Anticancer Activity of Toxins from Bee and Snake Venom—An Overview on Ovarian Cancer

Abstract
:1. Introduction
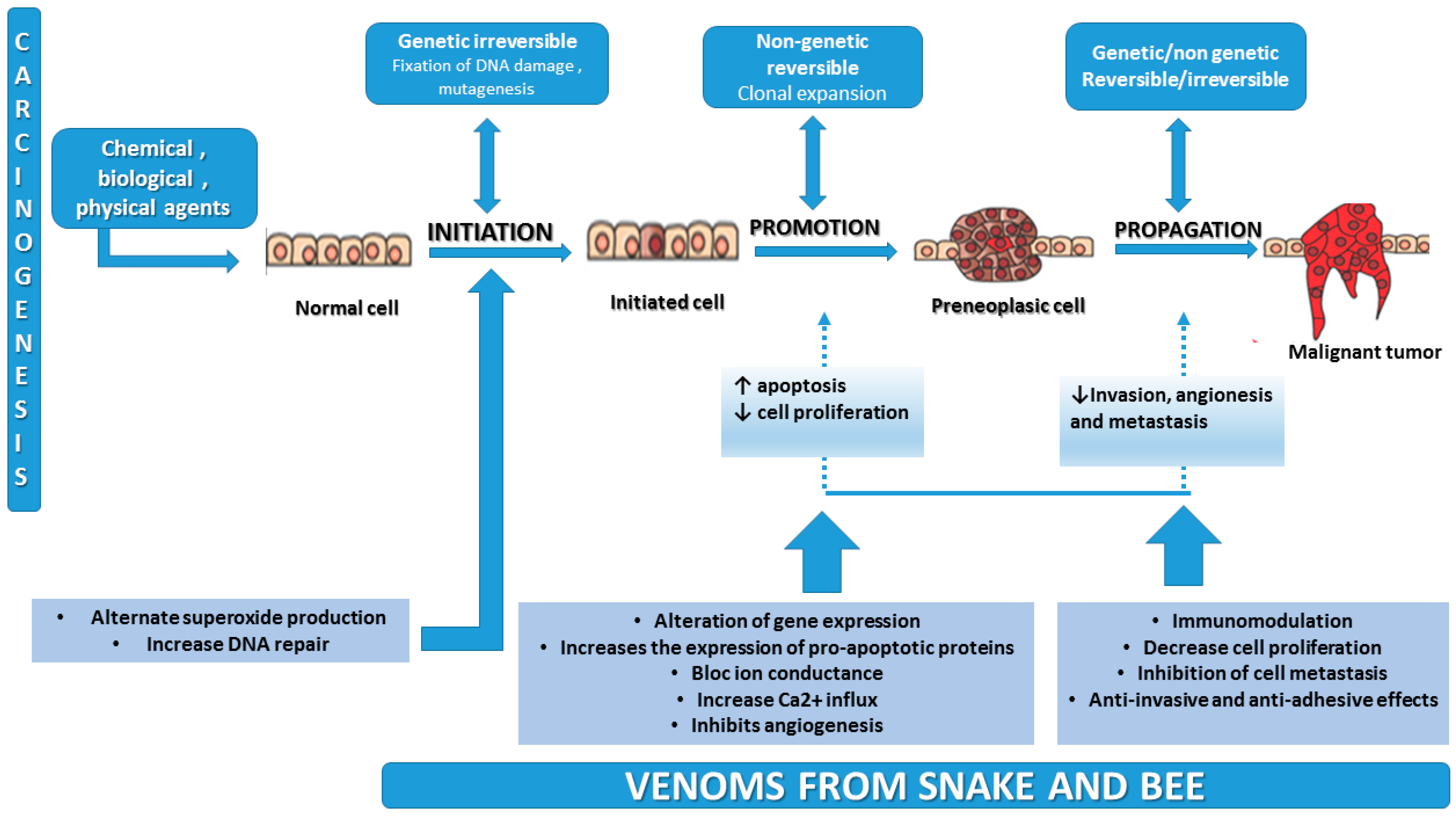
2. Carcinogenesis Mechanism
3. Toxins from Animal Venom—Compounds and Activity
3.1. Bee Venom
Compounds and Activity of Bee Venom
3.2. Snake Venom
- Neurotoxins act at the level of the central nervous system, causing breathing difficulties or heart failure. These toxins affect the cholinergic neurons and block the binding of acetylcholine. Another effect is the inhibition of ion movement through the cell membrane, which blocks the communication between neurons [52].
- The toxins from snake venom that cause the destruction of the red blood cells are known as hemotoxins and mainly affect the circulatory system, blood function, and muscle tissue (causing gangrene and scarring). Viperidae species members (copperhead, rattlesnake, and cotton head) possess hemotoxic venom while mambas, cobras, krait, sea snakes, and coral snakes have neurotoxic venom. However, some species contain both neurotoxins and hemotoxins.
- The third group, the cardiotoxins, are those toxins that affect the heart muscle, binding to the cells of the heart and blocking muscle contraction [53].
- One of the most important and researched toxins from snake venom is the cytotoxin. It targets specific cellular sites, affecting the cell membrane or interfering with the transport of substances or the transduction of signals across the membranes [54].
