
Distinctive Formation of Volatile Compounds in Fermented Rice Inoculated by Different Molds, Yeasts, and Lactic Acid Bacteria


Department of Food Science and Engineering, Ewha Womans University, Seoul 03760, Korea
*
Author to whom correspondence should be addressed.
Molecules 2019, 24(11), 2123; https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24112123
Submission received: 29 April 2019
/
Revised: 28 May 2019
/
Accepted: 4 June 2019
/
Published: 5 June 2019
(This article belongs to the Collection Recent Advances in Flavors and Fragrances)
Abstract
:Rice has been fermented to enhance its application in some foods. Although various microbes are involved in rice fermentation, their roles in the formation of volatile compounds, which are important to the characteristics of fermented rice, are not clear. In this study, diverse approaches, such as partial least squares-discriminant analysis (PLS-DA), metabolic pathway-based volatile compound formations, and correlation analysis between volatile compounds and microbes were applied to compare metabolic characteristics according to each microbe and determine microbe-specific metabolites in fermented rice inoculated by molds, yeasts, and lactic acid bacteria. Metabolic changes were relatively more activated in fermented rice inoculated by molds compared to other microbes. Volatile compound profiles were significantly changed depending on each microbe as well as the group of microbes. Regarding some metabolic pathways, such as carbohydrates, amino acids, and fatty acids, it could be observed that certain formation pathways of volatile compounds were closely linked with the type of microbes. Also, some volatile compounds were strongly correlated to specific microbes; for example, branched-chain volatiles were closely link to Aspergillus oryzae, while Lactobacillus plantarum had strong relationship with acetic acid in fermented rice. This study can provide an insight into the effects of fermentative microbes on the formation of volatile compounds in rice fermentation.
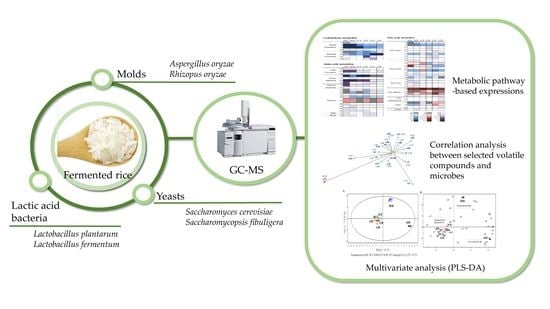
1. Introduction
Fermented rice products, such as alcoholic beverages (makgeolli, sake, and Chinese rice wine), fermented soybean pastes (deonjang and miso), and rice vinegar [1,2,3], have been consumed mainly due to their characteristic odors and tastes. During rice fermentation, fermentative microbes degrade macromolecules of food substrates (e.g., proteins, carbohydrates, and lipids) via diverse enzymatic metabolic pathways, providing a precursor pool of amino acids, sugars, and fatty acids that can participate in further biological and chemical reactions, so as to produce volatile compounds related to the characteristic tastes and odors [4]. Regarding previous studies on the main volatile compounds in fermented rice, Xiao et al. [5] reported the major volatile compounds in Chinese rice vinegars were furfural, acetic acid, ethyl acetate, 3-hydroxy-2-butanone, 3-methyl-1-butanol, isopentyl acetate, bezaldehyde, and phenylethyl alcohol. Son et al. [6] also compared the volatile compounds profiles in makgeolli fermented by Asepergillus oryzae and Saccharomycopsis fibuligera. Fusel alcohols, acetate esters, and ethyl esters were considered as the main volatile compounds of makgeolli.
On the other hand, the enzymes of microbes are specific to perform specialized catalytic reactions, forming different metabolites according to the type of microbes despite being exposed to identical culture conditions [7]. Diverse microbes (e.g., Aspergillus spp., Saccharomyces cerevisiae, non-Saccharomyces yeasts, and Lactobacillus spp.) have been commonly used as microbial starters in fermented rice products, such as makgeolli and rice vinegar [8,9]. In particular, molds such as Aspergillus spp. and Rhizopus spp. are preferentially used in rice fermentation and represented as an initial fermentative microbe due to their higher hydrolytic enzymes, such as amylase, protease, and glucoamylase [10,11]. On the other hand, yeasts, especially, Saccharomyces cerevisiae, are known for generating some volatile compounds, such as fusel alcohols and esters, in wine making, baking, and brewing [12]. In addition, lactic acid bacteria are also known as the characteristic fermentative microbes for producing organic acids, including acetic acid and lactic acid, so that acidification occurs in fermented foods [13]. The fermentative microbes mentioned above can also significantly affect the generation of volatile compounds in fermented rice. Thus, in order to investigate the quality of fermented rice, studies on the formation of characteristic volatile compounds based on microbial species should be carried out.
Although some studies on the formation of volatile compounds according to various microbes have been performed in rice fermentation [6,14], there is little research on the comprehensive determination of specific volatile compounds related to the range of microbes which are responsible for various rice fermentations. In this study, some approaches, such as partial least squares-discriminant analysis (PLS-DA) and metabolic pathway-based and correlation analysis, were applied to identify the characteristic patterns of volatile compound profiles distinguished from other species in fermented rice. The aim of this study was to compare the profiles of induced volatile compounds in fermented rice inoculated by diverse molds, yeasts, and lactic acid bacteria according to fermentative microbes. Solid phase microextraction-gas chromatography-mass spectrometry (SPME-GC-MS) was applied to obtain the volatile compound profiles of steamed and non-inoculated (control) or fermented rice. The mass spectrometry-based data sets were combined with correlation analysis to identify the differences in volatile metabolite profiles according to each microbe and to determine the microbe-specific volatile compounds in rice fermentation.
2. Results and Discussion
2.1. PLS-DA for Volatile Metabolite Profiles in Fermented Rice
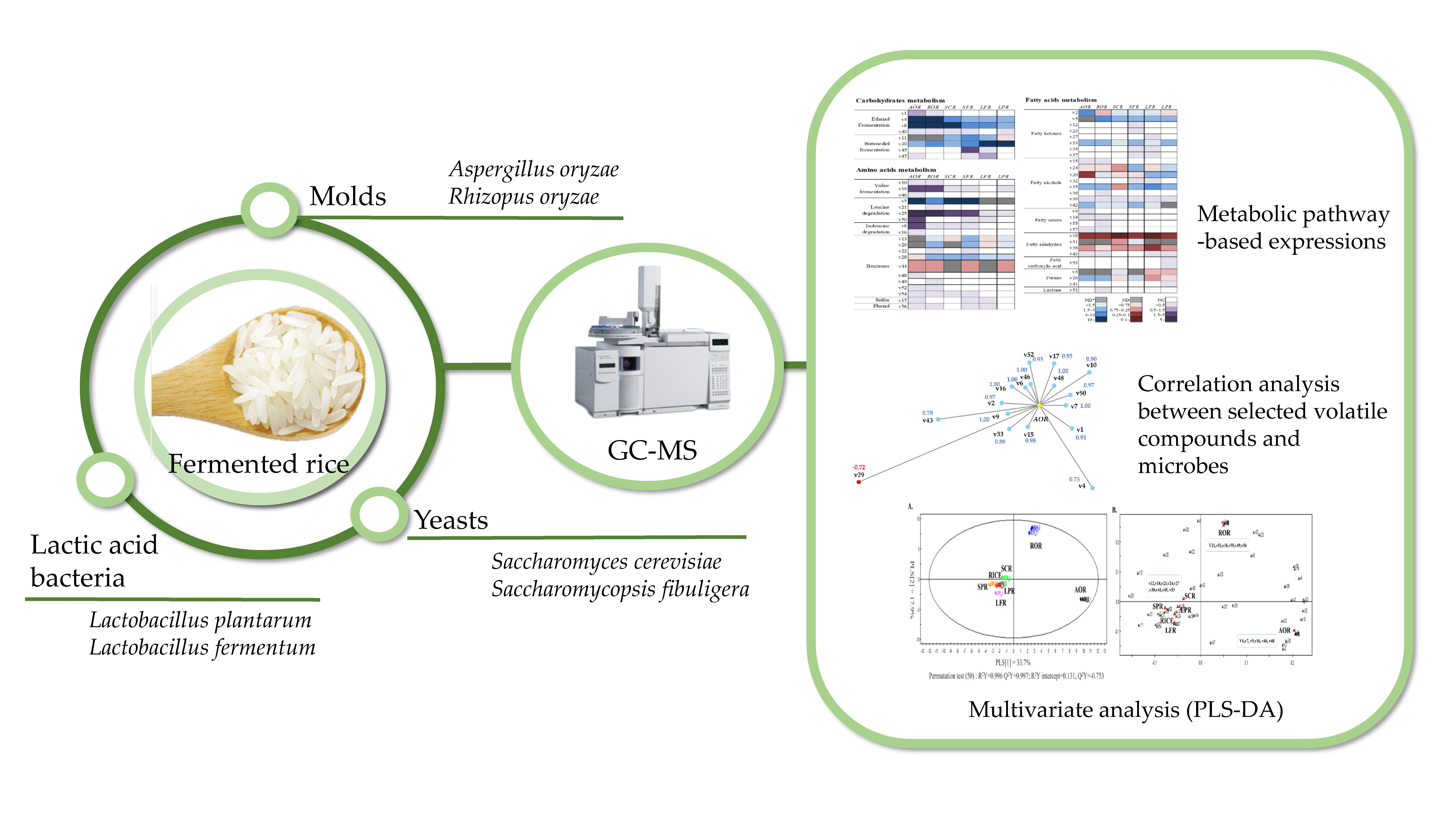
PLS-DA was performed to evaluate the differences among volatile compound profiles obtained from GC-MS data for different fermentation microbes (molds, yeasts, and lactic acid bacteria). The classification of different fermented samples and controls (RICE, non-inoculated and steamed rice), and also discriminant volatile compounds, are shown in Figure 1. The PLS-DA model explained 57.2% of total variance, such as 34.6% PLS [1] and 22.6% PLS [2]. The model was robust and predictable. The parameters of the cross-validation modeling for the third PLS component were R2Y = 0.996, and Q2Y = 0.997. Permutation testing (50 iterations) was also performed in order to further validate the model, after which R2 = 0.131 and Q2 = −0.753.
