Antioxidant Activity of Selected Polyphenolics in Yeast Cells: The Case Study of Montenegrin Merlot Wine

 , , ,
, , ,
Abstract
:1. Introduction
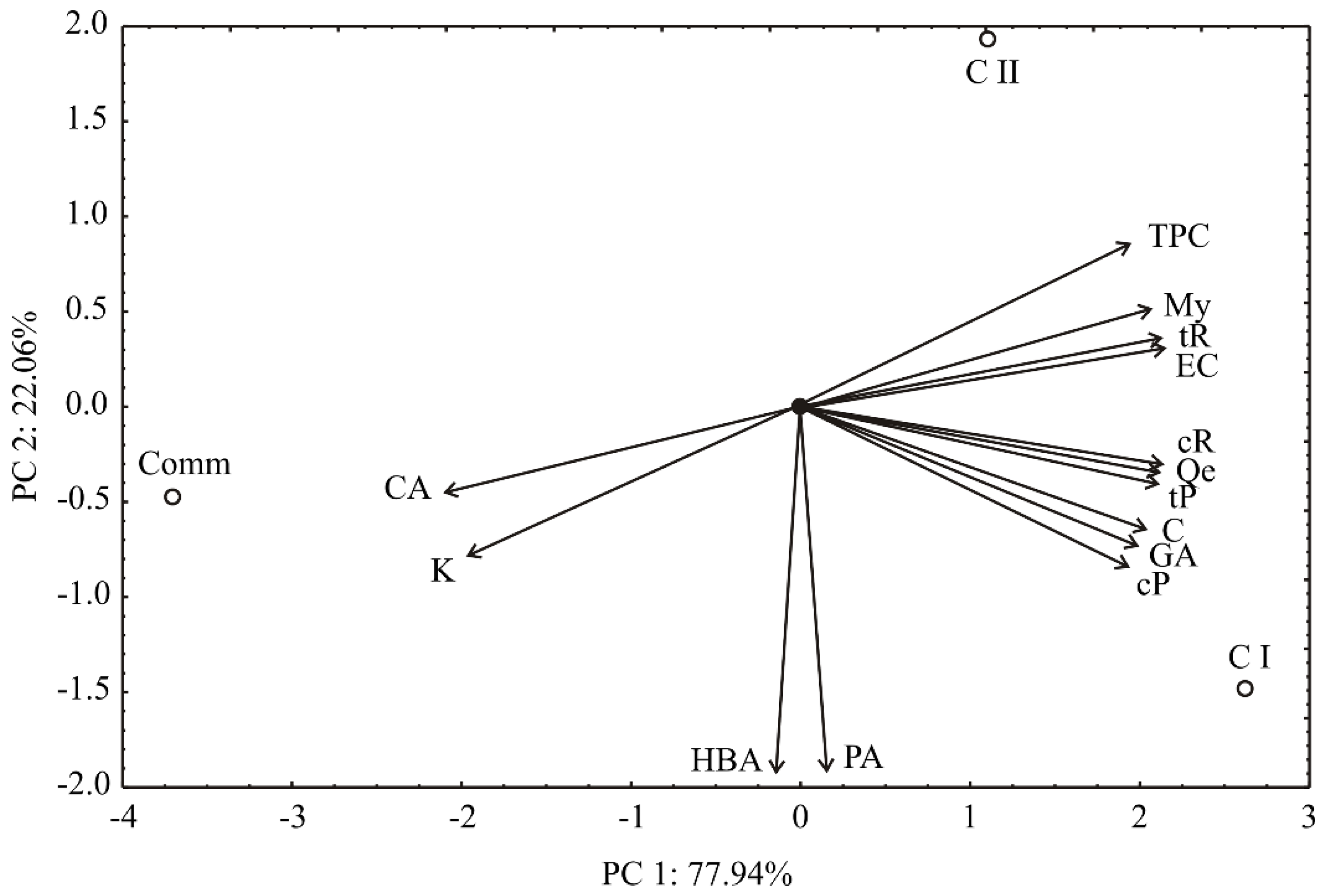
2. Results and Discussion
2.1. Phenolic Profile and In Vitro Antioxidant Activity
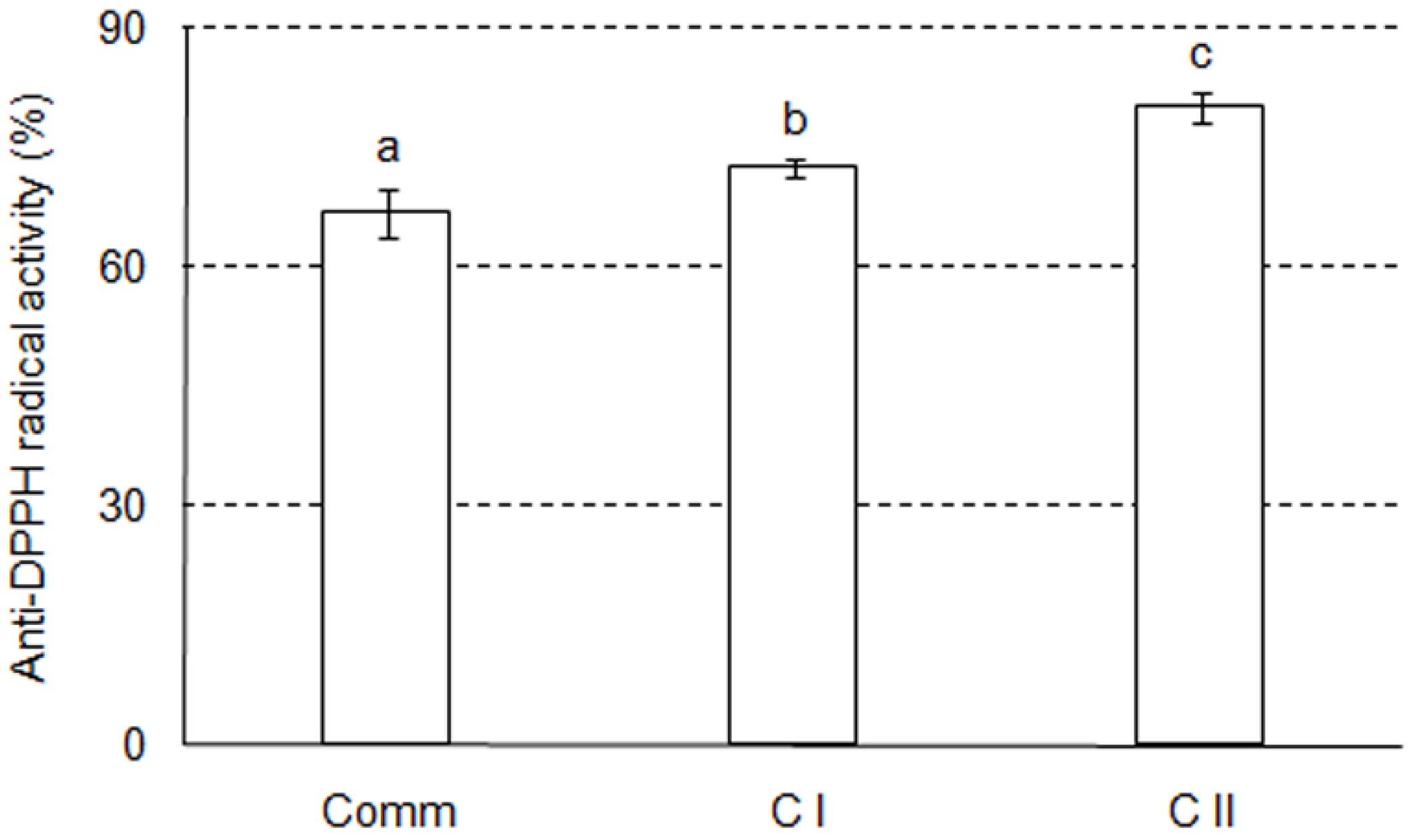
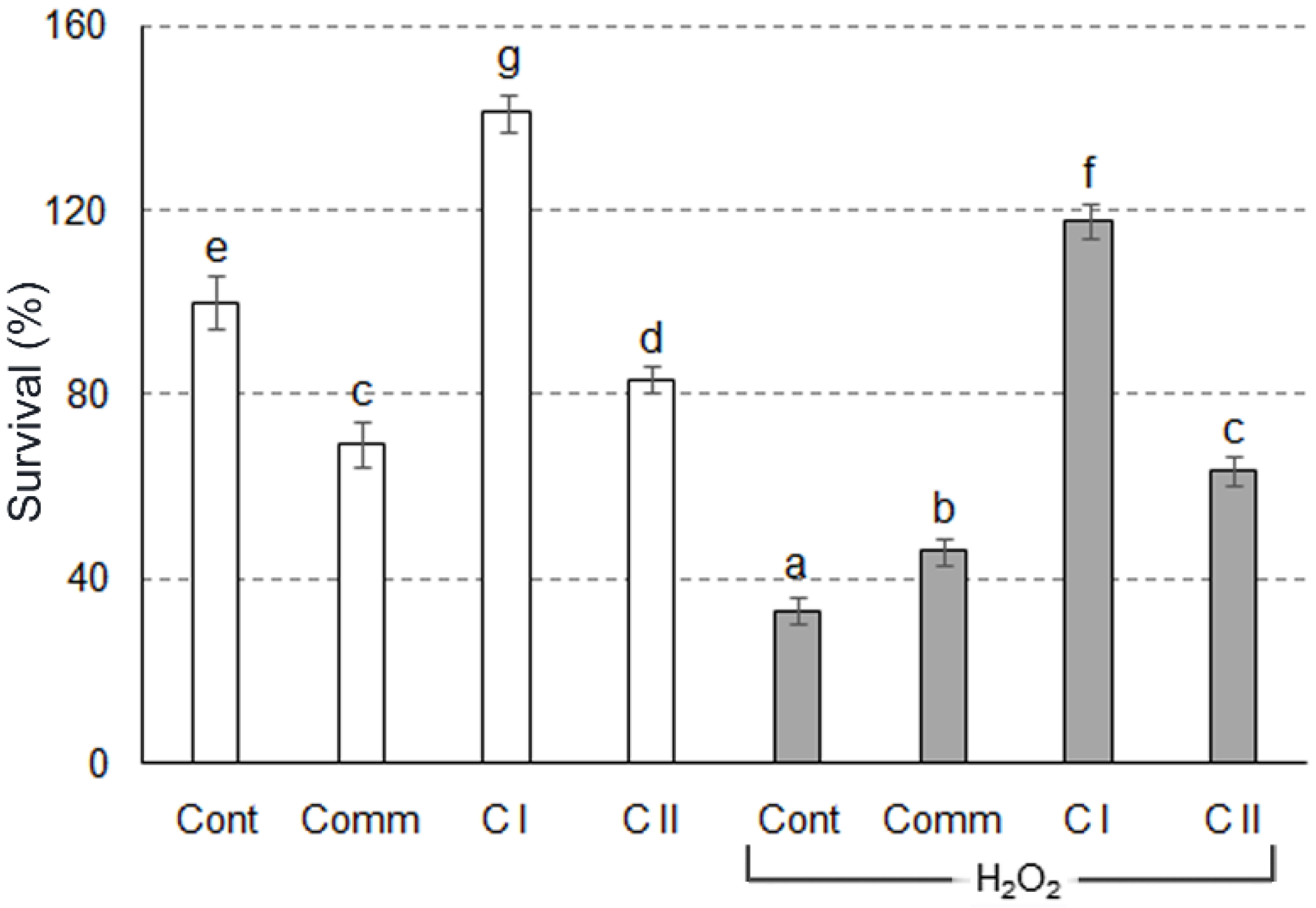
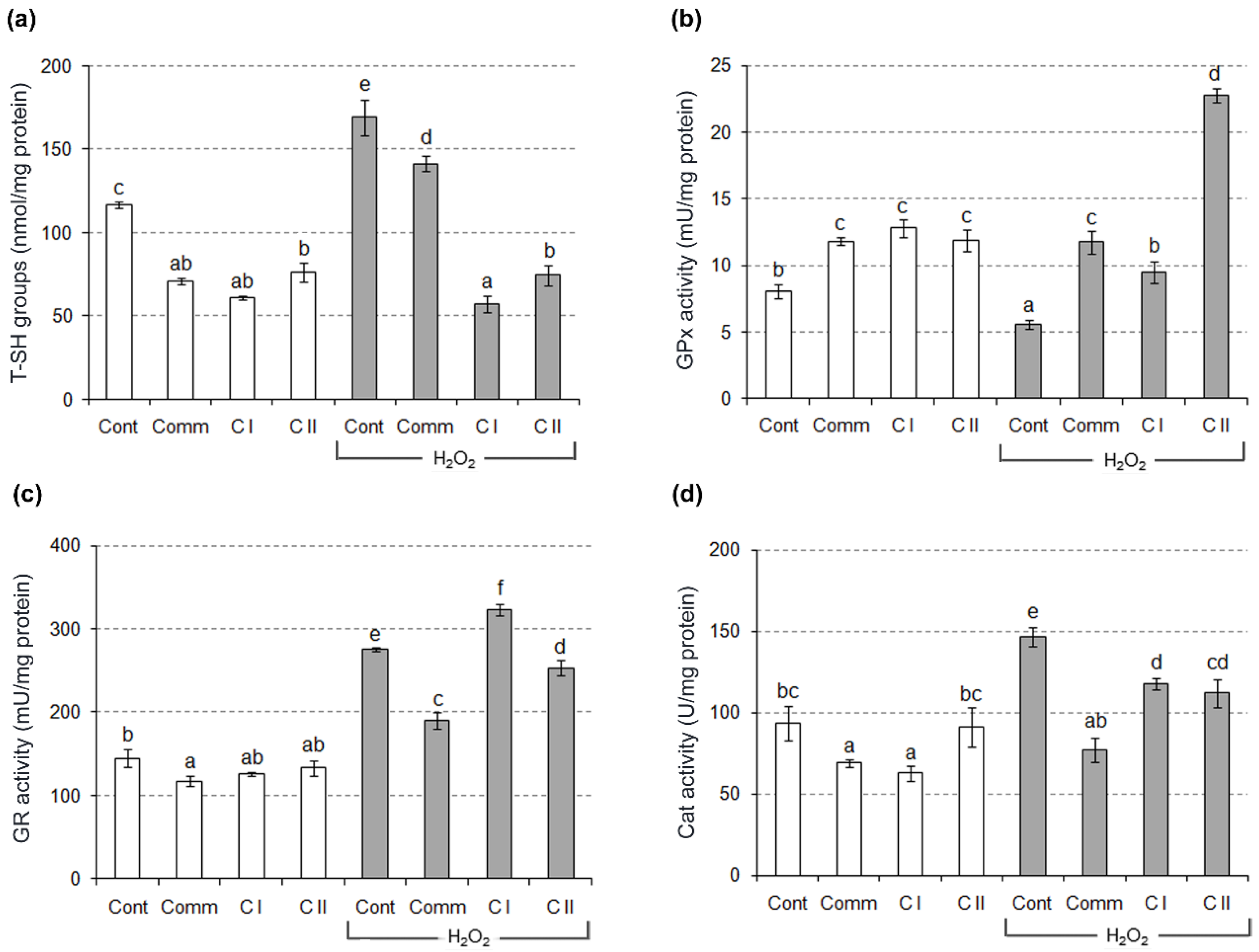
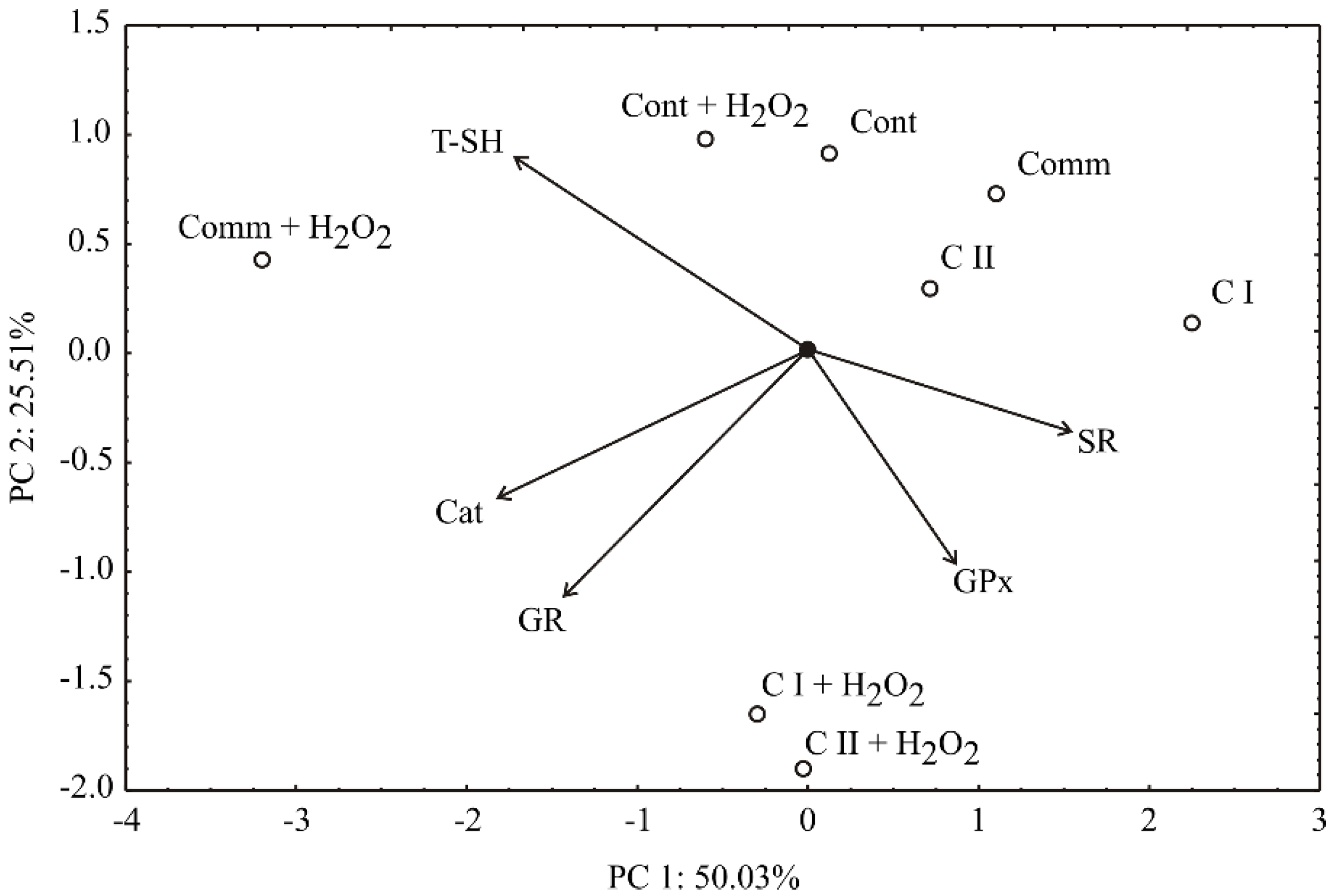
2.2. Antioxidant Activity in Cell Culture
2.3. Neurons in the ANN Hidden Layer
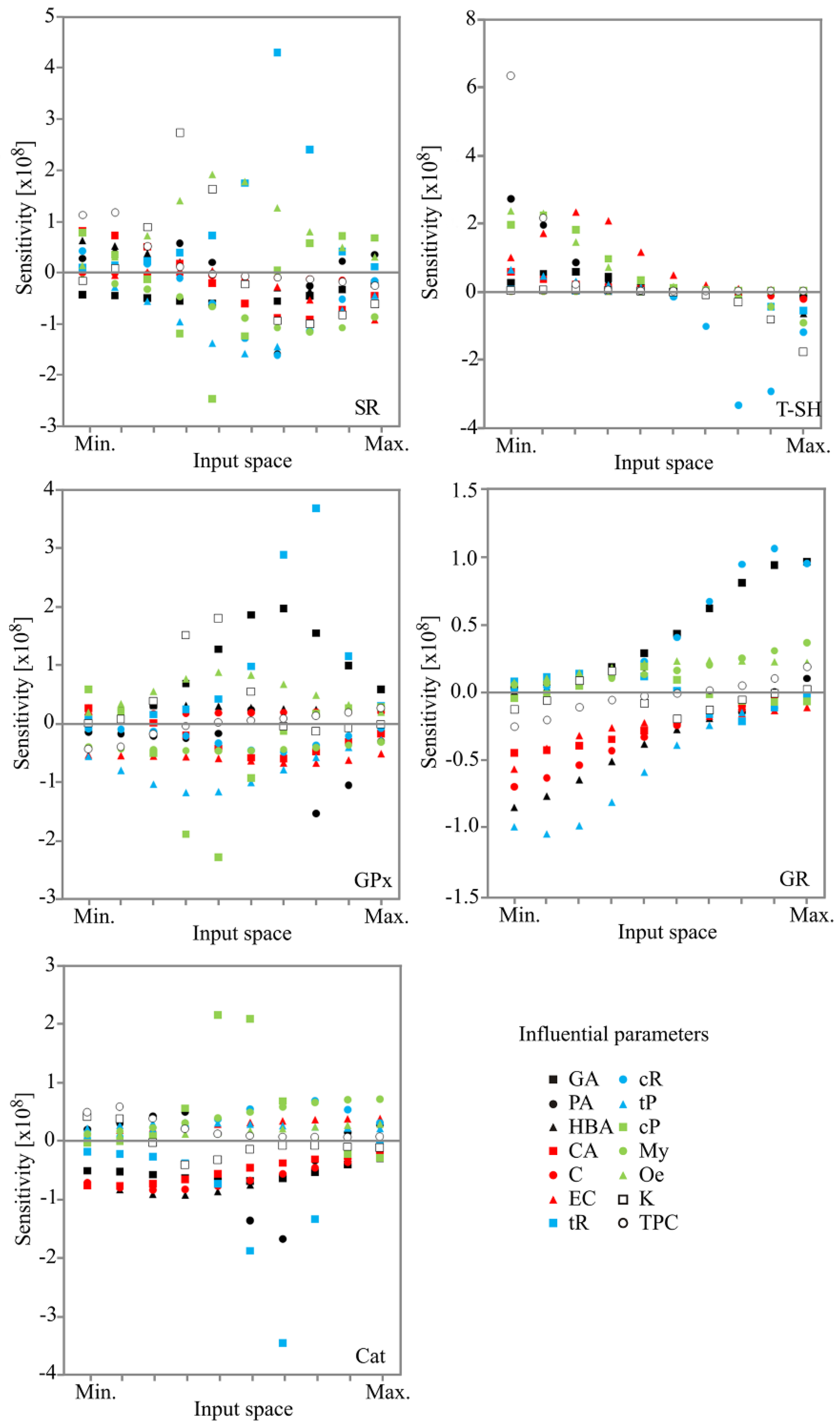
2.4. Sensitivity Analysis
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Wine Samples
3.3. Total Phenolic Content
3.4. Total Flavonoid Content
3.5. Total Monomeric Anthocyanin Content
3.6. Anti–DPPH Radical Activity
3.7. Cellular Assay for the Evaluation of Antioxidant Activity
3.8. Preparation of Cell–Free Extracts
3.9. Determination of T–SH Groups
3.10. Determination of GPx, GR and Cat Activity
3.11. Statistical Analyses
3.12. PCA Data Analysis
3.13. ANN Modeling
3.14. Sensitivity Analysis
3.15. Accuracy of the Models
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Giovinazzo, G.; Grieco, F. Functional Properties of Grape and Wine Polyphenols. Plant Foods Hum. Nutr. 2015, 70, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.H.; Jiang, Y.M.; Shi, J.; Tomás-Barberán, F.A.; Datta, N.; Singanusong, R.; Chen, S.S. Flavonoids in food and their health benefits. Plant Foods Hum. Nutr. 2004, 59, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell. Longev. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, R.; Miranda, A.; Vergara, L. Modulation of endogenous antioxidant system by wine polyphenols in human disease. Clin. Chim. Acta 2011, 412, 410–424. [Google Scholar] [CrossRef] [PubMed]
