the fungal commodification of Tibet’s rural economy. Econ. Bot. 2008, 62, 291–305. [Google Scholar] [CrossRef]
- Zhou, X.W.; Li, L.J.; Tian, E.W. Advances in research of the artificial cultivation of Ophiocordyceps sinensis in China. Crit. Rev. Biotechnol. 2014, 34, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.J.; Yin, D.H.; Li, L.; Xia, W.J. Investigation on quality difference between Cordyceps of Sichuan and those of Xizang. China J. Chin. Mater. Medica 2001, 26, 450–452. [Google Scholar]
- Cai, J.Z.; Wang, L.L. Research progress of artificial Cordyceps mycelium. Life Sci. Instrum. 2011, 9, 30–33. [Google Scholar]
- He, X.F.; Zhao, S.R. A review and prospect on the development of Cordyceps mycelium. Qinghai Med. J. 1999, 29, 60–61. [Google Scholar]
- Guo, L.X.; Xu, X.M.; Liang, F.R.; Yuan, J.P.; Peng, J.; Wu, C.F.; Wang, J.H. Morphological observations and fatty acid composition of indoor-cultivated Cordyceps sinensis at a high-altitude laboratory on Sejila Mountain, Tibet. PLoS ONE 2015, 10, e0126095. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xu, Z.L.; Lv, Y. The discrimination of Cordyceps sinensis and its adulterants. Jiangsu J. Agric. Sci. 2015, 43, 273–279. [Google Scholar]
- Pan, X.Y. Identification of Cordyceps sinensis by experience. China Pharm. 2006, 9, 380–381. [Google Scholar]
- Huang, B.; Cheng, Y.L.; Cao, X.J.; Li, Y.; Chen, R.; Cao, J.; Peng, C.; Wan, D.G.; Shen, C.H.; Guo, J.L. HPLC fingerprint of Cordyceps sinensis and its confused species and identification of common composition. Chin. Tradit. Herb. Drugs 2017, 48, 991–996. [Google Scholar]
- Caut, S.; Angulo, E.; Courchamp, F. Variation in discrimination factors (δ15N and δ13C): The effect of diet isotopic values and applications for diet reconstruction. J. Appl. Ecol. 2009, 46, 443–453. [Google Scholar] [CrossRef]
- Qiang, W.Y.; Wang, X.L.; Chen, T.; Feng, H.Y.; An, L.Z.; He, Y.Q.; Wang, G. Variations of stomatal density and carbon isotope values of Picea crassifolia at different altitudes in the Qilian Mountains. Trees-Struct. Funct. 2003, 17, 258–262. [Google Scholar]
- Camin, F.; Larcher, R.; Nicolini, G.; Bontempo, L.; Bertoldi, D.; Perini, M.; Schlicht, C.; Schellenberg, A.; Thomas, F.; Heinrich, K.; et al. Isotopic and elemental data for tracing the origin of European olive oils. J. Agric. Food Chem. 2010, 58, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Horacek, M.; Min, J.S.; Heo, S.C.; Soja, G. Discrimination between ginseng from Korea and China by light stable isotope analysis. Anal. Chim. Acta 2010, 682, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Kropf, U.; Golob, T.; Necemer, M.; Kump, P.; Korosec, M.; Bertoncelj, J.; Ogrinc, N. Carbon and nitrogen natural stable isotopes in Slovene honey: Adulteration and botanical and geographical aspects. J. Agric. Food Chem. 2010, 58, 12794–12803. [Google Scholar] [CrossRef] [PubMed]
