Extracellular Phytase Production by the Wine Yeast S. cerevisiae (Finarome Strain) during Submerged Fermentation


Abstract
:
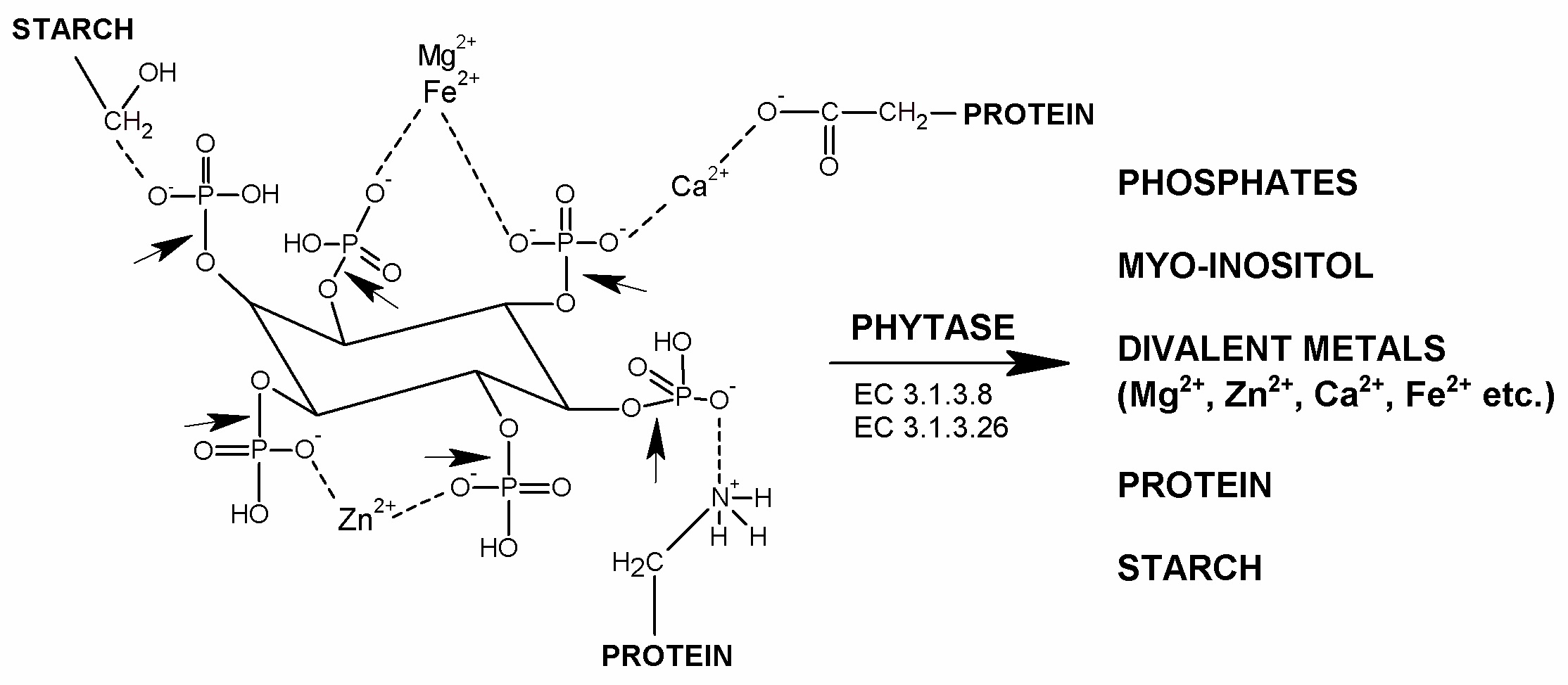
1. Introduction
2. Results and Discussion
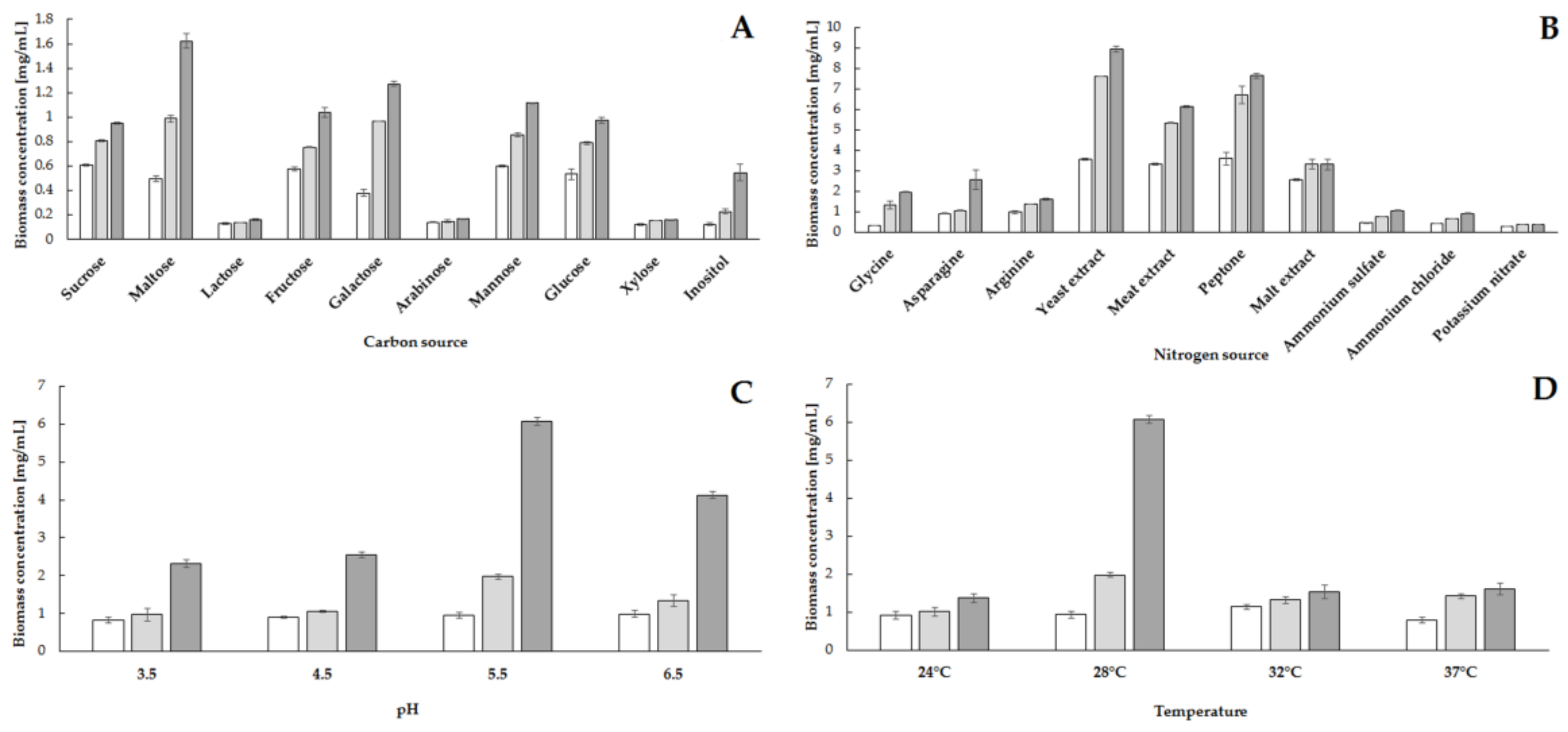
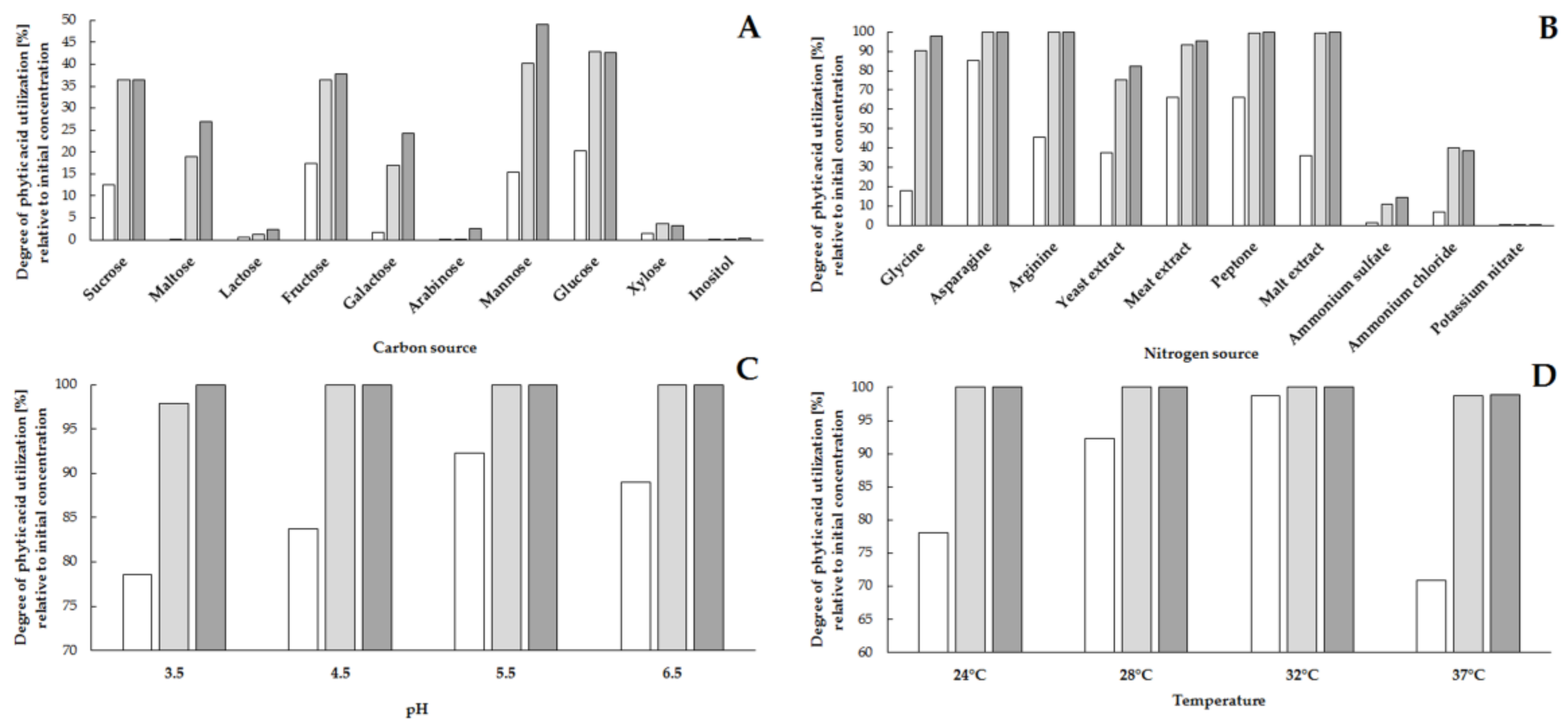
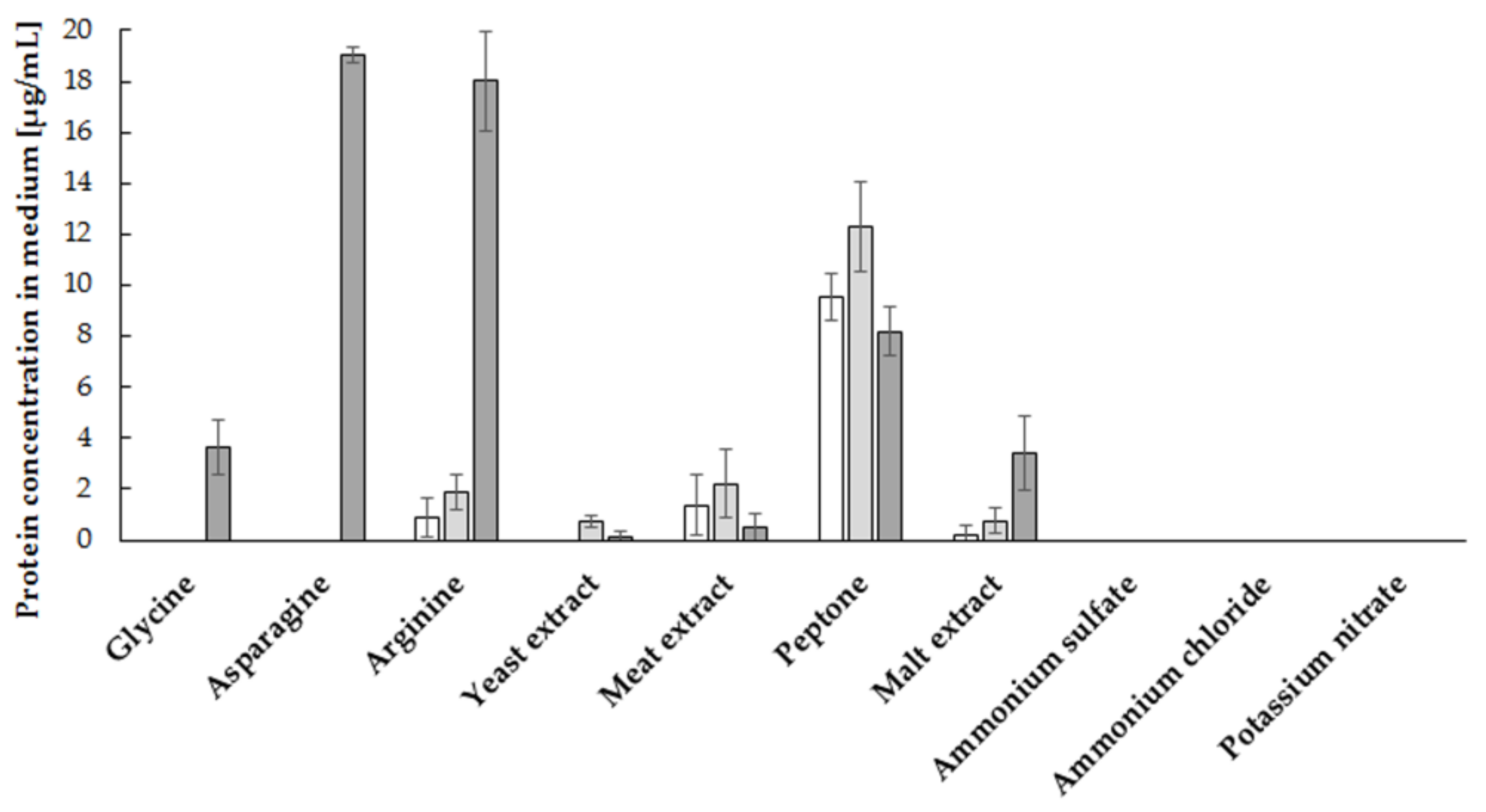
2.1. Influence of Medium Components and Culture Conditions on the Degree of Utilization of Phytic Acid
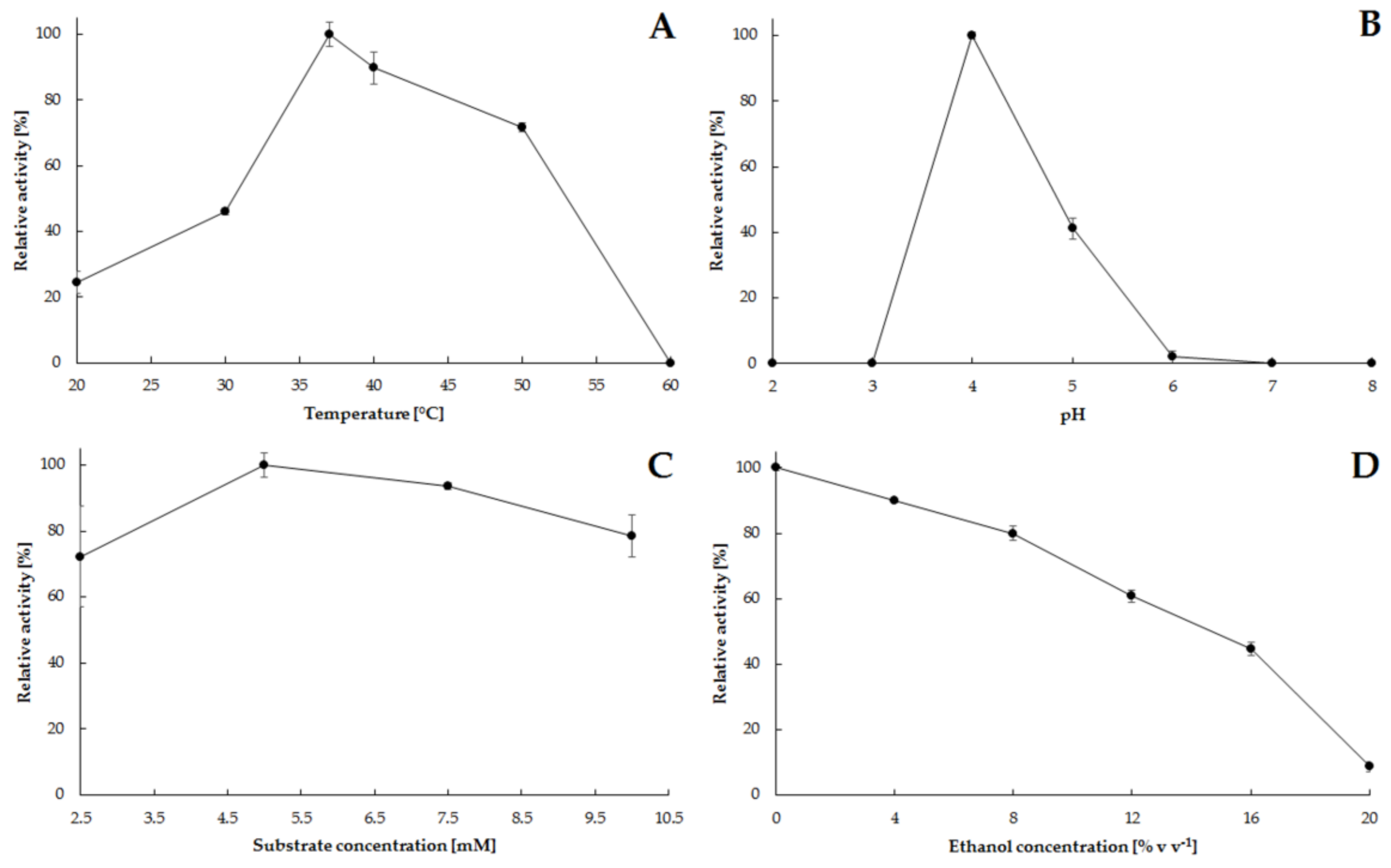
2.2. Catalytic Characteristics of Phytase Isolated from S. cerevisiae Strain Finarome
3. Materials and Methods
3.1. Yeast Strain and Inoculum Preparation
3.2. Culture Conditions Used for the Selection of the Composition of the Culture Media
3.3. Culture Conditions Used to Determine Catalytic Activity of Phytase
3.4. Analytical Methods
3.5. Determination of Catalytic Activity of Phytase
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Conflicts of Interest
Compliance with Ethics Requirements
References
- Boyce, A.; Walsh, G. Purification and characterisation of an acid phosphatase with phytase activity from Mucor hiemalis Wehmer. J. Biotechnol. 2007, 132, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Escobin-Mopera, L.; Ohtani, M.; Sekiguchi, S.; Sone, T.; Abe, A.; Tanaka, M.; Meevootisom, V.; Asano, K. Purification and characterization of phytase from Klebsiella pneumoniae 9-3B. J. Biosci. Bioeng. 