Computational Molecular Docking and X-ray Crystallographic Studies of Catechins in New Drug Design Strategies

Abstract
:1. Introduction
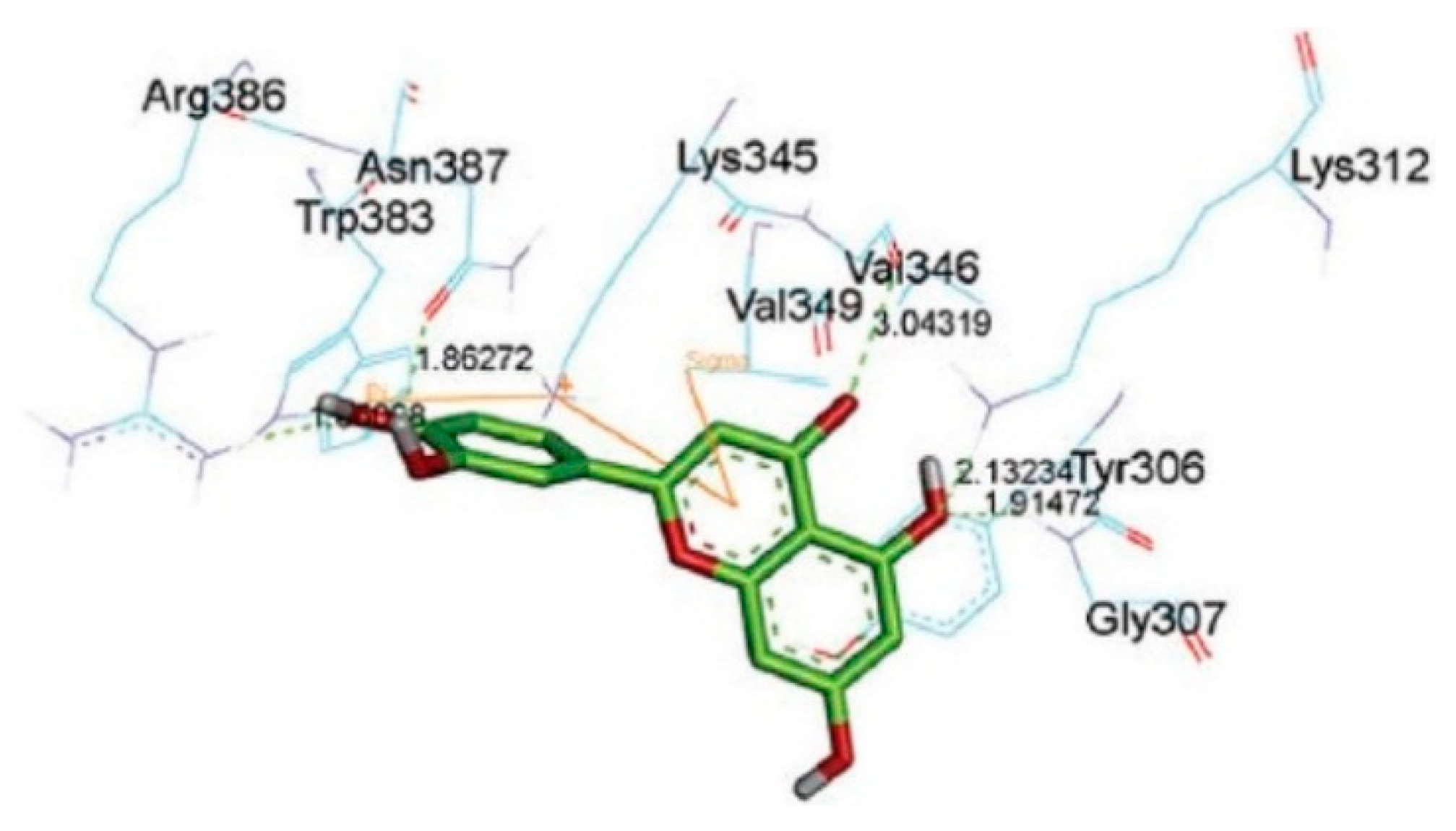
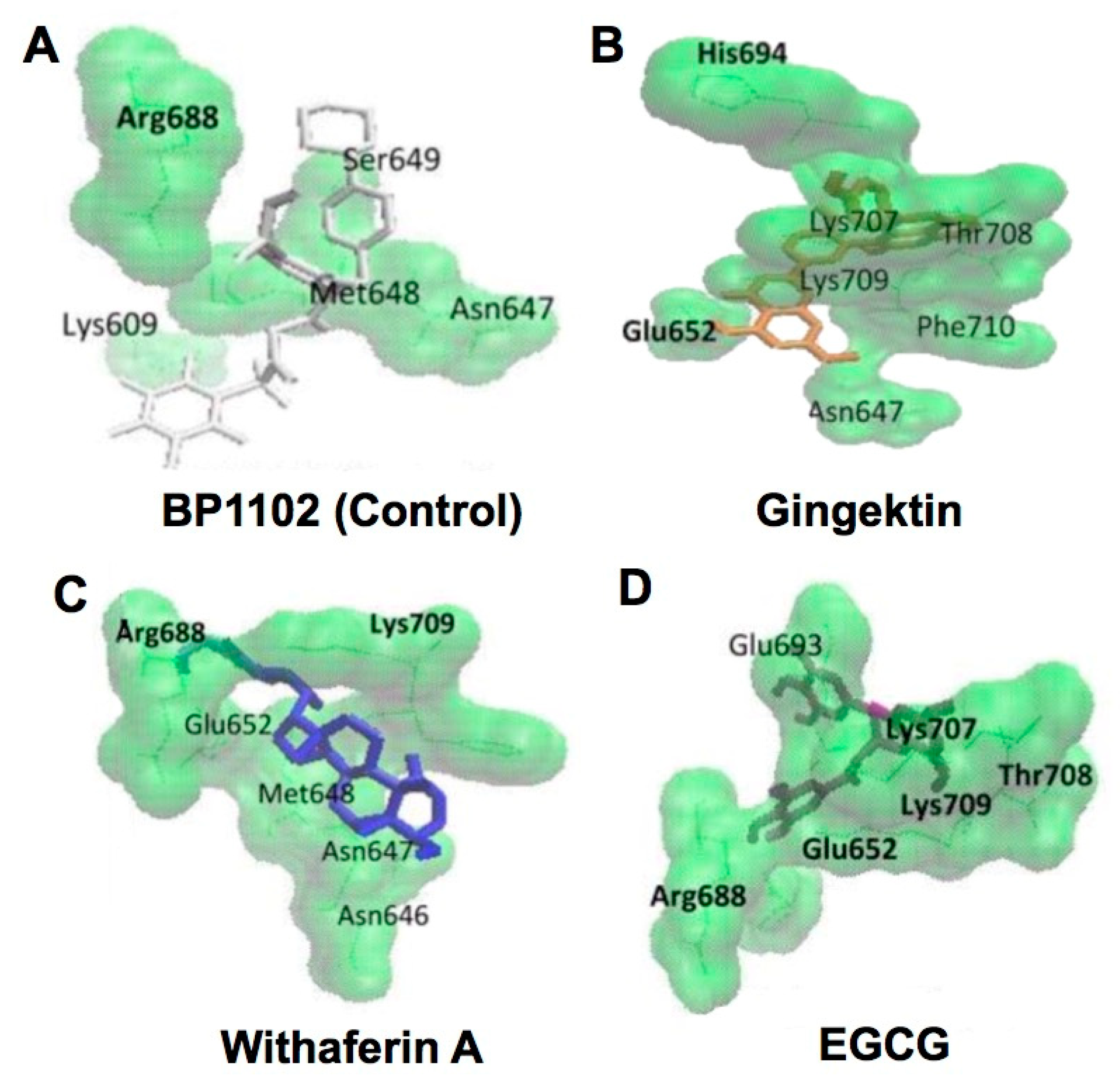
2. CMDA of Catechin-Protein Interaction
2.1. Interaction of Catechins and Cancer-Related Proteins
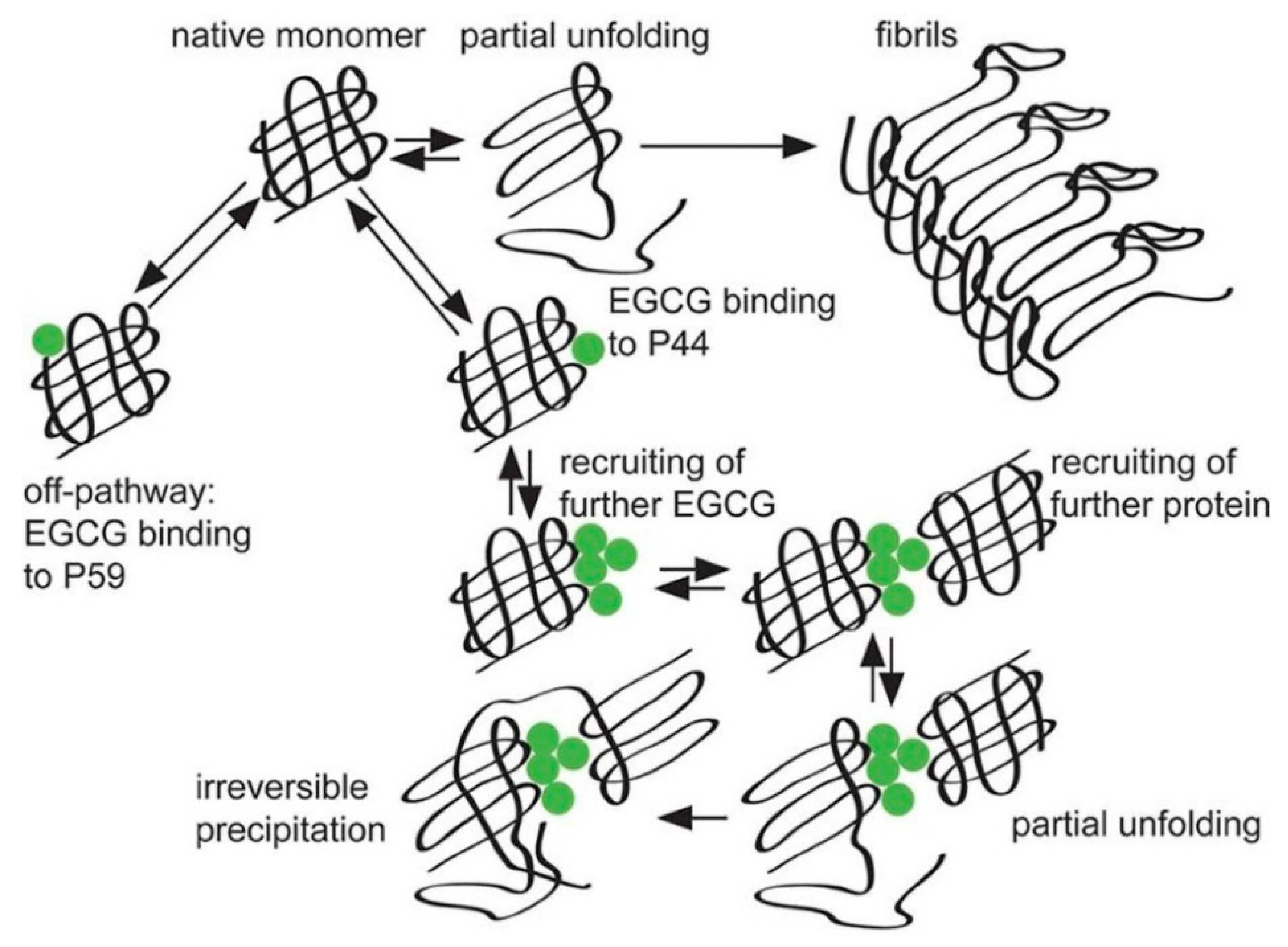
2.2. Interaction between Catechins and Proteins Related to Neurodegenerative Diseases
2.3. Interaction of Catechins and MetS-Related Proteins
2.4. Interaction of Catechins with Proteins Related to Infectious Diseases
2.5. Interaction of Catechins with Proteins Related to Immunological Disorders
2.6. Miscellaneous
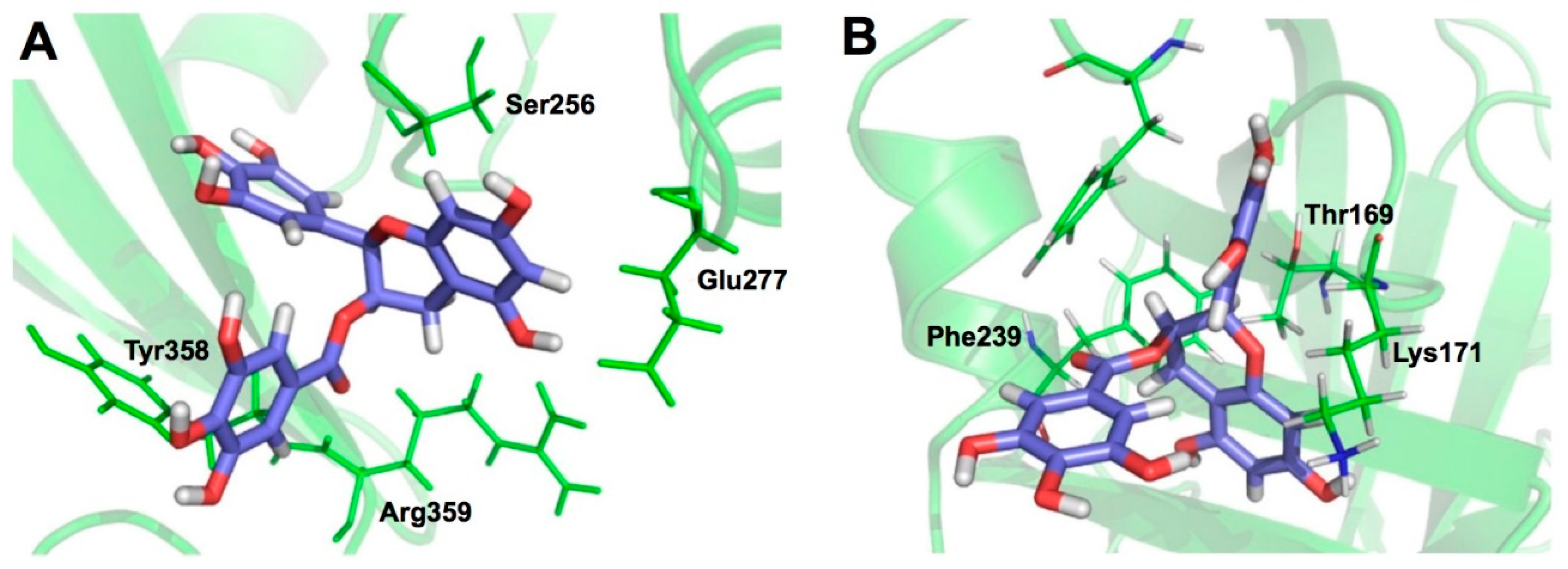
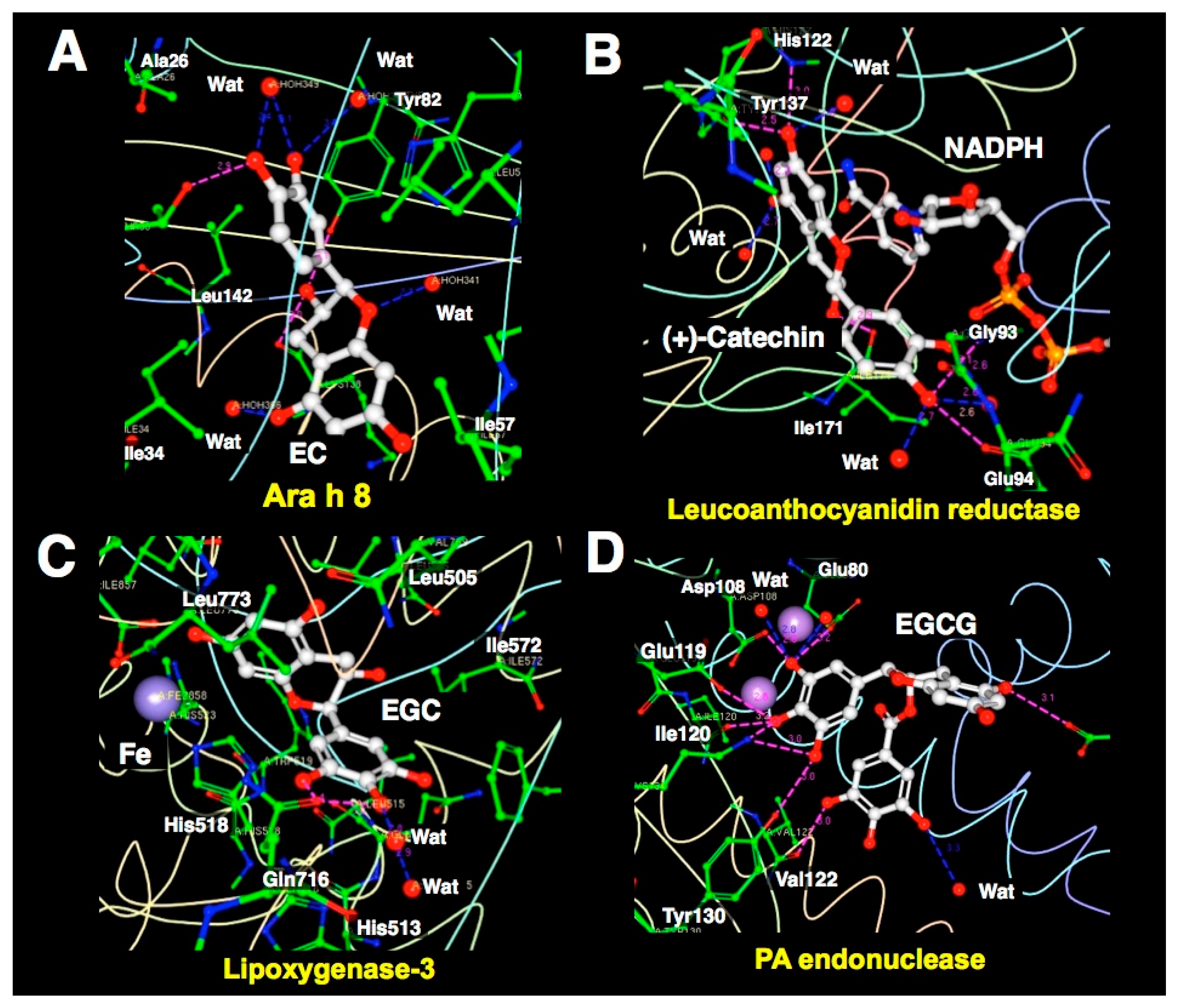
3. X-ray Crystallographic Analysis (XCA) of Protein-Catechin Complexes
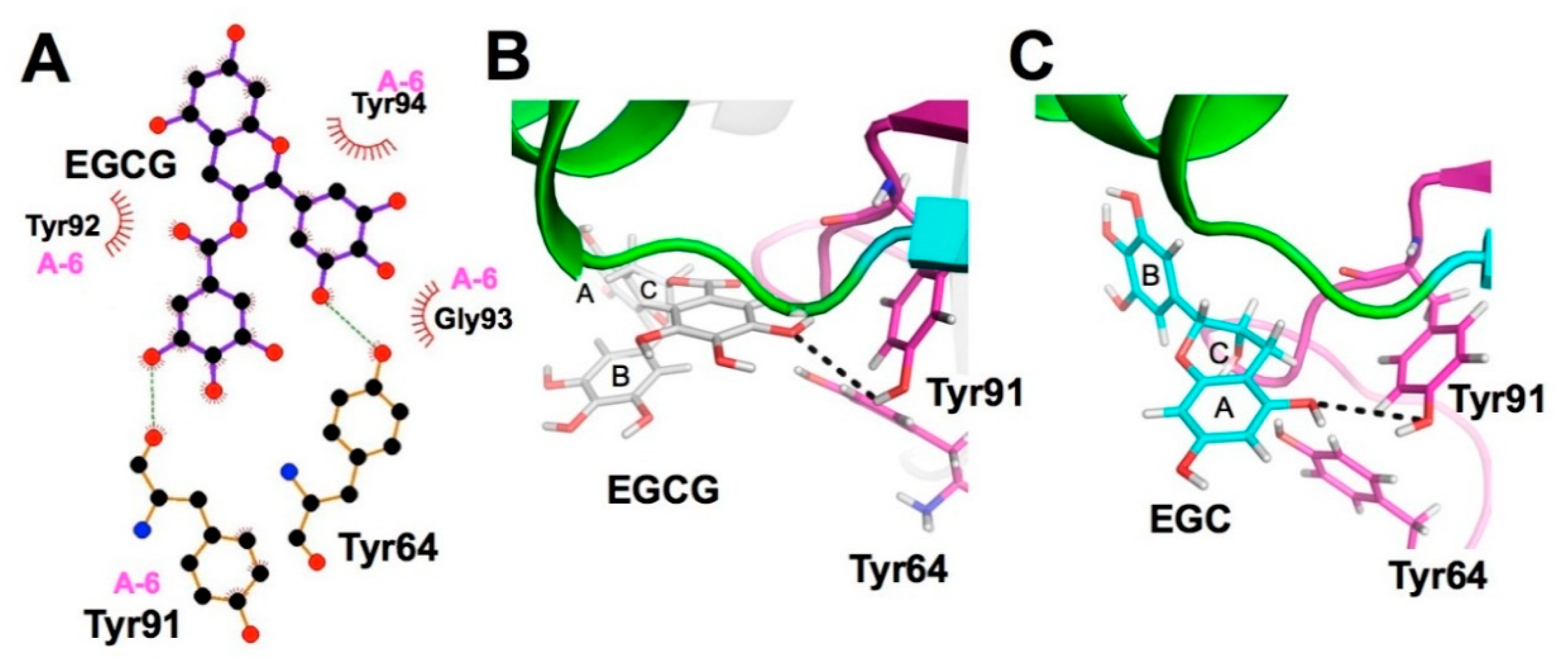
3.1. XCA of (+)-Catechin-and EC-Protein Complexes
3.2. XCA of EGC-Protein Complex
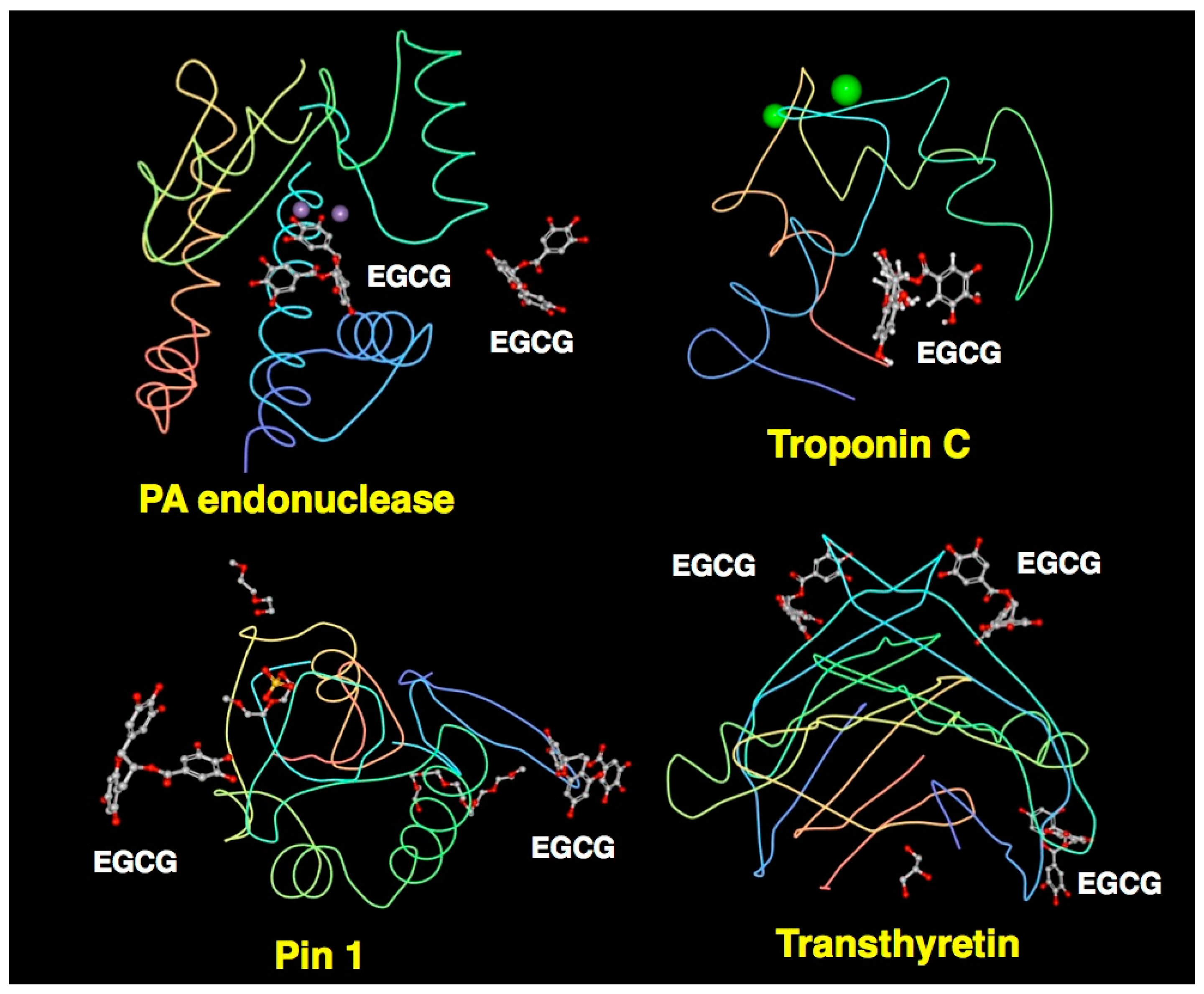
3.3. XCA of EGCG-Protein Complexes
4. Characteristics of Catechin-Protein Binding Interaction Revealed by XCA
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 3CLP | 3C-like protease |
| AChE | acetylcholinesterase |
| AD | Alzheimer’s disease |
| BChE | butyrylcholinesterase |
| CDK | cyclin-dependent kinase |
| CMDA | computational molecular docking analysis |
| CYP3A4 | cytochrome P-3A4 |
| DS | Down syndrome |
| EC | (−)-epicatechin |
| ECG | (−)-epicatechin gallate |
| EGC | (−)-epigallocatechin |
| EGCG | (−)-epigallocatechin-3-O-gallate |
| GCG | gallocatechin-3-O-gallate |
| GDH | glutamate dehydrogenase |
| GTCs | green tea catechins |
| HIV-1 | human immunodeficiency virus type 1 |
| HSOS | hepatic sinusoidal obstruction syndrome |
| Hsp | heat shock protein |
| IAV | influenza A virus |
| LPS | lipopolysaccharide |
| MAPK | p38 mitogen-activated protein kinase |
| MCT | monocrotaline |
| MD2 | myeloid differentiation factor 2 |
| MetS | metabolic syndrome |
| MMP | matrix metalloproteinase |
| MRSA | multidrug-resistant Staphylococcus aureus |
| NF | nuclear factor |
| NGF | nerve growth factor |
| Nrf2 | nuclear factor erythroid 2-related factor 2 |
| OPPA | oligopeptide ABC transporter binding lipoprotein |
| PBD | epicatechin-3-O-gallate-(4β→8)-epicatechin-3′-O-gallate |
| PD | Parkinson’s disease |
| PDB | protein data base |
| Pin1 | peptidyl prolyl cis/trans isomerase 1 |
| PKC | protein kinase C |
| PLY | pneumolysin |
| PPA | pancreatic α-amylase |
| PPO | polyphenol oxidase |
| ROS | reactive oxygen species |
| RSK2 | 90-kDa ribosomal S6 kinase |
| SA | serum albumin |
| SEA | Staphylococcal enterotoxin A |
| SIRT6 | sirtuin-6 |
| SOD1 | superoxide dismutase 1 |
| SrtA | sortase A |
| STAT | signal transducer and activator of transcription |
| SULT | sulfotransferase |
| TNF | tumor necrosis factor |
| TRAF6 | TNF receptor associated factor 6 |
| TLR4 | toll-like receptor 4 |
| VEGF | vascular endothelial growth factor |
References
- Suzuki, T.; Pervin, M.; Goto, S.; Isemura, M.; Nakamura, Y. Beneficial Effects of Tea and the Green Tea Catechin Epigallocatechin-3-gallate on Obesity. Molecules 2016, 21, 1305. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Zhang, J.; Zhang, L.; Huang, J.; Wang, Y. Mechanisms of body weight reduction and metabolic syndrome alleviation by tea. Mol. Nutr. Food Res. 2016, 60, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Wang, H. Cancer Preventive Activities of Tea Catechins. Molecules 2016, 21, 1679. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Miyoshi, N.; Hayakawa, S.; Imai, S.; Isemura, M.; Nakamura, Y. Health Benefits of Tea Consumption. In Beverage Impacts on Health and Nutrition; Wilson, T., Templ, N.J., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 29–47. ISBN 9783319236728. [Google Scholar]
- Saeki, K.; Hayakawa, S.; Nakano, S.; Ito, S.; Oishi, Y.; Suzuki, Y.; Isemura, M. In Vitro and In Silico Studies of the Molecular Interactions of Epigallocatechin-3-O-gallate (EGCG) with Proteins That Explain the Health Benefits of Green Tea. Molecules 2018, 23, 1295. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.R.; Bauer, B.A.; Vincent, A.; Limburg, P.J.; Wilson, T. Reading the tea leaves: Anticarcinogenic properties of (−)-epigallocatechin-3-gallate. Mayo Clin. Proc. 2007, 82, 725–732. [Google Scholar] [CrossRef]
- Lambert, J.D.; Hong, J.; Yang, G.Y.; Liao, J.; Yang, C.S. Inhibition of carcinogenesis by polyphenols: Evidence from laboratory investigations. Am. J. Clin. Nutr. 2005, 81 (Suppl. S1), 284S–291S. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, H.; Pervin, M.; Goto, S.; Isemura, M.; Nakamura, Y. Beneficial Effects of Plant Polyphenols on Obesity. Obes. Control Ther. 2017, 4, 1–16. [Google Scholar] [CrossRef]