In a score plot of PLS-DA, AOR (fermented by Aspergillus oryzae) and ROR (Rhyzopus oryzae) were clearly separated from SCR (Sacharomyces cerevisiae), SFR (Saccharomycopsis fibuligera), LFR (Lactobacillus fermentum), LPR (Lactobacillus plantarum), and control sample (RICE, non-fermented rice), and SCR, SFR, LFR, and LPR were closely placed near RICE. It can be assumed that A. oryzae and R. oryzae were more active in the changes of volatile compounds during rice fermentation than other microbes.
Table 1 lists the distinct volatile compounds from each microbe. A total of 21 significant variables contributing to discriminate samples (p-value >0.1 or <−0.1) were selected on the basis of a threshold of 1.0 on the variable importance in the projection (VIP) test. Some volatile metabolites derived from amino acids, such as 2-methylbutanal, 3-methylbutanal, ethyl 2-methylbutanoate, 2-methylpropanoic acid, and 2-phenylacetaldehyde, were highly correlated to AOR, while 3-methylbutyl acetate, 5-ethyloxolan-2-one, 3-ethoxypropan-1-ol, ethyl tetradecanoate, 1-phenylethanone, and 2-phenylethanol, which are mainly derived from amino acids and fatty acids, were representative of ROR. In addition, volatile metabolites derived from fatty acids and butanediol fermentation were major discriminant volatile compounds in other samples, such as RICE, SCR, SFR, LFR, and LPR.
Table 2 shows the identified volatile compounds with their relative peak areas and retention indices (RI). A total of 57 volatile compounds, such as eight carbohydrate-derived, 20 amino acid-derived, and 29 fatty acid-derived volatile compounds, were found in all samples.
During rice fermentation, the generation of volatile compounds varied according to each microbe in this study. In addition, we observed that the total contents of volatile compounds were much higher in AOR and ROR compared to other samples. The major volatile compounds generated by AOR were ethanol, 3-methylbutan-1-ol, 3-methylbutanal, and 2-methylbutanal, while 3-methylbutan-1-ol, ethanol, and 2-methylpropan-1-ol were the major volatile compounds in ROR. In addition, the contents of ethanol and 3-methylbutan-1-ol were higher than those of other volatile compounds in SCR, while butan-2,3-diol, butane-2,3-dione, and 3-methyl-butan-1-ol were the predominant volatile compounds in SFR. The level of 3-hydroxy-2-butanone was the most abundant in LFR. On the other hand, acetic acid and 3-hydroxy-2-butanone were the main metabolites of LPR, but overall formation of volatile compounds was weakly generated compared to other fermented samples. These results indicated that certain volatile compounds were generated at high levels, specific to each microbe. For example, the level of 3-methylbutan-1-ol in mold and yeast fermentation (AOR, ROR, SCR, and SFR) was higher, while that of 3-hydroxybutan-2-one was superior in lactic acid bacteria fermentation (LFR and LPR). In the following section, we compare the formation of volatile compounds based on the major metabolic processes, such as carbohydrate, amino acid, and fatty acid metabolism, to determine the metabolic characteristics of each microbe.
2.2. Volatile Compounds Based on Metabolic Pathways in Fermented Rice According to Each Microbe
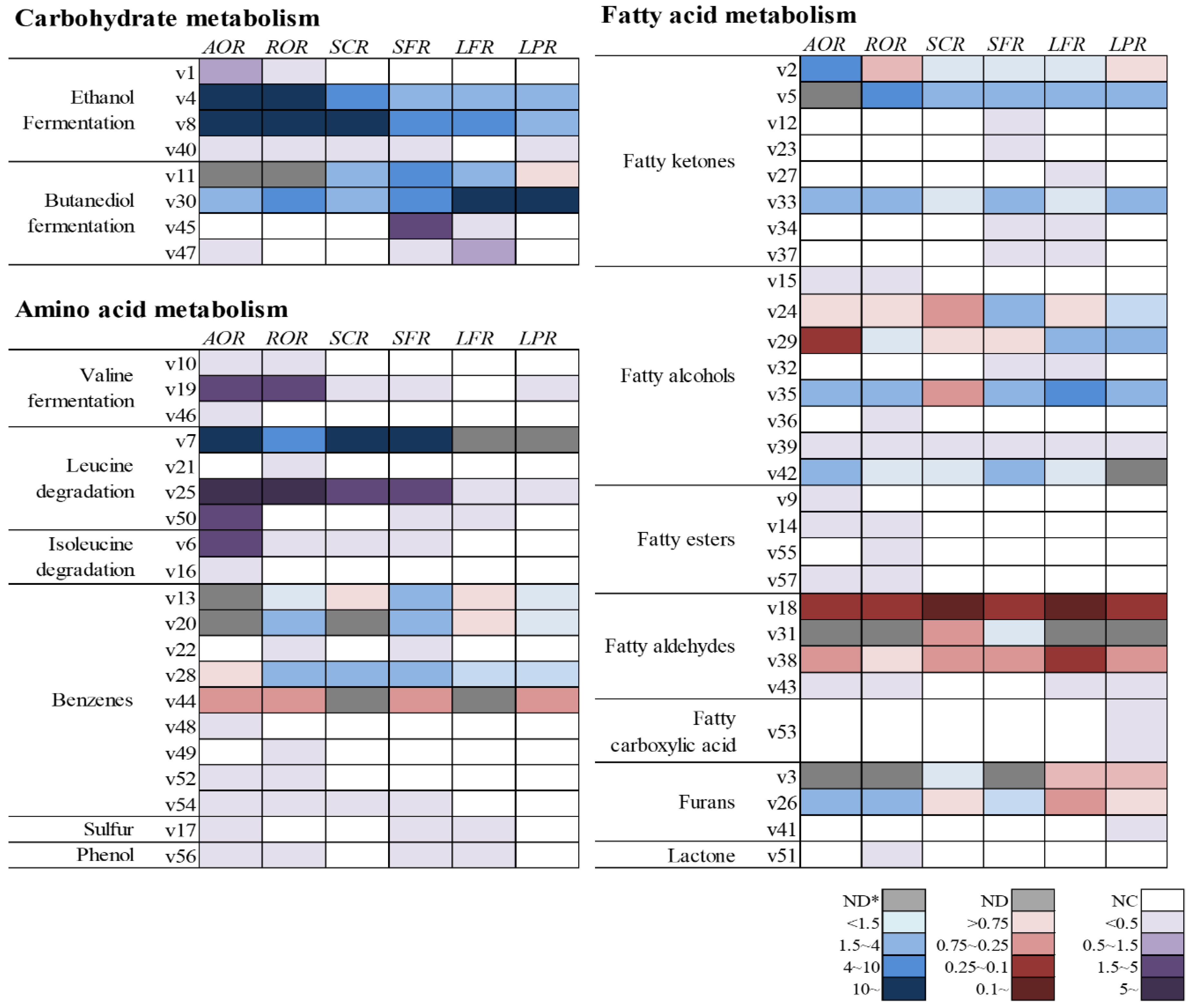
The formation of volatile compounds in fermented rice are visually represented by heatmaps in Figure 2. The fold changes of volatile compounds, which were calculated compared to the control (RICE), were classified by three main metabolic pathways, including carbohydrate, amino acid, and fatty acid metabolism (Table S1). Heatmaps were clustered by defined pathways provided from kyoto encyclopedia of genes and genomes (KEGG) pathways and previous studies [15,16,17]. The tendency to increase or decrease compared to the control was shown in five stages (blue; positive, red; negative, and gray; not detected), while the formation of volatile compounds which were not detected in the control but were newly produced during fermentation, were marked separately (purple; newly detected, white; not detected).