Compounds and Activity of Snake Venom
4. Effects of Bee and Snake Venom on Cancer Cells
4.1. The Effects of Bee Venom on Cancer Cells
4.2. The Effects of Snake Venom on Cancer Cells
5. Studies Regarding the Effects of Toxins from Bee and Snake Venom on Ovarian Cancer Cells
5.1. Bee Venom and Ovarian Carcinoma
5.2. Snake Venom and Ovarian Carcinoma
6. Conclusions and Future Perspectives
Author Contributions
Conflicts of Interest
Abbreviations
| AIF | apoptosis-inducing factor |
| Akt | protein kinase B |
| ASR | age standardized rate |
| Bax | BCL2-associated X protein |
| Bcl-2 | B-cell lymphoma 2 |
| bFGF | basic fibroblast growth factor |
| BJcuL | lectin from the venom of the snake Bothrops jararacussu |
| BjV | Bothrops jararaca venom |
| BV | bee venom |
| CN | contortrostatin |
| COX | cyclooxygenase |
| CTX-3 | cardiotoxin-3 |
| CVF | cobra venom factor |
| DR | death receptor |
| DU 145 | a prostate cancer cell line |
| EAT | Ehrlich ascites tumor |
| ECM | extra cellular matrix |
| EndoG | endonuclease G |
| ER | endoplasmic reticulum |
| ERK | extracellular signal-regulated kinase |
| GRP 78 | protein 78 |
| hIL-2 | human interleukin-2 |
| IL-2 | interleukin-2 |
| JAK2 | Janus-associated kinase 2 |
| JNK | c-Jun N-terminal kinases |
| LAAO | l-amino acid oxidase |
| LCN | liposomal encapsulation of contortrostatin |
| MAPK | mitogen-activated protein kinase |
| MCD | mast-cell degranulating peptide |
| MCF-7 | human breast adenocarcinoma cell line |
| MCF7- cells | human breast adenocarcinoma cell line |
| MhIL-2 | melittin human interleukine-2 |
| MMP | matrix metalloproteinases |
| NF-κB | nuclear factor-kappa B |
| NR | not reported |
| OVCAR 5 | human epithelial carcinoma cell line of the ovary |
| PLA2 | phospholipase A2 |
| RGD | arginyl-glycyl-aspartic acid |
| rhIl-2 | recombinant human IL-2 protein |
| STAT 3 | signal transducer and activator of transcription 3 |
| TCA | tricarboxylic acid |
| Th1 | lymphocyte T helper 1 |
| TNF-α | tumor necrosis factor-α |
| VEGF | vascular endothelial growth factor |
| VEGFR | vascular endothelial growth factor receptor |
References
- Reid, B.M.; Permuth, J.B.; Sellers, T.A. Epidemiology of ovarian cancer: A review. Cancer Biol. Med. 2017, 14, 9. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.A.; Manassaram-Baptiste, D.; Brooks, D.; Doroshenk, M.; Fedewa, S.; Saslow, D.; Brawley, O.W.; Wender, R. Cancer screening in the United States, 2015: A review of current American cancer society guidelines and current issues in cancer screening. CA Cancer J. Clin. 2015, 65, 30–54. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, I.; Chon, H.S.; Apte, S.M. The role of surgery in the management of epithelial ovarian cancer. Cancer Control 2011, 18, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.; Calò, V.; Bruno, L.; Rizzo, S.; Bazan Di Fede, G.V. Hereditary ovarian cancer. Crit. Rev. Oncol. Hematol. 2009, 69, 28–44. [Google Scholar] [CrossRef] [PubMed]
- Sabatier, R.; Finetti, P.; Cervera, N.; Birnbaum, D.; Bertucci, F. Gene expression profiling and prediction of clinical outcome in ovarian cancer. Crit. Rev. Oncol. Hematol. 2009, 72, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.; Visser-Grieve, S.; Yang, X. Tumour suppressor genes in chemotherapeutic drug response. Biosci. Rep. 2012, 32, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Serna, N.; Sánchez-García, L.; Unzueta, U.; Díaz, R.; Vázquez, E.; Mangues, R.; Villaverde, A. Protein-Based Therapeutic Killing for Cancer Therapies. Trends Biotechnol. 2017, 36, 318–335. [Google Scholar] [CrossRef] [PubMed]
- Mahadevappa, R.; Ma, R.; Kwok, H.F. Venom Peptides: Improving Specificity in Cancer Therapy. Trends Cancer 2017, 3, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Fantini, M.; Benvenuto, M.; Masuelli, L.; Frajese, G.V.; Tresoldi, I.; Modesti, A.; Bei, R. In vitro and in vivo antitumoral effects of combinations of polyphenols, or polyphenols and anticancer drugs: Perspectives on cancer treatment. Int. J. Mol. Sci. 2015, 16, 9236–9282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roleira, F.M.; Tavares-da-Silva, E.J.; Varela, C.L.; Costa, S.C.; Silva, T.; Garrido, J.; Borges, F. Plant derived and dietary phenolic antioxidants: Anticancer properties. Food Chem. 2015, 183, 235–258. [Google Scholar] [CrossRef] [PubMed]
- Gali-Muhtasib, H.; Hmadi, R.; Kareh, M.; Tohme, R.; Darwiche, N. Cell death mechanisms of plant-derived anticancer drugs: Beyond apoptosis. Apoptosis 2015, 20, 1531–1562. [Google Scholar] [CrossRef] [PubMed]
- Shani, K.H.; Zubair, M.; Rizwan, K.; Rasool, N.; Ercisli, S.; Mahmood, A.; Zia-Ul-Haq, M.; Dima, L.; Pascu, A.M. Compositional studies of oil and antioxidant capacity of oil and extracts of Diarthron vesiculosum. Oxid. Commun. 2015, 38, 1252–1264. [Google Scholar]
- Huang, H.; Chen, A.Y.; Rojanasakul, Y.; Rankin, G.O.; Chen, Y.C. Dietary compounds galangin and myricetin suppress ovarian cancer cell angiogenesis. J. Funct. Foods 2015, 15, 464–475. [Google Scholar] [CrossRef] [PubMed]
