Phenolic Compounds in Organic and Aqueous Extracts from Acacia farnesiana Pods Analyzed by ULPS-ESI-Q-oa/TOF-MS. In Vitro Antioxidant Activity and Anti-Inflammatory Response in CD-1 Mice

 ,
,  , , , ,
, , , ,  ,
, 
Abstract
:
1. Introduction
2. Results
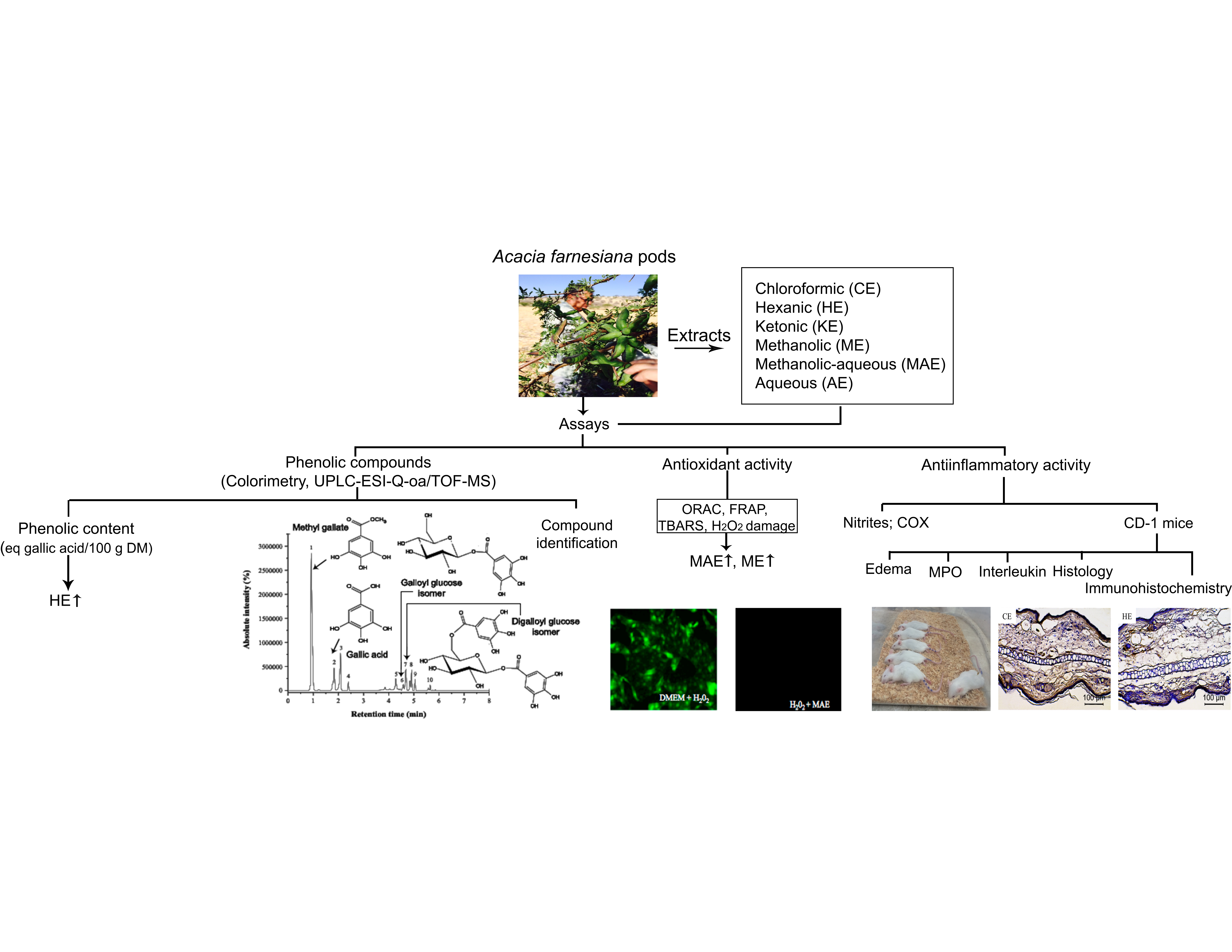
2.1. Total Polyphenols Content
2.2. AF Pod Extracts Analyzed by UPLC-ESI-Q-oa-/TOF-MS
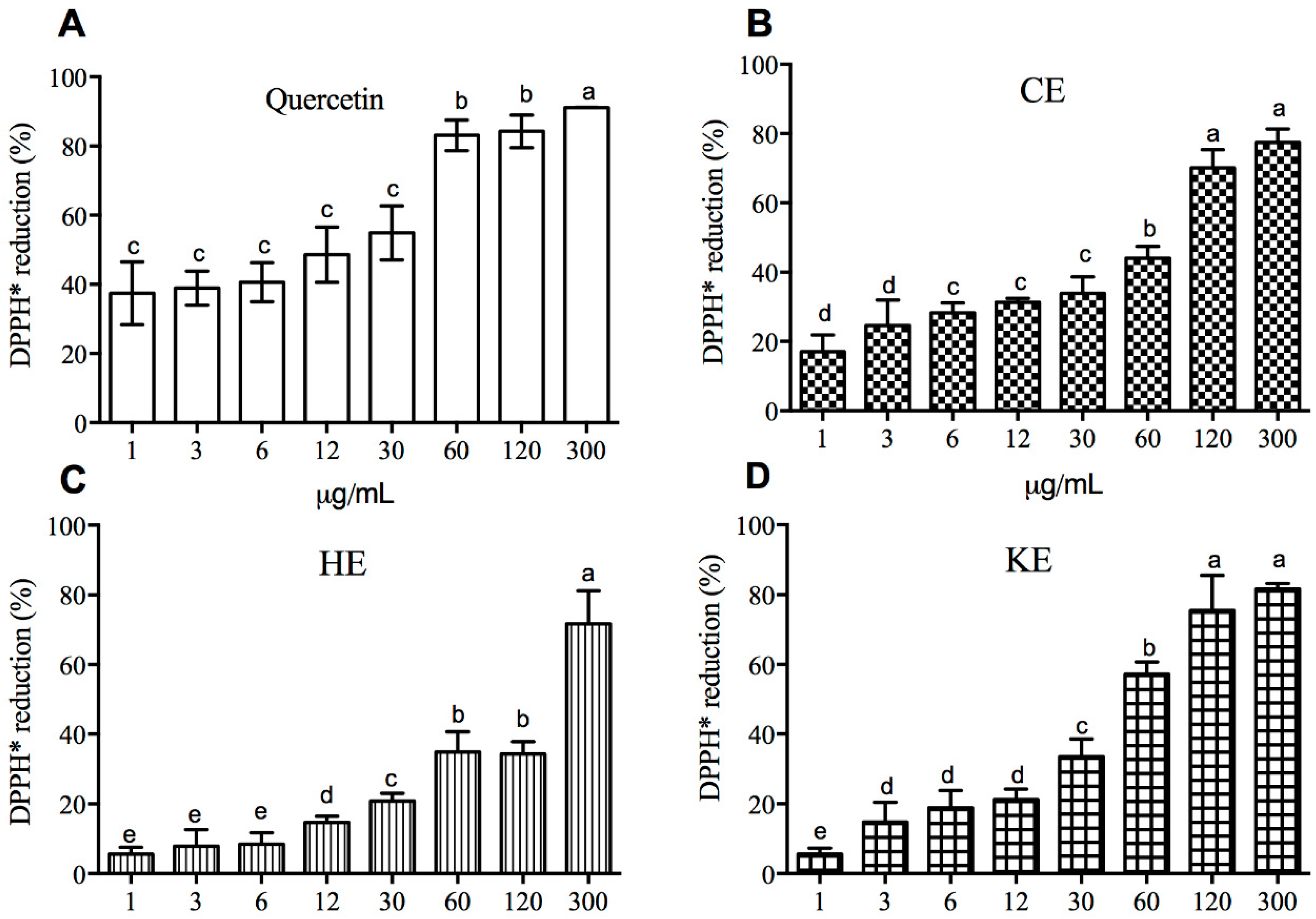
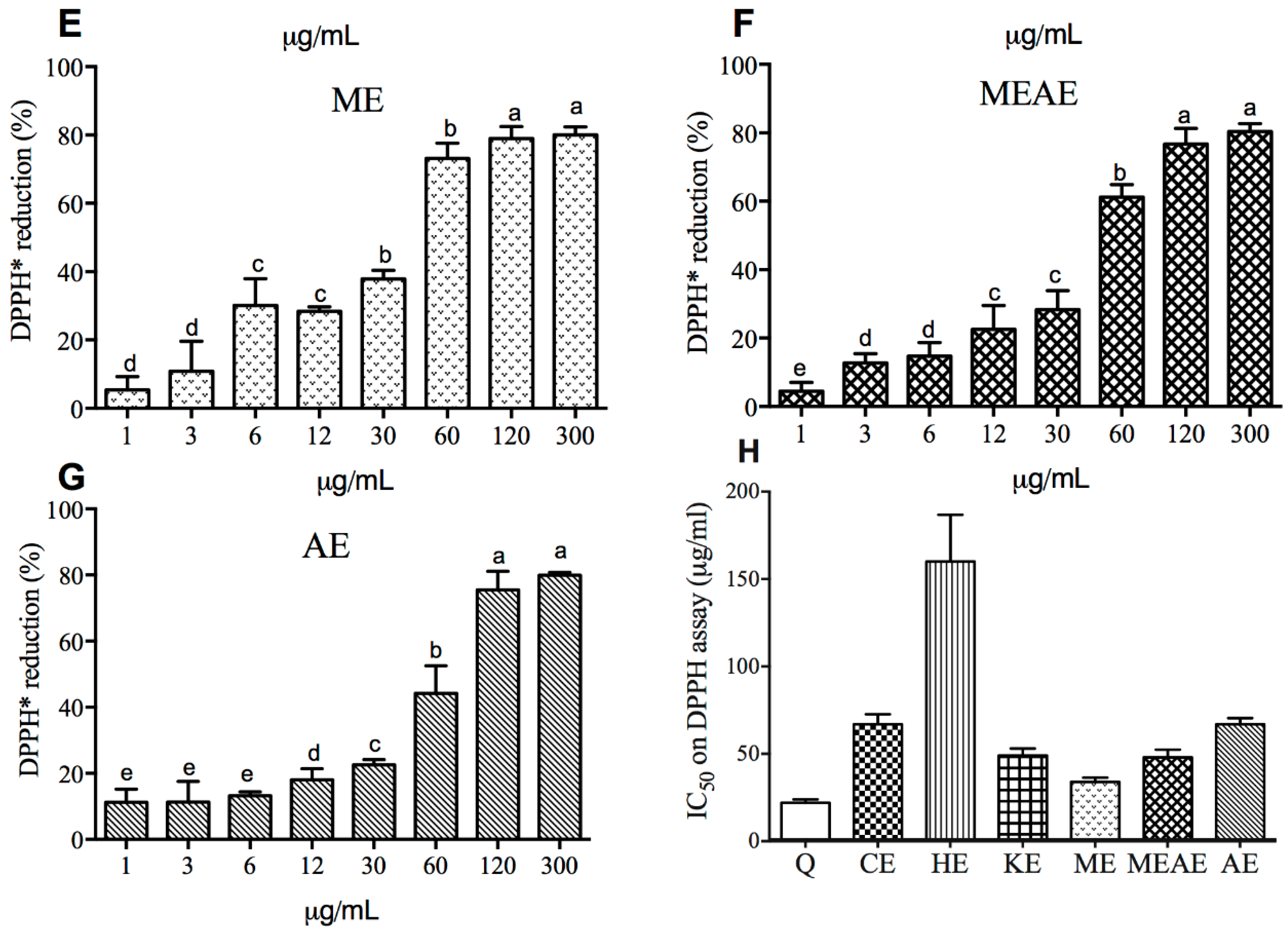
2.3. The Effect of the Extracts on the Elimination of Radicals Is Concentration-Dependent
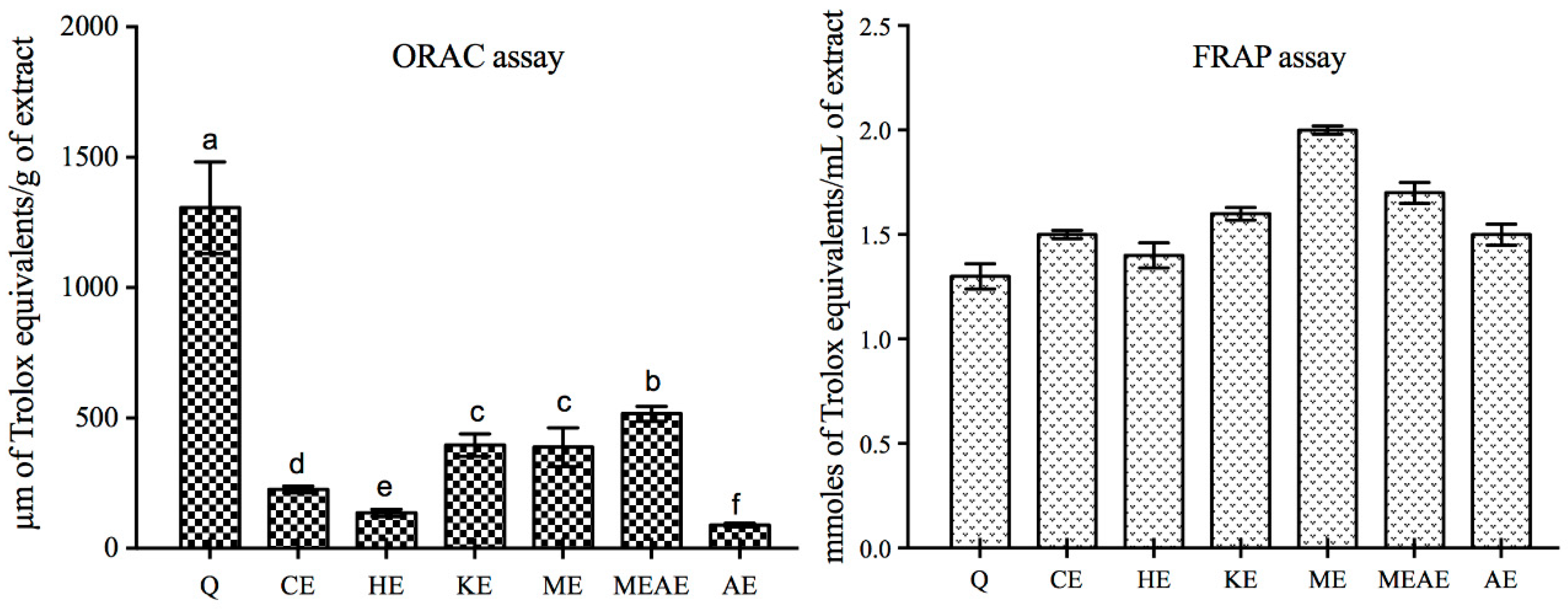
2.4. Quantitative Antioxidant Activity
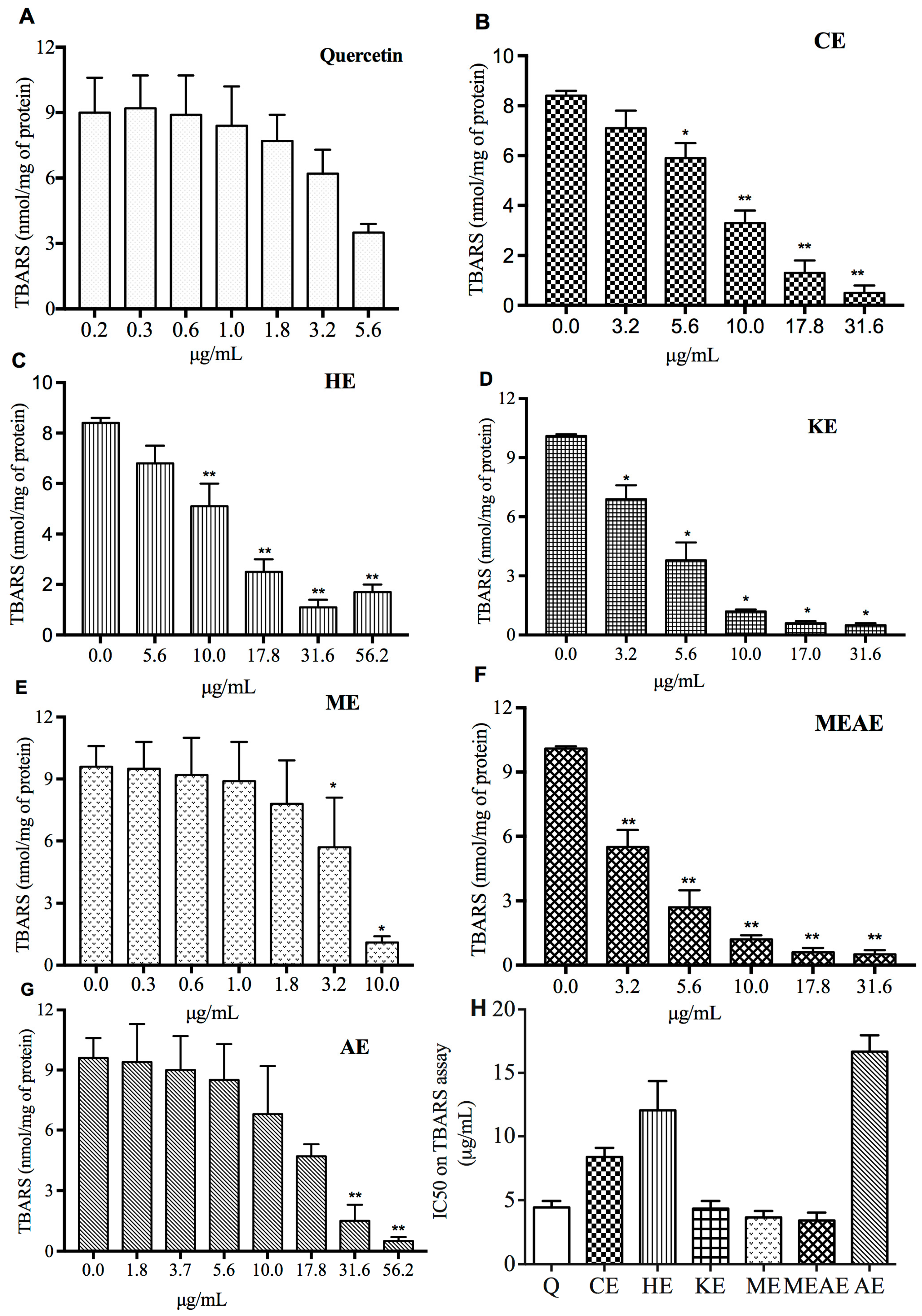
2.5. Lipid Peroxidation Is Inhibited by All Extracts in a Dose-Dependent Manner
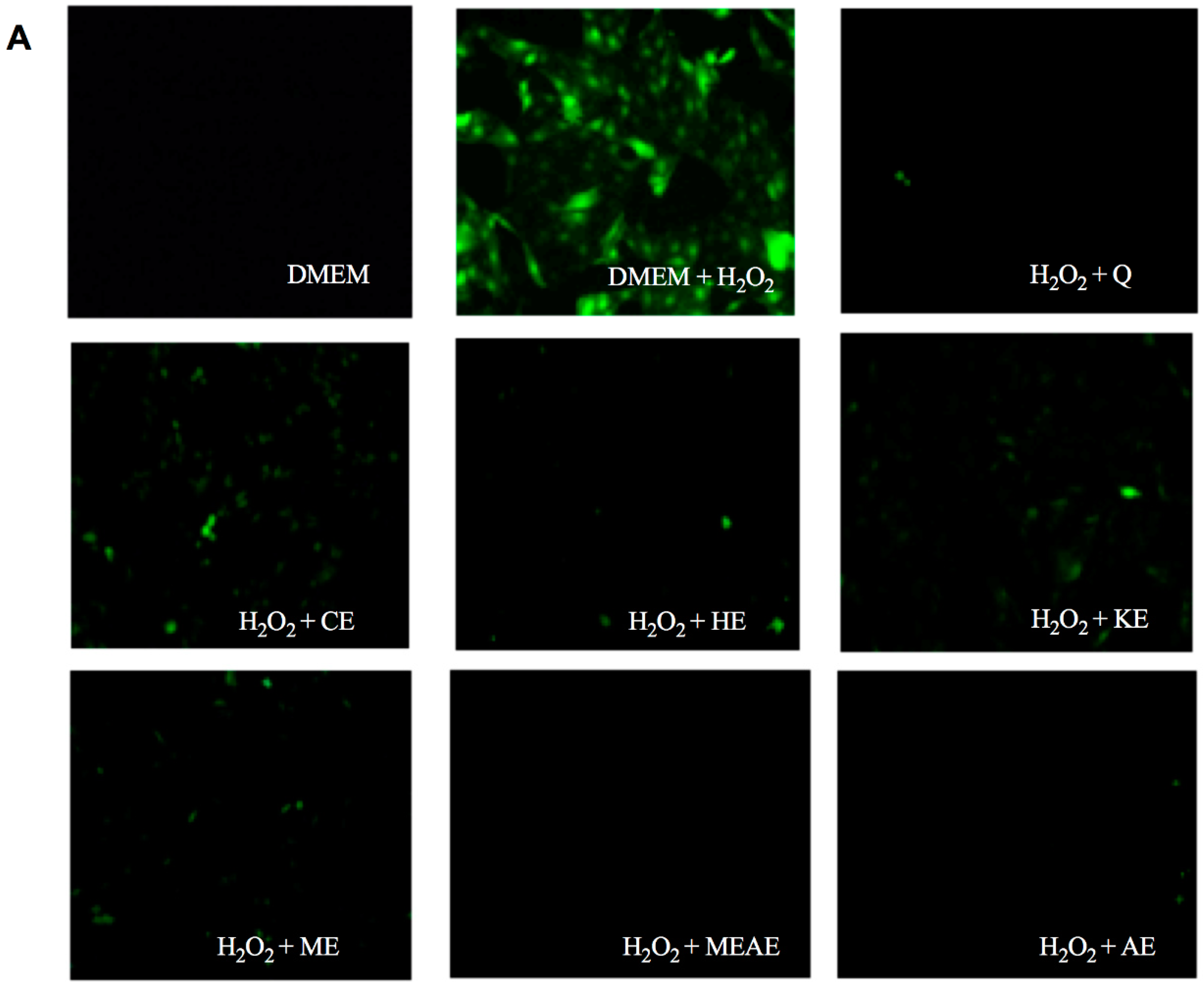
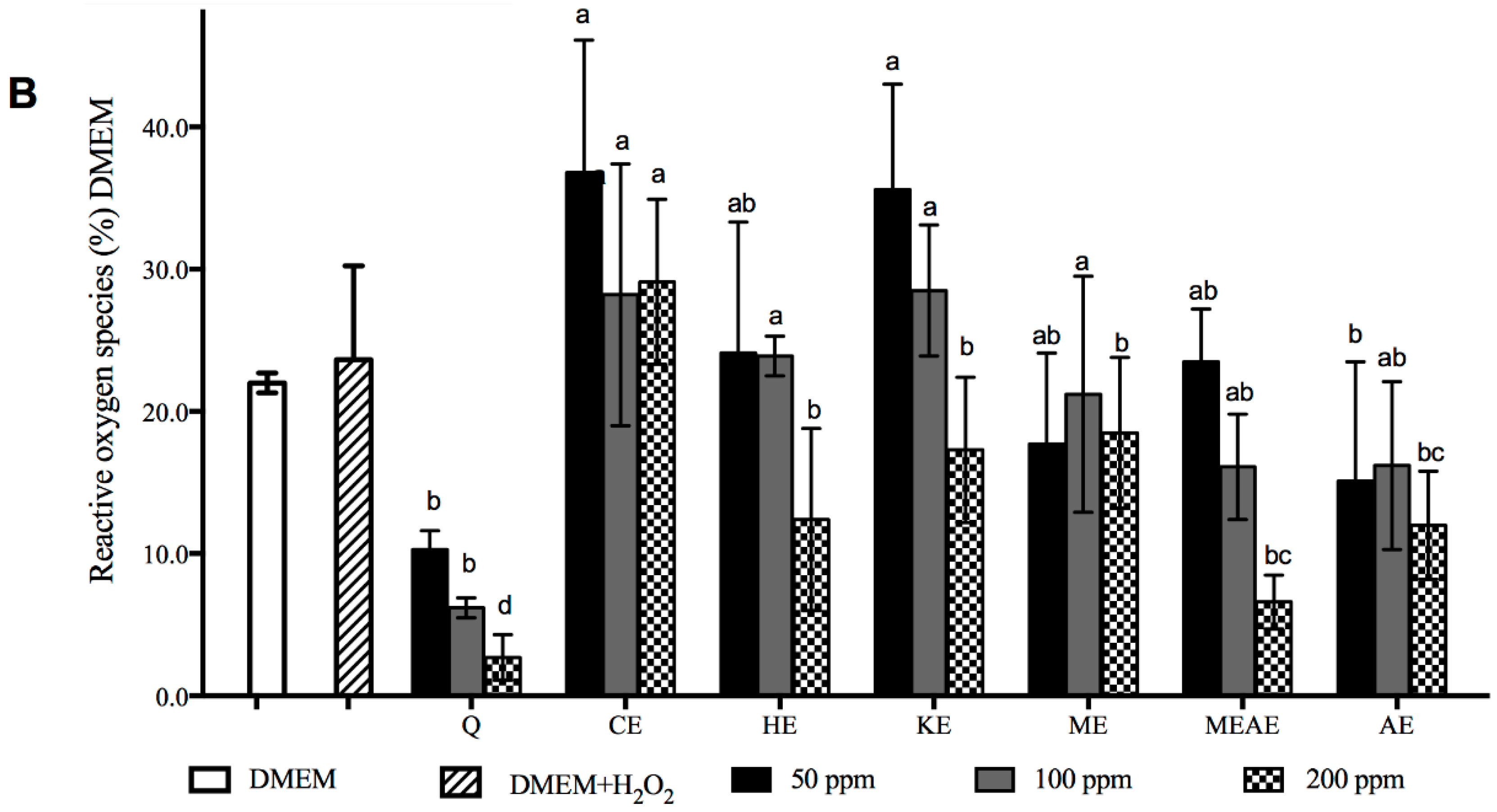
2.6. Acacia farnesiana Extracts Down Regulate Inflammation
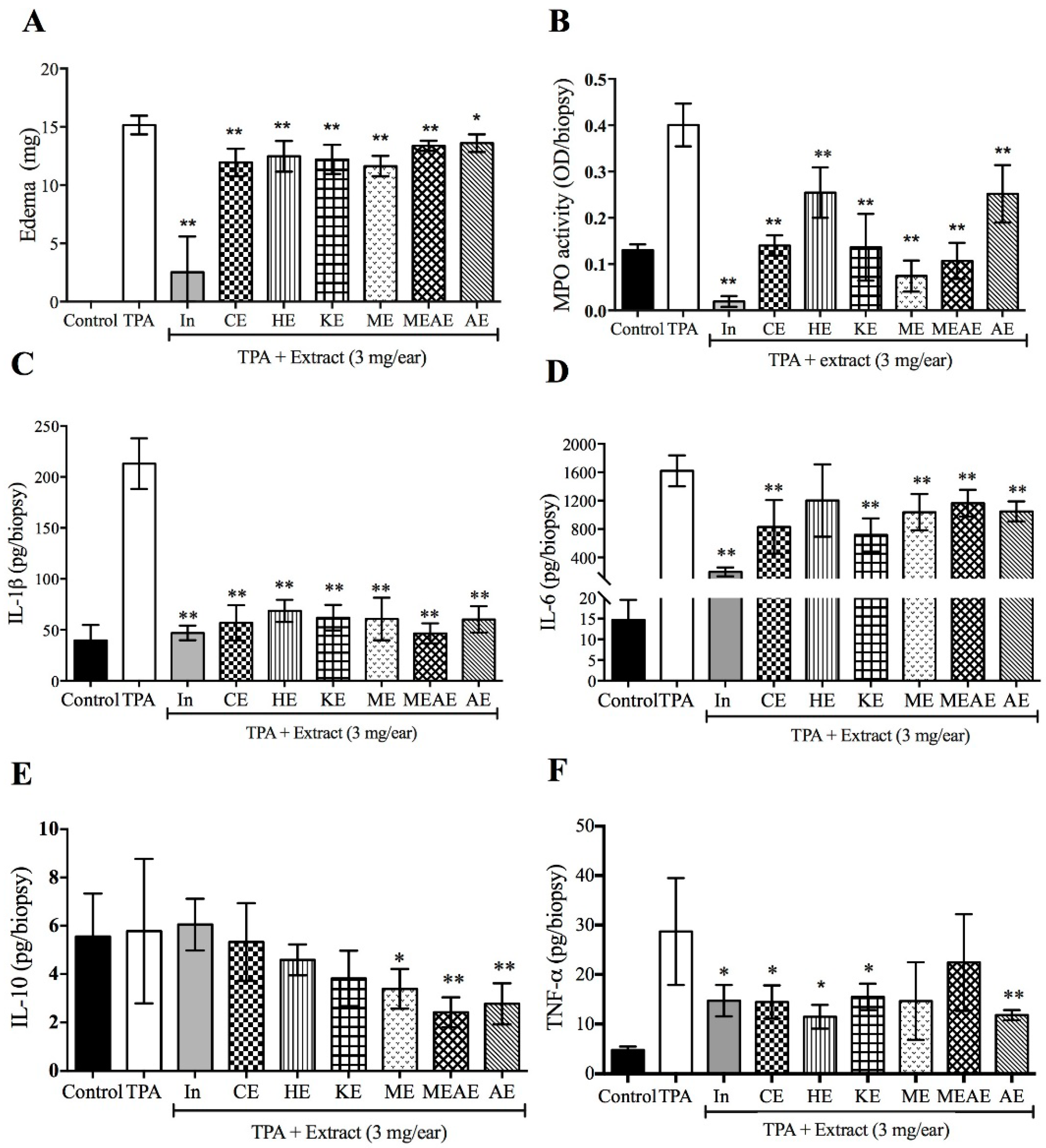
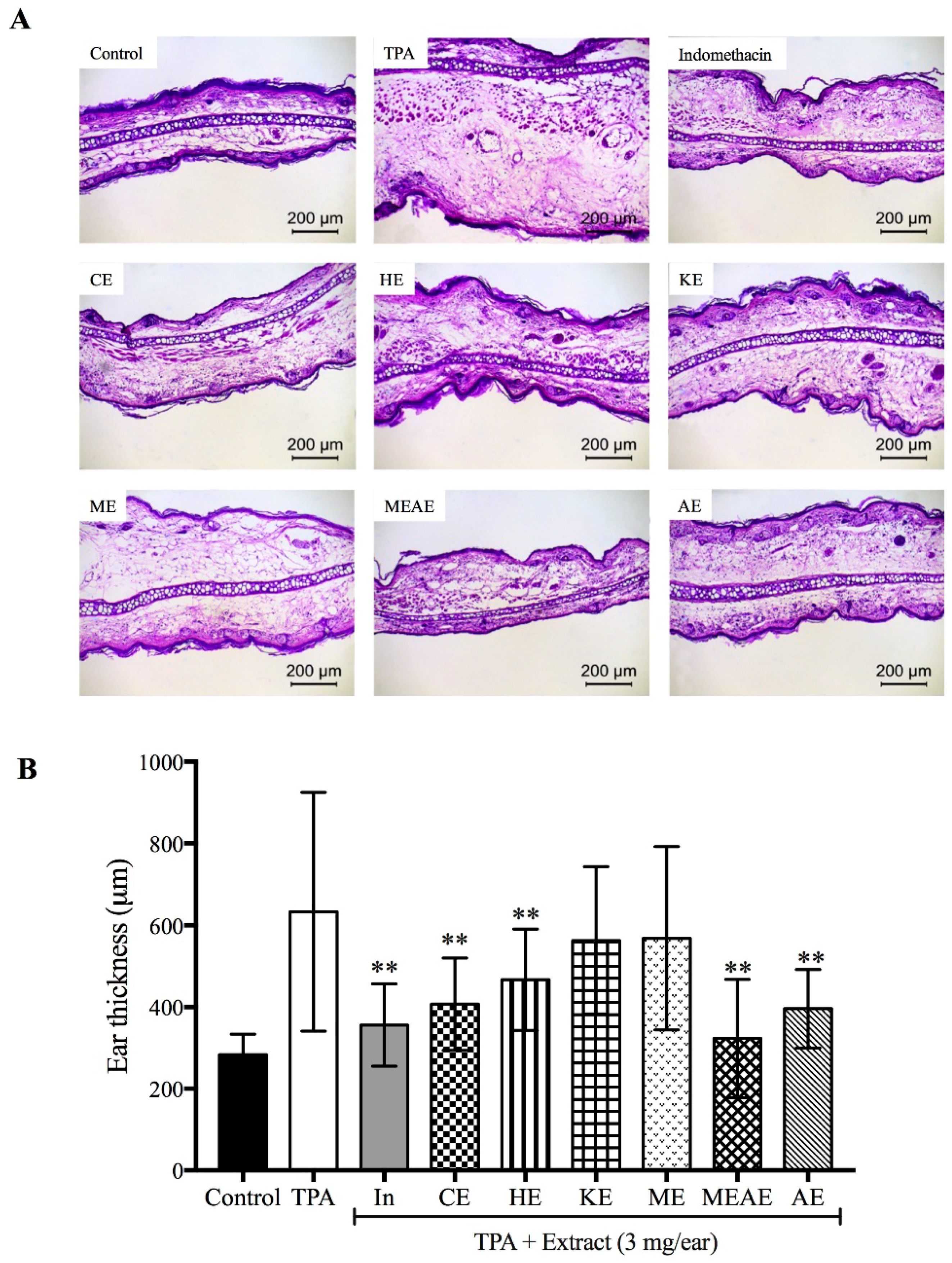
2.7. Effect of Acacia farnesiana Extracts on Mice Ear Swelling
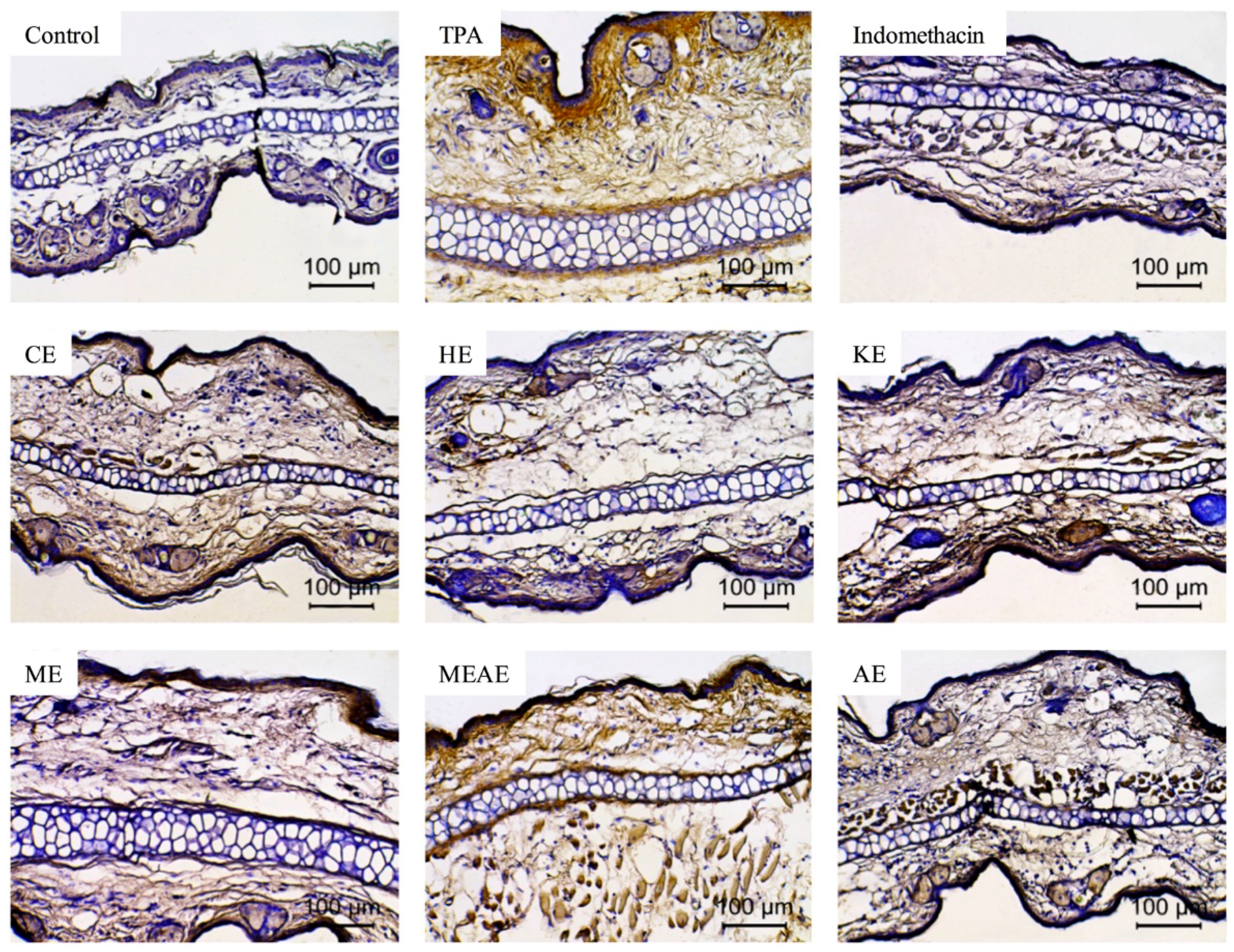
2.8. Effect of Acacia farnesiana Extracts on TNF-α-Expressing Cells
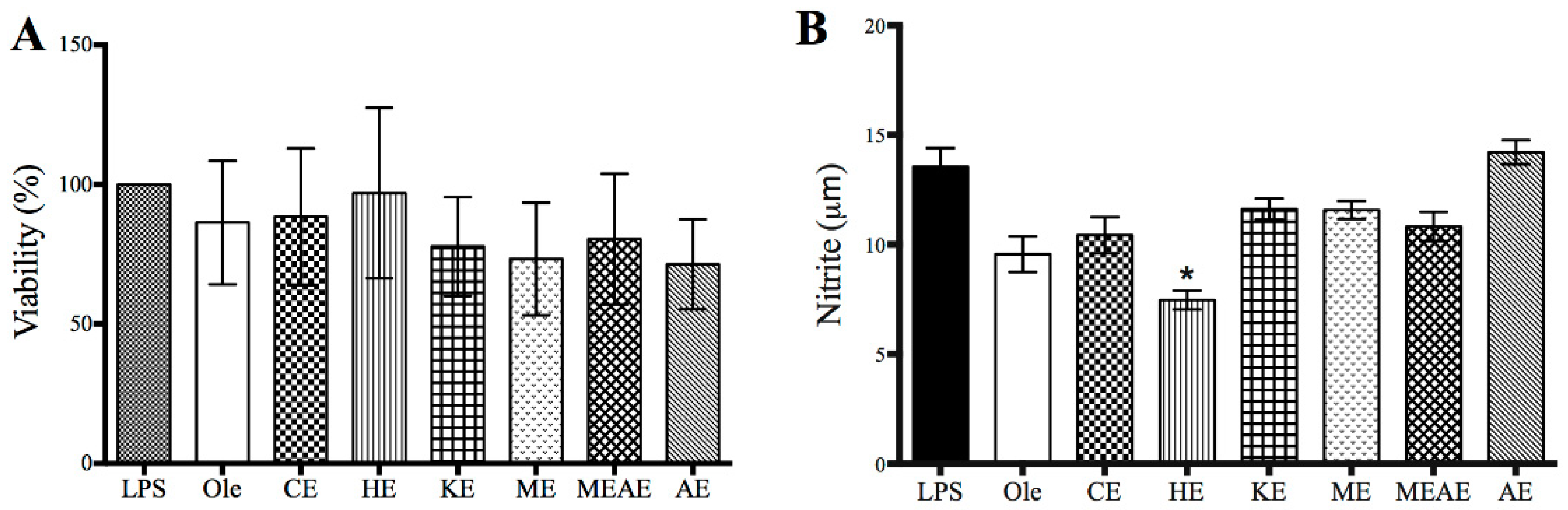
2.9. Effect of Acacia farnesiana Extracts on COX Activity, Cell Viability and Nitrite Production
3. Discussion
4. Materials and Methods
4.1. Vegetal Collection and Plant Extracts
4.2. Total Phenolic Content
4.3. UPLC-ESI-Q-oa/TOF-MS Analysis
4.4. Antioxidant Activity
4.5. Anti-Inflammatory Activity
4.5.1. Animal Experimental Set Up
4.5.2. Ear Edema Essay in CD-1 Mice Induced with 12-O-tetradecanoylphorbol Acetate (TPA)
4.5.3. Oxidative Enzyme Myeloperoxidase Assay (MPO)
4.5.4. Histological Analysis of Ear
4.5.5. Immunohistochemistry
4.5.6. Quantitation of IL-1β, IL-6, IL-10 and Tumor Necrosis Factor α (TNF-α) from Ear Extracts
4.6. In Vitro COX Inhibition and Nitrite Assay
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Halliwell, B. Free radicals and antioxidants: Updating a personal view. Nutr. Rev. 2012, 70, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, D.E.; Hurst, R.D. Polyphenolic phytochemicals–just antioxidants or much more? Cell. Mol. Life Sci. 2007, 64, 2900–2916. [Google Scholar] [CrossRef] [PubMed]
