Polyphenols from Acorn Leaves (Quercus liaotungensis) Protect Pancreatic Beta Cells and Their Inhibitory Activity against α-Glucosidase and Protein Tyrosine Phosphatase 1B

Abstract
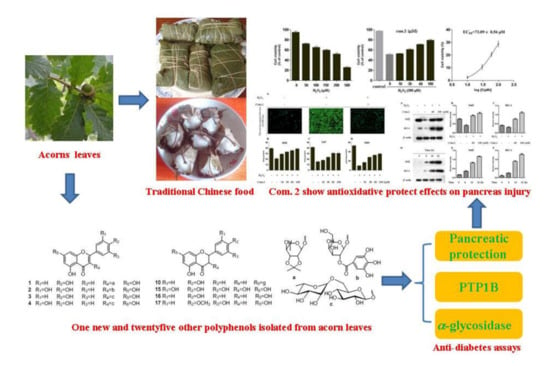
:
1. Introduction
2. Results and Discussion
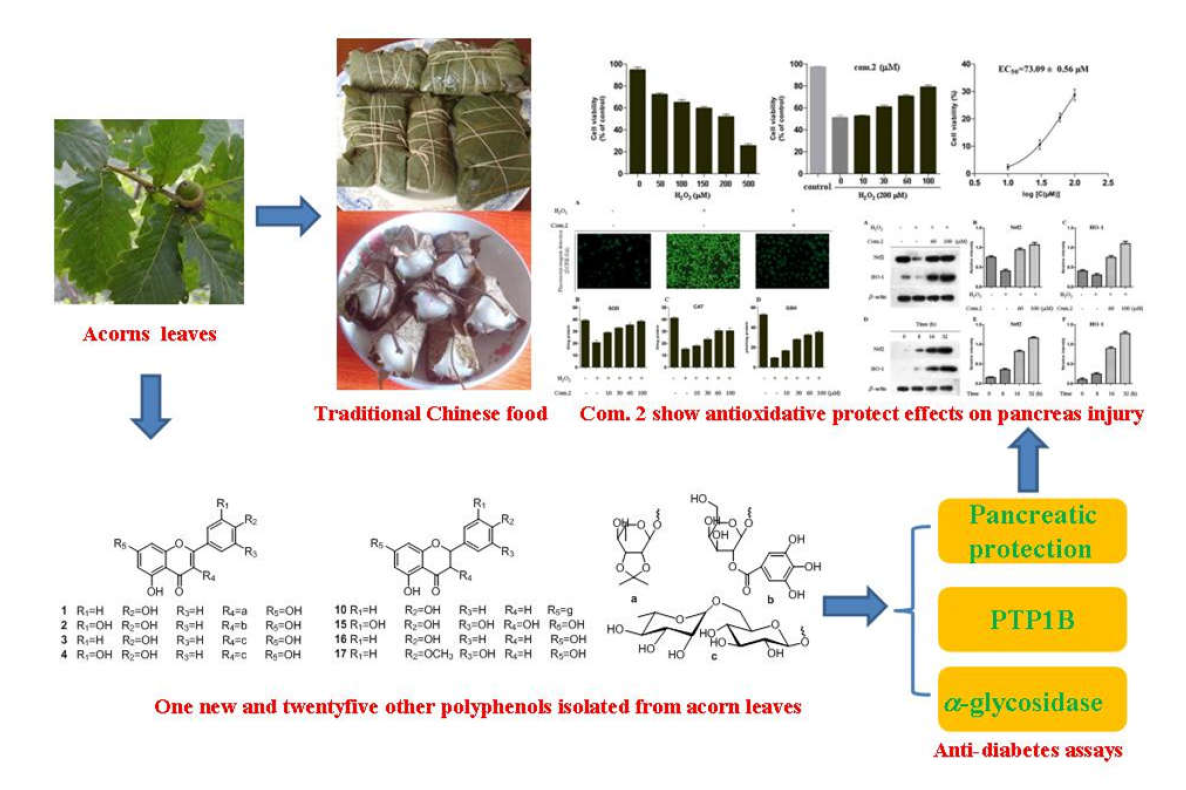
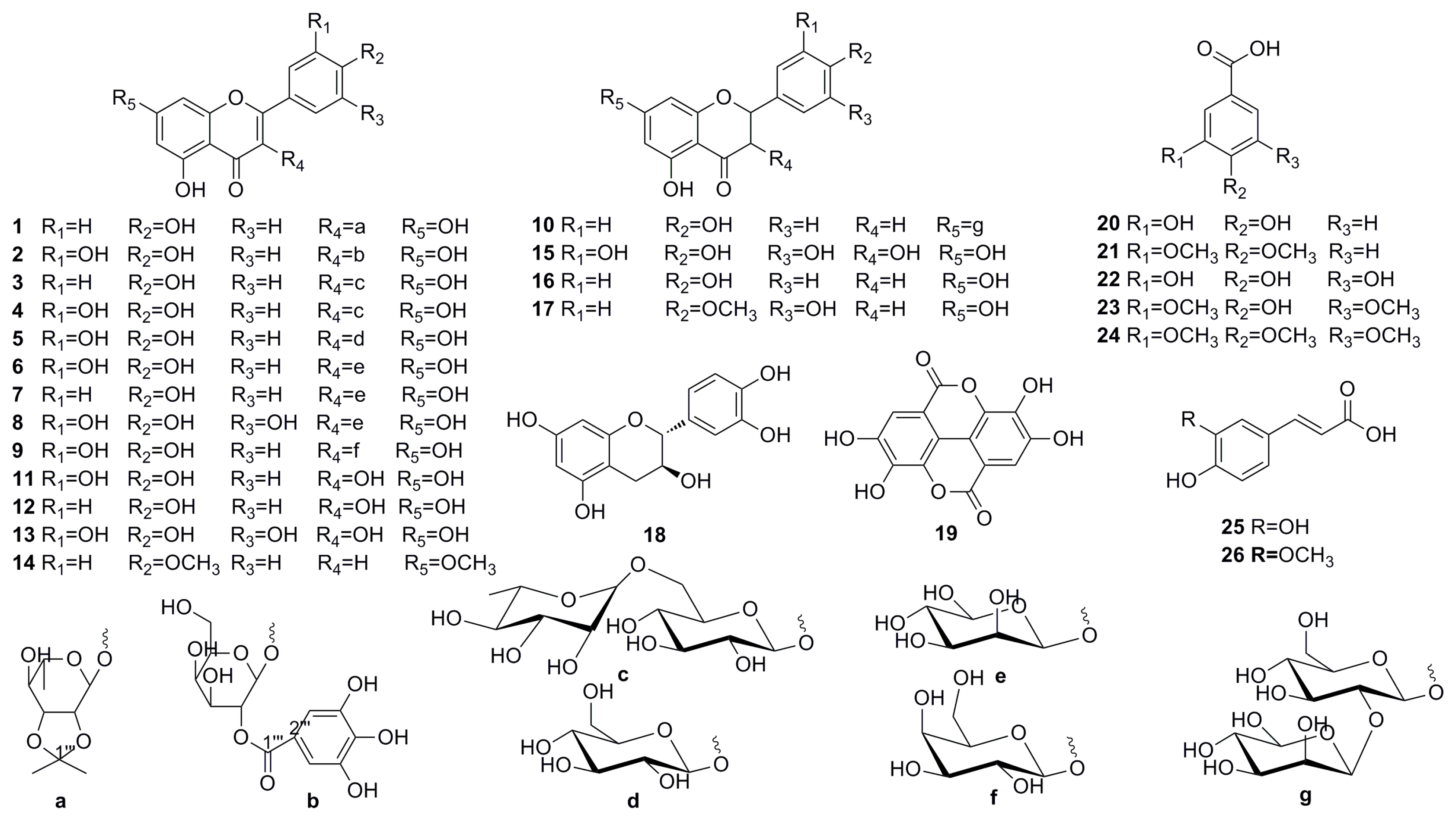
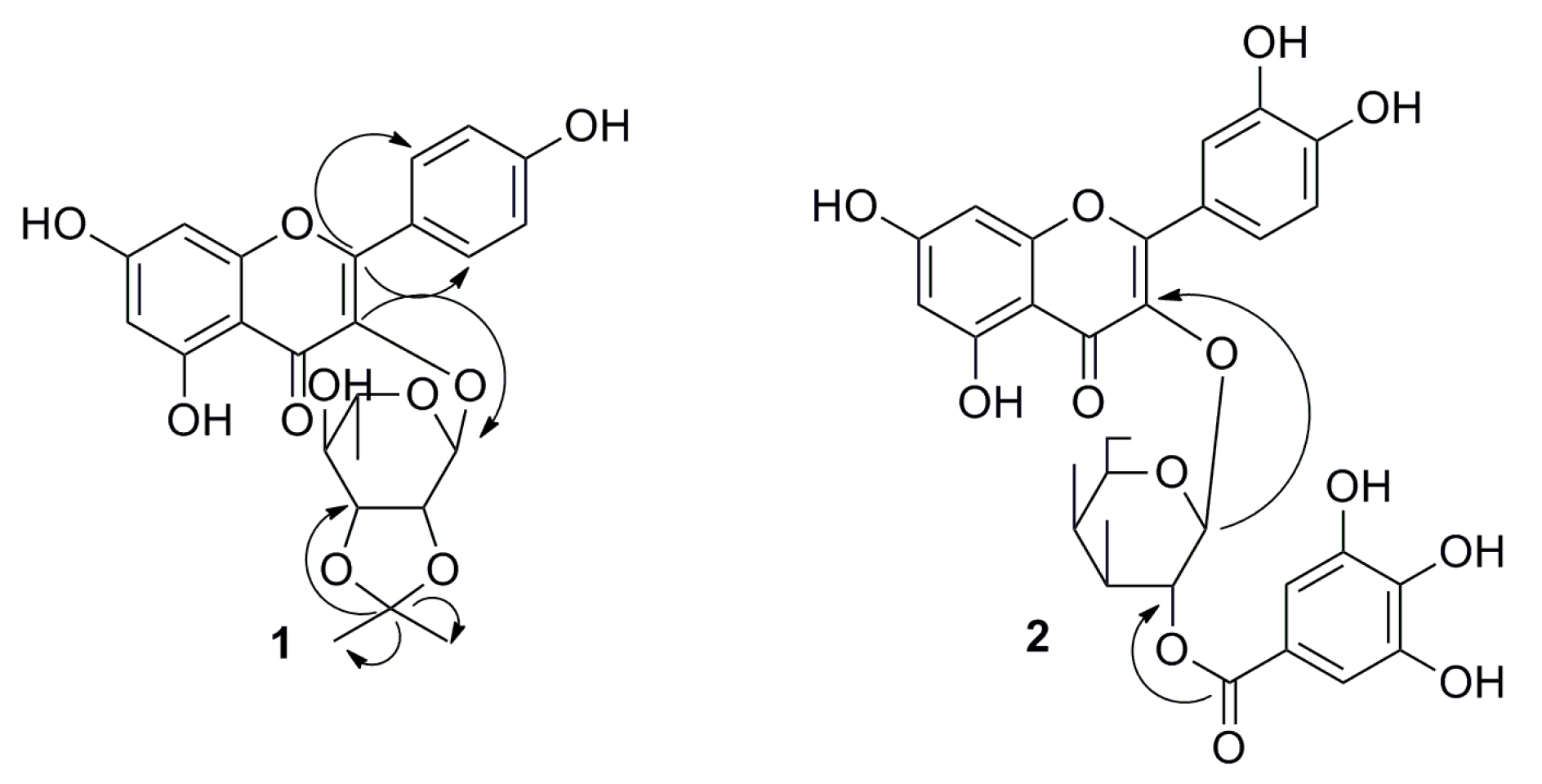
2.1. Phytochemical Studies of Acorn Leaves
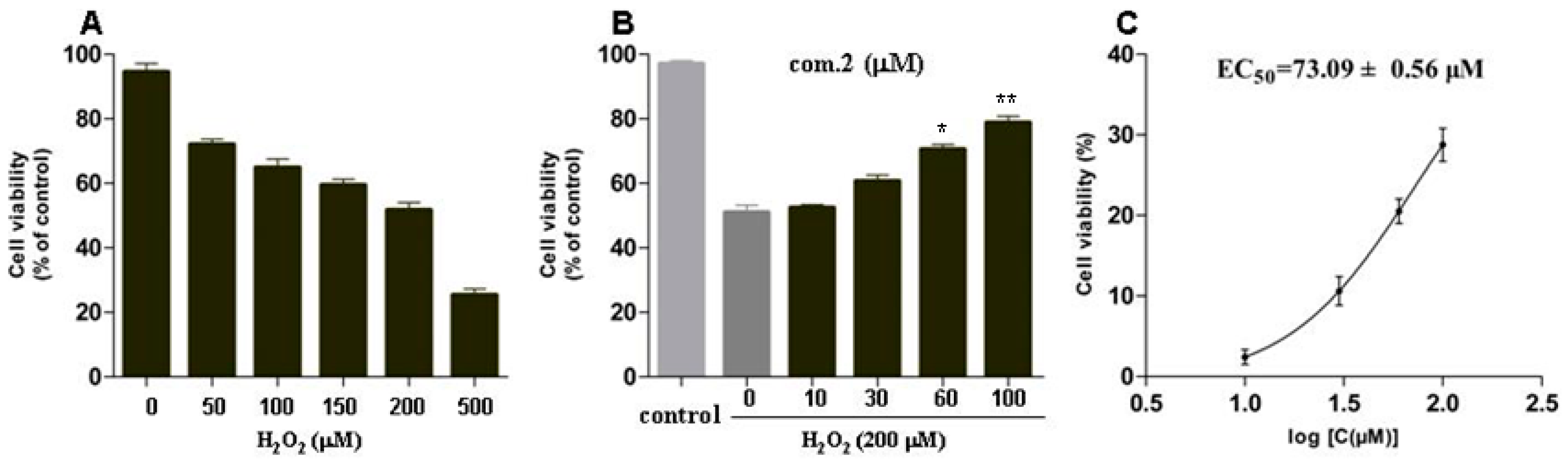
2.2. Protective Effects on Pancreatic Beta Cells (MIN6) Damaged by H2O2
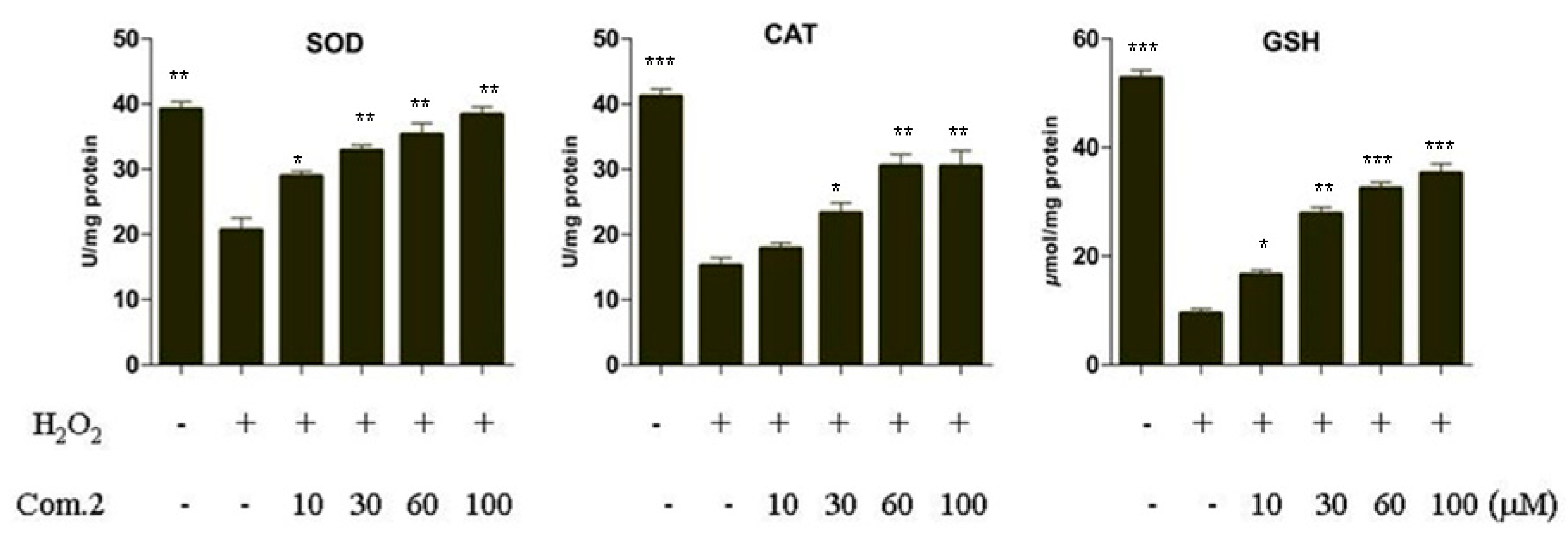
2.3. Compound 2 Enhanced the Activities of SOD, CAT, and GSH in Pancreatic Beta Cells (MIN6) Damaged by H2O2
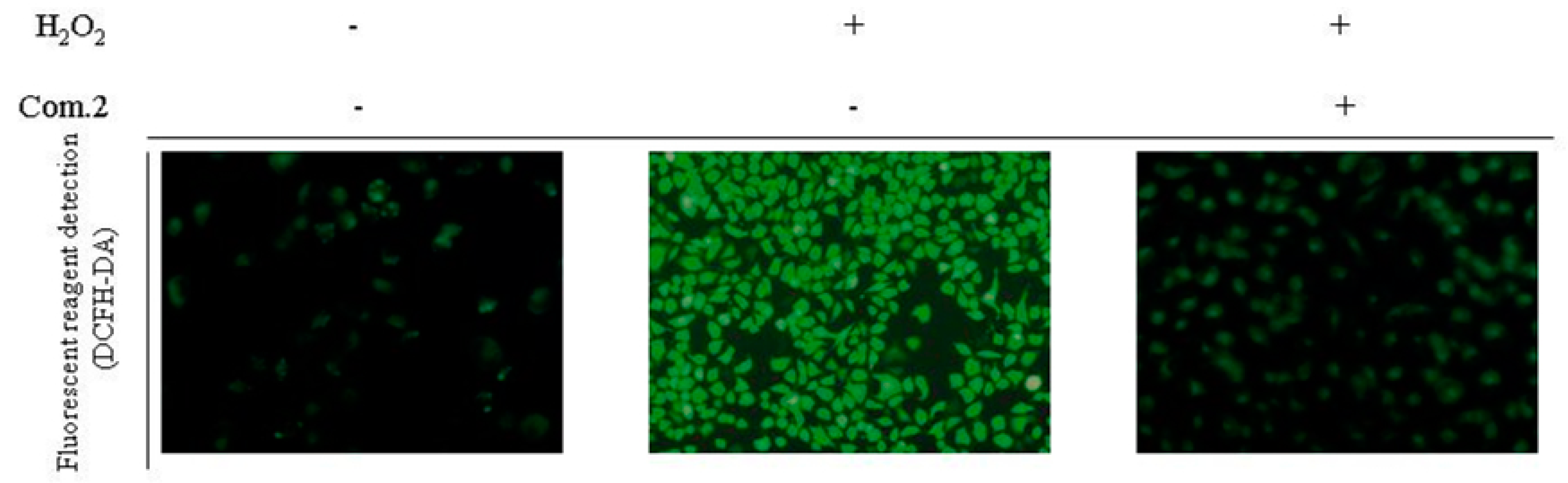
2.4. Compound 2 Reduced the Amount of Intracellular ROS
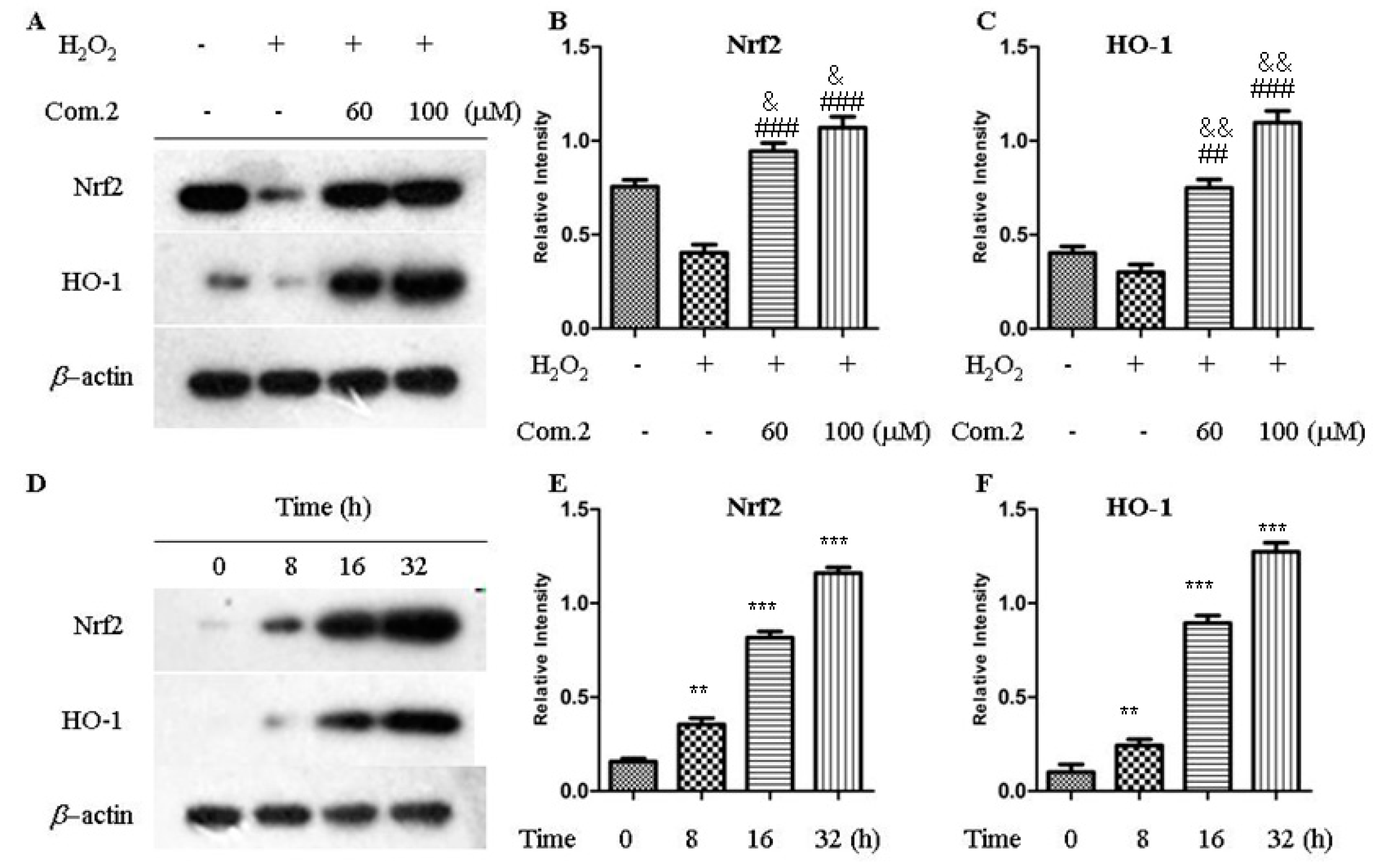
2.5. Compound 2 Activated Nrf2/HO-1 Pathway
2.6. PTP1B and α-Glucosidase Inhibition Assay
3. Materials and Methods
3.1. Reagents and Chemicals
3.2. Plant Material
3.3. Extraction and Isolation
3.4. Cell Culture and Cell Viability Assay
3.5. Determining CAT, SOD, and GSH
3.6. Determining the Amount of Intracellular ROS
3.7. Western Blot Analysis
3.8. Assay for PTP1B Inhibitory Activity
3.9. Assay for α-Glycosidase Inhibitory Activity