2012, 113, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Haefner, S.; Knietsch, A.; Scholten, E.; Braun, J.; Lohscheidt, M.; Zelder, O. Biotechnological production and applications of phytases. Appl. Microbiol. Biotechnol. 2005, 68, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Vats, P.; Banerjee, U. Production studies and catalytic properties of phytases (myo-inositolhexakisphosphate phosphohydrolases): An overview. Enzyme Microb. Tech. 2004, 15, 3–14. [Google Scholar] [CrossRef]
- Mikulski, D.; Kłosowski, G.; Rolbiecka, A. Influence of phytase and supportive enzymes applied during high gravity mash preparation on the improvement of technological indicators of the alcoholic fermentation process. Biomass Bioenergy 2015, 80, 191–202. [Google Scholar] [CrossRef]
- Mikulski, D.; Kłosowski, G. Phytic acid concentration in selected raw materials and analysis of its hydrolysis rate with the use of microbial phytases during the mashing process. J. Inst. Brew. 2015, 121, 213–218. [Google Scholar] [CrossRef]
- Bogar, B.; Szakacs, G.; Linden, J.C.; Pandey, A.; Tengerdy, R.P. Optimization of phytase production by solid substrate fermentation. J. Ind. Microbiol. Biotechnol. 2003, 30, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Vohra, A.; Satyanarayana, T. Statistical optimization of the medium components by response surface methodology to enhance phytase production by Pichia anomala. Process Biochem. 2002, 37, 999–1004. [Google Scholar] [CrossRef]