- Ehtesham, E.; Hayman, A.R.; McComb, K.A.; Van Hale, R.; Frew, R.D. Correlation of geographical location with stable isotope values of hydrogen and carbon of fatty acids from New Zealand milk and bulk milk powder. J. Agric. Food Chem. 2013, 61, 8914–8923. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.X.; Mai, Z.H.; Zhao, H.; Liu, H.; Tang, H.W. Application of stable carbon isotopes in detection of food adulteration and traceability. Modern Food Sci. Technol. 2016, 32, 281–290. [Google Scholar]
- Chen, D.; Yuan, J.; Xu, S.; Zhou, X.; Zhang, Y.; Xu, X.; Zou, Z.; Zhang, G.; Wang, J. Stable carbon isotope evidence for tracing the diet of the host Hepialus larva of Cordyceps sinensis in the Tibetan Plateau. Sci. China Ser. D Earth Sci. 2009, 52, 655–659. [Google Scholar] [CrossRef]
- Guo, L.X.; Hong, Y.H.; Zhou, Q.Z.; Zhu, Q.; Xu, X.M.; Wang, J.H. Fungus-larva relation in the formation of Cordyceps sinensis as revealed by stable carbon isotope analysis. Sci. Rep. 2017, 7, 7789. [Google Scholar] [CrossRef] [PubMed]
- Kubásek, J.; Urban, O.; Šantrůček, J. C4 plants use fluctuating light less efficiently than do C3 plants: A study of growth, photosynthesis and carbon isotope discrimination. Physiol. Plant. 2013, 149, 528–539. [Google Scholar] [CrossRef] [PubMed]
- Gleixner, G.; Danier, H.J.; Werner, R.A.; Schmidt, H.L. Correlations between the 13C content of primary and secondary plant products in different cell compartments and that in decomposing basidiomycetes. Physiol. Plant. 1993, 102, 1287–1290. [Google Scholar] [CrossRef]
- Ruess, L.; Tiunov, A.; Haubert, D.; Richnow, H.H.; Haggblom, M.M.; Scheu, S. Carbon stable isotope fractionation and trophic transfer of fatty acids in fungal based soil food chains. Soil Biol. Biochem. 2005, 37, 945–953. [Google Scholar] [CrossRef]
- Xia, F.; Chen, X.; Guo, M.Y.; Bai, X.H.; Liu, Y.; Shen, G.R.; Li, Y.L.; Lin, J.; Zhou, X.W. High-throughput sequencing-based analysis of endogenetic fungal communities inhabiting the Chinese Cordyceps reveals unexpectedly high fungal diversity. Sci. Rep. 2016, 6, 33437. [Google Scholar] [CrossRef] [PubMed]
- DeNiro, M.J.; Epstein, S. Mechanism of carbon isotope fractionation associated with lipid synthesis. Science 1977, 197, 261–263. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.M. Fractionation of carbon and hydrogen isotopes in biosynthetic processes. Stable Isot. Geochem. 2001, 43, 225–277. [Google Scholar] [CrossRef]