- Arora, J.P.; Singhal, V.K.; Chand, M.; Laxmi; Pal, C. Study of interaction between catechin and native and modified bovine serum albumin by physico-chemical methods. Indian J. Biochem. Biophys. 1989, 26, 14–18. [Google Scholar] [PubMed]
- Maiti, T.K.; Ghosh, K.S.; Dasgupta, S. Interaction of (−)-epigallocatechin-3-gallate with human serum albumin: fluorescence, fourier transform infrared, circular dichroism, and docking studies. Proteins 2006, 64, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Hagerman, A.E. Role of the flavan-3-ol and galloyl moieties in the interaction of (−)-epigallocatechin gallate with serum albumin. J. Agric. Food Chem. 2014, 62, 3768–3775. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Ueda-Wakagi, M.; Hayashibara, K.; Kitano, R.; Kawase, M.; Kaihatsu, K.; Kato, N.; Suhara, Y.; Osakabe, N.; Ashida, H. Substitution at the C-3 Position of Catechins Has an Influence on the Binding Affinities against Serum Albumin. Molecules 2017, 22, 314. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, Y.Y.; Sun, W.J.; Zhang, M.H.; Zheng, Y.F.; Shen, H.M.; Yang, J.; Zhu, X.Q. FBS or BSA Inhibits EGCG Induced Cell Death through Covalent Binding and the Reduction of Intracellular ROS Production. Biomed. Res. Int. 2016, 2016, 5013409. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Tang, H.; Xiao, J.; Cui, F.; Yang, K.; Li, J.; Zhao, Q.; Huang, Q.; Li, Y. Small-Angle X-ray Scattering Study of Protein Complexes with Tea Polyphenols. J. Agric. Food Chem. 2017, 65, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Bu, W.; Tang, Z.Y.; Sun, F.X.; Ye, S.L.; Liu, K.D.; Xue, Q.; Chen, J.; Gao, D.M. Effects of matrix metalloproteinase inhibitor BB-94 on liver cancer growth and metastasis in a patient-like orthotopic model LCI-D20. Hepatogastroenterology 1998, 45, 1056–1061. [Google Scholar] [PubMed]
- Sazuka, M.; Murakami, S.; Isemura, M.; Satoh, K.; Nukiwa, T. Inhibitory effects of green tea infusion on in vitro invasion and in vivo metastasis of mouse lung carcinoma cells. Cancer Lett. 1995, 98, 27–31. [Google Scholar] [CrossRef]
- Taniguchi, S.; Fujiki, H.; Kobayashi, H.; Go, H.; Miyado, K.; Sadano, H.; Shimokawa, R. Effect of (−)-epigallocatechin gallate, the main constituent of green tea, on lung metastasis with mouse B16 melanoma cell lines. Cancer Lett. 1992, 65, 51–54. [Google Scholar] [CrossRef]
- Gupta, S.; Hastak, K.; Ahmad, N.; Lewin, J.S.; Mukhtar, H. Inhibition of prostate carcinogenesis in TRAMP mice by oral infusion of green tea polyphenols. Proc. Natl. Acad. Sci. USA 2001, 98, 10350–10355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, A.; Nandy, S.K.; Sarkar, J.; Chakraborti, T.; Chakraborti, S. Inhibition of pro-/active MMP-2 by green tea catechins and prediction of their interaction by molecular docking studies. Mol. Cell. Biochem. 2017, 427, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, J.; Nandy, S.K.; Chowdhury, A.; Chakraborti, T.; Chakraborti, S. Inhibition of MMP-9 by green tea catechins and prediction of their interaction by molecular docking analysis. Biomed. Pharmacother. 2016, 84, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Maeda-Yamamoto, M.; Kawahara, H.; Tahara, N.; Tsuji, K.; Hara, Y.; Isemura, M. Effects of tea polyphenols on the invasion and matrix metalloproteinases activities of human fibrosarcoma HT1080 cells. J. Agric. Food Chem. 1999, 47, 2350–2354. [Google Scholar] [CrossRef] [PubMed]
- Koivunen, E.; Ristimaki, A.; Itkonen, O.; Osman, S.; Vuento, M.; Stenman, U.H. Tumor-associated trypsin participates in cancer cell-mediated degradation of extracellular matrix. Cancer Res. 1991, 51, 2107–2112. [Google Scholar] [PubMed]
- Cui, F.; Yang, K.; Li, Y. Investigate the binding of catechins to trypsin using docking and molecular dynamics simulation. PLoS ONE 2015, 10, e0125848. [Google Scholar] [CrossRef] [PubMed]
- Daniel, K.G.; Landis-Piwowar, K.R.; Chen, D.; Wan, S.B.; Chan, T.H.; Dou, Q.P. Methylation of green tea polyphenols affects their binding to and inhibitory poses of the proteasome beta5 subunit. Int. J. Mol. Med. 2006, 18, 625–632. [Google Scholar] [PubMed]
- Kongpichitchoke, T.; Chiu, M.T.; Huang, T.C.; Hsu, J.L. Gallic Acid Content in Taiwanese Teas at Different Degrees of Fermentation and Its Antioxidant Activity by Inhibiting PKCdelta Activation: In Vitro and in Silico Studies. Molecules 2016, 21, 1346. [Google Scholar] [CrossRef] [PubMed]