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Robertson, R.P. Oxidative stress and impaired insulin secretion in type 2 diabetes. Curr. Opin. Pharmacol. 2006, 6, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Dinkova-Kostova, A.T.; Holtzclaw, W.D.; Cole, R.N.; Itoh, K.; Wakabayashi, N.; Katoh, Y.; Yamamoto, M.; Talalay, P. Direct evidence that sulfhydryl groups of Keap1 are the sensors regulating induction of phase 2 enzymes that protect against carcinogens and oxidants. Proc. Natl. Acad. Sci. USA 2002, 99, 11908–11913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, L.; Johnson, J.A. Oxidative damage and the Nrf2-ARE pathway in neurodegenerative disease. Biochim. Biophys. Acta 2014, 1842, 1208–1218. [Google Scholar] [CrossRef] [PubMed]
- Kovac, S.; Angelova, P.R.; Holmström, K.M.; Zhang, Y.; Dinkova-Kostova, A.T.; Abramov, A.Y. Nrf2 regulates ROS production by mitochondria and NADPH oxidase. Biochim. Biophys. Acta 2015, 1850, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Krentz, A.J.; Bailey, C.J. Oral antidiabetic agents: Current oral in type 2 diabetes mellitus. Drugs 2005, 65, 385–411. [Google Scholar] [CrossRef] [PubMed]
- Krayt-Cohen, J.; Muller, W.J.; Elson, A.P. Protein-tyrosine phosphatase epsilon regulates signaling in a kinase-specific manner: Increasing coherence in tyrosine phosphatase signaling. J. Biol. Chem. 2008, 283, 4612–4621. [Google Scholar] [CrossRef] [PubMed]