- Liu, X.; Xu, S.P.; Wang, J.H.; Yuan, J.P.; Guo, L.X.; Li, X.; Huang, X.N. Characterization of ganoderma spore lipid by stable carbon isotope analysis: Implications for authentication. Anal. Bioanal. Chem. 2007, 388, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Hoefs, J. Stable Isotope Geochemistry, 6th ed.; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Sakuradani, E.; Ando, A.; Ogawa, J.; Shimizu, S. Improved production of various polyunsaturated fatty acids through filamentous fungus Mortierella alpina breeding. Appl. Microbiol. Biot. 2009, 84, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kilby, B.A. The biochemistry of the insect fat body. Adv. Insect Physiol. 1963, 1, 111–174. [Google Scholar]
- Stanley-Samuelson, D.W.; Jurenka, R.A.; Cripps, C.; Blomquist, G.J.; Renobales, M.D. Fatty acids in insects: Composition, metabolism, and biological significance. Arch. Insect Biochem. Physiol. 1988, 9, 1–33. [Google Scholar] [CrossRef]
- Araus, J.L.; Villegas, D.; Aparicio, N.; del Moral, L.F.G.; El Hani, S.; Rharrabti, Y.; Ferrio, J.P.; Royo, C. Environmental factors determining carbon isotope discrimination and yield in durum wheat under Mediterranean conditions. Crop Sci. 2003, 43, 170–180. [Google Scholar] [CrossRef]
- Wang, H.; Shan, Y.; Sun, Z. The utilization and analysis of present market situation for Cordyceps sinensis. Chin. Med. J. Res. Pract. 2016, 30, 83–86. [Google Scholar]
- DeNiro, M.J.; Epstein, S. You are what you eat (plus a few per mil): The carbon isotope cycle in food chains. Geol. Soc. Am. Abstr. Prog. 1976, 8, 834–835. [Google Scholar]
- Guo, L.X.; Xu, X.M.; Yuan, J.P.; Wu, C.F.; Wang, J.H. Characterization and authentication of significant Chinese edible oilseed oils by stable carbon isotope analysis. J. Am. Oil Chem. Soc. 2010, 87, 839–848. [Google Scholar] [CrossRef]
Sample Availability: The FAMEs for all samples and substitudes of O. sinensis are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Nos. | Location | Longitude | Latitude | Weight (mg) | Larva Length (cm) | Stroma Length (cm) | Altitude (m) | Relative Humidity (%) |
|---|---|---|---|---|---|---|---|---|
| YN1 | Deqin (Yunnan) | 99°32′20 E | 29°15′02 N | 164.1 | 3.23 | 4.45 | 4250 | 71 |
| YN2 | Deqin (Yunnan) | 99°32′20 E | 29°15′02 N | 530.0 | 4.95 | 2.13 | 4250 | 71 |
| YN3 | Deqin (Yunnan) | 99°32′20 E | 29°15′02 N | 492.3 | 3.71 | 3.26 | 4250 | 71 |
| HM1 | Heimahe (Qinghai) | 99°53′50 E | 36°58′50 N | 137.9 | 2.73 | 3.12 | 3310 | 49 |
| HM2 | Heimahe (Qinghai) | 99°53′50 E | 36°58′50 N | 154.9 | 2.57 | 4.61 | 3310 | 49 |