As shown in Figure 2, the heatmap involved in carbohydrate metabolism was more activated compared to other forms of metabolism during rice fermentation. Carbohydrates were closely linked to the formation of some volatile compounds, such as ethanol fermentation and butanediol fermentation. Ethanol originates from fermentative breakdown of glucose and other hexose via the Embden–Meyerhof–Parnas (EMP) pathway and some other pathways with decarboxylase and dehydrogenase [18]. By-products of ethanol fermentation, including acetic acid, ethyl acetate, and acetaldehyde, were found in this study. In particular, acetaldehyde and acetic acid were detected only in the fermented samples, with the exception of RICE (non-fermented). The increase of acetaldehyde was higher in AOR, while that of acetic acid was superior in LPR compared to other samples. On the other hand, the overall by-products of ethanol fermentation were more actively generated in AOR, ROR, and SCR. In particular, ethanol and ethyl acetate were generated at amounts approximately 200- and 25-fold higher, respectively, than those in the control.
The three-stage butanediol fermentation pathway begins with the combination of two molecules of pyruvate from glycolysis to form α-acetolactate by α-acetolactate synthase (α-ALS). α-Acetolactate is then decarboxylated to butane-2,3-dione (diacetyl) and 3-hydroxybutan-2-one (acetoin) by α-acetolactate decarboxylase (α-ALD) and, finally, 2,3-butanediol can be formed by the reduction of acetoin by 2,3-butanediol dehydrogenase (BDH) [19]. In this study, AOR, ROR, LFR, and LPR showed a higher generation of acetoin than other metabolites. In particular, lactic acid bacteria such as LFS and LPS were strongly associated with a higher production of acetoin, consistent with the results of a previous study [20]. On the other hand, the levels of diacetyl and acetoin in SCS and SPS were similar, while the formation of butan-2,3-diol was significantly increased in SPS.
Amino acid metabolism can be involved in the generation of some volatile compounds, such as sulfur-containing volatiles, branched-chain volatiles, and aromatic volatiles [21,22]. They come from the degradation of specific amino acids, such as sulfur-containing (methionine and cysteine), branched-chain (leucine, valine, and isoleucine), and aromatic ones (phenylalanine, tyrosine, and tryptophan). In this study, branched-chain volatiles, except for 3-methylbutan-1-ol, were detected only in the fermented samples. In particular, levels of 2-methylbutanal, 3-methylbutanal, 3-methylbutan-1-ol, and 3-methylbutanoic acid were greatly increased in AOR, while much lower levels of these compounds were produced by LPR and LFR compared to other samples. This result might be due to the absence of certain enzymes, such as transaminase, decarboxylase, and hydrogenase, or weaker activities of these enzymes in lactic acid bacteria than what is found in other microbes.
With regard to products of benzenes, the fold change of benzaldehyde was decreased or not found in all samples studied. It could be assumed that benzaldehyde can be converted to other benzenes, such as styrene, 2-phenylethanol, ethyl 2-phenylacetate, 1-phenylethanone, toluene, and ethylbenzene. In particular, 2-phenylethanol with a rose-like odor is commonly found in alcoholic beverages, such as makgeolli and sake [6,23]. 2-Phenylethanol can be converted to 2-phenylacetaldehyde and ethyl 2-phenylacetate by phenylacetaldehyde reductase and alcohol acetyltransferase, respectively [24]. In this study, the levels of 2-phenylacetaldehyde were greater in AOR than those of benzoic compounds, while that of 2-phenylethanol was higher in ROR, SCR and SFR. Regarding other benzoic volatile compounds, 4-ethenyl-2-methoxyphenol, 1-phenylethanone and styrene can be converted from cinnamic acid which is derived from phenylalanine degradation [25].
A compound called 4-ethenyl-2-methoxyphenol, which is known as the main volatile compound of rice and has a smoky odor, was the only phenolic volatile compound found in this study [26]. The formation of 4-ethenyl-2-methoxyphenol is characterized by a two-step process. Firstly, cinnmic acid is converted to hydroxycinnamic acid by esterase. Then, cinnamate decarboxylase catalyzes its transformation to 4-ethenyl-2-methoxyphenol [27]. In ROS, the formation of these benzoic volatiles was more activated compared to other samples, while that of styrene was decreased and 1-phenylethanone was not detected in AOR. The only sulfur-containing volatile compound in this study, (methyldisulfanyl)methane, could be produced from methionine. Methionine is degraded to methanethiol by methionine-γ-lyase, and then oxidized to (methyldisulfanyl)methane [22]. This compound, which had been reported as an off-odorant in yakju (Korean rice liquor) [28], was detected only in AOR, SFR, and LFR in this study.
During fermentation, lipids can be hydrolyzed to glycerol and fatty acids by lipase. The fatty acid metabolic pathway includes both fatty acids and their derivatives. There are various volatile compounds derived from fatty acids, including furans, lactones, and volatile compounds derived from fatty acids (aldehydes, acids, ketones, and esters). β-Oxidation, which is a main fatty acid breakdown pathway, converts fatty acids into two carbon segments (acetyl-CoA), then, further enzymatic activities, such as decarboxylation, oxidation, reduction, and so on, catalyze the formation of aldehydes, alcohols, ketones, acids, and esters [29]. Esters derived from fatty acids, such as ethyl propanoante, ethyl butanoate, ethyl tetradecanoate, and ethyl hexadecanoate, were only detected in AOR and ROR. These volatile compounds can be formed by esterification, combining fatty acids and ethanol, which is catalyzed by esterase [29]. A marked content of ethanol was observed in AOR, ROR, and SCR, while fatty acid esters were not found in SCR. It might be assumed that the different activity of esterase of the individual microbes could affect the formation of fatty acid esters. In addition, higher numbers of ketones were detected in SFR than in other samples in this study.
Lactones are mainly produced from unsaturated fatty acids, such as linoleic acid and oleic acid [30], and significantly contribute to the aroma of fermented rice products, such as rice beer and Chinese rice wine, as fruity and floral odor notes [31,32]. An important odor component of peach [33], 5-ethyloxolan-2-one (γ-hexalactone), was only detected in ROR.
2.3. Microbe-Specific Volatile Compounds in Fermented Rice
Fermented rice is enjoyed due to its characteristic aroma and taste [34] as well as its health benefits [35], which are not present in raw rice. In this study, a correlation analysis was performed to determine the microbe-specific volatile compounds involved in the flavor properties of fermented rice. Volatile compounds related to each microbe were selected using a threshold of 0.7 coefficient (r) (p-value < 0.05) (Table 3).
Relatively more diverse volatile compounds were linked to AOR compared to other samples. In particular, branched-chain volatiles, such as ethyl 2-methylbutanonoate, 2-methylpropanoic acid, 2-methylbutanal, 3-methylbutanal, 3-methylbutanoic acid, and ethyl 2-methylpropanoate, were more highly associated with AOR compared to other microbes. It can be explained that the activities of certain enzymes, such as protease, branched-chain amino acid aminotransferase, and so on, in A. oryzae were higher than in other microbes.