- Trinh, B.T.; Staerk, D.; Jäger, A.K. Screening for potential α-glucosidase and α-amylase inhibitory constituents from selected Vietnamese plants used to treat type 2 diabetes. J. Ethnopharmacol. 2016, 186, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Mendes, N.F.; Castro, G.; Guadagnini, D.; Tobar, N.; Cognuck, S.Q.; Elias, L.L.K.; Boer, P.A.; Prada, P.O. Knocking down amygdalar PTP1B in diet-induced obese rats improves insulin signaling/action, decreases adiposity and may alter anxiety behavior. Metab. Clin. Exp. 2017, 70, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jaroslaw, K.; Mariusz, W.; Rafal, Z.; Leslaw, J. The influence of acorn flour on rheological properties of gluten–free dough and physical characteristics of the bread. Eur. Food Res. Technol. 2015, 240, 1135–1143. [Google Scholar]
- Cantos, E.; Espin, J.; Lopez, C.; Barberan, F.A. Phenolic compounds and fatty acids from acorns (Quercus spp.): The main dietary constituent of free ranged iberian pigs. J. Agric. Food Chem. 2003, 51, 6248–6255. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Cao, J.Q.; Yue, J.Y.; Zhang, X.S.; Zhao, Y.Q. New triterpenoids from acorns of Quercus liaotungensis and their inhibitory activity against α-glucosidase, α-amylase and protein-tyrosine phosphatase 1B. J. Funct. Foods 2018, 41, 232–239. [Google Scholar] [CrossRef]
- Rocío, G.V.; Juan, C.E.; Francisco, A.T.B.; Nuria, E.R.G. Comprehensive characterization by LC-DAD-MS/MS of the phenolic composition of seven Quercus leaf teas. J. Food Compos. Anal. 2017, 63, 38–46. [Google Scholar]