- Kumazoe, M.; Sugihara, K.; Tsukamoto, S.; Huang, Y.; Tsurudome, Y.; Suzuki, T.; Suemasu, Y.; Ueda, N.; Yamashita, S.; Kim, Y.; et al. 67-kDa laminin receptor increases cGMP to induce cancer-selective apoptosis. J. Clin. Invest. 2013, 123, 787–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, S.; Basak, A.; Dasgupta, S. Synthesis and ribonuclease A inhibition activity of resorcinol and phloroglucinol derivatives of catechin and epicatechin: Importance of hydroxyl groups. Bioorg. Med. Chem. 2010, 18, 6538–6546. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.Y.; Yoon, H.; Ahn, S.; Kim, D.W.; Kim, S.H.; Koh, D.; Lee, Y.H.; Lim, Y. Chromenylchalcones showing cytotoxicity on human colon cancer cell lines and in silico docking with aurora kinases. Bioorg. Med. Chem. 2013, 21, 4250–4258. [Google Scholar] [CrossRef] [PubMed]
- Iftikhar, H.; Rashid, S. Molecular docking studies of flavonoids for their inhibition pattern against beta-catenin and pharmacophore model generation from experimentally known flavonoids to fabricate more potent inhibitors for Wnt signaling pathway. Pharmacogn. Mag. 2014, 10 (Suppl. S2), S264–S271. [Google Scholar] [CrossRef] [PubMed]
- Jayadeepa, R.M.; Ray, A.; Naik, D.; Sanyal, D.N.; Shah, D. Review and research analysis of computational target methods using BioRuby and in silico screening of herbal lead compounds against pancreatic cancer using R programming. Curr. Drug Metab. 2014, 15, 535–543. [Google Scholar] [PubMed]
- Dunna, N.R.; Bandaru, S.; Akare, U.R.; Rajadhyax, S.; Gutlapalli, V.R.; Yadav, M.; Nayarisseri, A. Multiclass comparative virtual screening to identify novel Hsp90 inhibitors: A therapeutic breast cancer drug target. Curr. Top. Med. Chem. 2015, 15, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Farhan, M.; Zafar, A.; Chibber, S.; Khan, H.Y.; Arif, H.; Hadi, S.M. Mobilization of copper ions in human peripheral lymphocytes by catechins leading to oxidative DNA breakage: A structure activity study. Arch. Biochem. Biophys. 2015, 580, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yao, K.; Chang, X.; Shim, J.H.; Kim, H.G.; Malakhova, M.; Kim, D.J.; Bode, A.M.; Dong, Z. Computational and Biochemical Discovery of RSK2 as a Novel Target for Epigallocatechin Gallate (EGCG). PLoS ONE 2015, 10, e0130049. [Google Scholar] [CrossRef] [PubMed]
- Abdulrahman, N.; Jaballah, M.; Poomakkoth, N.; Riaz, S.; Abdelaziz, S.; Issa, A.; Mraiche, F. Inhibition of p90 ribosomal S6 kinase attenuates cell migration and proliferation of the human lung adenocarcinoma through phospho-GSK-3beta and osteopontin. Mol. Cell. Biochem. 2016, 418, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Daniels, J.; Kadlubar, S. Sulfotransferase genetic variation: From cancer risk to treatment response. Drug Metab. Rev. 2013, 45, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Cook, I.; Wang, T.; Girvin, M.; Leyh, T.S. The structure of the catechin-binding site of human sulfotransferase 1A1. Proc. Natl. Acad. Sci. USA 2016, 113, 14312–14317. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.; Zhang, J.; Huang, Z.; Sheng, Y.; Ji, L. The involvement of Nrf2 antioxidant signalling pathway in the protection of monocrotaline-induced hepatic sinusoidal obstruction syndrome in rats by (+)-catechin hydrate. Free Radic. Res. 2018, 52, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Ashour, M.L.; Youssef, F.S.; Gad, H.A.; Wink, M. Inhibition of Cytochrome P450 (CYP3A4) Activity by Extracts from 57 Plants Used in Traditional Chinese Medicine (TCM). Pharmacogn. Mag. 2017, 13, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, L.; Xu, Y.; Jiang, S.; Shao, Y. Deciphering the binding behavior of flavonoids to the cyclin dependent kinase 6/cyclin D complex. PLoS ONE 2018, 13, e0196651. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T.; Saeed, M.E.M.; Mirghani, E.; Alim, A.; Yassin, Z.; Saeed, E.; Khalid, H.E.; Daak, S. Integration of phytochemicals and phytotherapy into cancer precision medicine. Oncotarget 2017, 8, 50284–50304. [Google Scholar] [CrossRef] [PubMed]
- Rahnasto-Rilla, M.; Tyni, J.; Huovinen, M.; Jarho, E.; Kulikowicz, T.; Ravichandran, S.; Bohr, V.A.; Ferrucci, L.; Lahtela-Kakkonen, M.; Moaddel, R. Natural polyphenols as sirtuin 6 modulators. Sci. Rep. 2018, 8, 4163. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, R.; Devi, A.; Mishra, S. Molecular docking and molecular dynamics studies reveal structural basis of inhibition and selectivity of inhibitors EGCG and OSU-03012 toward glucose regulated protein-78 (GRP78) overexpressed in glioblastoma. J. Mol. Model. 2015, 21, 272. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; He, Z.; Ermakova, S.; Zheng, D.; Tang, F.; Cho, Y.Y.; Zhu, F.; Ma, W.Y.; Sham, Y.; Rogozin, E.A.; et al. Direct inhibition of insulin-like growth factor-I receptor kinase activity by (−)-epigallocatechin-3-gallate regulates cell transformation. Cancer Epidemiol. Biomark. Prev. 2007, 16, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Moyle, C.W.; Cerezo, A.B.; Winterbone, M.S.; Hollands, W.J.; Alexeev, Y.; Needs, P.W.; Kroon, P.A. Potent inhibition of VEGFR-2 activation by tight binding of green tea epigallocatechin gallate and apple procyanidins to VEGF: Relevance to angiogenesis. Mol. Nutr. Food Res. 2015, 59, 401–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, J.H.; Choi, H.S.; Pugliese, A.; Lee, S.Y.; Chae, J.I.; Choi, B.Y.; Bode, A.M.; Dong, Z. (−)-Epigallocatechin gallate regulates CD3-mediated T cell receptor signaling in leukemia through the inhibition of ZAP-70 kinase. J. Biol. Chem. 2008, 283, 28370–28379. [Google Scholar] [CrossRef]
- Singh, P.; Bast, F. Screening of multi-targeted natural compounds for receptor tyrosine kinases inhibitors and biological evaluation on cancer cell lines, in silico and in vitro. Med. Oncol. 2015, 32, 233. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lei, Z.; Huang, Z.; Zhang, X.; Zhou, Y.; Luo, Z.; Zeng, W.; Su, J.; Peng, C.; Chen, X. Epigallocatechin-3-gallate(EGCG) suppresses melanoma cell growth and metastasis by targeting TRAF6 activity. Oncotarget 2016, 7, 79557–79571. [Google Scholar] [CrossRef]
- Jankun, J.; Selman, S.H.; Swiercz, R.; Skrzypczak-Jankun, E. Why drinking green tea could prevent cancer. Nature 1997, 387, 561. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.M.; Daniel, K.G.; Wang, Z.; Guida, W.C.; Chan, T.H.; Dou, Q.P. Docking studies and model development of tea polyphenol proteasome inhibitors: Applications to rational drug design. Proteins 2004, 54, 58–70. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Sun, Z.; Dong, S.; Liu, Y.; Liu, Y. Molecular interactions between (−)-epigallocatechin gallate analogs and pancreatic lipase. PLoS ONE 2014, 9, e111143. [Google Scholar] [CrossRef] [PubMed]
- Islam, B.; Sharma, C.; Adem, A.; Aburawi, E.; Ojha, S. Insight into the mechanism of polyphenols on the activity of HMGR by molecular docking. Drug Des. Devel. Ther. 2015, 9, 4943–4951. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Chen, T.S.; Lu, T. A comparative reverse docking strategy to identify potential antineoplastic targets of tea functional components and binding mode. Int. J. Mol. Sci. 2011, 12, 5200–5212. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.Z.; Wang, Y.; Ai, N.; Hou, Z.; Sun, Y.; Lu, H.; Welsh, W.; Yang, C.S. Tea polyphenol (−)-epigallocatechin-3-gallate inhibits DNA methyltransferase and reactivates methylation-silenced genes in cancer cell lines. Cancer Res. 2003, 63, 7563–7570. [Google Scholar] [PubMed]
- Van Aller, G.S.; Carson, J.D.; Tang, W.; Peng, H.; Zhao, L.; Copeland, R.A.; Tummino, P.J.; Luo, L. Epigallocatechin gallate (EGCG), a major component of green tea, is a dual phosphoinositide-3-kinase/mTOR inhibitor. Biochem. Biophys. Res. Commun. 2011, 406, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Kiss, A.; Becsi, B.; Kolozsvari, B.; Komaromi, I.; Kover, K.E.; Erdodi, F. Epigallocatechin-3-gallate and penta-O-galloyl-beta-D-glucose inhibit protein phosphatase-1. FEBS J. 2013, 280, 612–626. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ren, X.; Deng, C.; Yang, L.; Yan, E.; Guo, T.; Li, Y.; Xu, M.X. Mechanism of the inhibition of the STAT3 signaling pathway by EGCG. Oncol. Rep. 2013, 30, 2691–2696. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.P.; Huang, C.; Cui, Q.Y.; Yang, D.J.; Sun, K.; Chen, X.; Li, X.H. Meta-Analysis of the Association between Tea Intake and the Risk of Cognitive Disorders. PLoS ONE 2016, 11, e0165861. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, K.; Watanabe, Y.; Nakamura, K.; Sanpei, K.; Wakasugi, M.; Yokoseki, A.; Onodera, O.; Ikeuchi, T.; Kuwano, R.; Momotsu, T.; et al. Modifiable Factors Associated with Cognitive Impairment in 1, 143 Japanese Outpatients: The Project in Sado for Total Health (PROST). Dement. Geriatr. Cogn. Dis. Extra 2016, 6, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Unno, K.; Ishikawa, Y.; Takabayashi, F.; Sasaki, T.; Takamori, N.; Iguchi, K.; Hoshino, M. Daily ingestion of green tea catechins from adulthood suppressed brain dysfunction in aged mice. Biofactors 2008, 34, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Khalid, S.; Paul, S. Identifying a C-terminal ATP binding sites-based novel Hsp90-Inhibitor in silico: A plausible therapeutic approach in Alzheimer’s disease. Med. Hypotheses 2014, 83, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Ehrnhoefer, D.E.; Bieschke, J.; Boeddrich, A.; Herbst, M.; Masino, L.; Lurz, R.; Engemann, S.; Pastore, A.; Wanker, E.E. EGCG redirects amyloidogenic polypeptides into unstructured, off-pathway oligomers. Nat. Struct. Mol. Biol. 2008, 15, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Chebaro, Y.; Jiang, P.; Zang, T.; Mu, Y.; Nguyen, P.H.; Mousseau, N.; Derreumaux, P. Structures of Abeta17-42 trimers in isolation and with five small-molecule drugs using a hierarchical computational procedure. J. Phys. Chem. B 2012, 116, 8412–8422. [Google Scholar] [CrossRef] [PubMed]