- Đorđević, N.O.; Novaković, M.M.; Pejin, B.; Mutić, J.J.; Vajs, V.E.; Pajović, S.B.; Tešević, V.V. Comparative analytical study of the selected wine varieties grown in Montenegro. Nat. Prod. Res. 2017, 31, 1825–1830. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.-Y.; Ho, C.-T.; Chen, Y.-K. Biological actions and molecular effects of resveratrol, pterostilbene, and 3′-hydroxypterostilbene. J. Food Drug Anal. 2017, 25, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Ignea, C.; Dorobantu, C.M.; Mintoff, C.P.; Branza-Nichita, N.; Ladomery, M.R.; Kefalas, P.; Chedea, V.S. Modulation of the antioxidant/pro-oxidant balance, cytotoxicity and antiviral actions of grape seed extracts. Food Chem. 2013, 141, 3967–3976. [Google Scholar] [CrossRef] [PubMed]
- Martorell, P.; Forment, J.V.; De Llanos, R.; Montón, F.; Llopis, S.; González, N.; Genovés, S.; Cienfuegos, E.; Monzó, H.; Ramón, D. Use of Saccharomyces cerevisiae and Caenorhabditis elegans as model organisms to study the effect of cocoa polyphenols in the resistance to oxidative stress. J. Agric. Food Chem. 2011, 59, 2077–2085. [Google Scholar] [CrossRef] [PubMed]
- Sastre, J.; Pallardo, F.V.; Viña, J. Glutathione. In The Handbook of Environmental Chemistry; Barceló, D., Kostianoy, A.G., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; Volume 2, pp. 91–108. [Google Scholar]
- Jamieson, D.J. Oxidative stress responses of the yeast Saccharomyces cerevisiae. Yeast 1998, 14, 1511–1527. [Google Scholar] [CrossRef]
