Recovery of Antimicrobials and Bioaccessible Isoflavones and Phenolics from Soybean (Glycine max) Meal by Aqueous Extraction

 ,
,  ,
,  , and
, and
Abstract
:
1. Introduction
2. Results
2.1. Identification and Quantification of Bioactive Compounds in Soybean Meal Aqueous Extract and Their Bioaccessibility
2.2. Bioactivities of Soybean Meal Aqueous Extract
2.2.1. Antioxidant Activity
2.2.2. Antimicrobial Activity
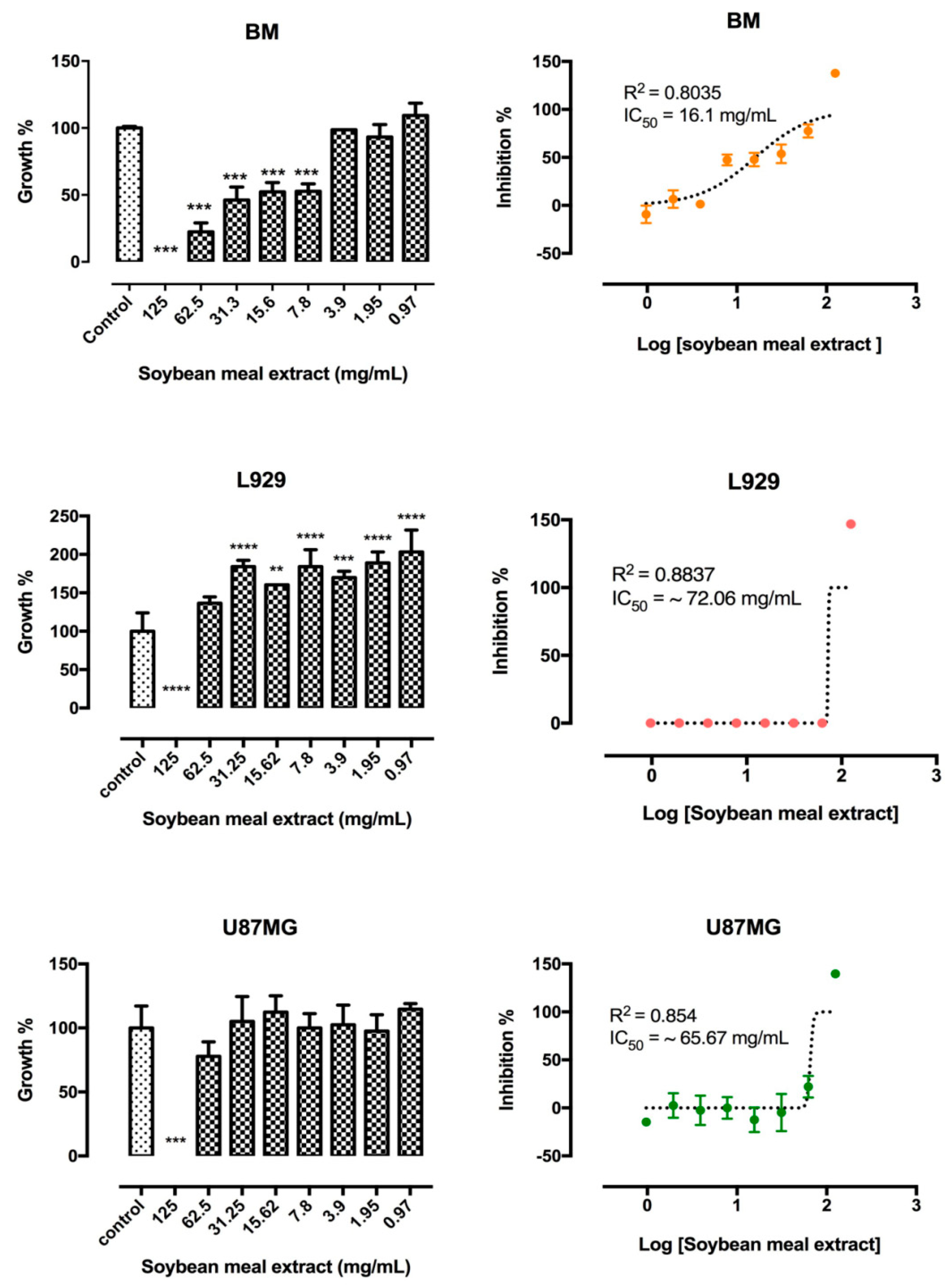
2.2.3. In Vitro Cytotoxicity Assay in Different Cell Lineages
3. Discussion
4. Materials and Methods
4.1. Organisms
4.2. Preparation of Soybean Meal Aqueous Extract
4.3. Identification and Quantification of Isoflavones Forms by LC-DAD-FLD
4.4. Identification and Quantification of Phenolic Compounds (PC)
4.5. Evaluation of the Antimicrobial Activity
4.6. Determination of Total Antioxidant Activity
4.6.1. Determination of FRAP
4.6.2. Determination of TEAC
4.6.3. Determination of TAP
4.6.4. Determination of ORAC
4.7. Lipid Peroxidation Inhibition
4.8. In Vitro Simulated Gastrointestinal Digestion
4.9. In Vitro Cytotoxicity Assays Against Murine and Human Cells
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zakir, M.M.; Freitas, I.R. Benefícios à saúde humana do consumo de isoflavonas presentes em produtos derivados da soja. J. Bioen. Food Sci. 2015, 2, 3–10. [Google Scholar] [CrossRef]
- Day, L. Proteins from land plants—Potential resources for human nutrition and food security. Trends Food Sci. Technol. 2013, 32, 25–42. [Google Scholar] [CrossRef]
- Wang, D.; Thakker, C.; Liu, P.; Bennett, G.N.; San, K.Y. Efficient production of free fatty acids from soybean meal carbohydrates. Biotechnol. Bioeng. 2015, 112, 2324–2333. [Google Scholar] [CrossRef] [PubMed]
- Villalobos, M.D.C.; Serradilla, M.J.; Martín, A.; Ordiales, E.; Ruiz-Moyano, S.; Córdoba, M.D.G. Antioxidant and antimicrobial activity of natural phenolic extract from defatted soybean flour by-product for stone fruit postharvest application. J. Sci. Food Agric. 2016, 96, 2116–2124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, R.; Zhao, W.; Hua, X.; Zhang, W. Application of high density steam flash-explosion in protein extraction of soybean meal. J. Food Engin. 2013, 116, 430–435. [Google Scholar] [CrossRef]
- Martino, H.S.D.; Cardoso, L.M.; Ribeiro, S.M.R.; Dantas, M.I.S.; Piovesan, N.D.; De Mejía, E. Nutritional and Bioactive Compounds of Soybean: Benefits on Human Health; Soybean and Health, IntechOpen: London, UK, 2011; pp. 465–488. [Google Scholar] [CrossRef]