On the other hand, some volatile compounds, such as esters derived from fatty acids (ethyl tetradecanoate and ethyl hexadecanoate) and lactone (5-ethyloxolan-2-one), were closely related with ROR. R. oryzae might have superior activities of some enzymes related to the formation of esters and lactones, such as lipase, esterase, lactonizing enzymes, and so on.
Regarding yeast fermentation, SFR was linked to a greater number of distinctive volatile compounds that can be distinguished from other fermented rice compared to SCR. Some ketones derived from fatty acids (3-methylpentan-2-one and 5-methylhexan-2-one) and volatiles derived from butanediol fermentation (diacetyl and butane-2,3-diol) were highly involved in SFR, while SCR was linked to only oxolan.
In this study, the content of ethanol induced by S. cerevisiae, which is the most commonly employed yeast for ethanol production [36], was highest, but other microbes, such as A. oryzae and R. oryzae, could also contribute to the formation of ethanol.
Some volatile compounds from lactic acid bacteria fermentation, such as acetoin (a yogurt-like odorant) and 4-ethenyl-2-methoxyphenol (a smoky odorant), were closely linked with LFR, while acids, such as hexanoic acid and acetic acid, were positively associated with LPR.
3. Material and Methods
3.1. Chemicals and Reagents
HPLC-grade methanol and water were obtained from J. T. Baker (Phillipsburg, NJ, USA) and sodium chloride was purchased from Sigma-Aldrich (St. Louis, MO, USA).
3.2. Sample Preparation
Fermentative microbes isolated from fermented foods, such as Aspergillus oryzae KACC46102 (AOR), Rhizopus oryzae KACC40242 (ROR), Saccharomyces cerevisiae KACC30008 (SCR), Saccharomycopsis fibuligera KACC47772 (SFR), Lactobacillus fermentum KACC 15,736 (LFR), and Lactobacillus plantarum KACC 124,401 (LPR), were supported from the Korean Agricultural Culture Collection (KACC, Jeonju-si, Jeollabuk-do, Korea).
Rice (Oryza sativa L.) was soaked in water for 16 h and cooked at 121 °C for 15 min before inoculating with 1 × 105 colony forming units (CFU)/g of microorganisms. Then, they were incubated at 30 °C for 48 h. They were kept in a deep freezer (below −70 °C) before analysis.
3.3. Volatile Compounds Analysis
The samples were kept in liquid nitrogen for 1 min, before grinding for 60 s using a miller (7011S, Waring Commercial, Stamford, CT, USA). Samples (2.5 g) with 5 mL saturated sodium chloride solution were put into a screw vial with screw cap (Ultraclean 18 mm, Agilent technologies, Santa Clara, CA, USA). Then, 1 μL of an internal standard (L-borneol (Sigma-Aldrich) 100 μg/mL, in methanol) was added into the vial for quantifying the volatile metabolites. Solid phase microextraction (SPME) was applied to extract volatile compounds in this study. It was maintained at 30 °C for 30 min for an equilibrium state. Then, volatile compounds were adsorbed for 30 min onto divinylbenzene/carboxen/polydimethylsiloxane (DVB/CAR/PDMS, 75 μm, Supelco, Bellefonte, PA, USA) coated SPME fiber that was inserted 20 mm into the sealed vial.
Gas chromatography (GC-MS) analysis was performed using a 7890A series gas chromatograph (Agilent Technologies), an CTC-PAL auto sampler (GC sampler 80, Agilent Technologies), and a 5975C mass selective detector (Agilent Technologies) equipped with a DB-WAX column (30 m length × 0.25 mm i.d. × 25 μm film thickness, J&W Scientific, Folsom, CA, USA). Oven temperature was initially held at 40 °C for 6 min and ramped to 82 °C at a rate of 6 °C/min, and then raised to 200 °C (1 min) at 4 °C/min. Helium, a carrier gas, constantly flowed at 0.8 mL/min. Injector and detector temperatures were 230 °C and 250 °C, respectively. The mass spectral data were obtained at 70 eV in electron ionization (EI), with a mass scan range of 35–350 amu at a rate of 4.5 scans/s. The identification of metabolites was positively confirmed by comparing mass spectral data and retention times to those of authentic standard compounds. Otherwise, they were identified with the basis of their mass spectral database (NIST08 and Wiley9n.1) and retention index (RI) values using the national institute of standards and technology (NIST) Chemistry Webbook. The contents of volatile compounds were calculated by comparing their peak areas to those of an internal standard compound.
3.4. Statistical Analysis
Analysis of variance (ANOVA) and correlation analysis were accomplished using IMB SPSS Statistics for Windows, version 25.0 (IBM Corp., Armonk, NY, USA) to evaluate statistical change and difference of volatile compound formations in each sample. The result of Duncan’s multi-range test was presented at the significant different level (p ≤ 0.05). A partial least squares-discriminant analysis (PLS-DA) was performed to discriminate samples based on the volatile compound profiles using SIMCA-P software (version 11.0, Umetrics, Umea, Sweden).
4. Conclusions
We compared the volatile compound profiles and the microbes that produced these compounds during rice fermentation. Diverse approaches, such as multivariable statistical analysis (PLS-DA), comparative analysis based on main pathways related to generate volatile compounds, and correlation analysis between volatile compounds and each microbe, were undertaken to find the key volatile compounds for discrimination of fermented rice samples inoculated by different microbes. In this study, 46 volatile compounds were classified by metabolisms strongly involved in the formation of volatile compounds, regardless of the type of microbes. Some findings corresponded to those of previous studies. For example, S. cerevisiae is well known as a higher producer of ethanol and fusel alcohols. In addition, 2-methylbutanal and 3-methylbutanal are major fermentative compounds, induced from fungi, such as A. oryzae [16], while lactic acid bacteria are superior for generating organic acids, including acetic acid, and butanediol fermentation [37]. However, we were able to investigate the characteristic features of volatile compound profiles in rice fermentation according to each microbe. The new findings that were revealed in this study showed that A. oryzae was highly linked with the formation of volatiles derived from amino acids, while R. oryzae was superior for generating volatiles derived from fatty acids. In addition, ester formation was closely linked with R. oryzae as well as yeasts, which are typical microbial starters for the production of esters [38]. In conclusion, these results can help in the rational selection of fermentative microbial starters in fermented rice to obtain the proper sensory qualities.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/1420-3049/24/11/2123/s1, Table S1: The fold changes of volatile compounds in fermented rice samples (purple boxes indicate newly detected volatile compounds).
Author Contributions
Conceptualization, Y.-S.K. and M.K.P.; Methodology and formal analysis, M.K.P.; Data curation, M.K.P.; Writing—Original Draft Preparation, M.K.P.; Writing—Review and Editing, Y.-S.K. and M.K.P.; Visualization, M.K.P.; Supervision, Y.-S.K.