- Guiné, R.P.F.; Barroca, M.J.; Gonçalves, F.J.; Alves, M.; Oliveira, S.; Mendes, M. Artificial neural network modelling of the antioxidant activity and phenolic compounds of bananas submitted to different drying treatments. Food Chem. 2015, 168, 454–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cimpoiu, C.; Cristea, V.M.; Hosu, A.; Sandru, M.; Seserman, L. Antioxidant activity prediction and classification of some teas using artificial neural networks. Food Chem. 2011, 127, 1323–1328. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Liu, W.; Tian, S. Artificial neural network optimization of Althaea rosea seeds polysaccharides and its antioxidant activity. Int. J. Biol. Macromol. 2014, 70, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, A.C.; Prieto, J.M. Application of artificial neural networks to the prediction of the antioxidant activity of essential oils in two experimental in vitro models. Food Chem. 2010, 118, 141–146. [Google Scholar] [CrossRef]
- Almeida, J.S. Predictive non-linear modeling of complex data by artificial neural networks. Curr. Opin. Biotechnol. 2002, 13, 72–76. [Google Scholar] [CrossRef]
- Đorđević, N.O.; Pejin, B.; Novaković, M.M.; Stanković, D.M.; Mutić, J.J.; Pajović, S.B.; Tešević, V.V. Some chemical characteristics and antioxidant capacity of novel Merlot wine clones developed in Montenegro. Sci. Hortic. 2017, 225, 505–511. [Google Scholar] [CrossRef]
- Badhani, B.; Sharma, N.; Kakkar, R. Gallic acid: A versatile antioxidant with promising therapeutic and industrial applications. RSC Adv. 2015, 5, 27540–27557. [Google Scholar] [CrossRef]