- Correa, L.B.; Pádua, T.A.; Seito, L.N.; Costa, T.E.M.M.; Silva, M.A.; Candéa, A.L.P.; Rosas, E.C.; Henriques, M.G. Anti-inflammatory effect of methyl gallate on experimental arthritis: Inhibition of neutrophil recruitment, production of inflammatory mediators, and activation of macrophages. J. Nat. Prod. 2016, 79, 1554–1566. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Valle, E.; Herrera-Ruiz, M.; Salgado, G.; Zamilpa, A.; Ocampo, M.; Aparicio, A.; Tortoriello, J.; Jiménez-Ferrer, E. Anti-inflammatory effect of 3-O-[(6′-O-Palmitoyl)-β-d-glucopyranosyl sitosterol] from Agave angustifolia on ear edema in mice. Molecules 2014, 19, 15624–15637. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.L.; Tong, Q.; Wang, W.Q.; Shi, C.Y.; Xiong, W.; Chen, J.; Liu, X.; Fang, J.G. Suppression of inflammatory responses by dihydromyricetin, a flavonoid from Ampelopsis grossedentata, via inhibiting the activation of NF-κB and MAPK signaling pathways. J. Nat. Prod. 2015, 78, 1689–1696. [Google Scholar] [CrossRef] [PubMed]
- Gasparrini, M.; Giampieri, F.; Forbes-Hernandez, T.Y.; Afrin, S.; Cianciosi, D.; Reboredo-Rodriguez, P.; Valera-Lopez, A.; Zhang, J.J.; Quiles, L.J.; Mezzetti, B.; et al. Strawberry extracts efficiently counteract inflammatory stress induced by the endotoxin lipopolysaccharide in Human Dermal Fibroblast. Food Chem. Toxicol. 2018, 114, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, M.; Nieto, A.; Marin, J.C.; Keck, A.S.; Jeffery, E.; Céspedes, C.L. Antioxidant activities of extracts from Barkleyanthus salicifolius (Asteraceae) and Penstemon gentianoides (Scrophulariaceae). J. Agri. Food Chem. 2005, 53, 5889–5895. [Google Scholar] [CrossRef]
- Oliveira, A.S.; Cercato, L.M.; de Santana Souza, M.T.; Melo, A.J.O.; Lima, B.D.S.; Duarte, M.C.; Araujo, A.A.S.; de Oliveira e Silva, A.M.; Camargo, E.A. The ethanol extract of Leonurus sibiricus L. induces antioxidant, antinociceptive and topical anti-inflammatory effects. J. Ethnopharmacol. 2017, 206, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Rauh, L.K.; Horinouchi, C.D.S.; Loddi, A.M.V.; Pietrovski, E.F.; Neris, R.; Souza-Fonseca-Guimarães, F.; Buchi, D.F.; Biavatti, M.W.; Otuki, M.F.; Cabrini, D.A. Effectiveness of Vernonia scorpioides ethanolic extract against skin inflammatory processes. J. Ethnopharmacol. 2011, 138, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, H.M.; Esmat, A. Antioxidant and anti-inflammatory activities of the major phenolics from Zygophyllum simplex L. J. Ethnopharmacol. 2017, 205, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Fortunato, L.R.; Alves, C.D.F.; Teixeira, M.M.; Rogerio, A.P. Quercetin: A flavonoid with the potential to treat asthma. Braz. J. Pharm. Sci. 2012, 48, 589–599. [Google Scholar] [CrossRef]