- Liberio, S.M.; Joanitti, G.A.; Fontes, W.; Castro, M.S. Anticancer peptides and proteins: A panoramic view. Protein Pept. Lett. 2013, 20, 380–391. [Google Scholar] [CrossRef] [PubMed]
- McLeod, H.L. Cancer pharmacogenomics: Early promise, but concerted effort needed. Science 2013, 339, 1563–1566. [Google Scholar] [CrossRef] [PubMed]
- Moga, M.A.; Dimienescu, O.G.; Arvatescu, C.A.; Mironescu, A.; Dracea, L.; Ples, L. The role of natural polyphenols in the prevention and treatment of cervical cancer—An overview. Molecules 2016, 21, 1055. [Google Scholar] [CrossRef] [PubMed]
- Chaisakul, J.; Hodgson, W.C.; Kuruppu, S.; Prasongsook, N. Effects of animal venoms and toxins on hallmarks of cancer. J. Cancer 2016, 7, 1571. [Google Scholar] [CrossRef] [PubMed]
- Son, D.J.; Leea, J.W.; Lee, Y.H.; Song, H.S.; Leea, C.K.; Hong, J.T. Therapeutic application of anti-arthritis, pain-releasing, and anti-cancer effects of bee venom and its constituent compounds. Pharmacol. Ther. 2007, 115, 246–270. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Lee, S.H.; Son, D.J.; Oh, K.W.; Kim, K.H.; Song, H.S.; Kim, G.J.; Oh, G.T.; Yoon, D.Y.; Hong, J.T. Antiarthritic effect of bee venom: Inhibition of inflammation mediator generation by suppression of NF-κB through interaction with the p50 subunit. Arthritis Rheum. 2004, 50, 3504–3515. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Nakatani, Y.; Atsumi, G.; Inoue, K.; Kudo, I. Regulatory functions of phospholipase A2. Crit. Rev. Immunol. 1997, 17, 225–283. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, J.P.; Jovanovic, D.; Fernandes, J.C.; Manning, P.; Connor, J.R.; Currie, M.G.; Di Battista, J.A.; Martel-Pelletier, J. Reduced progression of experimental osteoarthritis in vivo by selective inhibition of inducible nitric oxide synthase. Arthritis Rheum. 1998, 41, 1275–1286. [Google Scholar] [CrossRef]
- Jang, M.H.; Shin, M.C.; Lim, S.; Han, S.M.; Park, H.J.; Shin, I.; Lee, J.S.; Kim, K.A.; Kim, E.H.; Kim, C.J. Bee venom induces apoptosis and inhibits expression of cyclooxygenase-2 mRNA in human lung cancer cell line NCI-H1299. Jpn. J. Pharmacol. 2003, 91, 95–104. [Google Scholar] [CrossRef]
- Amin, A.R.; Attur, M.; Abramson, S.B. Nitric oxide synthase and cyclooxygenases: Distribution, regulation, and intervention in arthritis. Curr. Opin. Rheumatol. 1999, 11, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Ip, S.W.; Liao, S.S.; Lin, S.Y.; Lin, J.P.; Yang, J.S.; Lin, M.L.; Chen, G.W.; Lu, H.F.; Weillin, M.; Mihan, S.; et al. The role of mitochondria in bee venom-induced apoptosis in human breast cancer MCF7 cells. In Vivo 2008, 22, 237–245. [Google Scholar] [PubMed]
- Jo, M.; Park, M.H.; Kollipara, P.S.; An, B.J.; Song, H.S.; Han, S.B.; Kim, J.H.; Song, M.J.; Hong, J.T. Anti-cancer effect of bee venom toxin and melittin in ovarian cancer cells through induction of death receptors and inhibition of JAK2/STAT3 pathway. Toxicol. Appl. Pharm. 2012, 258, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Ip, S.W.; Chu, I.L.; Yu, C.S.; Chen, P.Y.; Ho, H.C.; Yang, J.S.; Huang, H.Y.; Chueh, F.S.; Lai, T.Y.; Chung, J.G. Bee venom induces apoptosis through intracellular Ca2+-modulated intrinsic death pathway in human bladder cancer cells. Int. J. Urol. 2012, 19, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.E.; Baek, Y.H.; Lee, M.H.; Choi, D.Y.; Park, D.S.; Lee, J.D. Bee venom inhibits tumor angiogenesis and metastasis by inhibiting tyrosine phosphorylation of VEGFR-2 in LLC-tumor-bearing mice. Cancer Lett. 2010, 292, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Yu, M.; He, Y.; Xiao, L.; Wang, F.; Song, C.; Sun, S.; Ling, C.; Xu, Z. Melittin prevents liver cancer cell metastasis through inhibition of the Rac1, dependent pathway. Hepathology 2008, 47, 1964–1973. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Choi, M.S.; Kwak, D.H.; Oh, K.W.; Yoon, D.Y.; Han, S.B.; Song, H.S.; Song, M.J.; Hong, J.T. Anti-cancer effect of bee venom in prostate cancer cells through activation of caspase pathway via inactivation of NF-κB. Prostate 2011, 71, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Shimpi, R.; Chaudhari, P.; Deshmukh, R.; Devare, S.; Bagad, Y.; Bhurat, M. A review: Pharmacotherapeutics of bee venom. World. J. Pharm. Pharm. Sci. 2016, 5, 656–667. [Google Scholar] [CrossRef]
- Lariviere, W.R.; Melzack, R. The bee venom test: A new tonic-pain test. Pain 1996, 66, 271–277. [Google Scholar] [CrossRef]
- Ali, E.M. Contributions of some biological activities of honey bee venom. J. Apic. Res. 2014, 53, 441–451. [Google Scholar] [CrossRef]
- Gevod, V.S.; Birdi, K.S. Melittin and the 8–26 fragment. Differences in ionophoric properties as measured by monolayer method. Biophys. J. 1984, 45, 1079–1083. [Google Scholar] [CrossRef]