- Xiao, M.; Ye, J.; Tang, X.; Huang, Y. Determination of soybean isoflavones in soybean meal and fermented soybean meal by micellar electro kinetic capillary chromatography (MECC). Food Chem. 2011, 126, 1488–1492. [Google Scholar] [CrossRef]
- Flachowsky, G.; Hünerberg, M.; Meyer, U.; Kammerer, D.R.; Carle, R.; Goerke, M.; Eklund, M. Isoflavone concentration of soybean meal from various origins and transfer of isoflavones into milk of dairy cows. J. Verbr. Lebensm. 2011, 6, 449–456. [Google Scholar] [CrossRef]
- Hooper, L.; Kroon, P.A.; Rimm, E.B.; Cohn, J.S.; Harvey, I.; Le Cornu, K.A.; Hall, W.L.; Ryder, J.J.; Cassidy, A. Flavonoids, flavonoid-rich foods, and cardiovascular risk: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2008, 88, 38–50. [Google Scholar] [CrossRef]
- Bursać, M.; Krstonošić, M.A.; Miladinović, J.; Malenčić, D.; Gvozdenović, L.; Cvejić, J.H. Isoflavone composition, total phenolic content and antioxidant capacity of soybeans with colored seed coat. Nat. Prod. Commun. 2017, 12, 527–532. [Google Scholar] [CrossRef]
- Budryn, G.; Gałązka-Czarnecka, I.; Brzozowska, E.; Grzelczyk, J.; Mostowski, R.; Żyżelewicz, D.; Cerón-Carrasco, J.P.; Pérez-Sánchez, H. Evaluation of estrogenic activity of red clover (Trifoliumpratense L.) sprouts cultivated under different conditions by content of isoflavones, calorimetric study and molecular modelling. Food Chem. 2018, 245, 324–336. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, B.W.; Kim, B.; Kim, H.T.; Ko, J.M.; Baek, I.Y.; Seo, W.T.; Kang, Y.M.; Cho, K.M. Changes in phenolic compounds (isoflavones and phenolic acids) and antioxidant properties in high-protein soybean (Glycine max L. cv. Saedanbaek) for different roasting conditions. J. Korean Soc. Appl. Biol. Chem. 2013, 56, 605–612. [Google Scholar] [CrossRef]
- Barnes, S. The biochemistry, chemistry and physiology of the isoflavones in soybeans and their food products. Lymphat. Res. Biol. 2010, 8, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Kuligowski, M.; Pawłowska, K.; Jasińska-Kuligowska, I.; Nowak, J. Isoflavone composition, polyphenols content and antioxidative activity of soybean seeds during tempeh fermentation. CyTA J. Food 2017, 15, 27–33. [Google Scholar] [CrossRef]