- Sathya, A.; Siddhuraju, P. Protective effect of bark and empty pod extracts from Acacia auriculiformis against paracetamol intoxicated liver injury and alloxan induced type II diabetes. Food Chem. Toxicol. 2013, 56, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Hernández, G.E. Aislamiento, caracterización estructural y determinación de las propiedades antibacterianas de los constituyentes de los frutos de Acacia farnesiana. Master’s Thesis, Facultad de Ciencias Químicas, Universidad Autónoma de Nuevo León, San Nicolás de los Garza, Mexico, 2017. [Google Scholar]
- Fernandez, P.M.S.; Villano, D.; Troncoso, A.M.; Garcia, P.M.C. Antioxidant activity of phenolic compounds: From in vitro results to in vivo evidence. Crit. Rev. Food Sci. Nutr. 2008, 48, 649–671. [Google Scholar] [CrossRef] [PubMed]
- Cuchillo, H.M.; Puga, D.C.; Wrage-Mönning, N.; Espinosa, M.J.G.; Montaño, B.S.; Navarro-Ocaña, A.; Ledesma, J.A.; Diaz, M.M.; Pérez-Gil, R.F. Chemical composition, antioxidant activity and bioactive compounds of vegetation species ingested by goats on semiarid rangelands. J. Anim. Feed Sci. 2013, 22, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Delgadillo, P.C.; Cuchillo, H.M.; Espinosa, M.J.G.; Medina, C.O.; Molina, J.E.; Díaz, M.M.; Álvarez, I.M.A.; Ledesma, S.J.A.; Pedraza-Chaverri, J. Antioxidant activity and protection against oxidative-induced damage of Acacia shaffneri and Acacia farnesiana pods extracts: In vitro and in vivo assays. BMC Complement. Altern. Med. 2015, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Akdis, M.; Burgler, S.; Crameri, R.; Eiwegger, T.; Fujita, H.; Gomez, E.; Klunker, S.; Meyer, N.; O’Mahony, L.; Palomares, O.; et al. Interleukins, from 1 to 37, and interferon-γ: Receptors, functions, and roles in diseases. J. Allergy Clin. Immunol. 2011, 127, 701–721. [Google Scholar] [CrossRef] [PubMed]
- García, J.S.A.; Verde, M.J.S.; Heredia, L.N. Traditional uses and scientific knowledge of medicinal plants from Mexico and Central America. J. Herbs Spices Med. Plants 2001, 8, 37–89. [Google Scholar] [CrossRef]
- Bae, S.; Kim, S.Y.; Do, M.H.; Lee, C.H.; Song, Y.J. 1,2,3,4,6-Penta-O-galloyl-ß-d-glucose, a bioactive compound in Elaeocarpus sylvestris extract, inhibits varicella-zoster virus replication. Antivir. Res. 2017, 144, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Ambigaipalan, P.; de Camargo, A.C.; Shahidi, F. Identification of phenolic antioxidants and bioactives of pomegranate seeds following juice extraction using HPLC-DAD-ESI-MSn. Food Chem. 2017, 221, 1883–1894. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Zhao, X.; Wang, P.; Jia, P.; Zhao, X.; Xiao, C.; Zheng, X. Metabolite characterization of Penta-O-galloyl-beta-d-glucose in rat biofluids by HPLC-QTOF-MS. Chin. Herb. Med. 2018, 10, 73–79. [Google Scholar] [CrossRef]