| HM3 | Heimahe (Qinghai) | 99°53′50 E | 36°58′50 N | 148.9 | 2.47 | 3.61 | 3310 | 49 |
| NQ1 | Naqu (Tibet) | 93°02′20 E | 31°55′02 N | 434.2 | 4.08 | 2.13 | 4800 | 50 |
| NQ2 | Naqu (Tibet) | 93°02′20 E | 31°55′02 N | 479.3 | 3.95 | 3.43 | 4800 | 50 |
| NQ3 | Naqu (Tibet) | 93°02′20 E | 31°55′02 N | 449.3 | 3.85 | 3.13 | 4800 | 50 |
| ML1 | Mila Mountain (Tibet) | 92°19′53 E | 29°50′21 N | 223.4 | 3.18 | 2.67 | 4825 | 46 |
| ML2 | Mila Mountain (Tibet) | 92°19′53 E | 29°50′21 N | 235.1 | 3.21 | 2.95 | 4825 | 46 |
| ML3 | Mila Mountain (Tibet) | 92°19′53 E | 29°50′21 N | 245.7 | 3.98 | 3.89 | 4825 | 46 |
| NM1 | Nam Co (Tibet) | 90°16′50 E | 30°35′18 N | 510.0 | 4.34 | 3.39 | 4805 | 53 |
| NM2 | Nam Co (Tibet) | 92°19′53 E | 29°50′21 N | 381.7 | 4.22 | 3.08 | 4805 | 53 |
| NM3 | Nam Co (Tibet) | 92°19′53 E | 29°50′21 N | 397.1 | 4.32 | 3.28 | 4805 | 53 |
| SJ1 | Sejila Mountain (Tibet) | 94°36′03 E | 29°35′36 N | 282.5 | 4.32 | 4.76 | 4241 | 79 |
| SJ2 | Sejila Mountain (Tibet) | 94°36′03 E | 29°35′36 N | 189.5 | 3.95 | 3.46 | 4241 | 79 |
| SJ3 | Sejila Mountain (Tibet) | 94°36′03 E | 29°35′36 N | 208.5 | 4.15 | 3.26 | 4241 | 79 |
| MZ1 | Maizhokunggar (Tibet) | 91°48′03 E | 29°50′36 N | 322.3 | 3.93 | 3.32 | 4241 | 46 |
| MZ2 | Maizhokunggar (Tibet) | 94°36′04 E | 29°35′37 N | 280.5 | 3.75 | 4.25 | 4241 | 46 |
| MZ3 | Maizhokunggar (Tibet) | 94°36′05 E | 29°35′38 N | 222.3 | 3.25 | 3.45 | 4241 | 46 |
| Sample Nos. | Bulk Sample | Neutral Lipids | Polar Lipids | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C16:0 | C18:0 | C18:1 | C18:2 | C16:0–C18:0 | C18:1–C18:0 | C18:1–C18:2 | C16:0 | C18:0 | C18:1 | C18:2 | C16:0–C18:0 | C18:1–C18:0 | C18:1–C18:2 | ||
| YN1 | −26.0 | −28.5 | −31.4 | −28.4 | −29.7 | 2.9 | 3.0 | 1.3 | −28.2 | −30.9 | −28.7 | −30.4 | 2.7 | 2.2 | 1.7 |
| YN2 | −25.5 | −27.9 | −30.3 | −27.8 | −28.3 | 2.4 | 2.5 | 0.5 | −28.2 | −30.6 | −28.7 | −29.2 | 2.4 | 1.9 | 0.5 |
| YN3 | −27.0 | −29.6 | −30.7 | −29.1 | −29.8 | 1.1 | 1.6 | 0.7 | −29.0 | −31.9 | −29.4 | −30.5 | 2.9 | 2.5 | 1.1 |
| YN AVR ± SD | −26.2 ± 0.8 | −28.7 ± 0.9 | −30.8 ± 0.6 | −28.4 ± 0.7 | −29.3 ± 0.8 | 2.1 | 2.4 | 0.9 | −28.5 ± 0.5 | −31.1 ± 0.7 | −28.9 ± 0.4 | −30.0 ± 0.7 | 2.6 | 2.2 | 1.1 |
| HM1 | −26.2 | −28.7 | −30.2 | −28.6 | −29.5 | 1.5 | 1.6 | 0.9 | −28.2 | −30.8 | −28.1 | −30.2 | 2.6 | 2.7 | 2.1 |
| HM2 | −26.2 | −28.7 | −30.1 | −29.5 | −30.2 | 1.4 | 0.6 | 0.7 | −29.7 | −31.9 | −28.7 | −29.7 | 2.2 | 3.2 | 1.0 |
| HM3 | −26.8 | −28.4 | −30.3 | −29.1 | −30.0 | 1.9 | 1.2 | 0.9 | −29.3 | −31.5 | −28.3 | −29.2 | 2.2 | 3.2 | 0.9 |