- Jose, A.G.I.; Nuria, E.R.G.; Ruben, F.G.L.; Luis, M.T.; Carlos, A.G.A.; Luz, A.O.M.; Cecilia, E.M.S.; Betsabe, H.S.; Juan, R.R. Physicochemical properties and antioxidant capacity of oak (Quercus resinosa) leaf infusions encapsulated by spray-drying. Food Biosci. 2013, 2, 31–38. [Google Scholar]
- Moreno-Jiménez, M.R.; Trujillo-Esquivel, F.; Gallegos-Corona, M.A.; Reynoso-Camacho, R.; González-Laredo, R.F.; Gallegos-Infante, J.A.; Rocha-Guzmán, N.E.; RamosGómez, M. Antioxidant, anti-inflammatory and anticarcinogenic activities of edible red oak (Quercus Spp.) infusions in rat colon carcinogenesis induced by 1,2-dimethylhydrazine. Food Chem. Toxicol. 2015, 80, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Burgos, J.A.; Ramírez-Mares, M.V.; Larrosa, M.M.; Gallegos-Infante, J.A.; González-Laredo, R.F.; Medina-Torres, L.; Rocha-Guzmán, N.E. Antioxidant, antimicrobial: Antitopoisomerase and gastroprotective effect of herbal infusions from four Quercus species. Ind. Crops Prod. 2013, 42, 57–62. [Google Scholar] [CrossRef]
- Rocha-Guzmán, N.E.; Gallegos-Infante, J.A.; González-Laredo, R.F.; Reynoso-Camacho, R.; Ramos-Gómez, M.; García-Gasca, T.; Rodríguez-Muñoz, E.; Guzmán-Maldonado, S.H.; Medina-Torres, L.; Luján-García, B.A. Antioxidant activity and genotoxic effect on HeLa cells of phytophenolic compounds from infusions of Quercus resinosa leaves. Food Chem. 2009, 115, 1320–1325. [Google Scholar] [CrossRef]