- Baião, D.D.S.; Freitas, C.S.; Gomes, L.P.; da Silva, D.; Correa, A.C.N.; Pereira, P.R.; Del Aguila, E.M.; Paschoalin, V.M.F. Polyphenols from root, tubercles and grains cropped in Brazil: Chemical and nutritional characterization and their effects on human health and diseases. Nutrients 2017, 9, 1044. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, H.; Pihlanto, A. Food-derived bioactive peptides-opportunities for designing future foods. Curr. Pharm. Des. 2003, 9, 1297–1308. [Google Scholar] [CrossRef] [PubMed]
- Bruce, K.J.; Karr-Lilienthal, L.K.; Zinn, K.E.; Pope, L.L.; Mahan, D.C.; Fastinger, N.D.; Watts, M.; Utterback, P.L.; Parsons, C.M.; Castaneda, E.O.; Ellis, M.; et al. Evaluation of the inclusion of soybean oil and soybean processing by-products to soybean meal on nutrient composition and digestibility in swine and poultry. J. Anim. Sci. 2006, 84, 1403–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Nawaz, H.; Pohorly, J.; Mittal, G.; Kakuda, Y.; Jiang, Y. Extraction of polyphenolics from plant material for functional foods—Engineering and technology. Food Rev. Int. 2005, 21, 139–166. [Google Scholar] [CrossRef]
- Nunes, R.S.C.; de Souza, C.P.; Pereira, K.S.; Del Aguila, E.M.; Paschoalin, V.M.F. Identification and molecular phylogeny of coagulase-negative staphylococci isolates from Minas Frescal cheese in southeastern Brasil: Superantigenic toxin production and antibiotic resistance. J. Dairy Sci. 2016, 99, 2641–2653. [Google Scholar] [CrossRef]
- Pinhati, F.R.; Del Aguila, E.M.; Tôrres, A.P.R.; Sousa, M.P.D.; Santiago, V.M.J.; Silva, J.T.; Paschoalin, V.M.F. Evaluation of the efficiency of deterioration of aromatic hydrocarbons by bacteria from wastewater treatment plant of oil refinery. Quim. Nova 2014, 37, 1269–1274. [Google Scholar] [CrossRef]
- Li, R.; Hettiarachchy, N.; Rayaprolu, S.; Eswaranandam, S.; Howe, B.; Davis, M.; Jha, A. Phenolics and antioxidant activity of saskatoon berry (Amelanchieralnifolia) pomace extract. J. Med. Food 2014, 17, 384–392. [Google Scholar] [CrossRef]
- Cho, K.M.; Ha, T.J.; Lee, Y.B.; Seo, W.D.; Kim, J.Y.; Ryu, H.W.; Jeong, S.H.; Kang, Y.M.; Lee, J.H. Soluble phenolics and antioxidant properties of soybean (Glycine max L.) cultivars with varying seed coat colours. J. Funct. Foods 2013, 5, 1065–1076. [Google Scholar] [CrossRef]
- Phenol-explorer. Available online: http://phenol-explorer.eu/search?utf8=%E2%9C%93&query=soybean&button=> (accessed on 22 June 2018).