| HM AVR ± SD | −26.4 ± 0.4 | −28.6 ± 0.2 | −30.2 ± 0.1 | −29.1 ± 0.5 | −29.9 ± 0.4 | 1.6 | 1.1 | 0.8 | −29.1 ± 0.8 | −31.4 ± 0.6 | −28.4 ± 0.3 | −29.7 ± 0.5 | 2.3 | 3.0 | 1.3 |
| NQ1 | −25.8 | −28.2 | −29.7 | −27.9 | −28.4 | 1.5 | 1.8 | 0.5 | −28.4 | −30.8 | −28.2 | −29.4 | 2.4 | 2.6 | 1.2 |
| NQ2 | −25.6 | −28.0 | −29.7 | −28.1 | −29.0 | 1.7 | 1.6 | 0.9 | −29.3 | −31.5 | −29.7 | −30.3 | 2.2 | 1.8 | 0.6 |
| NQ3 | −25.3 | −28.4 | −29.2 | −28.4 | −28.8 | 0.8 | 0.8 | 0.4 | −28.5 | −31.0 | −29.2 | −30.0 | 2.5 | 1.8 | 0.8 |
| NQ AVR ± SD | −25.6 ± 0.3 | −28.2 ± 0.2 | −29.5 ± 0.3 | −28.1 ± 0.3 | −28.7 ± 0.3 | 1.3 | 1.4 | 0.6 | −28.7 ± 0.5 | −31.1 ± 0.4 | −29.0 ± 0.8 | −29.9 ± 0.5 | 2.4 | 2.1 | 0.9 |
| ML1 | −26.7 | −29.3 | −30.7 | −30.2 | −30.7 | 1.4 | 0.5 | 0.5 | −30.1 | −31.4 | −30.9 | −31.1 | 1.3 | 0.5 | 0.2 |
| ML2 | −26.5 | −29.1 | −30.2 | −30.5 | −30.8 | 1.1 | -0.3 | 0.3 | −28.7 | −30.5 | −29.7 | −30.1 | 1.8 | 0.8 | 0.4 |
| ML3 | −26.7 | −29.3 | −30.2 | −29.5 | −29.9 | 0.9 | 0.7 | 0.4 | −29.8 | −31.7 | −28.1 | −29.8 | 1.9 | 3.6 | 1.7 |
| ML AVR ± SD | −26.6 ± 0.1 | −29.2 ± 0.1 | −30.4 ± 0.3 | −30.1 ± 0.5 | −30.5 ± 0.5 | 1.2 | 0.3 | 0.4 | −29.5 ± 0.7 | −31.2 ± 0.6 | −29.6 ± 1.4 | −30.3 ± 0.7 | 1.7 | 1.6 | 0.7 |
| NM1 | −25.8 | −28.2 | −29.5 | −28.8 | −29.8 | 1.3 | 0.7 | 1.0 | −28.8 | −30.9 | −27.5 | −29.2 | 2.1 | 3.4 | 1.7 |
| NM2 | −25.8 | −28.2 | −29.3 | −28.2 | −29.4 | 1.1 | 1.1 | 1.2 | −29.0 | −30.5 | −27.9 | −29.7 | 1.5 | 2.6 | 1.8 |
| NM3 | −25.5 | −27.8 | −28.8 | −28.5 | −29.0 | 1.0 | 0.3 | 0.5 | −29.4 | −30.1 | −27.9 | −29.4 | 0.7 | 2.2 | 1.5 |
| NM AVR ± SD | −25.7 ± 0.2 | −28.1 ± 0.2 | −29.2 ± 0.4 | −28.5 ± 0.3 | −29.4 ± 0.4 | 1.1 | 0.7 | 0.9 | −29.1 ± 0.3 | −30.5 ± 0.4 | −27.8 ± 0.2 | −29.4 ± 0.3 | 1.4 | 2.7 | 1.6 |
| SJ1 | −26.2 | −28.7 | −30.1 | −28.6 | −28.9 | 1.4 | 1.5 | 0.3 | −28.7 | −31.0 | −28.8 | −29.5 | 2.3 | 2.2 | 0.7 |
| SJ2 | −27.5 | −30.2 | −30.6 | −28.7 | −30.0 | 0.4 | 1.9 | 1.3 | −28.8 | −30.5 | −28.2 | −30.1 | 1.7 | 2.3 | 1.9 |
| SJ3 | −27.0 | −30.3 | −30.3 | −28.5 | −30.2 | 0.0 | 1.8 | 1.7 | −28.5 | −30.3 | −28.1 | −30.1 | 1.8 | 2.2 | 2.0 |
| SJ AVR ± SD | −26.9 ± 0.7 | −29.7 ± 0.9 | −30.3 ± 0.3 | −28.6 ± 0.1 | −29.7 ± 0.7 | 0.6 | 1.7 | 1.1 | −28.7 ± 0.2 | −30.6 ± 0.4 | −28.4 ± 0.4 | −29.9 ± 0.4 | 1.9 | 2.2 | 1.5 |
| MZ1 | −25.6 | −28.0 | −29.3 | −27.9 | −29.7 | 1.3 | 1.4 | 1.8 | −29.3 | −30.5 | −29.1 | −30.6 | 1.2 | 1.4 | 1.5 |
| MZ2 | −25.7 | −28.2 | −29.5 | −28.0 | −30.0 | 1.3 | 1.5 | 2.0 | −29.5 | −30.7 | −29.1 | −30.8 | 1.2 | 1.6 | 1.7 |
| MZ3 | −25.8 | −28.8 | −29.1 | −28.4 | −30.2 | 0.3 | 0.7 | 1.8 | −29.5 | −30.4 | −29.4 | −30.5 | 0.9 | 1.0 | 1.1 |