- Li, J.; Kuang, G.; Chen, X.; Zeng, R. Identification of chemical composition of leaves and flowers from Paeonia rockii by UHPLC-Q-Exactive Orbitrap HRMS. Molecules 2016, 21, 947. [Google Scholar] [CrossRef] [PubMed]
- Slatnar, A.; Mikulic-Petkovsek, M.; Stampar, F.; Veberic, R.; Solar, A. Identification and quantification of phenolic compounds in kernels, oil and bagasse pellets of common walnut (Juglans regia L.). Food Res. Int. 2015, 67, 255–263. [Google Scholar] [CrossRef]
- Tasioula-Margari, M.; Tsabolatidou, E. Extraction, separation, and identification of phenolic ompounds in virgin olive oil by HPLC-DAD and HPLC-MS. Antioxidants 2015, 4, 548–562. [Google Scholar] [CrossRef] [PubMed]
- Llorent-Martínez, E.J.; Spínola, V.; Castilho, P.C. Phenolic profiles of Lauraceae plant species endemic to Laurisilva forest: A chemotaxonomic survey. Ind. Crops Prod. 2017, 107, 1–12. [Google Scholar] [CrossRef]
- Spínola, V.; Pinto, J.; Castilho, P.C. Identification and quantification of phenolic compounds of selected fruits from Madeira Island by HPLC-DAD–ESI-MSn and screening for their antioxidant activity. Food Chem. 2015, 173, 14–30. [Google Scholar] [CrossRef] [PubMed]
- Salem, M.M.; Davidorf, H.F.; Abdel-Rahman, H.M. In vitro anti-uveal melanoma activity pf phenols compounds fron the Egyptian medicianla plant Acacia nilotica. Fitoterapia 2011, 82, 1279–1284. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.S.; Lin, C.R.; Du, Y.C.; Lübken, T.; Chiang, M.Y.; Chen, I.H.; Wu, C.C.; Hwang, T.L.; Chen, S.L.; Yen, M.H.; et al. Acasiane A and B and farnesirane A and B, diterpene derivatives from the roots of Acacia farnesiana. Planta Med. 2009, 75, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Zhu, R.; Wu, J.; Wu, J.; Xia, W.; Zhu, L.; Jiang, W.; Xiang, S.; Tan, Z. In vivo protective effect of betulinic acid on dexamethasone induced thymocyte apoptosis by reducing oxidative stress. Pharmacol. Rep. 2016, 68, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Karoune, S.; Falleh, H.; Kechebar, M.S.A.; Halis, Y.; Mkadmini, K.; Belhamra, M.; Rahmoune, C.; Ksouri, R. Evaluation of antioxidant activities of the edible and medicinal Acacia albida organs related to phenolic compounds. Nat. Prod. Res. 2015, 29, 452–454. [Google Scholar] [CrossRef] [PubMed]