- Rocha-Guzmán, N.E.; Medina-Medrano, J.R.; Gallegos-Infante, J.A.; González-Laredo, R.F.; Ramos-Gómez, M.; Reynoso-Camacho, R.; Guzmán-Maldonado, H.; GonzálezHerrera, S.M. Chemical evaluation, antioxidant capacity: And consumer acceptance of several oak infusions. J. Food Sci. 2012, 77, C162–C163. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.H.; Ryoo, I.J.; Kim, Y.H.; Choo, S.J.; Yoo, I.D. Free radical scavenging and antielastase activities of flavonoids from the fruits of Thuja orientalis. Arch. Pharm. Res. 2009, 32, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Pakulski, G.; Budzianowski, J. Quercetin and kaempferol glycosides of Dionaea muscipula from in vitro cultures. Planta Med. 1996, 62, 95–96. [Google Scholar] [CrossRef] [PubMed]
- Dash, R.; Acharya, C.; Bindu, P.C.; Kundu, S.C. Antioxidant potential of silk protein sericin against hydrogen peroxide-induced oxidative stress in skin fibroblasts. BMB Rep. 2008, 41, 236–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, D. Antioxidant therapies for hypolipidemia and hyperglycemia. Antioxid. Enzym. 2009, 284, 169–196. [Google Scholar]
- Fang, L.L.; Cao, J.Q.; Duan, L.L.; Tang, Y.; Zhao, Y.Q. Protein tyrosine phosphatase 1B (PTP1B) and α-glycosidase inhibitory activities of Schisandra chinensis (Turca.) Baill. J. Funct. Foods 2014, 9, 264–270. [Google Scholar] [CrossRef]