- Kaur, P.; Satyanarayana, T. Production of cell-bound phytase by Pichia anomala in an economical cane molasses medium: Optimization using statistical tools. Process Biochem. 2005, 40, 3095–3102. [Google Scholar] [CrossRef]
- Vohra, A.; Satyanarayana, T. Phytase production by the yeast, Pichia anomala. Biotechnol. Lett. 2001, 23, 551–554. [Google Scholar] [CrossRef]
- Kammoun, R.; Farhat, A.; Chouayekh, H.; Bouchaala, K.; Bejar, S. Phytase production by Bacillus subtilis US417 in submerged and solid state fermentation. Ann. Microbiol. 2012, 62, 155–164. [Google Scholar] [CrossRef]
- Singh, B.; Satyanarayana, T. Improved phytase production by a thermophilic mould Sporotrichum thermophile in submerged fermentation due to statistical optimization. Bioresour. Technol. 2008, 99, 824–830. [Google Scholar] [CrossRef] [PubMed]
- Ries, E.F.; Macedo, G.A. Improvement of phytase activity by a new Saccharomyces cerevisiae strain using statistical optimization. Enzyme Res. 2011, 11. [Google Scholar] [CrossRef]
- In, M.-J.; Soe, S.-W.; Oh, N.-S. Fermentative production and application of acid phytase by Saccharomyces cerevisiae CY strain. Afr. J. Biotechnol. 2008, 7, 3115–3120. [Google Scholar]