- Fernández, M.A.; de las Heras, B.; Garcia, M.D.; Sáenz, M.T.; Villar, A. New insights into the mechanism of action of the anti-inflammatory triterpene lupeol. J. Pharm. Pharmacol. 2001, 53, 1533–1539. [Google Scholar] [CrossRef] [PubMed]
- Adkins, Y.; Kelley, D.S. Mechanisms underlying the cardioprotective effects of omega-3 polyunsaturated fatty acids. J. Nutr. Biochem. 2010, 21, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Endo, J.; Arita, M. Cardioprotective mechanism of omega-3 polyunsaturated fatty acids. J. Cardiol. 2016, 67, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Romero, E.A.; Maldonado, M.A.; González, C.J.; Bahena, S.M.; Garduño, R.M.L.; Rodríguez, L.V.; Alvarez, L. Anti-inflammatory and antioxidative effects of six pentacyclic triterpenes isolated from the Mexican copal resin of Bursera copallifera. BMC Complement. Altern. Med. 2016, 16, 422. [Google Scholar] [CrossRef]
- Horinouchi, C.D.; Mendes, D.A.; Soley Bda, S.; Pietrovski, E.F.; Facundo, V.A.; Santos, A.R.S.; Cabrini, D.A.; Otuki, M.F. Combretum leprosum Mart. (Combretaceae): Potential as an antiproliferative and anti-inflammatory agent. J. Ethnopharmacol. 2013, 145, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Heredia, N.; Camacho, C.M.R.; García, S. Isolation, characterization and mode of antimicrobial action against Vibrio cholerae of methyl gallate isolated from Acacia farnesiana. J. App. Microbiol. 2013, 115, 1307–1316. [Google Scholar] [CrossRef] [PubMed]
- Crispo, J.A.G.; Piché, M.; Ansell, D.R.; Eibl, J.K.; Tai, I.T.; Kumar, A.; Ross, G.M.; Tai, T.C. Protective effects of methyl gallate on H2O2-induced apoptosis in PC12 cells. Biochem. Biophys. Res. Commun. 2010, 393, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Huang, D.; Hampsch-Woodill, M.; Flanagan, J.A.; Deemer, E.K. Analysis of antioxidant activities of common vegetables employing oxygen radical absorbance capacity (ORAC) and ferric reducing antioxidant power (FRAP) assays: A comparative study. J. Agric. Food Chem. 2002, 50, 3122–3128. [Google Scholar] [CrossRef] [PubMed]
- Liang, N.; Kitts, D.D. Antioxidant property of coffee components: Assessment of methods that define mechanisms of action. Molecules 2014, 19, 19180–19208. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Koren, E.; Kohen, R.; Ginsburg, I. Polyphenols enhance total oxidant-scavenging capacities of human blood by binding to red blood cells. Exp. Biol. Med. 2010, 235, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-throughput assay of oxygen radical absorbance capacity (ORAC) using a multichannel liquid handling system coupled with a microplate fluorescence reader in 96-well format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Hernández, F.K.; Cárdenas, R.N.; Pedraza, C.J.; Massieu, L. Calcium-dependent production of reactive oxygen species is involved in neuronal damage induced during glycolysis inhibition in cultured hippocampal neurons. J. Neurosci. Res. 2008, 86, 1768–1780. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Esterbauer, H.; Cheeseman, K.H. Determination of aldehydic lipid peroxidation products: Malonaldehyde and 4-hydroxynonenal. Methods Enzymol. 1990, 186, 407–421. [Google Scholar] [CrossRef] [PubMed]
- Del-Ángel, M.; Nieto, A.; Ramírez-Apan, T.; Delgado, G. Anti-inflammatory effect of natural and semi-synthetic phthalides. Eur. J. Pharmacol. 2015, 752, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Ota, H.; Sasagawa, S.; Sakatani, T.; Fujikura, T. Assay method for myeloperoxidase in human polymorphonuclear leukocytes. Anal. Biochem. 1983, 132, 345–352. [Google Scholar] [CrossRef]
- Bradley, P.P.; Priebat, D.A.; Christensen, R.D.; Rothstein, G. Measurement of cutaneous inflammation: Estimation of neutrophil content with an enzyme marker. J. Investig. Dermatol. 1982, 78, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Leal-Díaz, A.M.; Noriega, L.G.; Torre-Villalvazo, I.; Torres, N.; Alemán-Escondrillas, G.; López-Romero, P.; Sánchez-Tapia, M.; Aguilar-López, M.; Furuzawa-Carballeda, J.; Velázquez-Villegas, L.A.; et al. Aguamiel concentrate from Agave salmiana and its extracted saponins attenuated obesity and hepatic steatosis and increased Akkermansia muciniphila in C57BL/6 mice. Sci. Rep. 2016, 6, 34242. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Flores, S.; Hernandez-Molina, G.; Enriquez, A.B.; Faz-Muñoz, D.; Esquivel, Y.; Pacheco-Molina, C.; Furuzawa-Carballeda, J. Cytokines and effector/regulatory cells characterization in the physiopathology of cutaneous lupus erythematous: A cross-sectional study. Mediat. Inflamm. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Rt (min) | [M − H]− (m/z) | Molecular Formula | Dominant Ion Fragment | Assignment | Reference |
|---|---|---|---|---|---|---|
| Chloroformic | ||||||
| 1 | 0.866 | 387.0918 | - | 341.1011 (100) | Unknow | - |
| 2 | 0.935 | 331.0602 | C13H16O10 | 271.0561 (100) | Galloyl glucose isomer 1 | [19] |
| 3 | 1.826 | 169.0484 | C7H6O5 | 125.0678 (100) | Gallic acid | [20] |
| 4 | 1.912 | 331.0612 | C13H16O10 | 271.0566 (100) | Galloyl glucose isomer 2 | [19] |