- Lemkul, J.A.; Bevan, D.R. The role of molecular simulations in the development of inhibitors of amyloid beta-peptide aggregation for the treatment of Alzheimer’s disease. ACS Chem. Neurosci. 2012, 3, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Hyung, S.J.; DeToma, A.S.; Brender, J.R.; Lee, S.; Vivekanandan, S.; Kochi, A.; Choi, J.S.; Ramamoorthy, A.; Ruotolo, B.T.; Lim, M.H. Insights into antiamyloidogenic properties of the green tea extract (−)-epigallocatechin-3-gallate toward metal-associated amyloid-beta species. Proc. Natl. Acad. Sci. USA 2013, 110, 3743–3748. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Jamal, Q.M.; Shams, S.; Al-Wabel, N.A.; Siddiqui, M.U.; Alzohairy, M.A.; Al Karaawi, M.A.; Kesari, K.K.; Mushtaq, G.; Kamal, M.A. In Silico Analysis of Green Tea Polyphenols as Inhibitors of AChE and BChE Enzymes in Alzheimer’s Disease Treatment. CNS Neurol. Disord. Drug Targets 2016, 15, 624–628. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.X.; Li, G.Z.; Zhang, B.; Xia, Z.Y.; Zhang, M. Molecular evaluation of herbal compounds as potent inhibitors of acetylcholinesterase for the treatment of Alzheimer’s disease. Mol. Med. Rep. 2016, 14, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Azam, F.; Mohamed, N.; Alhussen, F. Molecular interaction studies of green tea catechins as multitarget drug candidates for the treatment of Parkinson’s disease: Computational and structural insights. Network 2015, 26, 97–115. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; He, J.; Chang, A.K.; Wang, Y.; Xu, L.; Chong, X.; Lu, X.; Sun, Y.; Xia, X.; Li, H.; et al. (−)-epigallocatechin-3-gallate inhibits fibrillogenesis of chicken cystatin. J. Agric. Food Chem. 2015, 63, 1347–1351. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, E.; Rajasekaran, R. Probing the inhibitory activity of epigallocatechin-gallate on toxic aggregates of mutant (L84F) SOD1 protein through geometry-based sampling and steered molecular dynamics. J. Mol. Graph. Model. 2017, 74, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Broski, S.M.; Spinner, R.J.; Howe, B.M.; Dispenzieri, A.; Johnson, G.B. 18F-Florbetapir and 18F-FDG PET/CT in Systemic Immunoglobulin Light Chain Amyloidosis Involving the Peripheral Nerves. Clin. Nucl. Med. 2016, 41, e115–e117. [Google Scholar] [CrossRef] [PubMed]
- Hora, M.; Carballo-Pacheco, M.; Weber, B.; Morris, V.K.; Wittkopf, A.; Buchner, J.; Strodel, B.; Reif, B. Epigallocatechin-3-gallate preferentially induces aggregation of amyloidogenic immunoglobulin light chains. Sci. Rep. 2017, 7, 41515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyganowska-Swiatkowska, M.; Matthews-Kozanecka, M.; Matthews-Brzozowska, T.; Skrzypczak-Jankun, E.; Jankun, J. Can EGCG Alleviate Symptoms of Down Syndrome by Altering Proteolytic Activity? Int. J. Mol. Sci. 2018, 19, 248. [Google Scholar] [CrossRef] [PubMed]
- Nagao, T.; Meguro, S.; Hase, T.; Otsuka, K.; Komikado, M.; Tokimitsu, I.; Yamamoto, T.; Yamamoto, K. A catechin-rich beverage improves obesity and blood glucose control in patients with type 2 diabetes. Obesity 2009, 17, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Rashid, A.M.; Lu, K.; Yip, Y.M.; Zhang, D. Averrhoa carambola L. peel extract suppresses adipocyte differentiation in 3T3-L1 cells. Food Funct. 2016, 7, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Jhong, C.H.; Riyaphan, J.; Lin, S.H.; Chia, Y.C.; Weng, C.F. Screening alpha-glucosidase and alpha-amylase inhibitors from natural compounds by molecular docking in silico. Biofactors 2015, 41, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Ulloa, A.; Mendez-Luna, D.; Beltran-Partida, E.; Castillo, C.; Guevara, G.; Ramirez-Sanchez, I.; Correa-Basurto, J.; Ceballos, G.; Villarreal, F. The effects of (−)-epicatechin on endothelial cells involve the G protein-coupled estrogen receptor (GPER). Pharmacol. Res. 2015, 100, 309–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, A.E.; Piegholdt, S.; Rabe, D.; Baenas, N.; Schloesser, A.; Eggersdorfer, M.; Stocker, A.; Rimbach, G. Epigallocatechin gallate affects glucose metabolism and increases fitness and lifespan in Drosophila melanogaster. Oncotarget 2015, 6, 30568–30578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brewer, R.A.; Gibbs, V.K.; Smith, D.L., Jr. Targeting glucose metabolism for healthy aging. Nutr. Healthy Aging 2016, 4, 31–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Gidley, M.J.; Warren, F.J. The mechanism of interactions between tea polyphenols and porcine pancreatic alpha-amylase: Analysis by inhibition kinetics, fluorescence quenching, differential scanning calorimetry and isothermal titration calorimetry. Mol. Nutr. Food Res. 2017, 61, 1700324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, B.; Watson, K.A.; McGuffin, L.J.; Lovegrove, J.A.; Gibbins, J.M. GRID and docking analyses reveal a molecular basis for flavonoid inhibition of Src family kinase activity. J. Nutr. Biochem. 