Funding
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (NRF-2017R1A2B4002233) and the High Value-added Food Technology Development Program (grant number 317035-03), funded by the Ministry of Agriculture, Food and Rural Affairs (MAFRA).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lee, S.-W.; Yoon, S.-R.; Kim, G.-R.; Woo, S.-M.; Jeong, Y.-J.; Yeo, S.-H.; Kim, K.-S.; Kwon, J.-H. Effect of nuruk and fermentation method on organic acid and volatile compounds in brown rice vinegar. Food Sci. Biotechnol. 2012, 21, 453–460. [Google Scholar] [CrossRef]
- Blandino, A.; Al-Aseeri, M.E.; Pandiella, S.S.; Cantero, D.; Webb, C. Cereal-based fermented foods and beverages. Food Res. Int. 2003, 36, 527–543. [Google Scholar] [CrossRef]
- Kum, S.J.; Yang, S.O.; Lee, S.M.; Chang, P.S.; Choi, Y.H.; Lee, J.J.; Hurh, B.S.; Kim, Y. Effects of Aspergillus species inoculation and their enzymatic activities on the formation of volatile components in fermented soybean paste (doenjang). J. Agric. Food Chem. 2015, 63, 1401–1418. [Google Scholar] [CrossRef] [PubMed]
- Park, M.K.; Cho, I.H.; Lee, S.; Choi, H.-K.; Kwon, D.-Y.; Kim, Y.-S. Metabolite profiling of cheonggukjang, a fermented soybeanpaste, during fermentation by gas chromatography-mass spectrometry and principal component analysis. Food Chem. 2010, 122, 1313–1319. [Google Scholar] [CrossRef]
- Xiao, Z.; Dai, S.; Niu, Y.; Yu, H.; Zhu, J.; Tian, H.; Gu, Y. Discrimination of Chinese vinegars based on headspace solid-phase microextraction-gas chromatography mass spectrometry of volatile compounds and multivariate analysis. J. Food Sci. 2011, 76, C1125–C1135. [Google Scholar] [CrossRef]
- Son, E.Y.; Lee, S.M.; Kim, M.J.; Seo, J.A.; Kim, Y.-S. Comparison of volatile and non-volatile metabolites in rice wine fermented by Koji inoculated with Saccharomycopsis fibuligera and Aspergillus oryzae. Food Res. Int. 2018, 109, 596–605. [Google Scholar] [CrossRef]
- Park, M.K.; Choi, H.-S.; Kim, Y.-S.; Cho, I.H. Change in profiles of volatile compounds from two types of Fagopyrum Esculentum (buckwheat) soksungjang during fermentation. Food Sci. Biotechnol. 2017, 26, 871–882. [Google Scholar] [CrossRef]
- Min, J.-H.; Kim, Y.-H.; Kim, J.H.; Choi, S.; Lee, J.-S.; Kim, H.-K. Comparison of microbial diversity of Korean commercial makgeolli showing high β-glucan content and high antihypertensive activity, respectively. Mycobiology 2012, 40, 138–141. [Google Scholar] [CrossRef]
- Haruta, S.; Ueno, S.; Egawa, I.; Hashiguchi, K.; Fujii, A.; Nagano, M.; Ishii, M.; Igarashi, Y. Succession of bacterial and fungal communities during a traditional pot fermentation of rice vinegar assessed by PCR-mediated denaturing gradient gel electrophoresis. Int. J. Food Microbiol. 2006, 109, 79–87. [Google Scholar] [CrossRef]
- Stark, D.; Münch, T.; Sonnleitner, B.; Marison, I.W.; von Stockar, U. Extractive bioconversion of 2-phenylethanol from l-phenylalanine by Saccharomyces cerevisiae. Biotechnol. Prog. 2002, 18, 514–523. [Google Scholar] [CrossRef]
- Kumar, S.; Sharma, N.S.; Saharan, M.R.; Singh, R. Extracellular acid protease from Rhizopus oryzae: Purification and characterization. Process Biochem. 2005, 40, 1701–1705. [Google Scholar] [CrossRef]
- Hazelwood, L.A.; Daran, J.M.; van Maris, A.J.; Pronk, J.T.; Dickinson, J.R. The Ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [PubMed]
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Kang, H.; Hwang, H.; Lee, J.E.; Kim, H.R. Quantitative analysis of volatile flavor components in Korean alcoholic beverage and Japanese sake using SPME-GC/MS. Food Sci. Biotechnol. 2016, 25, 979–985. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.E.; Lee, S.M.; Jang, E.S.; Shin, H.W.; Moon, B.S.; Lee, C.H. Metabolomic profiles of Aspergillus oryzae and Bacillus amyloliquefaciens during rice koji fermentation. Molecules 2016, 21, 773. [Google Scholar] [CrossRef]
- Seo, H.S.; Lee, S.M.; Singh, D.; Park, M.K.; Kim, Y.-S.; Shin, H.W.; Cho, S.A.; Lee, C.H. Evaluating the headspace volatolome, primary metabolites, and aroma characteristics of koji fermented with Bacillus amyloliquefaciens and Aspergillus oryzae. J. Microbiol. Biotechnol. 2018, 28, 1260–1269. [Google Scholar] [CrossRef]
- Boumba, V.A.; Ziavrou, K.S.; Vougiouklakis, T. Biochemical pathways generating post-mortem volatile compounds co-detected during forensic ethanol analyses. Forensic Sci. Int. 2008, 174, 133–151. [Google Scholar] [CrossRef]
- Rodrigues, F.; Ludovico, P.; Leão, C. Sugar metabolism in yeasts: An overview of aerobic and anaerobic glucose catabolism. In Biodiversity and Ecophysiology of Yeasts; Péter, G., Rosa, C., Eds.; Springer: Berlin, Germany, 2006; pp. 101–121. [Google Scholar]
- Vivijs, B.; Moons, P.; Geeraerd, A.H.; Aertsen, A.; Michiels, C.W. 2,3-Butanediol fermentation promotes growth of Serratia Plymuthica at low pH but not survival of extreme acid challenge. Int. J. Food Microbiol. 2014, 175, 36–44. [Google Scholar] [CrossRef]
- Pogačić, T.; Maillard, M.-B.; Leclerc, A.; Hervé, C.; Chuat, V.; Valence, F.; Thierry, A. Lactobacillus and Leuconostoc volatilomes in cheese conditions. Appl. Microbiol. Biotechnol. 2016, 100, 2335–2346. [Google Scholar] [CrossRef]
- Gonda, I.; Bar, E.B.; Portnoy, V.; Lev, S.; Burger, J.; Schaffer, A.A.; Tadmor, Y.; Gepstein, S.; Giovannoni, J.J.; Lewinsohn, N.K.E. Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L. Fruit. J. Exp. Bot. 2010, 61, 1111–1123. [Google Scholar] [CrossRef]
- Landaud, S.; Helinck, S.; Bonnarme, P. Formation of volatile sulfur compounds and metabolism of methionine and other sulfur compounds in fermented food. Appl. Microbiol. Biotechnol. 2008, 77, 1191–1205. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-M.; Shin, K.-J.; Lee, S.-J. Exploring nuruk aroma; identification of volatile compounds in commercial fermentation starters. Food Sci. Biotechnol. 2016, 25, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Masuo, S.; Osada, L.; Zhou, S.; Fujita, T.; Takaya, N. Aspergillus oryzae pathways that convert phenylalanine into the flavor volatile 2-phenylethanol. Fungal Genet. Biol. 2015, 77, 22–30. [Google Scholar] [CrossRef]
- Hirata, H.; Ohnishi, T.; Watanabe, N. Biosynthesis of floral scent 2-phenylethanol in rose flowers. Biosci. Biotechnol. Biochem. 2016, 80, 1865–1873. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Zhang, H.; Zhang, T.; Tamogami, S.; Chen, J.Y. Analysis of flavor volatiles of glutinous rice during cooking by combined gas chromatography–mass spectrometry with modified headspace solid-phase microextraction Method. J. Food Composit. Anal. 2009, 22, 347–353. [Google Scholar] [CrossRef]
- Šućur, S.; ČADEŽ, N.; KOŠMERL, T. Volatile phenols in wine: Control measures of Brettanomyces/Dekkera Yeasts. Acta Agric. Slov. 2016, 107, 453–472. [Google Scholar] [CrossRef]
- Kang, J.-E.; Kim, C.-W.; Yeo, S.-H.; Jeong, S.-T.; Kim, Y.-S.; Choi, H.-S. Effect of heat-treated Nuruk on the quality characteristics of aged Yakju. Food Sci. Biotechnol. 2018, 27, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.-Q.; Juwono, N.K.P.; Leong, S.S.J.; Chang, M.W. Production of fatty acid-derived valuable chemicals in synthetic microbes. Front. Bioeng. Biotechnol. 2014, 2, 78. [Google Scholar] [CrossRef] [PubMed]
- Romero-Guido, C.; Belo, I.; Ta, T.M.N.; Cao-Hoang, L.; Alchihab, M.; Gomes, N.; Thonart, P.; Teixeira, J.A.; Destain, J.; Waché, Y. Biochemistry of lactone formation in yeast and fungi and its utilisation for the production of flavour and fragrance compounds. Appl. Microbiol. Biotechnol. 2011, 89, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Mo, X.; Fan, W.; Xu, Y. Changes in volatile compounds of Chinese rice wine wheat Qu during fermentation and storage. J. Inst. Brewing 2009, 115, 300–307. [Google Scholar] [CrossRef]
- Lyu, J.; Nam, P.-W.; Lee, S.-J.; Lee, K.-G. Volatile compounds isolated from rice beers brewed with three medicinal plants. J. Inst. Brewing 2013, 119, 271–279. [Google Scholar] [CrossRef]
- Bacvonkralj, M.; Jug, T.; Komel, E.; Fajt, N.; JARNI, K.; Zivkovic, J.; Mujic, I.; Trutic, N. Effects of ripening degree and sample preparation on peach aroma profile characterization by headspace solid-phase microextraction. Turk. J. Agric. For. 2014, 38, 676–687. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Choi, S.J.; Kwak, J.; Kim, K.; Seo, M.; Moon, T.W.; Lee, Y.-W. Aspergillus oeyzae strains isolated from traditional Korean nuruk: Fermentation properties and influence on rice wine quality. Food Sci. Biotechnol. 2013, 22, 425–432. [Google Scholar] [CrossRef]
- Kalaivani, M.; Sabitha, R.; Kalaiselvan, V.; Rajasekaran, A. Health benefits and clinical impact of major nutrient, red yeast rice: A review. Food Bioproc. Tech. 2009, 3, 333–339. [Google Scholar] [CrossRef]
- Bhadana, B.; Chauhan, M. Bioethanol production using Saccharomyces cerevisiae with different perspectives: Substrates, growth variables, inhibitor reduction and immobilization. Ferment. Technol. 2016, 5, 3–6. [Google Scholar] [CrossRef]
- Steele, J.; Broadbent, J.; Kok, J. Perspectives on the contribution of lactic acid bacteria to cheese flavor development. Curr. Opin. Biotechnol. 2013, 24, 135–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pires, E.J.; Teixeira, J.A.; Brányik, T.; Vicente, A.A. Yeast: The soul of beer’s aroma—A review of flavour-active esters and higher alcohols produced by the brewing yeast. Appl. Microbiol. Biotechnol. 2014, 98, 1937–1949. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |
Figure 1.