| 5 | 2.169 | 331.0610 | C13H16O10 | 271.0572 (100) | Galloyl glucose isomer 3 | [19] |
| 6 | 2.512 | 493.0616 | - | 271.0562 (100) | Unknow | - |
| 7 | 4.673 | 183.0608 | C8H8O | 168.9363 (100) | Methyl gallate | [21] |
| 8 | 4.776 | 483.0230 | C20H20O14 | - | Digalloyl glucose isomer 1 | [22] |
| 9 | 4.982 | 483.0224 | C20H20O14 | - | Digalloyl glucose isomer 2 | [22] |
| Hexanic | ||||||
| 1 | 0.866 | 387.0918 | - | 341.1011 (100) | Unknow | - |
| 2 | 0.935 | 331.0602 | C13H16O10 | 271.0561 (100) | Galloyl glucose isomer 1 | [19] |
| 3 | 1.826 | 169.0484 | C7H6O5 | 125.0678 (100) | Gallic acid | [20] |
| 4 | 1.912 | 331.0612 | C13H16O10 | 271.0566 (100) | Galloyl glucose isomer 2 | [19] |
| 5 | 2.169 | 331.0610 | C13H16O10 | 271.0572 (100) | Galloyl glucose isomer 3 | [19] |
| 6 | 2.512 | 493.0616 | - | 271.0562 (100) | Unknow | - |
| 7 | 4.673 | 183.0608 | C8H8O | 168.9363 (100) | Methyl gallate | [21] |
| 8 | 4.776 | 483.0230 | C20H20O14 | - | Digalloyl glucose isomer 1 | [22] |
| 9 | 4.982 | 483.0224 | C20H20O14 | - | Digalloyl glucose isomer 2 | [22] |
| Ketonic | ||||||
| 1 | 0.917 | 331.0600 | C13H16O10 | 271.0560 (45) 211.0483 (1) | Galloyl glucose isomer 1 | [19] |
| 2 | 1.826 | 331.0605 | C13H16O10 | 271.0565 (100) | Galloyl glucose isomer 2 | [19] |
| 3 | 2.084 | 331.0597 | C13H16O10 | 271.0559 (100) | Galloyl glucose isomer 3 | [19] |
| 4 | 2.392 | 493.0606 | - | 271.0557 (17) 313.0538 (1) 169.0465 (1) | Unknow | - |
| 5 | 4.279 | 483.0223 | C20H20O14 | 183.0624 (100) | Digalloyl glucose isomer 1 | [23] |
| 6 | 4.588 | 183.0610 | C8H8O | 168.9363 (100) | Methyl gallate | [21] |
| 7 | 4.691 | 483.0228 | C20H20O14 | 271.0562 (15) 211.0498 (8) 169.0481 (49) 125.0660 (1) | Digalloyl glucose isomer 2 | [22] |
| 8 | 4.914 | 483.0219 | C20H20O14 | 271.0562 (36) 211.0494 (3) 169.0481 (28) 125.0646 (1) | Digalloyl glucose isomer 3 | [22] |
| 9 | 5.051 | 483.0226 | C20H20O14 | - | Digalloyl glucose isomer 4 | [22] |
| 10 | 5.651 | 401.0812 | - | - | Unknow | - |
| Methanolic | ||||||
| 1 | 0.832 | 331.0601 | C13H16O10 | 271.0561 (40) 169.0482 (1) 133.0557 (1) | Galloyl glucose isomer 1 | [19] |
| 2 | 1.072 | 331.0606 | C13H16O10 | 271.0565 (100) | Galloyl glucose isomer 2 | [19] |
| 3 | 1.603 | 483.0223 | C20H20O14 | 271.0487 (100) | Digalloyl glucose isomer 1 | [19] |
| 4 | 1.912 | 483.0223 | C20H20O14 | 271.0566 (100) | Digalloyl glucose isomer 2 | [19] |
| 5 | 2.581 | 183.0606 | C8H8O | 168.9363 (100) | Methyl gallate | [21] |
| Methanolic: Aqueous | ||||||
| 1 | 0.9 | 387.0913 | - | 377.0659 (40) 341.1000 (18) | Unknow | - |