2015, 26, 1156–1165. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Deng, J.; Li, W.; Lin, D.; Su, C.; Wang, M.; Li, X.; Abuaku, B.K.; Tan, H.; Wen, S.W. Impact of tea drinking upon tuberculosis: A neglected issue. BMC Public Health 2015, 15, 515. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P. Impact of green tea catechins on multidrug-resistant bacterial pathogens. In Health Benefits of Green Tea: An Evidence-Based Approach; Hara, Y., Yang, C.S., Isemura, M., Tomita, I., Eds.; CABI: Oxford, UK, 2016; pp. 147–156. ISBN 9781786392398. [Google Scholar]
- Sameshima, Y.; Ishida, Y.; Ono, Y.; Hujita, M.; Kuriki, Y. Green Tea Powder Enhances the Safety and Efficacy of Interferon α-2b Plus Ribavirin Combination Therapy in Chronic Hepatitis C Patients with a Very High Genotype 1 HCV Load; Isemura, M., Ed.; Research Signpost: Trivandrum, India, 2008; pp. 113–119. [Google Scholar]
- Song, J.M. Anti-infective potential of catechins and their derivatives against viral hepatitis. Clin. Exp. Vaccine Res. 2018, 7, 37–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.K.; Kumar, G.; Kapoor, M.; Surolia, A. Combined effect of epigallocatechin gallate and triclosan on enoyl-ACP reductase of Mycobacterium tuberculosis. Biochem. Biophys. Res. Commun. 2008, 368, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Aanandhi, M.V.; Bhattacherjee, D.; George, P.S.; Ray, A. Natural polyphenols down-regulate universal stress protein in Mycobacterium tuberculosis: An in-silico approach. J. Adv. Pharm. Technol. Res. 2014, 5, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Gradisar, H.; Pristovsek, P.; Plaper, A.; Jerala, R. Green tea catechins inhibit bacterial DNA gyrase by interaction with its ATP binding site. J. Med. Chem. 2007, 50, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Benedik, E.; Skrt, M.; Podlipnik, C.; Ulrih, N.P. Binding of flavonoids to staphylococcal enterotoxin B. Food Chem. Toxicol. 2014, 74, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shimamura, Y.; Utsumi, M.; Hirai, C.; Nakano, S.; Ito, S.; Tsuji, A.; Ishii, T.; Hosoya, T.; Kan, T.; Ohashi, N.; et al. Binding of Catechins to Staphylococcal Enterotoxin A. Molecules 2018, 23, 1125. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, M.; Shimatani, K.; Ozawa, T.; Shigemune, N.; Tomiyama, D.; Yui, K.; Katsuki, M.; Ikeda, K.; Nonaka, A.; Miyamoto, T. Mechanism for the antibacterial action of epigallocatechin gallate (EGCg) on Bacillus subtilis. Biosci. Biotechnol. Biochem. 2015, 79, 845–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.; Teng, Z.; Li, M.; Niu, X.; Wang, J.; Deng, X. Epigallocatechin gallate inhibits Streptococcus pneumoniae virulence by simultaneously targeting pneumolysin and sortase A. J. Cell. Mol. Med. 2017, 21, 2586–2598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamza, A.; Zhan, C.G. How can (−)-epigallocatechin gallate from green tea prevent HIV-1 infection? Mechanistic insights from computational modeling and the implication for rational design of anti-HIV-1 entry inhibitors. J. Phys. Chem. B 2006, 110, 2910–2917. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Woo, H.J.; Kang, H.K.; Nguyen, V.D.; Kim, Y.M.; Kim, D.W.; Ahn, S.A.; Xia, Y.; Kim, D. Flavonoid-mediated inhibition of SARS coronavirus 3C-like protease expressed in Pichia pastoris. Biotechnol. Lett. 2012, 34, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Derksen, A.; Hensel, A.; Hafezi, W.; Herrmann, F.; Schmidt, T.J.; Ehrhardt, C.; Ludwig, S.; Kuhn, J. 3-O-galloylated procyanidins from Rumex acetosa L. inhibit the attachment of influenza A virus. PLoS ONE 2014, 9, e110089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, J.; Kumari, A.; Somvanshi, P.; Grover, A.; Pai, S.; Sunil, S. In silico analysis of natural compounds targeting structural and nonstructural proteins of chikungunya virus. F1000Research 2017, 6, 1601. [Google Scholar] [CrossRef] [PubMed]