| MZ AVR ± SD | −25.7 ± 0.1 | −28.3 ± 0.4 | −29.3 ± 0.2 | −28.1 ± 0.3 | −30.0 ± 0.3 | 1.0 | 1.2 | 1.9 | −29.4 ± 0.1 | −30.5 ± 0.2 | −29.2 ± 0.2 | −30.6 ± 0.2 | 1.1 | 1.3 | 1.4 |
| Sample Nos. ** | Neutral Lipids | Polar Lipids | ||||||
|---|---|---|---|---|---|---|---|---|
| C16:0 | C18:0 | C18:1 | C18:2 | C16:0 | C18:0 | C18:1 | C18:2 | |
| SCMN 1 | −31.0 | −31.5 | −30.4 | −30.4 | −30.5 | −30.8 | −30.4 | −30.4 |
| SCMN 2 | −30.7 | −31.6 | −30.5 | −30.6 | −30.6 | −30.9 | −30.5 | −30.6 |
| SCMN 3 | −31.5 | −32.0 | −30.9 | −31.1 | −31.2 | −31.6 | −31.0 | −31.1 |
| SCMN AVR ± SD | −31.1 ± 0.4 | −31.7 ± 0.3 | −30.6 ± 0.3 | −30.7 ± 0.4 | −30.8 ± 0.4 | −31.1 ± 0.4 | −30.6 ± 0.4 | −30.7 ± 0.4 |
| SCMS 1 | −31.4 | −31.7 | −30.0 | −30.3 | −31.2 | −31.2 | −30.0 | −30.1 |
| SCMS 2 | −31.2 | −31.4 | −29.8 | −30.5 | −30.9 | −31.1 | −29.8 | −30.0 |
| SCMS 3 | −31.8 | −31.9 | −29.9 | −30.2 | −31.1 | −31.4 | −29.7 | −29.9 |
| SCMS AVR ± SD | −31.5 ± 0.3 | −31.7 ± 0.3 | −29.9 ± 0.1 | −30.3 ± 0.2 | −31.1 ± 0.2 | −31.2 ± 0.2 | −29.8 ± 0.2 | −30.0 ± 0.1 |
| FMH 1 | −14.7 | −16.2 | −16.4 | −16.6 | −14.5 | −15.9 | −16.4 | −16.5 |
| FMH 2 | −14.6 | −16.4 | −16.3 | −16.6 | −14.3 | −16.0 | −16.3 | −16.6 |
| FMH 3 | −14.9 | −16.6 | −16.7 | −16.8 | −14.5 | −16.3 | −16.8 | −16.9 |
| FMH AVR ± SD | −14.7 ± 0.2 | −16.4 ± 0.2 | −16.5 ± 0.2 | −16.7 ± 0.1 | −14.4 ± 0.1 | −16.1 ± 0.2 | −16.5 ± 0.3 | −16.7 ± 0.2 |
| FMP 1 | −24.5 | −24.8 | −25.0 | −26.5 | −24.2 | −24.4 | −25.3 | −26.2 |
| FMP 2 | −24.3 | −24.8 | −24.8 | −26.3 | −23.9 | −24.2 | −25.2 | −26.0 |
| FMP 3 | −24.2 | −24.6 | −24.7 | −26.4 | −24.0 | −24.3 | −25.1 | −26.1 |
| FMP AVR ± SD | −24.3 ± 0.2 | −24.7 ± 0.1 | −24.8 ± 0.2 | −26.4 ± 0.1 | −24.0 ± 0.2 | −24.3 ± 0.1 | −25.2 ± 0.1 | −26.1 ± 0.1 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, L.-X.; Xu, X.-M.; Hong, Y.-H.; Li, Y.; Wang, J.-H. Stable Carbon Isotope Composition of the Lipids in Natural Ophiocordyceps sinensis from Major Habitats in China and Its Substitutes. Molecules 2017, 22, 1567. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22091567
Guo L-X, Xu X-M, Hong Y-H, Li Y, Wang J-H. Stable Carbon Isotope Composition of the Lipids in Natural Ophiocordyceps sinensis from Major Habitats in China and Its Substitutes. Molecules. 2017; 22(9):1567. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22091567
Chicago/Turabian StyleGuo, Lian-Xian, Xiao-Ming Xu, Yue-Hui Hong, Yan Li, and Jiang-Hai Wang. 2017. "Stable Carbon Isotope Composition of the Lipids in Natural Ophiocordyceps sinensis from Major Habitats in China and Its Substitutes" Molecules 22, no. 9: 1567. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22091567