- Ma, R.; Mahadevappa, R.; Kwok, H.F. Venom-based peptide therapy: Insights into anti-cancer mechanism. Oncotarget 2017, 8, 100908–100930. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Jeong, Y.J.; Park, K.K.; Cho, H.J.; Chung, I.K.; Min, K.S.; Kim, M.; Lee, K.G.; Yeo, J.H.; Park, K.K.; et al. Melittin suppresses PMA-induced tumor cell invasion by inhibiting NF-κB and AP-1-dependent MMP-9 expression. Mol. Cells 2010, 29, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Cornara, L.; Biagi, M.; Xiao, J.; Burlando, B. Therapeutic properties of bioactive compounds from different honeybee products. Front. Pharmacol. 2017, 8, 412. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Jeong, Y.J.; Park, K.K.; Cho, H.I.; Chung, I.K.; Min, K.S.; Kim, M.; Lee, K.G.; Yeo, J.H.; Park, K.K.; et al. Melittin prevents liver cancer cell metastasis through inhibition of the Rac1-dependent pathway. Hepatology 2008, 47, 1964–1973. [Google Scholar] [CrossRef]
- Lazdunski, M.; Fosset, M.; Hugues, M.; Mourre, C.; Renaud, J.F.; Romey, G.; Schmid-Antomarchi, H. The apamin-sensitive Ca2+-dependent K+ channel molecular properties, differentiation and endogenous ligands in mammalian brain. Biochem. Soc. Symp. 1985, 50, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.M.; Maylie, J.; Adelman, J.P. Determinants of Apamin and d-tubocurarine block in SK potassium channels. J. Biol. Chem. 1997, 272, 23195–23200. [Google Scholar] [CrossRef] [PubMed]
- Modzelewska, B.; Kostrzewska, A.; Sipowicz, M.; Kleszczewski, T.; Batra, S. Apamin inhibits NO-induced relaxation of the spontaneous contractile activity of the myometrium from non-pregnant women. Reprod. Biol. Endocrinol. 2003, 1. [Google Scholar] [CrossRef]
- Buku, A. Mast cell degranulating peptide: A prototypic peptide in allergy and inflammation. Peptides 1999, 20, 415–420. [Google Scholar] [CrossRef]
- Mousli, M.; Bueb, J.L.; Bronner, J.C.; Rouot, B.; Landry, Y. G protein activation: A receptor independent mode of action for cationic amphiphilic neuropeptides, and venom peptides. Trends Pharmacol. Sci. 1990, 11, 358–362. [Google Scholar] [CrossRef]
- Buku, A.; Mendlowitz, M.; Condie, B.A.; Price, J.A. Partial alanine scan of mast cell degranulating peptide (MCD): Importance of the histidineand arginine-residues. J. Pept. Sci. 2004, 10, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Pak, S.C. Chemical composition of bee venom. In Bee Products-Chemical and Biological Properties; Springer: Cham, Switzerland, 2017; pp. 279–285. [Google Scholar]
- Eze, O.B.L.; Nwodo, O.F.C.; Ogugua, V.N. Therapeutic effect of honey bee venom. J. Pharm. Chem. Biol. Sci. 2016, 4, 48–53. [Google Scholar]
- Oršolić, N. Bee venom in cancer therapy. Cancer Metast. Rev. 2012, 31, 173–194. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.; Choudhury, S.R.; Saha, A.; Mishra, R.; Giri, B.; Biswas, A.K.; Debnath, A.; Gomes, A. A heat stable protein toxin (drCT-I) from the Indian Viper (Daboia russelli russelli) venom having antiproliferative, cytotoxic and apoptotic activities. Toxicon 2007, 49, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Leon, G.; Sanchez, L.; Hernandez, A.; Villalta, M.; Herrera, M.; Segura, A.; Estrada, R.; Maria Gutierrez, J. Immune response towards snake venoms. Inflamm. Allergy Drug Targets 2011, 10, 381–398. [Google Scholar] [CrossRef] [PubMed]
- Tashima, A.K.; Sanz, L.; Camargo, A.C.; Serrano, S.M.; Calvete, J.J. Snake venomics of the Brazilian pitvipers, Bothrops cotiara and Bothrops fonsecai. Identification of taxonomy markers. J. Proteom. 2008, 71, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Doley, R.; Kini, M. Protein complexes in snake venom. Cell. Mol. Life Sci. 2009, 66, 2851–2871. [Google Scholar] [CrossRef] [PubMed]
- Vonk, F.J.; Jackson, K.; Doley, R.; Madaras, F.; Mirtschin, P.J.; Vidal, N. Snake venom: From fieldwork to the clinic. Bioessays 2011, 33, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Chien, C.M.; Lu, M.C.; Lu, Y.J.; Wu, Z.Z.; Lin, S.R. Cardiotoxin III induces apoptosis in K562 cells through a mitochondrial-mediated pathway. Clin. Exp. Pharmacol. Physiol. 2005, 32, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Morita, T. Snake venom components affecting blood coagulation and the vascular system: Structural similarities and marked diversity. Curr. Pharm. Des. 2007, 13, 2872–2886. [Google Scholar] [CrossRef] [PubMed]
- Vyas, V.K.; Brahmbhatt, K.; Bhatt, H.; Parmar, U. Therapeutic potential of snake venom in cancer therapy: Current perspectives. Asian Pac. J. Trop. Biomed. 2013, 3, 156–162. [Google Scholar] [CrossRef]