- Fatima, K.; Mathew, S.; Suhail, M.; Ali, A.; Damanhouri, G.; Azhar, E.; Qadri, I. Docking studies of Pakistani HCV NS3 helicase: A possible antiviral drug target. PLoS ONE 2014, 9, e106339. [Google Scholar] [CrossRef] [PubMed]
- Cherubin, P.; Garcia, M.C.; Curtis, D.; Britt, C.B.; Craft, J.W., Jr.; Burress, H.; Berndt, C.; Reddy, S.; Guyette, J.; Zheng, T.; et al. Inhibition of Cholera Toxin and Other AB Toxins by Polyphenolic Compounds. PLoS ONE 2016, 11, e0166477. [Google Scholar] [CrossRef] [PubMed]
- Ismail, N.A.; Jusoh, S.A. Molecular Docking and Molecular Dynamics Simulation Studies to Predict Flavonoid Binding on the Surface of DENV2 E Protein. Interdiscip. Sci. 2017, 9, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Jena, L.; Galande, S.; Daf, S.; Mohod, K.; Varma, A.K. Elucidating Molecular Interactions of Natural Inhibitors with HPV-16 E6 Oncoprotein through Docking Analysis. Genomics Inform. 2014, 12, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Masuda, S.; Maeda-Yamamoto, M.; Usui, S.; Fujisawa, T. ‘Benifuuki’ green tea containing o-methylated catechin reduces symptoms of Japanese cedar pollinosis: A randomized, double-blind, placebo-controlled trial. Allergol. Int. 2014, 63, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Leichsenring, A.; Backer, I.; Furtmuller, P.G.; Obinger, C.; Lange, F.; Flemmig, J. Long-Term Effects of (−)-Epigallocatechin Gallate (EGCG) on Pristane-Induced Arthritis (PIA) in Female Dark Agouti Rats. PLoS ONE 2016, 11, e0152518. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Umar, S.; Riegsecker, S.; Chourasia, M.; Ahmed, S. Regulation of Transforming Growth Factor beta-Activated Kinase Activation by Epigallocatechin-3-Gallate in Rheumatoid Arthritis Synovial Fibroblasts: Suppression of K(63) -Linked Autoubiquitination of Tumor Necrosis Factor Receptor-Associated Factor 6. Arthritis Rheumatol. 2016, 68, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Fechtner, S.; Singh, A.; Chourasia, M.; Ahmed, S. Molecular insights into the differences in anti-inflammatory activities of green tea catechins on IL-1beta signaling in rheumatoid arthritis synovial fibroblasts. Toxicol. Appl. Pharmacol. 2017, 329, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Ognjenovic, J.; Stojadinovic, M.; Milcic, M.; Apostolovic, D.; Vesic, J.; Stambolic, I.; Atanaskovic-Markovic, M.; Simonovic, M.; Velickovic, T.C. Interactions of epigallo-catechin 3-gallate and ovalbumin, the major allergen of egg white. Food Chem. 2014, 164, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Vesic, J.; Stambolic, I.; Apostolovic, D.; Milcic, M.; Stanic-Vucinic, D.; Cirkovic Velickovic, T. Complexes of green tea polyphenol, epigalocatechin-3-gallate, and 2S albumins of peanut. Food Chem. 2015, 185, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Li, R.; Li, N.; Zhang, J.; Li, Y.; Gong, P.; Gao, D.; Liu, H.; Zhang, Y. Anti-inflammatory effect of procyanidin B1 on LPS-treated THP1 cells via interaction with the TLR4-MD-2 heterodimer and p38 MAPK and NF-kappaB signaling. Mol. Cell. Biochem. 2015, 407, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Shraberg, J.; Rick, S.W.; Rannulu, N.; Cole, R.B. A study of procyanidin binding to Histatin 5 using Electrospray Ionization Tandem Mass Spectrometry (ESI-MS/MS) and molecular simulations. Phys. Chem. Chem. Phys. 2015, 17, 12247–12258. [Google Scholar] [CrossRef] [PubMed]
- Nokthai, P.; Lee, V.S.; Shank, L. Molecular modeling of peroxidase and polyphenol oxidase: Substrate specificity and active site comparison. Int. J. Mol. Sci. 2010, 11, 3266–3276. [Google Scholar] [CrossRef] [PubMed]
- Fukuhara, N.; Kawabata, T. HOMCOS: A server to predict interacting protein pairs and interacting sites by homology modeling of complex structures. Nucleic Acids Res 2008, 36, W185–W189. [Google Scholar] [CrossRef] [PubMed]
- Hurlburt, B.K.; Offermann, L.R.; McBride, J.K.; Majorek, K.A.; Maleki, S.J.; Chruszcz, M. Structure and function of the peanut panallergen Ara h 8. J. Biol. Chem. 2013, 288, 36890–36901. [Google Scholar] [CrossRef] [PubMed]
- Mauge, C.; Granier, T.; d’Estaintot, B.L.; Gargouri, M.; Manigand, C.; Schmitter, J.M.; Chaudiere, J.; Gallois, B. Crystal structure and catalytic mechanism of leucoanthocyanidin reductase from Vitis vinifera. J. Mol. Biol. 2010, 397, 1079–1091. [Google Scholar] [CrossRef] [PubMed]
- Casanal, A.; Zander, U.; Munoz, C.; Dupeux, F.; Luque, I.; Botella, M.A.; Schwab, W.; Valpuesta, V.; Marquez, J.A. The strawberry pathogenesis-related 10 (PR-10) Fra a proteins control flavonoid biosynthesis by binding to metabolic intermediates. J. Biol. Chem. 2013, 288, 35322–35332. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, M.; Chen, P.; Narayan, S.; Matschinsky, F.M.; Bennett, M.J.; Stanley, C.A.; Smith, T.J. Green tea polyphenols control dysregulated glutamate dehydrogenase in transgenic mice by hijacking the ADP activation site. J. Biol. Chem. 2011, 286, 34164–34174. [Google Scholar] [CrossRef] [PubMed]