- Xu, P.; Wu, J.; Zhang, Y.; Chen, H.; Wang, Y.F. Physicochemical characterization of puerh tea polysaccharides and their antioxidant and α-glycosidase inhibition. J. Funct. Foods 2014, 6, 545–554. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 1 | Position | 2 | ||
|---|---|---|---|---|---|
| δc | δH, (J in Hz) | δc | δH, (J in Hz) | ||
| 2 | 157.2 | 2 | 158.1 | ||
| 3 | 133.4 | 3 | 135.1 | ||
| 4 | 177.5 | 4 | 179.1 | ||
| 5 | 161.2 | 5 | 163.1 | ||
| 6 | 98.8 | 6.19, d (2.0) | 6 | 99.7 | 6.16, d (1.4) |
| 7 | 164.4 | 7 | 165.7 | ||
| 8 | 93.8 | 6.40, d (2.0) | 8 | 94.5 | 6.33, d (1.4) |
| 9 | 156.5 | 9 | 158.3 | ||
| 10 | 104.0 | 10 | 105.8 | ||
| 1′ | 120.3 | 1′ | 123.1 | ||
| 2′ | 130.6 | 7.78, d (8.8) | 2′ | 117.2 | 7.64, d (1.6) |
| 3′ | 115.5 | 6.75, d (8.8) | 3′ | 145.9 | |
| 4′ | 160.2 | 4′ | 149.7 | ||
| 5′ | 115.5 | 6.75, d (8.8) | 5′ | 116.2 | 6.78, d (8.5) |
| 6′ | 130.6 | 7.78, d (8.8) | 6′ | 123.0 | 7.49, dd (8.5, 1.6) |
| 1″ (Rha) | 97.7 | 5.63, br.s | 1″ (Gal) | 101.2 | 5.68, d (7.9) |
| 2″ | 75.2 | 4.41, d (5.8) | 2″ | 74.6 | 5.44, t-like |
| 3″ | 77.8 | 3.83, dd (5.8, 7.5) | 3″ | 73.5 | 3.82, dd (3.3, 9.9) |
| 4″ | 68.1 | 3.02 m | 4″ | 70.5 | 3.92, d (3.2) |
| 5″ | 72.3 | 2.92 m | 5″ | 77.5 | 3.59, t-like |
| 6″ | 16.5 | 0.66, d (6.2) | 6″ | 62.1 | 3.68, m |
| 1‴ | 108.2 | 1‴ (galloyl) | 168.2 | ||