Partial least squares-discriminant analysis (PLS-DA) score plot for volatile compounds profiles in fermented rice inoculated by different microbes. (A) Score plot, (B) loading plot. AOR = Aspergillus oryzae, ROR = Rhyzopus oryzae, SCR = Sacharomyces cerevisiae, SFR = Saccharomycopsis fibuligera, LFR = Lactobacillus fermentum, LPR = Lactobacillus plantarum, RICE = controls (non-fermented rice).
Figure 1.
Partial least squares-discriminant analysis (PLS-DA) score plot for volatile compounds profiles in fermented rice inoculated by different microbes. (A) Score plot, (B) loading plot. AOR = Aspergillus oryzae, ROR = Rhyzopus oryzae, SCR = Sacharomyces cerevisiae, SFR = Saccharomycopsis fibuligera, LFR = Lactobacillus fermentum, LPR = Lactobacillus plantarum, RICE = controls (non-fermented rice).

Figure 2.
Metabolic pathway-based volatile compound expressions of fermented rice. The fold change of each volatile compound was calculated by comparing to control (RICE): ND; not detected, NC; not changed. Each square represents the fold change (log2 level) against the control based on the color gradient as shown in the legend (blue; positive, red; negative, and gray; not detected). The formation of volatile compounds which were not detected in the control but were newly produced during fermentation were marked separately (purple; newly detected, white; not detected). Sample abbreviations are shown below; AOR (fermented by Aspergillus oryzae), ROR (Rhyzopus oryzae), SCR (Sacharomyces cerevisiae), SFR (Saccharomycopsis fibuligera), LFR (Lactobacillus fermentum), LPR (Lactobacillus plantarum), and RICE (non-fermented rice).
Figure 2.
Metabolic pathway-based volatile compound expressions of fermented rice. The fold change of each volatile compound was calculated by comparing to control (RICE): ND; not detected, NC; not changed. Each square represents the fold change (log2 level) against the control based on the color gradient as shown in the legend (blue; positive, red; negative, and gray; not detected). The formation of volatile compounds which were not detected in the control but were newly produced during fermentation were marked separately (purple; newly detected, white; not detected). Sample abbreviations are shown below; AOR (fermented by Aspergillus oryzae), ROR (Rhyzopus oryzae), SCR (Sacharomyces cerevisiae), SFR (Saccharomycopsis fibuligera), LFR (Lactobacillus fermentum), LPR (Lactobacillus plantarum), and RICE (non-fermented rice).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Discriminant volatile compounds of different samples.
| No. 1 | Volatile Compound | Sample 2 |
|---|---|---|
| v6 | 2-Methylbutanal | AOR |
| v7 | 3-Methylbutanal | |
| v9 | Ethyl propanoate | |
| v16 | Ethyl 2-methylbutanoate | |
| v46 | 2-Methylpropanoic acid | |
| v48 | 2-Phenylacetaldehyde | |
| v21 | 3-Methylbutyl acetate | ROR |
| v51 | 5-Ethyloxolan-2-one | |
| v36 | 3-Ethoxypropan-1-ol | |
| v55 | Ethyl tetradecanoate | |
| v49 | 1-Phenylethanone | |
| v54 | 2-Phenylethanol | |
| v12 | 3-Methylpentan-2-one | RICE/SCR/SPR/LFR/LPR |
| v18 | Hexanal | |
| v23 | 5-Methylhexan-2-one | |
| v24 | Butan-1-ol | |
| v27 | 3-Hydroxy-3-methylbutan-2-one | |
| v30 | 3-Hydroxybutan-2-one | |
| v41 | Furan-2-carbaldehyde | |
| v45 | Butane-2,3-diol | |
| v53 | Hexanoic acid |
1 Numbered as in the order of retention indices (RI). 2 All abbreviations are shown below; AOR (fermented by Aspergillus oryzae), ROR (Rhyzopus oryzae), SCR (Sacharomyces cerevisiae), SFR (Saccharomycopsis fibuligera), LFR (Lactobacillus fermentum), LPR (Lactobacillus plantarum), and RICE (non-fermented rice).
Table 2.
Volatile compounds identified in fermented rice inoculated by different microbes.