- Aguiar, C.L.; Haddad, R.; Eberlin, M.N.; Carrao-Panizzi, M.C.; Tsai, S.; Mand Park, Y.K. Thermal behavior of malonyl glucoside isoflavones in soybean flour analyzed by RPHPLC/DAD and eletrospray ionization mass spectrometry. LWT Food Sci. Technol. 2012, 48, 114–119. [Google Scholar] [CrossRef]
- Pandjaitan, N.; Hettiarachchy, N.; Ju, Z.Y.; Crandall, P.; Sneller, C.; Dombek, D. Evaluation of genistin and genistein contents in soybean varieties and soy protein concentrate prepared with 3 basic methods. J. Food Sci. 2000, 65, 399–402. [Google Scholar] [CrossRef]
- Mujić, I.; Šertović, E.; Jokić, S.; Sarić, Z.; Alibabić, V.; Vidović, S.; Živković, J. Isoflavone content and antioxidant properties of soybean seeds. Croat. J. Food Sci. Technol. 2011, 3, 16–20. [Google Scholar] [CrossRef]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Heinonen, S.M.; Wähälä, K.; Adlercreutz, H. Metabolism of isoflavones in human subjects. Phytochem. Rev. 2002, 1, 175–182. [Google Scholar] [CrossRef]
- Sánchez-Calvo, J.M.; Rodríguez-Iglesias, M.A.; Molinillo, J.M.; Macías, F.A. Soy isoflavones and their relationship with microflora: Beneficial effects on human health in equol producers. Phytochem. Rev. 2013, 12, 979–1000. [Google Scholar] [CrossRef]
- Shahidi, F.; Yeo, J.D. Insoluble-bound phenolics in food. Molecules 2016, 21, 1216. [Google Scholar] [CrossRef]
- Pratt, D.E.; Birac, P.M. Source of antioxidant activity of soybeans and soy products. J. Food Sci. 1979, 44, 1720–1722. [Google Scholar] [CrossRef]
- Dueñas, M.; Hernández, T.; Lamparski, G.; Estrella, I.; Muñoz, R. Bioactive phenolic compounds of soybean (Glycine max cv. Merit): Modifications by different microbiological fermentations. Pol. J. Food Nutr. Sci. 2012, 62, 241–250. [Google Scholar] [CrossRef]
- Saura-Calixto, F.; Serrano, J.; Goñi, I. Intake and bioaccessibility of total polyphenols in a whole diet. Food Chem. 2007, 101, 492–501. [Google Scholar] [CrossRef] [Green Version]
- Fernández-García, E.; Carvajal-Lérida, I.; Pérez-Gálvez, A. In vitro bioaccessibility assessment as a prediction tool of nutritional efficiency. Nutr. Res. 2009, 29, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Campos-Vega, R.; Vázquez-Sánchez, K.; López-Barrera, D.; Loarca-Piña, G.; Mendoza-Díaz, S.; Oomah, B.D. Simulated gastrointestinal digestion and in vitro colonic fermentation of spent coffee (Coffeaarabica L.): Bioaccessibility and intestinal permeability. Food Res. Int. 2015, 77, 156–161. [Google Scholar] [CrossRef]