| 2 | 1.02 | 331.0603 | C13H16O10 | 271.0562 (100) | Galloyl glucose isomer 1 | [19] |
| 3 | 1.946 | 169.0483 | C7H6O5 | 125.0661 (100) | Gallic acid | [19] |
| 4 | 2.118 | 331.0606 | C13H16O10 | 271.0565 (100) | Galloyl glucose isomer 2 | [19] |
| 5 | 2.41 | 331.0611 | C13H16O10 | 271.0569 (100) | Galloyl glucose isomer 3 | [19] |
| 6 | 2.821 | 493.0619 | - | 271.0559 (100) | Unknow | - |
| 7 | 4.965 | 183.0604 | C8H8O | 168.9363 (100) | Methyl gallate | [21] |
| 8 | 5.171 | 483.0225 | C20H20O14 | 271.0564 (100) | Digalloyl glucose isomer 1 | [22] |
| Aqueous | ||||||
| 1 | 0.8666 | 195.0799 | - | 165.0759 (100) | Hydroxytyrosol acetate | [24] |
| 2 | 1.02 | 133.0546 | - | 115.0463 (100) | unknow | - |
| 3 | 1.26 | 191.0494 | C7H12O6 | 111.0524 (100) | Quinic acid | [25] |
| 4 | 1.398 | 295.0470 | C13H12O8 | - | Caffeoylmalic acid | [26] |
| 5 | 1.929 | 169.0478 | C7H6O5 | 125.0657 (100) | Gallic acid | [23] |
| 6 | 4.879 | 183.0609 | C8H8O | 168.9363 (100) | Methyl gallate | [21] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Claudia, D.P.; Mario, C.-H.; Arturo, N.O.; Omar Noel, M.-C.; Antonio, N.C.; Teresa, R.A.; Zenón Gerardo, L.-T.; Margarita, D.M.; Marsela Alejandra, Á.-I.; Yessica Rosalina, C.M.; et al. Phenolic Compounds in Organic and Aqueous Extracts from Acacia farnesiana Pods Analyzed by ULPS-ESI-Q-oa/TOF-MS. In Vitro Antioxidant Activity and Anti-Inflammatory Response in CD-1 Mice. Molecules 2018, 23, 2386. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23092386
Claudia DP, Mario C-H, Arturo NO, Omar Noel M-C, Antonio NC, Teresa RA, Zenón Gerardo L-T, Margarita DM, Marsela Alejandra Á-I, Yessica Rosalina CM, et al. Phenolic Compounds in Organic and Aqueous Extracts from Acacia farnesiana Pods Analyzed by ULPS-ESI-Q-oa/TOF-MS. In Vitro Antioxidant Activity and Anti-Inflammatory Response in CD-1 Mice. Molecules. 2018; 23(9):2386. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23092386
Chicago/Turabian StyleClaudia, Delgadillo Puga, Cuchillo-Hilario Mario, Navarro Ocaña Arturo, Medina-Campos Omar Noel, Nieto Camacho Antonio, Ramírez Apan Teresa, López-Tecpoyotl Zenón Gerardo, Díaz Martínez Margarita, Álvarez-Izazaga Marsela Alejandra, Cruz Martínez Yessica Rosalina, and et al. 2018. "Phenolic Compounds in Organic and Aqueous Extracts from Acacia farnesiana Pods Analyzed by ULPS-ESI-Q-oa/TOF-MS. In Vitro Antioxidant Activity and Anti-Inflammatory Response in CD-1 Mice" Molecules 23, no. 9: 2386. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23092386