Female Reproductive Performance in the Mouse: Effect of Oral Melatonin

 , ,
, , 
Abstract
:1. Introduction
2. Results
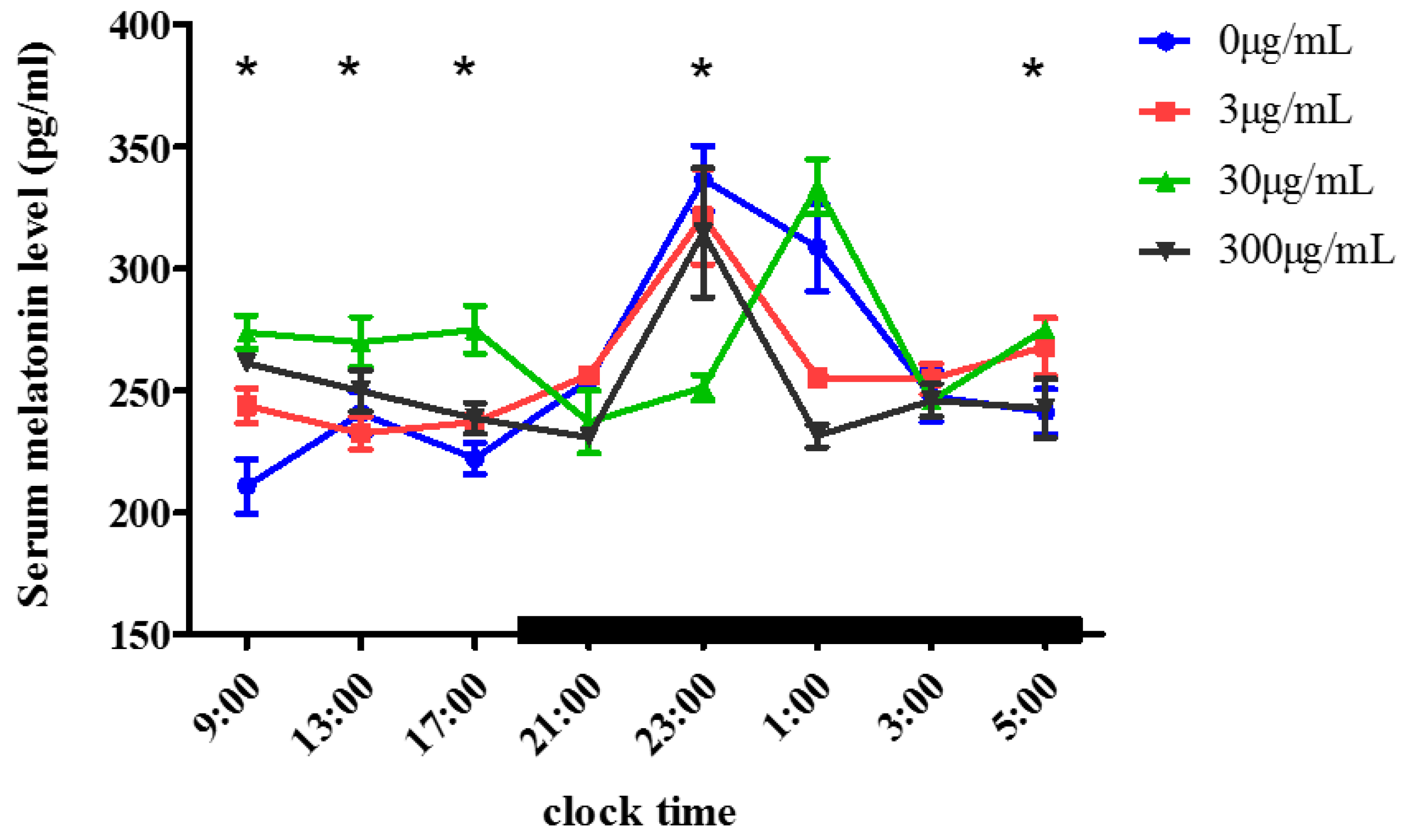
2.1. Effect of Melatonin on the Serum Melatonin Levels
2.2. Effect of Melatonin on Litter Size and Birth Weight of Pups
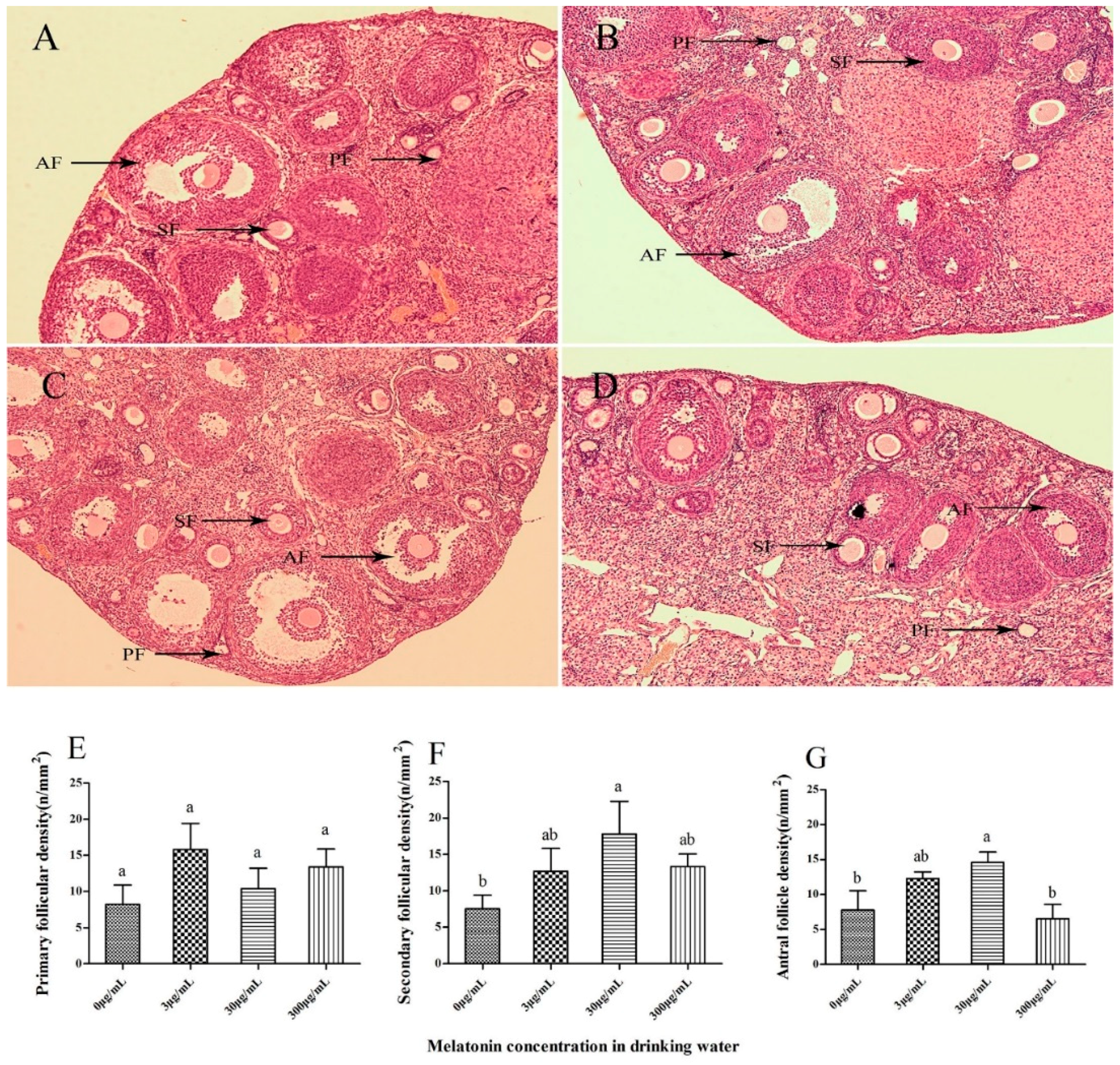
2.3. Effect of Melatonin on Follicular Development in Mouse Ovaries
2.4. Effect of Melatonin on Blastocyst Formation
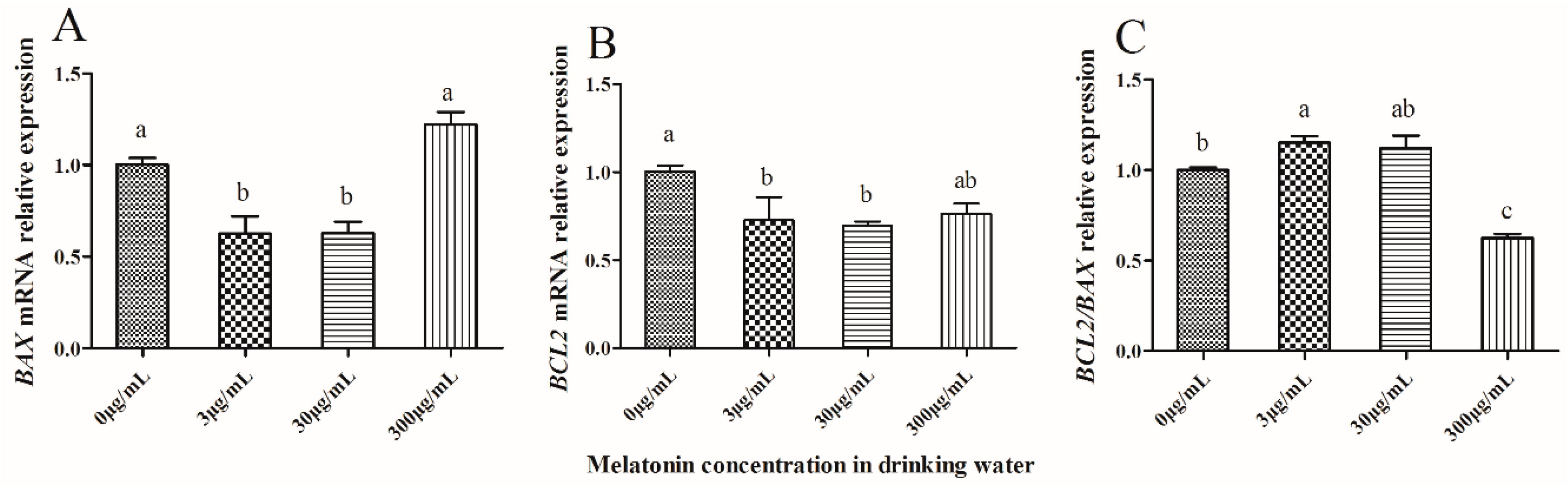
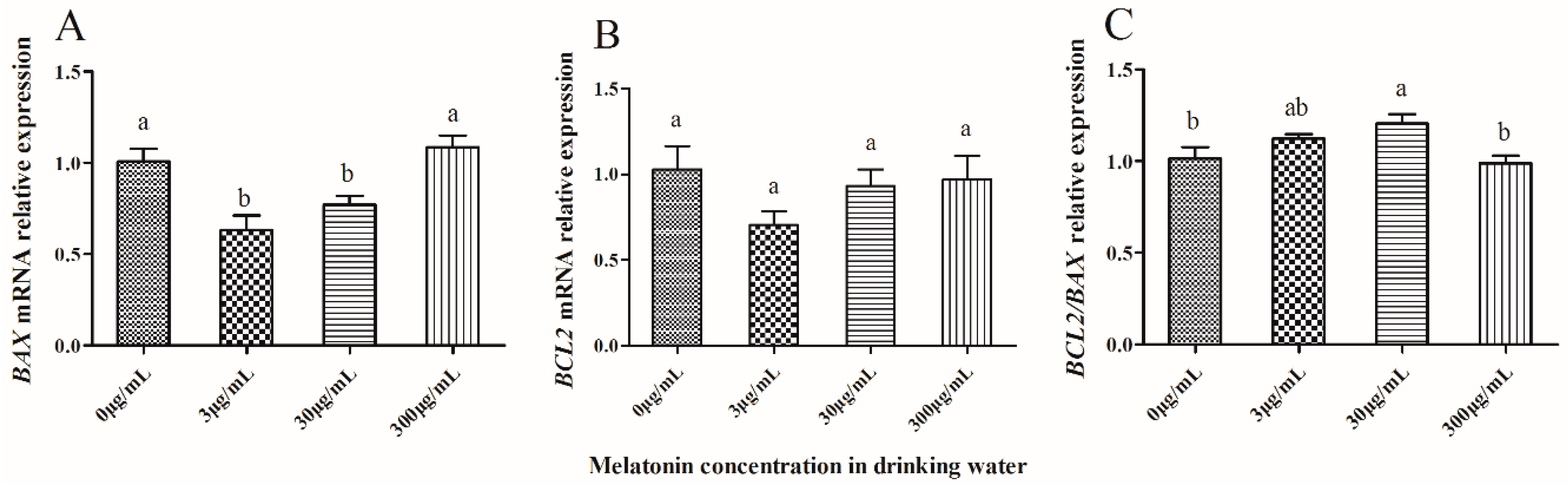
2.5. Effect of Melatonin on Expression of Apoptosis-Related Genes in IVF-Derived Mouse Embryos
3. Discussion
4. Materials and Methods
4.1. Animals and Drinking Water
4.2. Serum Melatonin Assay
4.3. HE Staining and Histological Analysis
4.4. Litter Size and Litter Birth Weights
4.5. In Vitro Fertilization (IVF) and In Vitro Embryo Culture
4.6. Quantitative Polymerase Chain Reaction (Q-PCR)
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GnRH | gonadotropin-releasing hormone |
| hCG | human chorionic gonadotropin |
| HE | hematoxylin and eosin |
| HTF | human tubal fluid |
| IVF | in vitro fertilization |
| MT | Melatonin |
| Q-PCR | Quantitative polymerase chain reaction |
| ROS | reactive oxygen species |
References
- Lerner, A.B.; Case, J.D.; Mori, W.; Wright, M.R. Melatonin in peripheral nerve. Nature 1959, 183, 1821. [Google Scholar] [CrossRef] [PubMed]
- Lincoln, G.A.; Clarke, I.J. Photoperiodically-induced cycles in the secretion of prolactin in hypothalamo-pituitary disconnected rams: Evidence for translation of the melatonin signal in the pituitary gland. J. Neuroendocrinol. 1994, 6, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Fiske, V.M.; Parker, K.L.; Ulmer, R.A.; Ow, C.H.; Aziz, N. Effect of melatonin alone or in combination with human chorionic gonadotropin or ovine luteinizing hormone on the in vitro secretion of estrogens or progesterone by granulosa cells of rats. Endocrinology 1984, 114, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Woo, M.M.; Tai, C.J.; Kang, S.K.; Nathwani, P.S.; Pang, S.F.; Leung, P.C. Direct action of melatonin in human granulosa-luteal cells. J. Clin. Endocrinol. Metab. 2001, 86, 4789–4797. [Google Scholar] [CrossRef] [PubMed]