- Liu, Y.; Pukala, T.L.; Musgrave, I.F.; Williams, D.M.; Dehle, F.C.; Carver, J.A. Gallic acid is the major component of grape seed extract that inhibits amyloid fibril formation. Bioorg. Med. Chem. Lett. 2013, 23, 6336–6340. [Google Scholar] [CrossRef] [PubMed]
- Jara-Palacios, M.J.; Hernanz, D.; Escudero-Gilete, M.L.; Heredia, F.J. The use of grape seed byproducts rich in flavonoids to improve the antioxidant potential of red wines. Molecules 2016, 21, 1526. [Google Scholar] [CrossRef] [PubMed]
- Pasetto, S.; Pardi, V.; Murata, R.M. Anti-HIV-1 activity of flavonoid myricetin on HIV-1 infection in a dual-chamber in vitro model. PLoS ONE 2014, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Chopra, M.; Fitzsimons, P.E.; Strain, J.J.; Thurnham, D.I.; Howard, A.N. Nonalcoholic red wine extract and quercetin inhibit LDL oxidation without affecting plasma antioxidant vitamin and carotenoid concentrations. Clin. Chem. 2000, 46, 1162–1170. [Google Scholar] [PubMed]
- Bhat, K.P.L.; Kosmeder, J.W.; Pezzuto, J.M. Biological Effects of Resveratrol. Antioxid. Redox Signal. 2001, 3, 1041–1064. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Kiyozuka, Y.; Uemura, Y.; Senzaki, H.; Shikata, N.; Hioki, K.; Tsubura, A. Resveratrol inhibits human breast cancer cell growth and may mitigate the effect of linoleic acid, a potent breast cancer cell stimulator. J. Cancer Res. Clin. Oncol. 2001, 127, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Rao, C.V.; Desai, D.; Simi, B.; Kulkarni, N.; Amin, S.; Reddy, B.S. Inhibitory Effect of Caffeic Acid Esters on Azoxymethane-induced Biochemical Changes and Aberrant Crypt Foci Formation in Rat Colon. Cancer Res. 1993, 53, 4182–4188. [Google Scholar] [PubMed]