- Ojeda, P.G.; Ramírez, D.; Alzate-Morales, J.; Caballero, J.; Kaas, Q.; González, W. Computational Studies of Snake Venom Toxins. Toxins 2017, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Muller, V.D.; Russo, R.R.; Cintra, A.C.; Sartim, M.A.; Alves-Paiva, R.D.; Figueiredo, L.T.; Sampaio, S.V.; Aquino, V.H. Crotoxin and phospholipases A2 from Crotalus durissus terrificus showed antiviral activity against dengue and yellow fever viruses. Toxicon 2012, 59, 507–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubbard, S.; Choudhary, S.; Maus, E.; Shukla, D.; Swenson, S.; Markland, F.S.; Tiwari, V. Contortrostatin, a homodimeric disintegrin isolated from snake venom inhibits herpes simplex virus entry and cell fusion. Antivir. Ther. 2012, 17, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Samy, R.P.; Gopalakrishnakone, P.; Stiles, B.G. Snake venom phospholipases A2: A novel tool against bacterial diseases. Curr. Med. Chem. 2012, 19, 6150–6162. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Bao-Jueng, W.U.; Pei-Hsiu, K. Antibacterial and membrane-damaging activities of β-bungarotoxin B chain. J. Pept. Sci. 2013, 19, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yamane, E.S.; Fernando, C.B.A.; Oliveira, E.B. Unraveling the antifungal activity of a South American rattlesnake toxin crotamine. Biochimie 2013, 95, 231–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo, J.C.; Vargas, L.J.; Segura, C. In vitro antiplasmodial activity of phospholipases A2 and a phospholipase homologue isolated from the venom of the snake Bothrops asper. Toxins 2012, 4, 1500–1516. [Google Scholar] [CrossRef] [PubMed]
- De Vieira Santos, M.M.; Sant’Ana, C.D.; Giglio, J.R.; da Silva, R.J.; Sampaio, S.V.; Soares, A.M.; Fecchio, D. Antitumor effect of an L-amino acid oxidase isolated from Bothrops jararaca snake venom. Basic Clin. Pharmacol. Toxicol. 2008, 102, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.S.; Georgieva, D.; Genov, N.; Murakami, M.T.; Sinha, M.; Kumar, R.P.; Kaur, P.; Kumar, S.; Dey, S.; Sharma, S.; et al. Enzymatic toxin from snake venom: Structural characterization and mechanism of catalysis. FEBS J. 2011, 278, 4544–4576. [Google Scholar] [CrossRef] [PubMed]
- Cohen, O.; Kronman, C.; Chitlaru, T.; Ordentlich, A.; Velan, B.; Shafferman, A. Effect of chemical modification of recombinant human acetylcholinesterase by polyethylene glycol on its circulatory longevity. Biochem. J. 2011, 357, 795–802. [Google Scholar] [CrossRef]
- Lokeshwar, V.B.; Selzer, M.G. Hyalurondiase: Both a tumor promoter and suppressor. Semin. Cancer Biol. 2008, 18, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Kemparaju, K.; Girish, K.S. Snake venom hyaluronidase: A therapeutic target. Cell Biochem. Funct. 2006, 24, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, R.S.; Izidoro, L.F.; de Oliveira, R.J., Jr.; Sampaio, S.V.; Soares, A.M.; Rodrigues, V.M. Snake venom phospholipases A2: A new class of antitumor agents. Protein Pept. Lett. 2009, 16, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Pan, H.; Liao, K.; Yang, M.; Huang, C. Snake Venom PLA2, a Promising Target for Broad-Spectrum Antivenom Drug Development. BioMed Res. Int. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Waheed, H.F.; Moin, S.I.; Choudhary, M. Snake venom: From deadly toxins to life-saving therapeutics. Curr. Med. Chem. 2017, 24, 1874–1891. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.W.; Tsai, T.S.; Tsai, I.H.; Lin, Y.S.; Tu, M.C. Prey envenomation does not improve digestive performance in Taiwanese pit vipers (Trimeresurus gracilis and T. stejnegeri stejnegeri). Comp. Biochem. Physiol. 2009, 152, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Armugam, A.; Cher, C.D.; Lim, K.; Koh, D.C.; Howells, D.W.; Jeyaseelan, K. A secretory phospholipase A2-mediated neuroprotection and anti-apoptosis. BMC Neurosci. 2009, 10, 120. [Google Scholar] [CrossRef] [PubMed]
- Pawelek, P.D.; Cheah, J.; Coulombe, R.; Macheroux, P.; Ghisla, S.; Vrielink, A. The structure of L-amino acid oxidase reveals the substrate trajectory into an enantiomerically conserved active site. EMBO J. 2000, 19, 4204–4215. [Google Scholar] [CrossRef] [PubMed]
- Koh, D.C.I.; Armugam, A.; Jeyaseelan, K. Snake venom components and their applications in biomedicine. Cell. Mol. Life Sci. 2006, 63, 3030–3041. [Google Scholar] [CrossRef] [PubMed]
- Fatima, L.; Fatah, C. Pathophysiological and Pharmacological Effects of Snake Venom Components: Molecular Targets. J. Clin. Toxicol. 2014, 4. [Google Scholar] [CrossRef]
- Gajski, G.; Garaj-Vrhovac, V. Melittin: A lytic peptide with anticancer properties. Environ. Toxicol. Pharm. 2013, 36, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.; Giralt, E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: Melittin, apamin and mastoparan. Toxins 2015, 7, 1126–1150. [Google Scholar] [CrossRef] [PubMed]
- Holle, L.; Song, W.; Holle, E.; Wei, Y.; Wagner, T.; Yu, X. A matrix metalloproteinase 2 cleavable melittin/avidin conjugate specifically target tumor cells in vitro and in vivo. Int. J. Oncol. 2003, 22, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.O.; Park, S.Y.; Heo, M.S.; Kim, K.C.; Park, C.; Ko, W.S.; Choi, Y.H.; Kim, G.Y. Key regulators in bee venom-induced apoptosis are Bcl-2 and caspase-3 in human leukemic U937 cells through downregulation of ERK and Akt. Int. Immnunol. 2006, 6, 1796–1807. [Google Scholar] [CrossRef] [PubMed]