- Sirotkin, A.V.; Schaeffer, H.J. Direct regulation of mammalian reproductive organs by serotonin and melatonin. J. Endocrinol. 1997, 154, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J. Melatonin and human reproduction. Ann. Med. 1998, 30, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.P.; Gogenur, I.; Rosenberg, J.; Reiter, R.J. The Safety of Melatonin in Humans. Clin. Drug Investig. 2016, 36, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J. Melatonin: Clinical relevance. Best Pract. Res. Clin. Endocrinol. Metab. 2003, 17, 273–285. [Google Scholar] [CrossRef]
- Tamura, H.; Kawamoto, M.; Sato, S.; Tamura, I.; Maekawa, R.; Taketani, T.; Aasada, H.; Takaki, E.; Nakai, A.; Reiter, R.J.; et al. Long-term melatonin treatment delays ovarian aging. J. Pineal Res. 2017, 62, e12381. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Peng, W.; Yin, S.; Zhao, J.; Fu, B.; Zhang, J.; Mao, T.; Wu, H.; Zhang, Y. Melatonin improves age-induced fertility decline and attenuates ovarian mitochondrial oxidative stress in mice. Sci. Rep. 2016, 6, 35165. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, T.; Hashimoto, S.; Ito, K.; Nakaoka, Y.; Matsumoto, K.; Hosoi, Y.; Morimoto, Y. Oral melatonin supplementation improves oocyte and embryo quality in women undergoing in vitro fertilization-embryo transfer. Gynecol. Endocrinol. 2014, 30, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, H.; Sun, J.; Tian, Y.; Li, C. Effect of melatonin on the peripheral T lymphocyte cell cycle and levels of reactive oxygen species in patients with premature ovarian failure. Exp. Ther. Med. 2016, 12, 3589–3594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Tian, X.; Zhang, L.; Tan, D.; Reiter, R.J.; Liu, G. Melatonin promotes the in vitro development of