- Brito, A.; Areche, C.; Sepúlveda, B.; Kennelly, E.J.; Simirgiotis, M.J. Anthocyanin Characterization, Total Phenolic Quantification and Antioxidant Features of Some Chilean Edible Berry Extracts. Molecules 2014, 19, 10936–10955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radovanović, A.; Radovanović, B.; Jovančićević, B. Free radical scavenging and antibacterial activities of southern Serbian red wines. Food Chem. 2009, 117, 326–331. [Google Scholar] [CrossRef]
- Pojer, E.; Mattivi, F.; Johnson, D.; Stockley, C.S. The case for anthocyanin consumption to promote human health: A review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 483–508. [Google Scholar] [CrossRef]
- Niki, E. Assessment of antioxidant capacity in vitro and in vivo. Free Radic. Biol. Med. 2010, 49, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Slatnar, A.; Jakopic, J.; Stampar, F.; Veberic, R.; Jamnik, P. The Effect of Bioactive Compounds on In Vitro and In Vivo Antioxidant Activity of Different Berry Juices. PLoS ONE 2012, 7, e47880. [Google Scholar] [CrossRef] [PubMed]
- Hugo, P.C.; Gil-Chávez, J.; Sotelo-Mundo, R.R.; Namiesnik, J.; Gorinstein, S.; González-Aguilar, G.A. Antioxidant interactions between major phenolic compounds found in “Ataulfo” mango pulp: Chlorogenic, gallic, protocatechuic and vanillic acids. Molecules 2012, 17, 12657–12664. [Google Scholar] [CrossRef]
- Baroni, M.V.; Di Paola Naranjo, R.D.; García-Ferreyra, C.; Otaiza, S.; Wunderlin, D.A. How good antioxidant is the red wine? Comparison of some in vitro and in vivo methods to assess the antioxidant capacity of Argentinean red wines. LWT Food Sci. Technol. 2012, 47, 1–7. [Google Scholar] [CrossRef]