- Pandey, A.; Szakacs, G.; Soccol, C.R.; Rodriguez-Leon, J.A.; Soccol, V.T. Production, purification and properties of microbial phytase. Bioresour. Technol. 2001, 77, 203–214. [Google Scholar] [CrossRef]
- Mikulski, D.; Kłosowski, G. Evaluation of phytic acid utilization by S. cerevisiae strains used in fermentation processes and biomass production. J. Basic Microbiol. 2017, 57, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Walker, G.M. Yeast: Physiology and Biotechnology; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2000; pp. 52–53. [Google Scholar]
- Qasim, S.S.; Shakir, K.A.; Al-Shaibani, A.B.; Walsh, M.K. Optimization of culture conditions to produce phytase from Aspergillus tubingensis SKA. Food Nutr. Sci. 2017, 8, 733–745. [Google Scholar] [CrossRef]
- Vohra, A.; Satyanarayana, T. A cost-effective cane molasses medium for enhanced cell-bound phytase production by Pichia anomala. J. Appl. Microbiol. 2004, 97, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Roehr, M. Biotechnology of Ethanol; WILEY-VCH Verlag GmbH: Weinheim, Germany, 2001; p. 52. [Google Scholar]
- Guo, M.; Hang, H.; Zhu, T.; Zhuang, Y.; Chu, J.; Zhang, S. Effect of glycosylation on biochemical characterization of recombinant phytase expressed in Pichia pastoris. Enzyme Microb. Tech. 2008, 42, 340–345. [Google Scholar] [CrossRef]