| 2‴ | 26.2 | 1.28 s | 2‴ | 121.6 | |
| 3‴ | 27.9 | 1.35 s | 3‴, 7‴ | 110.6 | 7.13, s |
| – | – | – | 4‴, 6‴ | 146.3 | |
| – | – | – | 5‴ | 139.8 | |
| Compound | Cell Viability | PTP1B | α-Glucosidase |
|---|---|---|---|
| % of Control a | IC50 | IC50 | |
| 1 | 50.49 ± 1.68 | 80.19 ± 5.28 b | 60.52 ± 4.19 b |
| 2 | 81.52 ± 2.66 | 5.56 ± 0.38 b | 8.59 ± 1.52 b |
| 3 | 56.22 ± 3.98 | 24.89 ± 2.39 b | 30.06 ± 3.19 b |
| 4 | 59.27 ± 4.83 | 20.56 ± 2.01 b | 25.33 ± 5.19 b |
| 5 | 55.13 ± 2.78 | 82.87 ± 8.61 b | 53.98 ± 3.09 b |
| 6 | 57.19 ± 4.68 | 81.59 ± 7.09 b | 52.09 ± 4.65 b |
| 7 | 55.21 ± 3.19 | 98.01 ± 8.21 b | 59.11 ± 5.06 b |
| 8 | 59.98 ± 4.61 | 70.52 ± 5.25 b | 49.37 ± 5.21 b |
| 9 | 58.99 ± 3.68 | 78.42 ± 6.15 b | 52.17 ± 5.91 b |
| 10 | 53.29 ± 2.76 | 89.56 ± 7.06 b | 69.38 ± 5.81 b |
| 11 | 65.35 ± 5.35 | 4.16 ± 0.56 b | 1.61 ± 0.06 b |
| 12 | 63.77 ± 4.15 | 3.92 ± 0.36 b | 2.73 ± 0.33 b |
| 13 | 67.29 ± 3.16 | 3.53 ± 0.92 b | 0.52 ± 0.09 b |
| 14 | 70.17 ± 3.26 | >100 b | >100 b |
| 15 | 69.71 ± 3.07 | 9.58 ± 1.82 b | 7.56 ± 1.99 b |
| 16 | 63.29 ± 3.18 | 15.38 ± 2.76 b | 10.96 ± 1.78 b |
| 17 | 72.27 ± 4.69 | 20.16 ± 3.02 b | 15.28 ± 1.77 b |
| 18 | 57.66 ± 4.56 | 51.11 ± 5.92 b | 63.93 ± 5.12 b |
| 19 | 68.51 ± 3.78 | 1.03 ± 0.12 b | 9.45 ± 1.62 b |
| 20 | 53.26 ± 4.13 | 81.16 ± 7.39 b | 45.08 ± 3.52 b |
| 21 | 49.79 ± 3.98 | >100 b | >100 b |
| 22 | 55.49 ± 4.49 | 79.07 ± 7.99 b | 43.58 ± 5.09 b |