- Lingua, M.S.; Fabani, M.P.; Wunderlin, D.A.; Baroni, M.V. In vivo antioxidant activity of grape, pomace and wine from three red varieties grown in Argentina: Its relationship to phenolic profile. J. Funct. Foods 2016, 20, 332–345. [Google Scholar] [CrossRef]
- Saltelli, A.; Annoni, P. How to avoid a perfunctory sensitivity analysis. Environ. Model. Softw. 2010, 25, 1508–1517. [Google Scholar] [CrossRef]
- Sadi, G.; Bozan, D.; Yildiz, H.B. Redox regulation of antioxidant enzymes: Post-translational modulation of catalase and glutathione peroxidase activity by resveratrol in diabetic rat liver. Mol. Cell. Biochem. 2014, 393, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Ourique, G.M.; Finamor, I.A.; Saccol, E.M.H.; Riffel, A.P.K.; Pês, T.S.; Gutierrez, K.; Gonçalves, P.B.D.; Baldisserotto, B.; Pavanato, M.A.; Barreto, K.P. Resveratrol improves sperm motility, prevents lipid peroxidation and enhances antioxidant defences in the testes of hyperthyroid rats. Reprod. Toxicol. 2013, 37, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Radović, B.R.; Tešević, V.T.; Kodžulović, V.K.; Maraš, V.M. Resveratrol concentration in ‘Vranac’ wines. Vitis 2015, 54, 169–171. [Google Scholar]
- Lee, E.J.; Nomura, N.; Patil, B.S.; Yoo, K.S. Measurement of total phenolic content in wine using an automatic Folin-Ciocalteu assay method. Int. J. Food Sci. Technol. 2014, 49, 2364–2372. [Google Scholar] [CrossRef]