- Russell, P.M.; Hewish, D.; Carter, T.; Sterling-Levis, K.; Ow, K.; Hattarki, M.; Doughty, L.; Guthrie, R.; Shapira, D.; Molloy, P.L.; et al. Cytotoxic properties of immunoconjugates containing melittin-like peptide 101 against prostate cancer: In vitro and in vivo studies. Cancer Immunol. Immunother. 2004, 53, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Ling, C.Q.; Li, B.; Zhang, C.; Zhu, D.Z.; Huang, X.Q.; Gu, W.; Li, S.X. Inhibitory effect of recombinant adenovirus carrying melittin gene on hepatocellular carcinoma. Ann. Oncol. 2005, 16, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Oršolić, N.; Šver, L.; Verstovšek, S.; Terzić, S.; Bašić, I. Inhibition of mammary carcinoma cell proliferation in vitro and tumor growth in vivo by bee venom. Toxicon 2003, 41, 861–870. [Google Scholar] [CrossRef]
- Li, B.; Gu, W.; Zhang, C.; Huang, X.; Han, K.; Ling, C. Growth arrest and apoptosis of the human hepatocellular carcinoma cell line BEL-7402 induced by melittin. Onkologie 2006, 29, 367–371. [Google Scholar] [CrossRef] [PubMed]
- DeWys, W.D.; Kwaan, H.C.; Bathina, S. Effect of defibrination on tumor growth and response to chemotherapy. Cancer Res. 1976, 36, 3584–3587. [Google Scholar] [PubMed]
- Borkow, G.; Chaim-Matyas, A.; Ovadia, M. Binding of cytotoxin P4 from Naja nigricollis nigricollis to B16F10 melanoma and WEHI-3B leukemia cells. FEMS Microbiol. Lett. 1992, 105, 139–145. [Google Scholar] [CrossRef]
- Song, J.K.; Jo, M.R.; Park, M.H.; Song, H.S.; An, B.J.; Song, M.J.; Han, S.B.; Hong, J.T. Cell growth inhibition and induction of apoptosis by snake venom toxin in ovarian cancer cell via inactivation of nuclear factor κB and signal transducer and activator of transcription 3. Arch. Pharm. Res. 2012, 35, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Braganca, B.M. Biologically active components of cobra venom in relation to cancer research. Indian J. Med. Res. 1976, 64, 1197–1207. [Google Scholar] [PubMed]
- Braganca, B.M.; Hospattankar, A.V. Potentiating action of cobra venom cytotoxin on the antitumour effects of an alkylating agent (melphalan). Eur. J. Cancer 1978, 14, 707–712. [Google Scholar] [CrossRef]
- Yang, S.H.; Chien, C.M.; Lu, M.C.; Lin, Y.H.; Hu, X.W.; Lin, S.R. Up-regulation of Bax and endonuclease G, and down-modulation of Bcl-XL involved in cardiotoxin III-induced apoptosis in K562 cells. Exp. Mol. Med. 2006, 38, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Chien, C.M.; Chang, L.S.; Lin, S.R. Cardiotoxin III-induced apoptosis is mediated by Ca2+-dependent caspase-12 activation in K562 cells. J. Biochem. Mol. Toxicol. 2008, 22, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Chien, C.M.; Yang, S.H.; Chang, L.S.; Lin, S.R. Involvement of both endoplasmic reticulum- and mitochondria-dependent pathways in cardiotoxin III-induced apoptosis in HL-60 cells. Clin. Exp. Pharmacol. Physiol. 2008, 35, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Chien, C.M.; Yang, S.H.; Yang, C.C.; Chang, L.S.; Lin, S.R. Cardiotoxin III induces c-jun N-terminal kinase-dependent apoptosis in HL-60 human leukaemia cells. Cell Biochem. Funct. 2008, 26, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.L.; Su, J.C.; Chien, C.M.; Chuang, P.W.; Chang, L.S.; Lin, S.R. Down-regulation of the JAK2/PI3K-mediated signaling activation is involved in Taiwan cobra cardiotoxin III-induced apoptosis of human breast MDA-MB-231 cancer cells. Toxicon 2010, 55, 1263–1273. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.C.; Lin, K.L.; Chien, C.M.; Chang, L.S.; Lin, S.R. Effects of cardiotoxin III on NF-κB function, proliferation, and apoptosis in human breast MCF-7 cancer cells. Oncol. Res. 2009, 17, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Trikha, M.; De Clerck, Y.A.; Markland, F.S. Contortrostatin, a snake venom disintegrin, inhibits β1 integrin-mediated human metastatic melanoma cell adhesion and blocks experimental metastasis. Cancer Res. 1994, 54, 4993–4998. [Google Scholar] [PubMed]
- Jakubowski, P.; Calvete, J.J.; Eblec, J.A.; Lazarovici, P.; Marcinkiewicz, C. Identification of inhibitors of α2β1 integrin, members of C-lectin type proteins, in Echis sochureki venom. Toxicol. Appl. Pharm. 2013, 269, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Lucena, S.E.; Romo, K.; Suntravat, M.; Sánchez, E.E. Anti-angiogenic activities of two recombinant disintegrins derived from the Mohave and Prairie rattlesnakes. Toxicon 2014, 78, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.Y.; Lee, H.; You, W.K.; Chung, K.H.; Kim, D.S.; Song, K. The snake venom disintegrin salmosin induces apoptosis by disassembly of focal adhesions in bovine capillary endothelial cells. Biochem. Biophys. Res. Commun. 2003, 302, 502–508. [Google Scholar] [CrossRef]