- Bai, F.W.; Zhao, X.Q. High Gravity Ethanol Fermentations and Yeast Tolerance. In Microbial Stress Tolerance for Biofuels; Liu, Z.L., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 117–137. [Google Scholar]
- Haros, M.; Bielecka, M.; Sanz, Y. Phytase activity as a novel metabolic feature in Bifidobacterium. FEMS Microbiol. Lett. 2005, 247, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Wyss, M.; Brugger, R.; Kronenberger, A.; Rémy, R.; Fimbel, R.; Oesterhelt, G.; Lehmann, M.; van Loon, A.P.G.M. Biochemical characterization of fungal phytase (myo-inositol hexakisphosphate phosphohydrolases): Catalytic properties. Appl. Environ. Microb. 1999, 65, 367–373. [Google Scholar]
- In, M.-J.; Seo, S.-W.; Kim, D.C.; Oh, N.-S. Purification and biochemical properties of an extracellular acid phytase produced by the Saccharomyces cerevisiae CY strain. Process Biochem. 2009, 44, 122–126. [Google Scholar] [CrossRef]
- Li, X.; Chi, Z.; Liu, Z.; Li, J.; Wang, X.; Hirimuthugoda, N.Y. Purification and characterization of extracellular phytase from a marine yeast Kodamaea ohmeri BG3. Mar. Biotechnol. 2008, 10, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Casey, A.; Walsh, G. Purification and characterization of extracellular phytase from Aspergillus niger ATCC 9142. Bioresour. Technol. 2003, 86, 183–188. [Google Scholar] [CrossRef]