| No. 1 | Volatile Compound | RI 2 | Relative Peak Area (Mean ± SD) 3 | ID 4 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| RICE 5 | AOR | ROR | SCR | SFR | LFR | LPR | ||||
| Carbohydrate metabolism | ||||||||||
| Ethanol fermentation | ||||||||||
| v1 | Acetaldehyde | <800 | ND 6a 7 | 0.723 ± 0.009c | 0.300 ± 0.006b | NDa | NDa | NDa | NDa | B |
| v4 | Ethyl acetate | 885 | 0.013 ± 0.002a | 0.438 ± 0.014f | 0.337 ± 0.007e | 0.092 ± 0.006d | 0.025 ± 0.003ab | 0.052 ± 0.005c | 0.031 ± 0.002b | A |
| v8 | Ethanol | 931 | 0.080 ± 0.007a | 19.811 ± 0.558b | 31.903 ± 0.914c | 33.447 ± 1.427d | 0.466 ± 0.021a | 0.700 ± 0.021a | 0.167 ± 0.003a | A |
| v40 | Acetic acid | 1454 | NDa | 0.058 ± 0.002c | 0.152 ± 0.005e | 0.038 ± 0.002b | 0.096 ± 0.006d | NDa | 0.485 ± 0.014f | A |
| Butanediol fermentation | ||||||||||
| v11 | Butane-2,3-dione | 971 | 0.187 ± 0.010b | NDa | NDa | 0.487 ± 0.017c | 1.259 ± 0.055e | 0.719 ± 0.022d | 0.165 ± 0.003b | A |
| v30 | 3-Hydroxybutan-2-one | 1279 | 0.037 ± 0.002a | 0.069 ± 0.002ab | 0.270 ± 0.002b | 0.107 ± 0.004ab | 0.265 ± 0.012b | 12.172 ± 0.310d | 0.781 ± 0.024c | A |
| v45 | Butane-2,3-diol | 1542 | NDa | NDa | NDa | NDa | 1.657 ± 0.073c | 0.163 ± 0.008b | NDa | A |
| v47 | Butane-1,3-diol | 1579 | NDa | 0.257 ± 0.005c | NDa | NDa | 0.195 ± 0.010b | 0.507 ± 0.016d | NDa | A |
| Amino acid metabolism | ||||||||||
| Valine degradation | ||||||||||
| v6 | 2-Methylbutanal | 909 | NDa | 1.485 ± 0.040c | 0.014 ± 0.003a | 0.070 ± 0.006b | 0.011 ± 0.000a | NDa | NDa | A |
| v16 | Ethyl 2-methylbutanoate | 1050 | NDa | 0.110 ± 0.002b | NDa | NDa | NDa | NDa | NDa | A |
| Leucine degradation | ||||||||||
| v7 | 3-Methylbutanal | 913 | 0.011 ± 0.002a | 3.824 ± 0.106d | 0.108 ± 0.001b | 0.237 ± 0.008c | 0.125 ± 0.007b | NDa | NDa | A |
| v21 | 3-Methylbutyl acetate | 1119 | NDa | NDa | 0.042 ± 0.002b | NDa | NDa | NDa | NDa | A |
| v25 | 3-Methylbutan-1-ol | 1211 | NDa | 14.935 ± 0.420c | 39.594 ± 1.129d | 1.298 ± 0.055b | 1.03 ± 0.045b | 0.036 ± 0.005a | 0.080 ± 0.001a | A |
| v50 | 3-Methylbutanoic acid | 1683 | NDa | 1.314 ± 0.035d | NDa | NDa | 0.324 ± 0.015c | 0.191 ± 0.008b | NDa | A |
| Isoleucine degradation | ||||||||||
| v10 | Ethyl 2-methylpropanoate | 959 | NDa | 0.462 ± 0.011c | 0.204 ± 0.004b | NDa | NDa | NDa | NDa | A |
| v19 | 2-Methylpropan-1-ol | 1100 | NDa | 2.439 ± 0.067d | 2.359 ± 0.059c | 0.216 ± 0.009b | 0.032 ± 0.004a | NDa | 0.026 ± 0.002a | A |
| v46 | 2-Methylpropanoic acid | 1575 | NDa | 0.368 ± 0.008b | NDa | NDa | NDa | NDa | NDa | A |
| Benzenes | ||||||||||
| v13 | Toluene | 1033 | 0.102 ± 0.019c | NDa | 0.146 ± 0.005d | 0.082 ± 0.003b | 0.260 ± 0.012e | 0.082 ± 0.006b | 0.110 ± 0.001c | A |
| v20 | Ethylbenzene | 1115 | 0.027 ± 0.002bc | NDa | 0.064 ± 0.007d | NDa | 0.071 ± 0.005d | 0.024 ± 0.004b | 0.032 ± 0.002c | A |
| v22 | 1,4-Xylene | 1130 | NDa | NDa | 0.046 ± 0.002b | NDa | 0.053 ± 0.005c | NDa | NDa | A |
| v28 | Styrene | 1250 | 0.074 ± 0.004a | 0.068 ± 0.002a | 0.190 ± 0.004f | 0.145 ± 0.006e | 0.136 ± 0.007d | 0.088 ± 0.006b | 0.106 ± 0.001c | A |
| v44 | Benzaldehyde | 1517 | 0.178 ± 0.010e | 0.083 ± 0.001c | 0.048 ± 0.008b | NDa | 0.107 ± 0.006d | NDa | 0.110 ± 0.001d | A |
| v48 | 2-Phenylacetaldehyde | 1637 | NDa | 0.222 ± 0.004b | NDa | NDa | NDa | NDa | NDa | A |
| v49 | 1-Phenylethanone | 1645 | NDa | NDa | 0.039 ± 0.008b | NDa | NDa | NDa | NDa | A |
| v52 | Ethyl 2-phenylacetate | 1785 | NDa | 0.117 ± 0.002c | 0.035 ± 0.002b | NDa | NDa | NDa | NDa | A |
| v54 | 2-Phenylethanol | >1900 | NDa | 0.020 ± 0.002b | 0.455 ± 0.004e | 0.131 ± 0.005d | 0.038 ± 0.004c | NDa | NDa | B |
| Sulfur-containing volatile compounds | ||||||||||
| v17 | (methyldisulfanyl)Methane | 1065 | NDa | 0.114 ± 0.002d | NDa | NDa | 0.024 ± 0.004b | 0.023 ± 0.004b | 0.030 ± 0.002c | A |
| Phenol | ||||||||||
| v56 | 4-Ethenyl-2-methoxyphenol | >1900 | NDa | 0.021 ± 0.002b | 0.020 ± 0.003b | NDa | 0.048 ± 0.005c | 0.069 ± 0.005d | NDa | B |
| Fatty acid metabolism | ||||||||||
| Fatty Ketones | ||||||||||
| v2 | Propan-2-one | 812 | 0.227 ± 0.014b | 1.040 ± 0.010f | 0.103 ± 0.001a | 0.270 ± 0.009c | 0.337 ± 0.015e | 0.316 ± 0.012d | 0.218 ± 0.005b | A |
| v5 | Butan-2-one | 901 | 0.044 ± 0.002b | NDa | 0.372 ± 0.008f | 0.109 ± 0.006e | 0.106 ± 0.006e | 0.085 ± 0.006d | 0.068 ± 0.001c | A |
| v12 | 3-Methylpentan-2-one | 1012 | NDa | NDa | NDa | NDa | 0.175 ± 0.009b | NDa | NDa | A |
| v23 | 5-Methylhexan-2-one | 1136 | NDa | NDa | NDa | NDa | 0.02 ± 0.003b | NDa | NDa | A |
| v27 | 3-Hydroxy-3-methylbutan-2-one | 1239 | NDa | NDa | NDa | NDa | NDa | 0.078 ± 0.006b | NDa | C |
| v33 | 6-Methylhept-5-en-2-one | 1334 | 0.033 ± 0.002a | 0.094 ± 0.001e | 0.051 ± 0.008bc | 0.045 ± 0.002b | 0.063 ± 0.005d | 0.037 ± 0.005a | 0.055 ± 0.001c | A |
| v34 | 3-Hydroxypentan-2-one | 1337 | NDa | NDa | NDa | NDa | 0.060 ± 0.005b | 0.079 ± 0.006c | NDa | C |
| v37 | Nonan-2-one | 1386 | NDa | NDa | NDa | NDa | 0.043 ± 0.004c | 0.012 ± 0.000b | NDa | A |
| Fatty alcohols | ||||||||||
| v15 | Propan-1-ol | 1041 | NDa | 0.447 ± 0.01c | 0.085 ± 0.007b | NDa | NDa | NDa | NDa | A |
| v24 | Butan-1-ol | 1150 | 0.081 ± 0.007c | 0.081 ± 0.001c | 0.069 ± 0.007b | 0.030 ± 0.002a | 0.159 ± 0.008e | 0.069 ± 0.005b | 0.121 ± 0.001d | A |
| v29 | Pentan-1-ol | 1254 | 0.147 ± 0.009cd | 0.040 ± 0.002a | 0.151 ± 0.005d | 0.118 ± 0.005b | 0.138 ± 0.007c | 0.240 ± 0.010e | 0.246 ± 0.006e | A |
| v32 | 3-Methylbut-2-en-1-ol | 1323 | NDa | NDa | NDa | NDa | 0.030 ± 0.004b | 0.027 ± 0.004b | NDa | C |
| v35 | Hexan-1-ol | 1357 | 0.102 ± 0.011b | 0.321 ± 0.007e | 0.311 ± 0.002e | 0.069 ± 0.003a | 0.218 ± 0.011c | 0.501 ± 0.016f | 0.294 ± 0.007d | A |
| v36 | 3-Ethoxypropan-1-ol | 1377 | NDa | NDa | 0.055 ± 0.008b | NDa | NDa | NDa | NDa | A |
| v39 | Oct-1-en-3-ol | 1454 | 0.029 ± 0.002b | 0.050 ± 0.002d | 0.035 ± 0.002c | 0.040 ± 0.002c | 0.058 ± 0.005e | 0.030 ± 0.004b | NDa | A |