- Skrzypczak-Jankun, E.; Zhou, K.; Jankun, J. Inhibition of lipoxygenase by (−)-epigallocatechin gallate: X-ray analysis at 2.1 A reveals degradation of EGCG and shows soybean LOX-3 complex with EGC instead. Int. J. Mol. Med. 2003, 12, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Robertson, I.M.; Li, M.X.; Sykes, B.D. Solution structure of human cardiac troponin C in complex with the green tea polyphenol, (−)-epigallocatechin 3-gallate. J. Biol. Chem. 2009, 284, 23012–23023. [Google Scholar] [CrossRef] [PubMed]
- Miyata, M.; Sato, T.; Kugimiya, M.; Sho, M.; Nakamura, T.; Ikemizu, S.; Chirifu, M.; Mizuguchi, M.; Nabeshima, Y.; Suwa, Y.; et al. The crystal structure of the green tea polyphenol (−)-epigallocatechin gallate-transthyretin complex reveals a novel binding site distinct from the thyroxine binding site. Biochemistry 2010, 49, 6104–6114. [Google Scholar] [CrossRef] [PubMed]
- Urusova, D.V.; Shim, J.H.; Kim, D.J.; Jung, S.K.; Zykova, T.A.; Carper, A.; Bode, A.M.; Dong, Z. Epigallocatechin-gallate suppresses tumorigenesis by directly targeting Pin1. Cancer Prev. Res. 2011, 4, 1366–1377. [Google Scholar] [CrossRef] [PubMed]
- Kowalinski, E.; Zubieta, C.; Wolkerstorfer, A.; Szolar, O.H.; Ruigrok, R.W.; Cusack, S. Structural analysis of specific metal chelating inhibitor binding to the endonuclease domain of influenza pH1N1 (2009) polymerase. PLoS Pathog. 2012, 8, e1002831. [Google Scholar] [CrossRef] [PubMed]












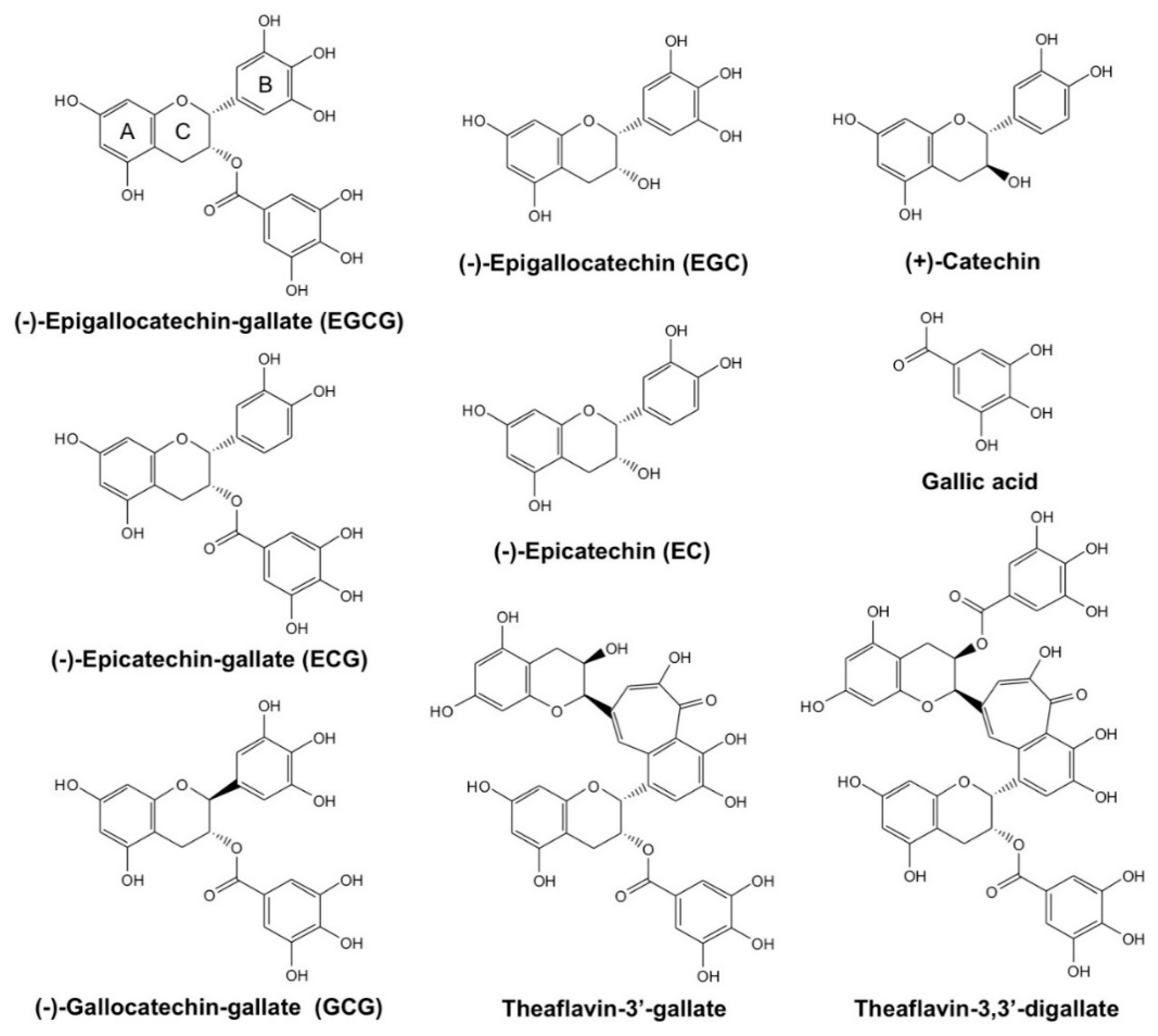{kind=link}
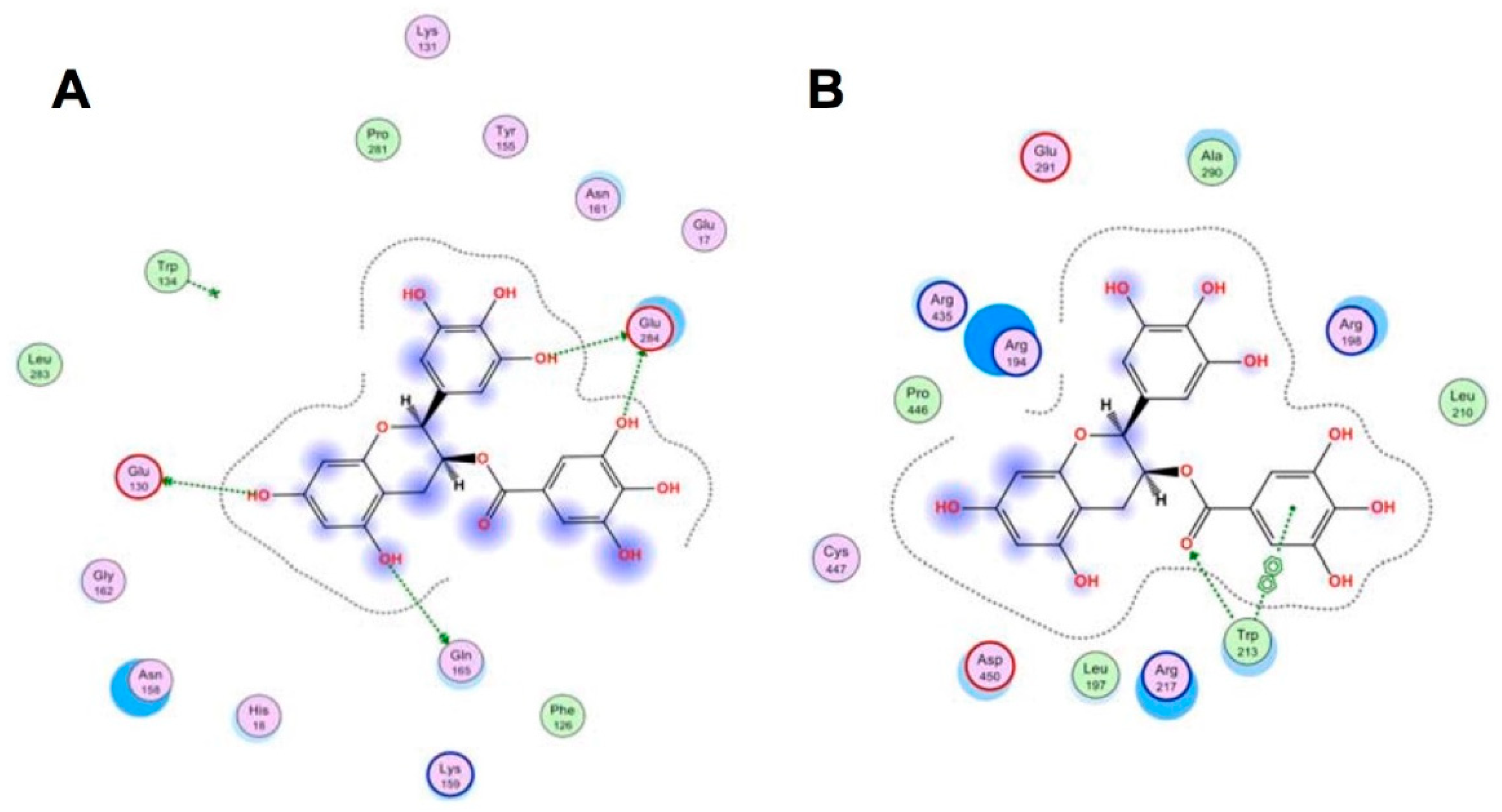{kind=link}
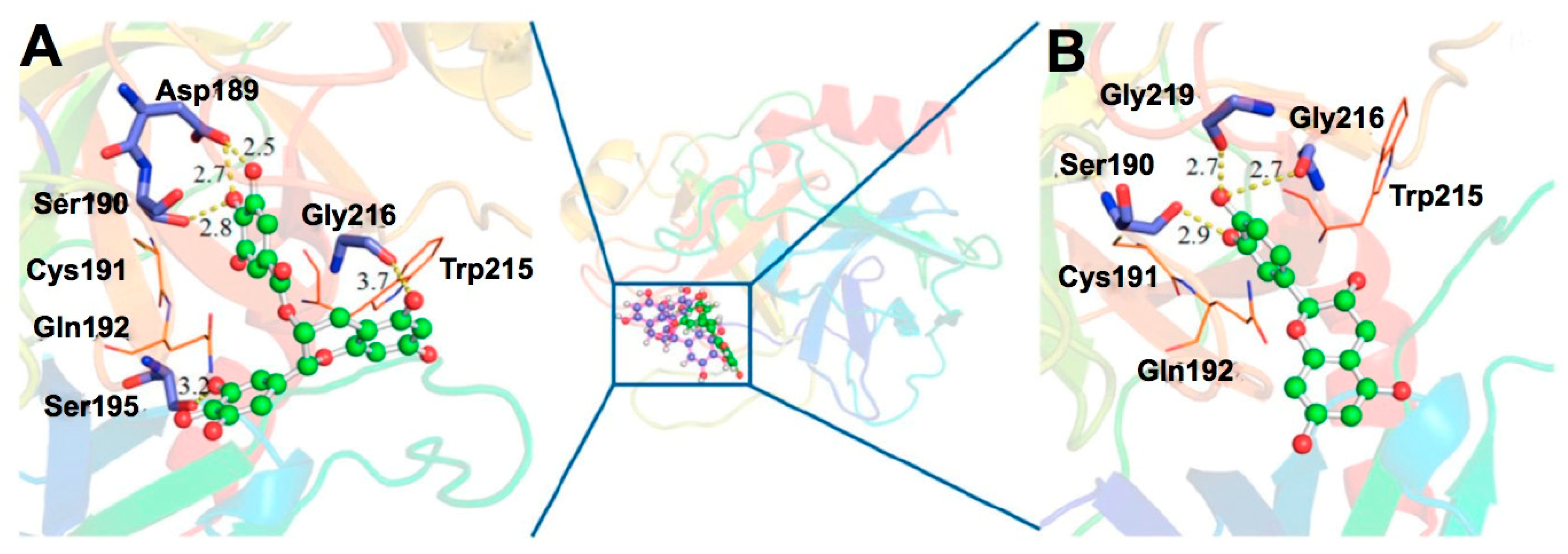{kind=link}
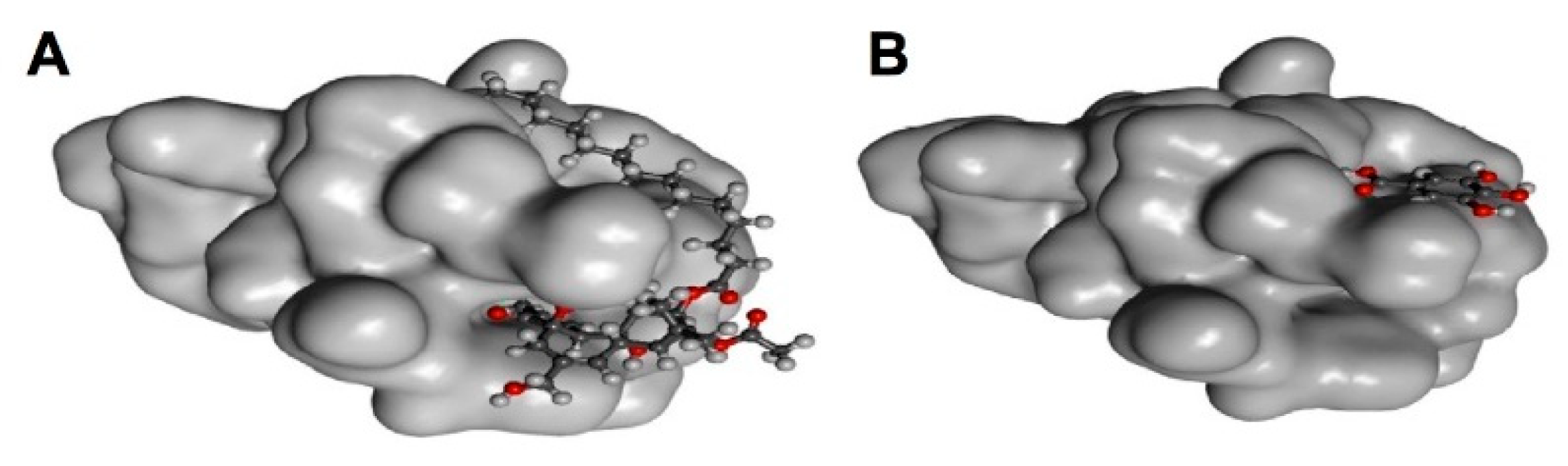{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PDB ID | Protein | No. of Sites | Ligand | Resolution | Binding Characteristics Defined by Ligand Explorer (4.2.0) | Head Author (Year) | Reference | |||
|---|---|---|---|---|---|---|---|---|---|---|
| No. of Hydrogen Bonds | Contact Residues and Molecules * | No. of Hydrophobic Contacts | Contact Residues and Molecules | |||||||
| 4MA6 | Ara h 8 | 1 | EC | 2.0 | 7 | T30(s), Y82(s), 4 waters | 6 | L, L, I, T, Y, H, K | Hurlburt, B. K. (2013) | [110] |
| 3I52 | Leucoanthocyanidin reductase | 1 | (+)-catechin | 2.3 | 11 | H122(m), Y137(s), I171(m), G93(m), E94(s), 5 waters | 6 | M, G, G, F, H | Mauge, C. (2010) | [111] |
| 4C9I | PR-10 Fra a 1E | 1 | (+)-catechin | 3.1 | 1 **** | D154(s) | 6 | L, G, H, K, D | Casanal, A. (2013) | [112] |
| 4C94 | PR-10 Fra a 3 | 1 | (+)-catechin | 3.0 | 5 | H70(s), 4 waters | 4 | L, R | Casanal, A. (2013) | [112] |
| 3QMU | Glutamate dehydrogenase | 1 | EGC | 3.6 | 4 **** | V120(m), R396(s), R459(s), R491(s) | 6 | V, A, H, K, D | Li. C. (2011) | [5,113] |
| 1JNQ | Lipoxygenase-3 | 1 | EGC (EGCG) ** | 2.1 | 4 | Q716(s), 2 waters | 18 | V, I, L, L, H, H, F, W, Q, Fe | Skrzypczak-Jankun, E. (2003) | [114] |
| 2KDH | Troponin C | 1 | EGCG | n. d *** | 0 **** | - | 7 | M, V, L, F, F | Robertson, I.M. (2009) | [115] |
| 3NG5 | Transthyretin mutant V30M | 2 | EGCG | 1.7 | 5 | D18(s), V20(m), 5 waters | 7 | V, L, I, R | Miyata, M. (2010) | [5,116] |
| 7 | V32(m), W41(m), S46(s), E72(s), 3 waters | 10 | P, F, H, E | |||||||
| 3OOB | Pin1 | 2 | EGCG | 1.9 | 4 | 2 waters, R17(m,s) | 7 | R, Ligand | Urusova, D.V. (2011) | [5,117] |
| 5 | Water R60(s), W73(m), D112(m), S114(m) | 2 | S, W | |||||||
| 4AWM | PA endonuclease | 1 | EGCG | 2.6 | 12 | E26(s), E80(s), D108(s), E119(s), I120(m), V122(m), K134(s), Y130(s), water | 4 | A, Y, H, I | Kowalinski, E. (2012) | [118] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakano, S.; Megro, S.-i.; Hase, T.; Suzuki, T.; Isemura, M.; Nakamura, Y.; Ito, S. Computational Molecular Docking and X-ray Crystallographic Studies of Catechins in New Drug Design Strategies. Molecules 2018, 23, 2020. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23082020
Nakano S, Megro S-i, Hase T, Suzuki T, Isemura M, Nakamura Y, Ito S. Computational Molecular Docking and X-ray Crystallographic Studies of Catechins in New Drug Design Strategies. Molecules. 2018; 23(8):2020. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23082020
Chicago/Turabian StyleNakano, Shogo, Shin-ichi Megro, Tadashi Hase, Takuji Suzuki, Mamoru Isemura, Yoriyuki Nakamura, and Sohei Ito. 2018. "Computational Molecular Docking and X-ray Crystallographic Studies of Catechins in New Drug Design Strategies" Molecules 23, no. 8: 2020. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23082020