- Swenson, S.; Costa, F.; Minea, R.; Sherwin, R.P.; Ernst, W.; Fujii, G.; Yang, D.; Markland, F.S., Jr. Intravenous liposomal delivery of the snake venom disintegrin contortrostatin limits breast cancer progression. Mol. Cancer Ther. 2004, 3, 499–511. [Google Scholar] [PubMed]
- Lin, K.L.; Chien, C.M.; Hsieh, C.Y.; Tsai, P.C.; Chang, L.S.; Lin, S.R. Antimetastatic potential of cardiotoxin III involves inactivation of PI3K/Akt and p38 MAPK signaling pathways in human breast cancer MDA-MB-231 cells. Life Sci. 2012, 90, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Feofanov, A.V.; Sharonov, G.V.; Astapova, M.V.; Rodionov, D.I.; Utkin, Y.N.; Arseniev, A.S. Cancer cell injury by cytotoxins from cobra venom is mediated through lysosomal damage. Biochem. J. 2005, 390, 11–18. [Google Scholar] [CrossRef] [PubMed]
- De Vieira Santos, M.M.; da Silva, R.J.; da Silva, M.G.; Fecchio, D. Subpopulations of mononuclear leukocytes associate with inhibition of Ehrlich ascites tumor growth by treatment with Bothrops jararaca venom. Mediat. Inflamm. 2004, 13, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Markland, F.S.; Swenson, S. Snake venom metalloproteinases. Toxicon 2013, 62, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Maria, D.A.; da Silva, M.G.L.; Correia, M.C.; Ruiz, I.R.G. Antiproliferative effect of the jararhagin toxin on B16F10 murine melanoma. BMC Complement. Altern. Med. 2014, 14, 446. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.G.; Jin, Y.; Lee, W.H.; Zhang, Y. A snake venom metalloproteinase that inhibited cell proliferation and induced morphological changes of ECV304 cells. Toxicon 2006, 47, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Bittencourt, M.; Carvalho, D.D.; Gagliardi, A.R.; Collins, D.C. The effect of a lectin from the venom of the snake, Bothrops jararacussu, on tumor cell proliferation. Anticancer Res. B 1999, 19, 4023–4025. [Google Scholar]
- Sarray, S.; Srairi, N.; Luis, J.; Marvaldi, J.; El Ayeb, M.; Marrakchi, N. Lebecetin, a C-lectin protein from the venom of Macrovipera lebetina that inhibits platelet aggregation and adhesion of cancerous cells. Haemostasis 2001, 31, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Dubashi, B.; Reddy, K.S.; Jain, P. Weekly paclitaxel in ovarian cancer-the latest success story. Curr. Oncol. 2011, 18, 16–17. [Google Scholar] [CrossRef] [PubMed]
- Al-Bahlani, S.; Fraser, M.; Wong, A.Y.; Sayan, B.S.R.; Melino, G.; Tsang, B.K. P73 regulates cisplatin-induced apoptosis in ovarian cancer cells via a calcium/calpain-dependent mechanism. Oncogene 2011, 30, 4219–4230. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.M.; Cui, M.H.; Su, M.M. Inhibitory effect of melittin on ovarian cancer. Chin. J. Lab. Diagn. 2007, 8, 013. [Google Scholar]
- Alizadehnohi, M.; Nabiuni, M.; Nazari, Z.; Safaeinejad, Z.; Irian, S. The synergistic cytotoxic effect of cisplatin and honey bee venom on human ovarian cancer cell line A2780cp. J. Venom Res. 2012, 3, 22. [Google Scholar] [PubMed]
- Amini, E.; Baharara, J.; Nikdel, N.; Abdollahi, F.S. Cytotoxic and Pro-Apoptotic Effects of Honey Bee Venom and Chrysin on Human Ovarian Cancer Cells. Asia Pac. J. Med. Toxicol. 2015, 4, 68–73. [Google Scholar] [CrossRef]
- Markland, F.S.; Shieh, K.; Zhou, Q.; Golubkov, V.; Sherwin, R.P.; Richters, V.; Sposto, R. A novel snake venom disintegrin that inhibits human ovarian cancer dissemination and angiogenesis in an orthotopic nude mouse model. Haemostasis 2001, 31, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Swenson, S.; Costa, F.; Ernst, W.; Fujii, G.; Markland, F.S. Contortrostatin, a snake venom disintegrin with anti-angiogenic and anti-tumor activity. Pathophysiol. Haemost. Thromb. 2005, 34, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.J.; Jeon, O.H.; Kim, D.S. Saxatilin, a snake venom disintegrin, regulates platelet activation associated with human vascular endothelial cell migration and invasion. J. Vasc. Res. 2007, 44, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Jang, Y.J.; Jeon, O.H.; Kim, D.S. Saxatilin, a snake venom disintegrin, suppresses TNF-α-induced ovarian cancer cell invasion. J. Biochem. Mol. Biol. 2007, 40, 290–294. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, D.D.; Schmitmeier, S.; Novello, J.C.; Markland, F.S. Effect of BJcuL (a lectin from the venom of the snake Bothrops jararacussu) on adhesion and growth of tumor and endothelial cells. Toxicon 2001, 39, 1471–1476. [Google Scholar] [CrossRef]
- Alonezi, S.; Tusiimire, J.; Wallace, J.; Dufton, M.J.; Parkinson, J.A.; Young, J.C.; Clements, C.J.; Park, J.K.; Jeon, J.W.; Ferro, V.A.; et al. Metabolomic profiling of the synergistic effects of melittin in combination with cisplatin on ovarian cancer cells. Metabolites 2017, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Lee, S.J.; Park, E.K.; Lee, Y.S.; Yoon, J.H.; Hur, S.Y.; Song, M.J. Anti-Tumor Activity of Melittin in Ovarian Cancer Cell Lines. 2012. Available online: http://www.papersearch.net/thesis/article.asp?key=3199828 (accessed on 22 July 2017).