| 23 | 51.22 ± 3.10 | >100 b | >100 b |
| 24 | 50.62 ± 2.91 | >100 b | >100 b |
| 25 | 52.56 ± 3.58 | 50.19 ± 6.01 b | 60.09 ± 5.29 b |
| 26 | 52.96 ± 2.86 | 70.99 ± 7.19 b | 90.89 ± 9.69 b |
| 75% EtOH extract | 64.16 ± 5.06 | 40.16 ± 4.53 c | 15.63 ± 2.11 c |
| Na3VO4 | – | 28.91 ± 2.78 b 5.32 ± 0.51 c | |
| Acarbose | – | 5.90 ± 0.98 b 3.81 ± 0.63 c |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Wang, X.; Yue, J.; Sun, Y.; Zhang, X.; Zhao, Y. Polyphenols from Acorn Leaves (Quercus liaotungensis) Protect Pancreatic Beta Cells and Their Inhibitory Activity against α-Glucosidase and Protein Tyrosine Phosphatase 1B. Molecules 2018, 23, 2167. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23092167
Xu J, Wang X, Yue J, Sun Y, Zhang X, Zhao Y. Polyphenols from Acorn Leaves (Quercus liaotungensis) Protect Pancreatic Beta Cells and Their Inhibitory Activity against α-Glucosidase and Protein Tyrosine Phosphatase 1B. Molecules. 2018; 23(9):2167. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23092167
Chicago/Turabian StyleXu, Jing, Xude Wang, Jiayin Yue, Yuanyuan Sun, Xiaoshu Zhang, and Yuqing Zhao. 2018. "Polyphenols from Acorn Leaves (Quercus liaotungensis) Protect Pancreatic Beta Cells and Their Inhibitory Activity against α-Glucosidase and Protein Tyrosine Phosphatase 1B" Molecules 23, no. 9: 2167. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23092167