pronuclear embryos and increases the efficiency of blastocyst implantation in murine. J. Pineal Res. 2013, 55, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Berlinguer, F.; Leoni, G.G.; Succu, S.; Spezzigu, A.; Madeddu, M.; Satta, V.; Bebbere, D.; Contreras-Solis, I.; Gonzalez-Bulnes, A.; Naitana, S. Exogenous melatonin positively influences follicular dynamics, oocyte developmental competence and blastocyst output in a goat model. J. Pineal Res. 2009, 46, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Cebrian-Serrano, A.; Salvador, I.; Raga, E.; Dinnyes, A.; Silvestre, M.A. Beneficial effect of melatonin on blastocyst in vitro production from heat-stressed bovine oocytes. Reprod. Domest. Anim. 2013, 48, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.M.; Tian, X.Z.; Zhou, G.B.; Wang, L.; Gao, C.; Zhu, S.E.; Zeng, S.M.; Tian, J.H.; Liu, G.S. Melatonin exists in porcine follicular fluid and improves in vitro maturation and parthenogenetic development of porcine oocytes. J. Pineal Res. 2009, 47, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Park, S.M.; Lee, E.; Kim, J.H.; Jeong, Y.I.; Lee, J.Y.; Park, S.W.; Kim, H.S.; Hossein, M.S.; Jeong, Y.W.; et al. Anti-apoptotic effect of melatonin on preimplantation development of porcine parthenogenetic embryos. Mol. Reprod. Dev. 2008, 75, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Ma, T.; Shi, J.; Zhang, Z.; Wang, J.; Zhu, K.; Li, Y.; Yang, M.; Song, Y.; Liu, G. Melatonin and its receptor MT1 are involved in the downstream reaction to luteinizing hormone and participate in the regulation of luteinization in different species. J. Pineal Res. 2016, 61, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhuo, Z.Y.; Shi, W.Q.; Tan, D.X.; Gao, C.; Tian, X.Z.; Zhang, L.; Zhou, G.B.; Zhu, S.E.; Yun, P.; et al. Melatonin Promotes Superovulation in Sika Deer (Cervus nippon). Int. J. Mol. Sci. 2014, 15, 12107–12118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forcada, F.; Abecia, J.A.; Cebrian-Perez, J.A.; Muino-Blanco, T.; Valares, J.A.; Palacin, I.; Casao, A. The