- Bouayed, J.; Hoffmann, L.; Bohn, T. Total phenolics, flavonoids, anthocyanins and antioxidant activity following simulated gastro-intestinal digestion and dialysis of apple varieties: Bioaccessibility and potential uptake. Food Chem. 2011, 128, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Roque, M.J.; Rojas-Graü, M.A.; Elez-Martínez, P.; Martín-Belloso, O. Soymilk phenolic compounds, isoflavones and antioxidant activity as affected by in vitro gastrointestinal digestion. Food Chem. 2013, 136, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 93 intervention studies. Am. J. Clin. Nutr. 2005, 81, 243S–255S. [Google Scholar] [CrossRef] [PubMed]
- Ryan, L.; Prescott, S.L. Stability of the antioxidant capacity of twenty-five commercially available fruit juices subjected to an in vitro digestion. Intl. J. Food Sci. Technol. 2010, 45, 1191–1197. [Google Scholar] [CrossRef]
- Silva, F.D.O.; Perrone, D. Characterization and stability of bioactive compounds from soybean meal. LWT-Food Sci. Technol. 2015, 63, 992–1000. [Google Scholar] [CrossRef]
- Xu, B.; Chang, S.K.C. Characterization of phenolic substances and antioxidant properties of food soybeans grown in the North Dakota-Minnesota region. J. Agric. Food Chem. 2008, 56, 9102–9113. [Google Scholar] [CrossRef]
- Fernandes, M.S.; Lima, F.S.; Rodrigues, D.; Handa, C.; Guelfi, M.; Garcia, S.; Ida, E.I. Evaluation of the isoflavone and total phenolic contents of kefir-fermented soymilk storage and after the in vitro digestive system simulation. Food Chem. 2017, 229, 373–380. [Google Scholar] [CrossRef]
- Shahidi, F.; Zhong, Y. Novel antioxidants in food quality preservation and health promotion. Eur. J. Lipid Sci. Technol. 2010, 112, 930–940. [Google Scholar] [CrossRef]
- Abutheraa, R.; Hettiarachchy, N.; Kumar-Phillips, G.; Horax, R.; Chen, P.; Morawicki, R.; Kwon, Y.M. Antimicrobial activities of phenolic extracts derived from seed coats of selected soybean varieties. J. Food Sci. 2017, 82, 731–737. [Google Scholar] [CrossRef]
- Del Aguila, E.M.; Gomes, L.P.; Freitas, C.S.; Pereira, P.R.; Paschoalin, V.M.F. Natural Antimicrobials in Food Processing: Bacteriocins, Peptides and Chitooligosaccharides. Front. Anti-Infect. Drug Discov. 2017, 5, 55–108. [Google Scholar] [CrossRef]
- Lopes, J.; Damasceno, J.L.; Oliveira, P.F.; Guedes, A.P.; Tavares, D.C.; Deflon, V.M.; Lopes, N.P.; Pivatto, M.; Batista, A.A.; Maia, P.S.; et al. Ruthenium (II) Complexes containing anti-inflammatory drugs as ligands: Synthesis, characterization and in vitro cytotoxicity activities on cancer cell lines. J. Braz. Chem Soc. 2015, 26, 1838–1847. [Google Scholar] [CrossRef]