- Olstorpe, M.; Schnürer, J.; Passoth, V. Screening of yeast strains for phytase activity. FEMS Yeast Res. 2009, 9, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Sonderegger, M.; Sauer, U. Evolutionary engineering of Saccharomyces cerevisiae for anaerobic growth on xylose. Appl. Environ. Microb. 2003, 69, 1990–1998. [Google Scholar] [CrossRef]
- Park, H.-R.; Ahn, H.-J.; Kim, S.-H.; Lee, C.-H.; Byun, M.-W.; Lee, G.-W. Determination of the phytic acid levels in infant foods using different analytical methods. Food Control 2006, 17, 727–732. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid sensitive method for the quantitation of quantities of protein utilizing the princuple of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Shimizu, M. Purification and characterization of phytase from Bacillus subtilis (natto) N-77. Biosci. Biotechnol. Biochem. 1992, 58, 1266–1269. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagents | Relative Activity (%) | |
|---|---|---|
| 5 mM | 10 mM | |
| Control | 100.0 ± 1.3 | 100.0 ± 0.5 |
| ZnSO4 | 113.0 ± 0.6 | 84.7 ± 0.3 |
| KCl | 104.7 ± 5.6 | 102.6 ± 2.3 |
| CaCl2 | 112.0 ± 1.4 | 111.8 ± 0.3 |
| CuCl2 | 91.6 ± 1.1 | 61.4 ± 0.4 |
| MgSO4 | 125.0 ± 1.3 | 106.1 ± 0.7 |
| MnSO4 | 120.8 ± 2.0 | 106.7 ± 1.3 |
| FeSO4 | 112.6 ± 1.3 | 93.8 ± 0.0 |
| AgSO4 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| NiCl2 | 125.1 ± 0.0 | 105.3 ± 0.5 |