- Stanisavljević, N.; Samardžić, J.; Janković, T.; Šavikin, K.; Mojsin, M.; Topalović, V.; Stevanović, M. Antioxidant and antiproliferative activity of chokeberry juice phenolics during in vitro simulated digestion in the presence of food matrix. Food Chem. 2015, 175, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, J.; Lindsay, R.H. Estimation of Total, Protein-Bound, and Nonprotein Sulfhydryl Groups in Tissue with Ellman’s Reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Havir, E.A.; McHale, N.A. Biochemical and developmental characterization of multiple forms of catalase in tobacco leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Abdi, H.; Williams, L.J. Principal component analysis. Wiley Interdiscip. Rev. Comput. Stat. 2010, 2, 433–459. [Google Scholar] [CrossRef] [Green Version]
- Pezo, L.L.; Ćurčić, B.L.; Filipović, V.S.; Nićetin, M.R.; Koprivica, G.B.; Mišljenović, N.M.; Lević, L.B. Artificial neural network model of pork meat cubes osmotic dehydration. Hem. Ind. 2013, 67, 465–475. [Google Scholar] [CrossRef]
- Hu, X.; Weng, Q. Estimating impervious surfaces from medium spatial resolution imagery using the self-organizing map and multi-layer perceptron neural networks. Remote Sens. Environ. 2009, 113, 2089–2102. [Google Scholar] [CrossRef]
- Smith, J.T. Neural network verification. In Methods and Procedures for the Verification and Validation of Artificial Neural Networks; Taylor, B.J., Ed.; Springer Science + Business Media: New York, NY, USA, 2006; pp. 109–169. [Google Scholar]
Sample Availability: Not available. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Content | Comm | C I | C II | |
|---|---|---|---|---|
| Total phenolic content | TPC | 2988.6 ± 142.2 a | 3255.7 ± 90.8 a,b | 3337.9 ± 115.4 b |
| Total flavonoid content | TFC | 376.0 ± 11.2 a | 393.2 ± 5.8 a | 397.2 ± 9.2 a |