- Su, M.; Chang, W.; Cui, M.; Lin, Y.; Wu, S.; Xu, T. Expression and anticancer activity analysis of recombinant human uPA1-43-melittin. Int. J. Oncol. 2015, 46, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Chang, W.; Cui, M.; Lin, Y.; Wu, S.; Xu, T. Expression and purification of recombinant ATF-mellitin, a new type fusion protein targeting ovarian cancer cells, in P. pastoris. Oncol. Rep. 2016, 35, 1179–1185. [Google Scholar] [CrossRef] [PubMed]
- Lipps, B.V. Novel snake venom proteins cytolytic to cancer cells in vitro and in vivo systems. J. Venom. Anim. Toxins 1999, 5, 172–183. [Google Scholar] [CrossRef]


{kind=link}
| Compound | Biological Effects | |
|---|---|---|
| Peptides | Melittin | anti-inflammatory, immunostimulatory, immunosuppressive, antibacterial, antifungal, antiviral, cytotoxic effect, ↑ the activity of phospholipase A2, anti-atherosclerotic, endosomolytic, stimulates smooth muscles, activates the hypophysis and adrenal glands, ↑ capillary permeability by ↑ blood circulation and ↓ blood pressure, ↓ blood coagulation |
| Apamin | cytotoxic effect, anti-inflammatory, anti-serotonin action, immunosuppressive, activates the hypophysis and adrenal glands, nociceptive effect | |
| MCD peptide | lyses mast cells, releases histamine, serotonin and heparin, ↑ capillary permeability, anti-inflammatory, analgesic effect, simulates the central nervous system | |
| Adolapine | inhibits PLA2 activity, inhibits COX activity, ↓ inflammation and ↓ pain, antipyretic, inhibits the aggregation of erythrocytes | |
| Protease inhibitor | inhibits the activity of trypsin, chymotrypsin, plasmin, thrombin, ↓ inflammation, anti-rheumatic | |
| Procamine A, B, Secapine, Panime, Minimine, Tertiapine, Cardiopep, Melittin F | ||
| Proteins (Enzymes) | Phospholipase A2 | immunomodulatory, anti-inflammatory, destroys phospholipids and dissolves the cell membrane of blood bodies; ↓ the blood coagulation and blood pressure, prevents neuronal cell death caused by prion peptides, cytotoxic effects against cancer cells, antitumor effects |
| Phospholipase B | detoxicating activity | |
| Hyaluronidase | catalyzes the hydrolysis of proteins, dilates blood vessels and ↑ permeability, causing an ↑ in blood circulation; immune response, tissue spread activity | |
| Glucosidase, Acid phosphomonoesterase | ||
| Biogenic Amines | Histamines | dilates blood vessels, ↑ the permeability of blood capillaries and ↑ blood circulation; stimulates smooth muscles |
| Dopamine, Norepinephrine | ||
| Others | Carbohydrates, r-Aminobutyric acid, B-aminoisobutyric acid | |
| Proteins | Enzymes | Compound | Major Activity |
| Acetylcholinesterase | Paralysis | ||
| Bglucosaminidase | Tissue damage | ||
| Phosphoesterase | Anticoagulant, paralysis | ||
| Phospholipase A2 | Membrane damage, anticoagulant | ||
| Hyaluronidase | ECM damage, apoptosis | ||
| l-amino acid oxidase | Platelet effect, anticoagulant | ||
| Snake venom metalloproteases | Anticoagulant, cell damage | ||
| Snake venom serine proteases | Anticoagulant, fibrinogenemia | ||
| Non-Enzymes | Protein C activator | Anticoagulant | |
| Growth factors (INGF, VEGF) | Endothelial damage, edema | ||
| Inhibitor of the prothrombinase complex formation | Anticoagulant | ||
| Lectins | Platelet effect | ||
| Precursors of bioactive peptides | Smooth muscle inhibitor | ||
| Von Willebrand factor binding proteins | Anticoagulant | ||
| Cysteine-rich secretory proteins | Anticoagulant | ||
| Peptides | Cytotoxic, cardiotoxic, myotoxic, neurotoxic | Anticoagulant, inflammatory | |
| Disintegrins | Apoptosis, myotoxicity | ||
| Natriuretic | Platelet effect, vascular | ||
| Bradykinin potentiator | Hypotensive | ||
| Organic Compounds | Biogenic amines | Serotonin, histamine | |
| Amino acids, carbohydrates, citrate, nucleosides | |||
| Inorganic Compounds | Calcium, cobalt, copper, iron, phosphorus, potassium, magnesium, sodium, zinc | ||
| Study | Compound | Mechanism | Cancer Cell | Results |
|---|---|---|---|---|
| Alizadehnohi et al. [111] | Melittin | induces apoptosis in cisplatin-resistant ovarian cancer cells |
|
|
| Alonezi et al. [118] | Melittin | compared the effects of melittin in combination with cisplatin |
|
|
| Amini et al. [112] | Bee venom and chrysin | cytotoxic and pro-apoptotic effects of BV and chrysin |
|
|
| Holle et al. [78] | Melittin/avidin conjugate | cytotoxic effects |
|
|
| Jo et al. [26] | Melittin | inhibits cell growth through enhancement of DR expressions |
|
|
| Lee et al. [119] | Melittin | suppresses the proliferation and growth of tumor cells |
|
|
| Liu et al. [29] | Melittin-MhIL-2 fusion protein | inhibits cell growth and proliferation of ovarian carcinoma |
|
|
| Su et al. [120] | Recombinant human Upa1-43-melittin | inhibits growth of ovarian cancer cells |
|
|
| Su et al. [121] | ATF-melittin | cytolytic activity |
|
|
| Xu et al. [110] | Melittin | inhibits the growth and activity of proliferation of ovarian cancer |
|
|
| Study | Compound | Species | Mechanism | Cancer Cell | Results |
|---|---|---|---|---|---|
| Markland et al. [113] | Contortrostatin | Agkistrodon contortrix |
|
|
|
| Swenson et al. [114] | Contortrostatin | Agkistrodon contortrix |
|
|
|
| Lipps et al. [122] | Atroporin and Kaotree | Crotalus atrox Naja naja kaouthia |
|
|
|
| Kim et al. [116] | Saxatilin | NR |
|
|
|
| Carvalho et al. [117] | BJcuL | Bothrops jararacussu |
|
|
|
| Song et al. [86] | NR | Vipera lebetina turanica |
|
|
|
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moga, M.A.; Dimienescu, O.G.; Arvătescu, C.A.; Ifteni, P.; Pleş, L. Anticancer Activity of Toxins from Bee and Snake Venom—An Overview on Ovarian Cancer. Molecules 2018, 23, 692. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23030692
Moga MA, Dimienescu OG, Arvătescu CA, Ifteni P, Pleş L. Anticancer Activity of Toxins from Bee and Snake Venom—An Overview on Ovarian Cancer. Molecules. 2018; 23(3):692. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23030692
Chicago/Turabian StyleMoga, Marius Alexandru, Oana Gabriela Dimienescu, Cristian Andrei Arvătescu, Petru Ifteni, and Liana Pleş. 2018. "Anticancer Activity of Toxins from Bee and Snake Venom—An Overview on Ovarian Cancer" Molecules 23, no. 3: 692. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23030692