effect of melatonin implants during the seasonal anestrus on embryo production after superovulation in aged high-prolificacy Rasa Aragonesa ewes. Theriogenology 2006, 65, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, M.I.; Abecia, J.A.; Forcada, F.; Casao, A. Effects of exogenous melatonin on in vivo embryo viability and oocyte competence of undernourished ewes after weaning during the seasonal anestrus. Theriogenology 2010, 74, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Waldhauser, F.; Waldhauser, M.; Lieberman, H.R.; Deng, M.H.; Lynch, H.J.; Wurtman, R.J. Bioavailability of oral melatonin in humans. Neuroendocrinology 1984, 39, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Li, N.; Bo, L.; Xu, Z. Melatonin and hypothalamic-pituitary-gonadal axis. Curr. Med. Chem. 2013, 20, 2017–2031. [Google Scholar] [CrossRef] [PubMed]
- Fernando, S.; Rombauts, L. Melatonin: Shedding light on infertility?—A review of the recent literature. J. Ovarian Res. 2014, 7, 98. [Google Scholar] [CrossRef] [PubMed]
- Myers, M.; Britt, K.L.; Wreford, N.G.; Ebling, F.J.; Kerr, J.B. Methods for quantifying follicular numbers within the mouse ovary. Reproduction 2004, 127, 569–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haghjooy Javanmard, S.; Ziaei, A.; Ziaei, S.; Ziaei, E.; Mirmohammad-Sadeghi, M. The effect of preoperative melatonin on nuclear erythroid 2-related factor 2 activation in patients undergoing coronary artery bypass grafting surgery. Oxid. Med. Cell. Longev. 2013, 2013, 676829. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, B.; Kuribayashi, Y.; Murai, K.; Amemiya, A.; Itoh, M.T. The effect of melatonin on in vitro fertilization and embryo development in mice. J. Pineal Res. 2000, 28, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Goto, Y.; Noda, Y.; Mori, T.; Nakano, M. Increased generation of reactive oxygen species in embryos cultured in vitro. Free Radic Biol. Med. 1993, 15, 69–75. [Google Scholar] [CrossRef]