| CoCl2 | 108.5 ± 0.4 | 106.7 ± 1.6 |
| HgSO4 | 11.7 ± 7.3 | 0.0 ± 0.0 |
| CdSO4 | 109.2 ± 1.4 | 88.0 ± 0.6 |
| Reagents | Relative Activity (%) |
|---|---|
| 5 mM | |
| Control | 100.0a ± 1.1 |
| CaCl2 + MnSO4 | 97.1a ± 1.1 |
| CaCl2 + NiCl2 | 100.6a ± 1.5 |
| CaCl2 + MgSO4 | 98.3a ± 1.1 |
| CaCl2 + CoCl2 | 100.3a ± 0.2 |
| MnSO4 + NiCl2 | 99.2a ± 1.0 |
| MnSO4 + MgSO4 | 97.3a ± 1.3 |
| MnSO4 + CoCl2 | 98.3a ± 1.1 |
| MgSO4 + NiCl2 | 98.9a ± 0.8 |
| MgSO4 + CoCl2 | 97.4a ± 2.5 |
| NiCl2 + CoCl2 | 100.3a ± 0.6 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kłosowski, G.; Mikulski, D.; Jankowiak, O. Extracellular Phytase Production by the Wine Yeast S. cerevisiae (Finarome Strain) during Submerged Fermentation. Molecules 2018, 23, 848. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23040848
Kłosowski G, Mikulski D, Jankowiak O. Extracellular Phytase Production by the Wine Yeast S. cerevisiae (Finarome Strain) during Submerged Fermentation. Molecules. 2018; 23(4):848. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23040848
Chicago/Turabian StyleKłosowski, Grzegorz, Dawid Mikulski, and Oliwia Jankowiak. 2018. "Extracellular Phytase Production by the Wine Yeast S. cerevisiae (Finarome Strain) during Submerged Fermentation" Molecules 23, no. 4: 848. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23040848