| v42 | 2-Ethylhexan-1-ol | 1493 | NDa | 0.044 ± 0.002cd | 0.056 ± 0.008e | 0.046 ± 0.002d | 0.099 ± 0.006f | 0.031 ± 0.004b | 0.038 ± 0.002bc | A |
| Fatty esters | ||||||||||
| v9 | Ethyl propanoate | 950 | NDa | 0.240 ± 0.005b | NDa | NDa | NDa | NDa | NDa | A |
| v14 | Ethyl butanoate | 1034 | NDa | 0.271 ± 0.005c | 0.251 ± 0.003b | NDa | NDa | NDa | NDa | A |
| v55 | Ethyl tetradecanoate | >1900 | NDa | NDa | 0.051 ± 0.008b | NDa | NDa | NDa | NDa | B |
| v57 | Ethyl hexadecanoate | >1900 | NDa | 0.098 ± 0.001b | 0.321 ± 0.002c | NDa | NDa | NDa | NDa | B |
| Fatty aldehydes | ||||||||||
| v18 | Hexanal | 1076 | 1.305 ± 0.086d | 0.190 ± 0.003b | 0.242 ± 0.003b | 0.105 ± 0.004a | 0.249 ± 0.012b | 0.071 ± 0.005a | 0.201 ± 0.004b | A |
| v31 | Octanal | 1284 | 0.026 ± 0.002c | NDa | NDa | 0.018 ± 0.001b | 0.029 ± 0.004d | NDa | NDa | A |
| v38 | Nonanal | 1390 | 0.265 ± 0.017e | 0.182 ± 0.003c | 0.215 ± 0.003d | 0.192 ± 0.008c | 0.193 ± 0.01c | 0.051 ± 0.005a | 0.080 ± 0.001b | A |
| v43 | Decanal | 1496 | NDa | 0.069 ± 0.002d | 0.040 ± 0.008c | NDa | NDa | 0.028 ± 0.004b | 0.025 ± 0.002b | A |
| Fatty carboxylic acid | ||||||||||
| v53 | Hexanoic acid | 1865 | NDa | NDa | NDa | NDa | NDa | NDa | 0.075 ± 0.001b | A |
| Furans | ||||||||||
| v3 | Oxolane | 855 | 0.137 ± 0.008d | NDa | NDa | 0.189 ± 0.007e | NDa | 0.037 ± 0.005b | 0.068 ± 0.001c | A |
| v26 | 2-Pentylfuran | 1227 | 0.045 ± 0.002c | 0.089 ± 0.001f | 0.071 ± 0.007e | 0.036 ± 0.002ab | 0.062 ± 0.005d | 0.029 ± 0.004a | 0.040 ± 0.002bc | A |
| v41 | Furan-2-carbaldehyde | 1459 | 0.021 ± 0.002b | NDa | NDa | NDa | NDa | NDa | NDa | A |
| Lactone | ||||||||||
| v51 | 5-Ethyloxolan-2-one | 1694 | NDa | NDa | 0.094 ± 0.007b | NDa | NDa | NDa | NDa | A |
1 Numbered as in the order of retention indices (RI). 2 RI; Retention indices were determined using n-alkanes (C7–C22) as external standards. 3 Mean values of relative peak area to that of internal standard ± standard deviation (SD). 4 Tentative identification was performed as follows: A, mass spectrum (MS) and retention index (RI) were consistent with those of the mass spectrum database and MS agree with the authentic compound (positive identification, PI); B, MS and PI; C, MS and RI. 5 All abbreviations are shown below; AOR (fermented by Aspergillus oryzae), ROR (Rhyzopus oryzae), SCR (Sacharomyces cerevisiae), SFR (Saccharomycopsis fibuligera), LFR (Lactobacillus fermentum), LPR (Lactobacillus plantarum), and RICE (non-fermented rice).6 Not detected. 7 There are significant differences (p < 0.05) among samples fermented by microbes (AOR, ROR, SCR, SFR, LFR, and LPR) and non-fermented rice (RICE) using Duncan’s multiple comparison test between the samples, shown by having different letter in lowercase.
Table 3.
Distinctive volatile compounds of fermented rice samples.
| AOR 3 | ROR | ||||
|---|---|---|---|---|---|
| No.1 | r-Value 2 | Volatile Compound | No. | r-Value | Volatile Compound |
| v16 | 1.00 | Ethyl 2-methylbutanoate | v21 | 1.00 | 3-Methylbutyl acetate |
| v48 | 1.00 | 2-Phenylacetaldehyde | v51 | 1.00 | 5-Ethyloxolan-2-one |
| v9 | 1.00 | Ethyl propanoate | v36 | 0.99 | 3-Ethoxypropan-1-ol |
| v46 | 1.00 | 2-Methylpropanoic acid | v55 | 0.99 | Ethyl tetradecanoate |
| v6 | 1.00 | 2-Methylbutanal | v49 | 0.98 | 1-Phenylethanone |
| v7 | 1.00 | 3-Methylbutanal | v54 | 0.96 | 2-Phenylethanol |
| v15 | 0.98 | Propan-1-ol | v57 | 0.95 | Ethyl hexadecanoate |
| v2 | 0.97 | Propan-2-one | v5 | 0.95 | Butan-2-one |
| v50 | 0.97 | 3-Methylbutanoic acid | v25 | 0.93 | 3-Methylbutan-1-ol |
| v52 | 0.95 | Ethyl 2-phenylacetate | v28 | 0.75 | Styrene |
| v17 | 0.95 | (methyldisulfanyl)Methane | |||
| v1 | 0.91 | Acetaldehyde | |||
| v10 | 0.90 | Ethyl 2-methylpropanoate | |||
| v33 | 0.88 | 6-Methylhept-5-en-2-one | |||
| v43 | 0.78 | Decanal | |||
| v4 | 0.75 | Ethyl acetate | |||
| v29 | −0.72 | Pentan-1-ol | |||
| SPR | SCR | ||||
| v12 | 1.00 | 3-Methylpentan-2-one | v3 | 0.93 | Oxolane |
| v45 | 0.99 | Butane-2,3-diol | |||
| v23 | 0.99 | 5-Methylhexan-2-one | |||
| v37 | 0.96 | Nonan-2-one | |||
| v42 | 0.93 | 2-Ethylhexan-1-ol | |||
| v13 | 0.83 | Toluene | |||
| v31 | 0.82 | Octanal | |||
| v11 | 0.82 | Butane-2,3-dione | |||
| v24 | 0.76 | Butan-1-ol | |||
| v22 | 0.70 | 1,4-Xylene | |||
| LFR | LPR | ||||
| v30 | 1.00 | 3-Hydroxybutan-2-one | v53 | 1.00 | Hexanoic acid |
| v27 | 1.00 | 3-Hydroxy-3-methylbutan-2-one | v40 | 0.96 | Acetic acid |
| v47 | 0.83 | Butane-1,3-diol | |||
| v56 | 0.76 | 4-Ethenyl-2-methoxyphenol | |||
| v34 | 0.75 | 3-Hydroxypentan-2-one | |||
| v35 | 0.74 | Hexan-1-ol | |||
1 Numbered as in the order of retention indices. 2 The correlation coefficient r was selected as a significant difference (r > 0.7 or r < −0.7) between a microbe and the levels of volatile compounds. 3 Sample abbreviations are shown below; AOR (fermented by Aspergillus oryzae), ROR (Rhyzopus oryzae), SCR (Saccharomyces cerevisiae), SFR (Saccharomycopsis fibuligera), LFR (Lactobacillus fermentum), LPR (Lactobacillus plantarum), and RICE (non-fermented rice).
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Park, M.K.; Kim, Y.-S. Distinctive Formation of Volatile Compounds in Fermented Rice Inoculated by Different Molds, Yeasts, and Lactic Acid Bacteria. Molecules 2019, 24, 2123. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24112123
AMA Style
Park MK, Kim Y-S. Distinctive Formation of Volatile Compounds in Fermented Rice Inoculated by Different Molds, Yeasts, and Lactic Acid Bacteria. Molecules. 2019; 24(11):2123. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24112123
Chicago/Turabian StylePark, Min Kyung, and Young-Suk Kim. 2019. "Distinctive Formation of Volatile Compounds in Fermented Rice Inoculated by Different Molds, Yeasts, and Lactic Acid Bacteria" Molecules 24, no. 11: 2123. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24112123