- Rodriguez, C.; Mayo, J.C.; Sainz, R.M.; Antolin, I.; Herrera, F.; Martin, V.; Reiter, R.J. Regulation of antioxidant enzymes: A significant role for melatonin. J. Pineal Res. 2004, 36, 1–9. [Google Scholar] [CrossRef] [PubMed]
- De Leon, P.M.; Campos, V.F.; Corcini, C.D.; Santos, E.C.; Rambo, G.; Lucia, T., Jr.; Deschamps, J.C.; Collares, T. Cryopreservation of immature equine oocytes, comparing a solid surface vitrification process with open pulled straws and the use of a synthetic ice blocker. Theriogenology 2012, 77, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Maity, P.; Bindu, S.; Dey, S.; Goyal, M.; Alam, A.; Pal, C.; Reiter, R.; Bandyopadhyay, U. Melatonin reduces indomethacin-induced gastric mucosal cell apoptosis by preventing mitochondrial oxidative stress and the activation of mitochondrial pathway of apoptosis. J. Pineal Res. 2009, 46, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Dehghani-Mohammadabadi, M.; Salehi, M.; Farifteh, F.; Nematollahi, S.; Arefian, E.; Hajjarizadeh, A.; Parivar, K.; Nourmohammadi, Z. Melatonin modulates the expression of BCL-xl and improve the development of vitrified embryos obtained by IVF in mice. J. Assist. Reprod. Genet. 2014, 31, 453–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, K. Cell death in the mammalian blastocyst. Mol. Hum. Reprod. 1997, 3, 919–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oltvai, Z.N.; Milliman, C.L.; Korsmeyer, S.J. Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that accelerates programmed cell death. Cell 1993, 74, 609–619. [Google Scholar] [CrossRef]
- Sheikhi, M.; Hultenby, K.; Niklasson, B.; Lundqvist, M.; Hovatta, O. Clinical grade vitrification of human ovarian tissue: An ultrastructural analysis of follicles and stroma in vitrified tissue. Hum. Reprod. 2011, 26, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Gougeon, A. Dynamics of follicular growth in the human: A model from preliminary results. Hum. Reprod. 1986, 1, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group (Oral MT) | No. of Oocytes Used for IVF | No. of Oocytes Developed to | ||||