- Fonseca, N.D.; Villar, M.P.M.; Donangelo, C.M.; Perrone, D. Isoflavones and soya saponins in soy infant formulas in Brazil: Profile and estimated consumption. Food Chem. 2014, 143, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Inada, K.O.P.; Oliveira, A.A.; Revorêdo, T.B.; Martins, A.B.N.; Lacerda, E.C.Q.; Freire, A.S.; Braz, B.F.; Santelli, R.E.; Torres, A.G.; Perrone, D.; et al. Screening of the chemical composition and occurring antioxidants in jabuticaba (Myrciariajaboticaba) and jussara (Euterpe edulis) fruits and their fractions. J. Funct. Foods 2015, 17, 422–433. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- da Silva, D.V.; Silva, F.O.; Perrone, D.; Pierucci, A.P.T.R.; Conte-Junior, C.A.; Alvares, T.S.; Del Aguila, E.M.; Paschoalin, V.M.F. Physicochemical, nutritional, and sensory analyses of a nitrate-enriched beetroot gel and its effects on plasmatic nitric oxide and blood pressure. Food Nutr. Res. 2016, 60, 1–9. [Google Scholar] [CrossRef]
- Zulueta, A.; Esteve, M.J.; Frígola, A. ORAC and TEAC assays comparison to measure the antioxidant capacity of food products. Food Chem. 2009, 114, 310–316. [Google Scholar] [CrossRef]
- Samaranayaka, A.G.P.; Li-Chan, E.C.Y. Pacific hake (Merlucciusproductus) fish protein hydrolysates with antioxidative properties. Food Chem. 2008, 107, 768–776. [Google Scholar] [CrossRef]
- Oomen, A.G.; Rompelberg, C.J.M.; Bruil, M.A.; Dobbe, C.J.G.; Pereboom, D.P.K.H.; Sips, A.J.A.M. Development of an in vitro digestion model for estimating the bioaccessibility of soil contaminants. Arch. Environ. Contam. Toxicol. 2003, 44, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Sagratini, G.; Caprioli, G.; Maggi, F.; Font, G.; Giardinà, D.; Mañes, J.; Meca, G.; Ricciutelli, M.; Sirocchi, V.; Torregiani, E.; Vittori, S. Determination of soyasaponins I and βg in raw and cooked legumes by solid phase extraction (SPE) coupled to liquid chromatography (LC)–mass spectrometry (MS) and assessment of their bioaccessibility by an in vitro digestion model. J. Agric. Food Chem. 2013, 61, 1702–1709. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zheng, Y.L.; Hyde, W.; Hendrich, S.; Murphy, P.A. Human fecal metabolism of soyasaponin I. J. Agric. Food Chem. 2004, 52, 2689–2696. [Google Scholar] [CrossRef] [PubMed]
- Mosele, J.I.; Macià, A.; Romero, M.P.; Motilva, M.J.; Rubió, L. Application of in vitro gastrointestinal digestion and colonic fermentation models to pomegranate products (juice, pulp and peel extract) to study the stability and catabolism of phenolic compounds. J. Funct. Foods 2015, 14, 529–540. [Google Scholar] [CrossRef]
- McDonald, J.A.; Schroeter, K.; Fuentes, S.; Heikamp-deJong, I.; Khursigara, C.M.; de Vos, W.M.; Allen-Vercoe, E. Evaluation of microbial community reproducibility, stability and composition in a human distal gut chemostat model. J. Microbiol. Methods 2013, 95, 167–174. [Google Scholar] [CrossRef] [PubMed]
- McMillian, M.K.; Li, L.; Parker, J.B.; Patel, L.; Zhong, Z.; Gunnett, J.W.; Johnson, M.D. An improved resazurin-based cytotoxicity assay for hepatic cells. Cell Biol. Toxicol. 2002, 18, 157–173. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |



{kind=link}
{kind=link}
{kind=link}
| Gastrointestinal Digestion (mg/100 g of Dry Weight) | Colonic Fermentation (mg/100 g of Dry Weight) | ||||||
|---|---|---|---|---|---|---|---|
| Isoflavones structures | PD | OD | GD | ID | 4 h | 24 h | 48 h |
| 6-O-Acetyl | |||||||
| Acetyldaidzin | 3.0 ± 0.02 c | 3.0 ± 0.02 c | 1.3 ± 0.01 d | 1.2 ± 0.01 d | 1.8 ± 0.02 d | 10.1 ± 0.02 a | 6.8 ± 0.02 b |
| Acetylgenistin | 3.0 ± 0.01 a | 2.0 ± 0.02 b | 2.0 ± 0.01 b | 2.5 ± 0.03 b | 2.7 ± 0.01 a | 3.4 ± 0.01 a | 3.5 ± 0.02 a |
| Acetylglycitin | 0.6 ± 0.01 a | 0.3 ± 0.02 a | 0.3 ± 0.01 a | 0.2 ± 0.03 a | 0.8 ± 0.01 a | nd | nd |
| Total | 6.6 | 5.3 | 3.6 | 3.9 | 5.3 | 13.5 | 10.3 |
| Aglycones | |||||||
| Daidzein | 3.0 ± 0.01 d | 2.0 ± 0.01 e | 1.1 ± 0.01 f | 4.0 ± 0.01 d | 21.1 ± 0.05 a | 12.7 ± 0.04 b | 7.1 ± 0.01 c |
| Genistein | 5.0 ± 0.03 b | 3.0 ± 0.01 c | 2.0 ± 0.02 d | 2.2 ± 0.02 d | 11.8 ± 0.05 a | 5.8 ± 0.02 b | 4.6 ± 0.01 b |
| Glycitein | 0.3 ± 0.01 d | 0.2 ± 0.01 d | 1.0 ± 0.01 e | 1.9 ± 0.01 e | 24.7 ± 0.06 a | 15.7 ± 0.05 b | 7.1 ± 0.01 c |
| Total | 8.3 | 5.2 | 4.1 | 8.1 | 57.6 | 34.2 | 18.8 |
| 7-O-Glucoside | |||||||
| Daidzin | 35.0 ± 0.02 a | 34.0 ± 0.01 a | 33.0 ± 0.02 a | 10.0 ± 0.01 b | nd | nd | nd |
| Genistin | 49.0 ± 0.02 a | 48.0 ± 0.02 a | 48.5 ± 0.01 a | 11.0 ± 0.01 b | nd | nd | nd |
| Glycitin | 16.0 ± 0.03 a | 15.0 ± 0.01 a | 14.5 ± 0.02 b | 4.0 ± 0.03 c | nd | nd | nd |
| Total | 100.0 | 97.0 | 96.0 | 25.0 | - | - | - |
| 6-O-Malonyl | |||||||
| Malonyldaidzin | 7.0 ± 0.03 a | 6.0 ± 0.03 a | 3.0 ± 0.001 b | 1.0 ± 0.03 c | nd | nd | nd |
| Malonylgenistin | 13.0 ± 0.01 b | 16.0 ± 0.02 a | 5.0 ± 0.001 c | 2.0 ± 0.04 d | nd | nd | nd |
| Malonylglycitin | 4.0 ± 0.01 a | 3.0 ± 0.03 a | 1.0 ± 0.001 b | 0.3 ± 0.03 c | nd | nd | nd |
| Total | 24.0 | 25.0 | 9.0 | 3.3 | - | - | - |
| Total Isoflavones | 138.9 | 132.5 | 112.7 | 40.3 | 62.9 | 47.7 | 29.1 |
| Phenolic Compounds | PD | OD | GD | ID |
|---|---|---|---|---|
| mg/100 g of Dry Weight | ||||
| Hydroxybenzoics | ||||
| Salicylic acid | 3.8 ± 0.01 a | 2.0 ± 0.2 b | 2.0. ± 0.01 b | 2.1 ± 0.2 b |
| Caffeic acid | 6.1 ± 0.1 a | 2.3 ± 0.2 d | 3.0 ± 0.2 c | 3.8 ± 0.3 b |
| Ferulic acid | 0.3 ± 0.02 b | 0.9 ± 0.01 a | 0.9 ± 0.01 a | 1.0 ± 0.02 a |
| Gallic acid | 7.7 ± 0.1 a | 1.1 ± 0.0 d | 2.9 ± 1.3 c | 4.8 ± 0.30 b |
| Syringic acid | 8.1 ± 0.1 a | 1.1 ± 0.01 d | 2.0 ± 0.3 c | 4.3 ± 0.3 b |
| Vanillic acid | 9.1 ± 0.2 a | 1.3 ± 0.02 d | 3.6 ± 0.6 c | 6.2 ± 1.6 b |
| 4-hydroxybenzoic acid | 5.1 ± 0.1 a | 1.2 ± 0.1 c | 3.1 ± 0.9 b | 1.7 ± 0.4 c |
| Total | 40.2 | 9.9 | 17.5 | 23.9 |
| Hydroxycinnamics | ||||