| Total monomeric anthocyanin content | TMA | 215.2 ± 4.1 a | 203.6 ± 4.5 a | 230.3 ± 6.6 b |
| χ2 | RMSE | MBE | MPE | r2 | Skew. | Kurt. | Mean | SD | Var. | |
|---|---|---|---|---|---|---|---|---|---|---|
| SR | 29.678 | 5.136 | 0.713 | 4.230 | 0.976 | −0.565 | 2.797 | 0.713 | 5.183 | 26.867 |
| T–SH | 1115.430 | 31.488 | −2.696 | 0.950 | 0.957 | −0.607 | 0.630 | −2.696 | 31.970 | 1022.080 |
| GPx | 0.027 | 0.156 | 0.005 | 1.013 | 0.923 | 0.688 | −0.183 | 0.005 | 0.159 | 0.025 |
| GR | 2.935 | 1.615 | −0.103 | 1.116 | 0.960 | 0.034 | −1.049 | −0.103 | 1.643 | 2.698 |
| Cat | 6.120 | 2.332 | −0.004 | 2.426 | 0.967 | 0.642 | 0.635 | −0.004 | 2.377 | 5.649 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Đorđević, N.O.; Todorović, N.; Novaković, I.T.; Pezo, L.L.; Pejin, B.; Maraš, V.; Tešević, V.V.; Pajović, S.B. Antioxidant Activity of Selected Polyphenolics in Yeast Cells: The Case Study of Montenegrin Merlot Wine. Molecules 2018, 23, 1971. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23081971
Đorđević NO, Todorović N, Novaković IT, Pezo LL, Pejin B, Maraš V, Tešević VV, Pajović SB. Antioxidant Activity of Selected Polyphenolics in Yeast Cells: The Case Study of Montenegrin Merlot Wine. Molecules. 2018; 23(8):1971. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23081971
Chicago/Turabian StyleĐorđević, Neda O., Nevena Todorović, Irena T. Novaković, Lato L. Pezo, Boris Pejin, Vesna Maraš, Vele V. Tešević, and Snežana B. Pajović. 2018. "Antioxidant Activity of Selected Polyphenolics in Yeast Cells: The Case Study of Montenegrin Merlot Wine" Molecules 23, no. 8: 1971. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23081971