|---|---|---|---|---|---|---|
| 2-Cell Embryos (%) | 4-Cell Embryos (%) | Early Blastocyst (%) | Expanded Blastocyst (%) | Hatched Blastocyst (%) | ||
| 0 μg/mL | 146 | 137(93.48 ± 5.70) | 133(90.06 ± 3.02) ab | 127(86.23 ± 4.74) a | 119(81.11 ± 4.46) a | 107(72.10 ± 8.13) bc |
| 3 μg/mL | 152 | 145(95.78 ± 2.78) | 143(93.35 ± 3.23) a | 138(91.08 ± 3.19) a | 135(87.73 ± 3.88) a | 131(83.89 ± 6.46) ab |
| 30 μg/mL | 180 | 170(95.38 ± 5.81) | 169(94.65 ± 5.38) a | 158(88.51 ± 7.82) a | 155(87.10 ± 8.89) a | 152(85.70 ± 8.10) a |
| 300 μg/mL | 110 | 94(87.5 ± 12.8) | 87(82.80 ± 15.18) b | 72(67.66 ± 14.34) b | 68(63.62 ± 12.83) b | 64(60.32 ± 17.24) c |
| Gene | Assay ID | Primer Seq (5′–3′) | Product Length | Tm (°C) |
|---|---|---|---|---|
| BCL2 | NM_009741.5 | F: AGGATTGTGGCCTTCTTTGA | 120 | 60 |
| R: CAGATGCCGGTTCAGGTACT | ||||
| BAX | NM_007527.3 | F: TGGAGATGAACTGGACAGCA | 117 | 60 |
| R: TGAAGTTGCCATCAGCAAAC | ||||
| GAPDH | NM_001289726.1 | F: AGAACATCATCCCTGCATCC | 124 | 63 |
| R: AGATCCACGACGGACACATT |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Wang, D.; Wu, Z.; Pan, B.; Yang, H.; Zeng, C.; Zhang, M.; Liu, G.; Han, H.; Zhou, G. Female Reproductive Performance in the Mouse: Effect of Oral Melatonin. Molecules 2018, 23, 1845. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23081845
Zhao X, Wang D, Wu Z, Pan B, Yang H, Zeng C, Zhang M, Liu G, Han H, Zhou G. Female Reproductive Performance in the Mouse: Effect of Oral Melatonin. Molecules. 2018; 23(8):1845. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23081845
Chicago/Turabian StyleZhao, Xiaoxue, Dian Wang, Zhenzheng Wu, Bo Pan, Haoxuan Yang, Changjun Zeng, Ming Zhang, Guoshi Liu, Hongbing Han, and Guangbin Zhou. 2018. "Female Reproductive Performance in the Mouse: Effect of Oral Melatonin" Molecules 23, no. 8: 1845. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23081845