| 5-caffeoylquinic chlorogenic acid | 3.5 ± 0.2 a | 1.0 ± 0.1 c | 2.4 ± 0.3 b | 1.3 ± 0.1 c |
| p-coumaric acid | 2.0 ± 0.1 a | 1.0 ± 0.01 b | 1.1 ± 0.1 b | 1.2 ± 0.1 b |
| 4-hydroxyphenylacetic acid | 3.4 ± 0.1 a | 1.0 ± 0.1 c | 1.6 ± 0.1 b | 1.0 ± 0.1 c |
| Sinapic acid | 2.7 ± 0.1 a | 1.0 ± 0.1 b | 1.1 ± 0.1 b | 1.0 ± 0.1 b |
| Total | 11.6 | 4.0 | 6.2 | 4.5 |
| Flavonones and Flavonols | ||||
| Hesperidin | 9.1 ± 0.4 a | 1.2 ± 0.2 d | 7.1 ± 0.6 b | 4.2 ± 0.6 c |
| Kaempferol | 0.4 ± 0.01 a | 0.9 ± 0.1 a | 0.9 ± 0.1 a | 0.9 ± 0.1 a |
| Myricetin | 5.0 ± 0.01 a | 1.3 ± 0.1 c | 1.7 ± 0.1 c | 3.2 ± 0.6 b |
| Naringin | 2.5 ± 0.5 a | 0.9 ± 0.1 c | 1.0 ± 0.1 c | 1.6 ± 0.8 b |
| Rutin | 4.9 ± 0.1 a | 1.8 ± 0.1 c | 2.1 ± 0.9 b | 2.6 ± 0.1 b |
| Total | 21.9 | 6.1 | 12.8 | 12.5 |
| Total phenolics | 73.7 | 20.0 | 36.5 | 40.9 |
| Microorganisms | Soybean Meal Aqueous Extract Concentration (mg/mL) | Inhibition (%) |
|---|---|---|
| Gram-positive | ||
| Listeria innocua ATCC 33090 | 150 | 21 ± 2.1 |
| coagulase-negative Staphylococcus saprophyticus KT955005 * | 75 | 100 ± 0.0 |
| Staphylococcus aureus ATCC 14458 | 120 | 100 ± 0.0 |
| Gram-negative | ||
| Acinetobacter genomospecies 3 ** | 12.5 | 100 ± 0.0 |
| Aeromonas hydrophila FDA110-36 | 90 | 100 ± 0.0 |
| A. hydrophila ATCC 7966 | 50 | 100 ± 0.0 |
| Escherichia coli DH5 alfa | 35 | 100 ± 0.0 |
| E. coli ATCC 43895 | 100 | 100 ± 0.0 |
| Salmonella enterica ATCC 12325 | 75 | 100 ± 0.0 |
| S. enterica ATCC 29934 | 75 | 100 ± 0.0 |
| Vibrio parahemolyticus ATCC 17802 | 100 | 100 ± 0.0 |
| Pseudomonas fluorescens ATCC 13525 | 150 | 100 ± 0.0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
S. Freitas, C.; Alves da Silva, G.; Perrone, D.; A. Vericimo, M.; Dos S. Baião, D.; R. Pereira, P.; M. F. Paschoalin, V.; M. Del Aguila, E. Recovery of Antimicrobials and Bioaccessible Isoflavones and Phenolics from Soybean (Glycine max) Meal by Aqueous Extraction. Molecules 2019, 24, 74. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24010074
S. Freitas C, Alves da Silva G, Perrone D, A. Vericimo M, Dos S. Baião D, R. Pereira P, M. F. Paschoalin V, M. Del Aguila E. Recovery of Antimicrobials and Bioaccessible Isoflavones and Phenolics from Soybean (Glycine max) Meal by Aqueous Extraction. Molecules. 2019; 24(1):74. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24010074
Chicago/Turabian StyleS. Freitas, Cyntia, Genilton Alves da Silva, Daniel Perrone, Mauricio A. Vericimo, Diego Dos S. Baião, Patrícia R. Pereira, Vânia M. F. Paschoalin, and Eduardo M. Del Aguila. 2019. "Recovery of Antimicrobials and Bioaccessible Isoflavones and Phenolics from Soybean (Glycine max) Meal by Aqueous Extraction" Molecules 24, no. 1: 74. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24010074