
Polyphenol Effects on Splenic Cytokine Response in Post-Weaning Contactin 1-Overexpressing Transgenic Mice




Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
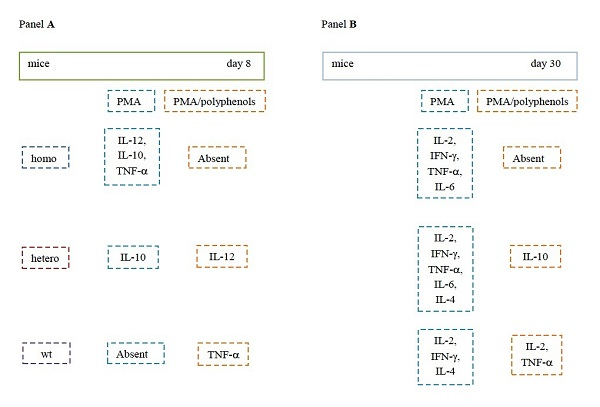
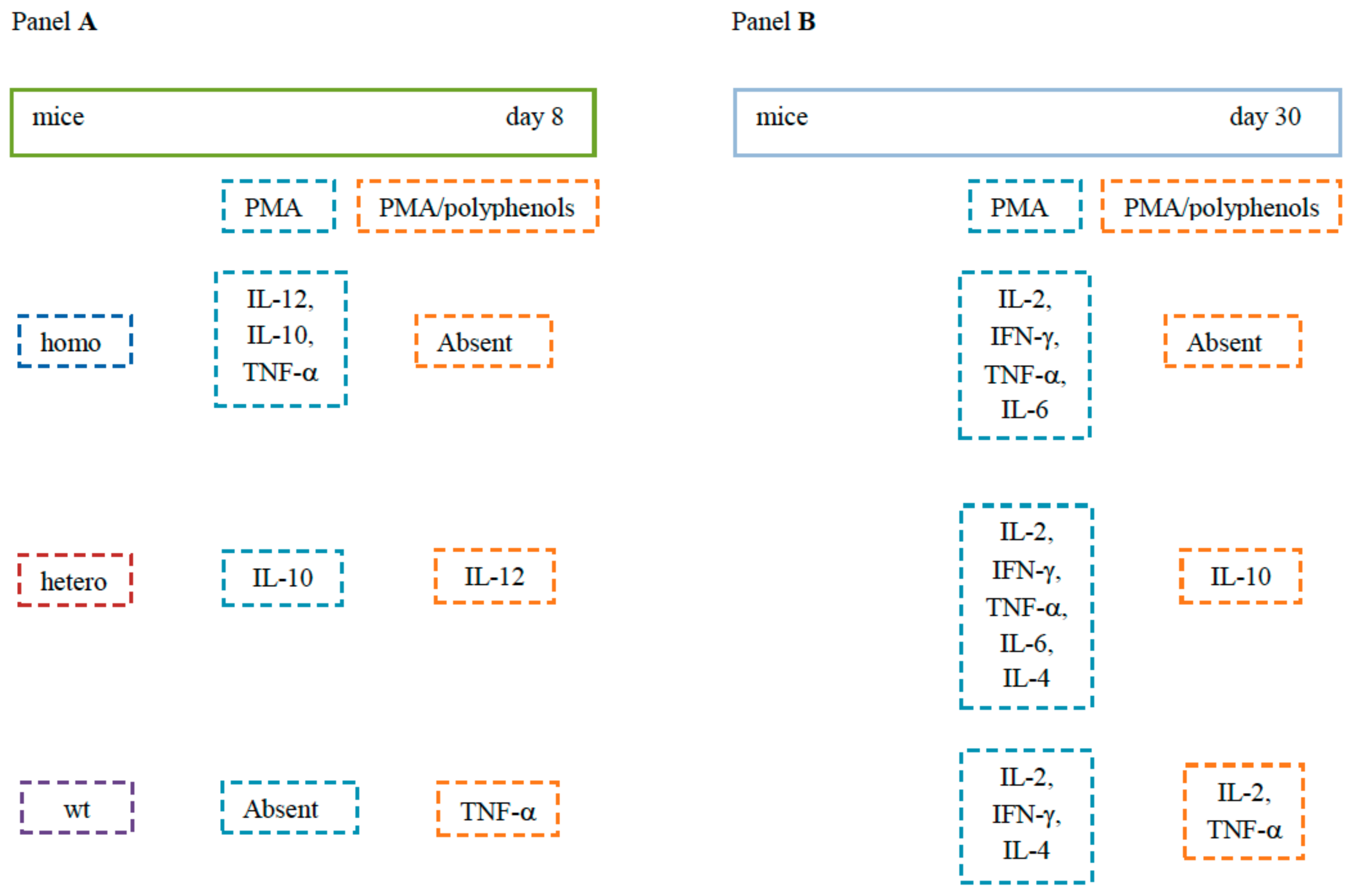
2. Results
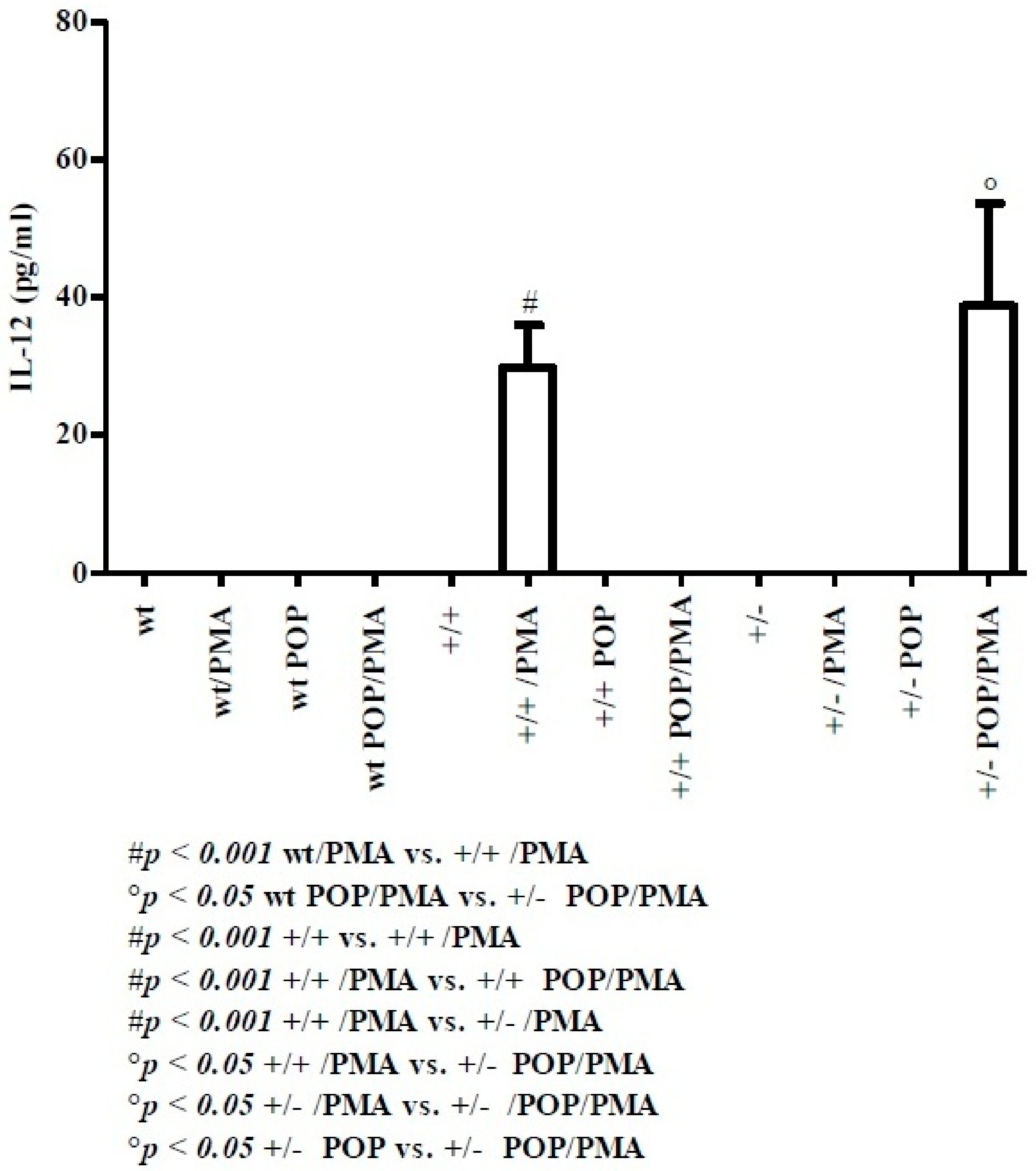
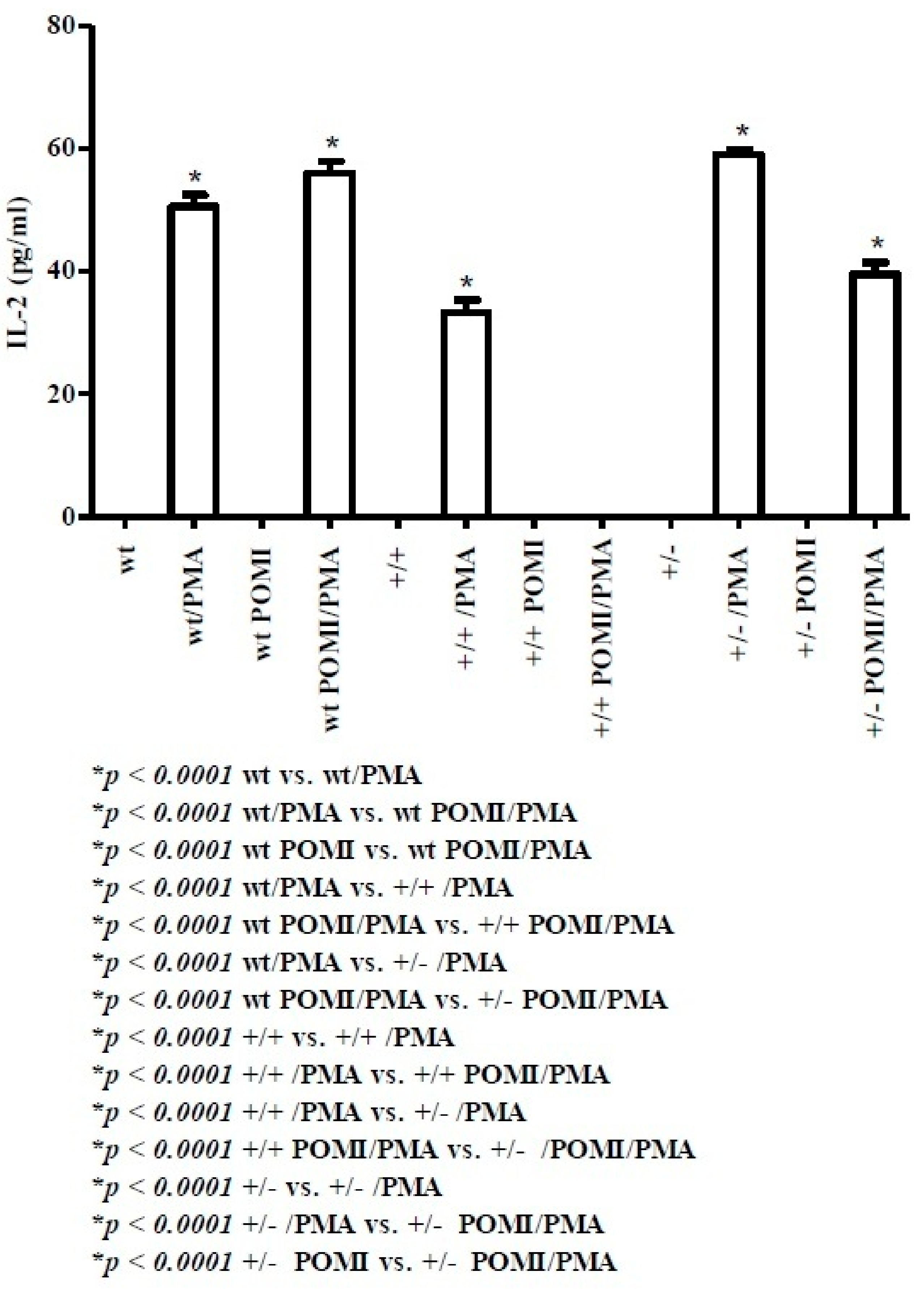
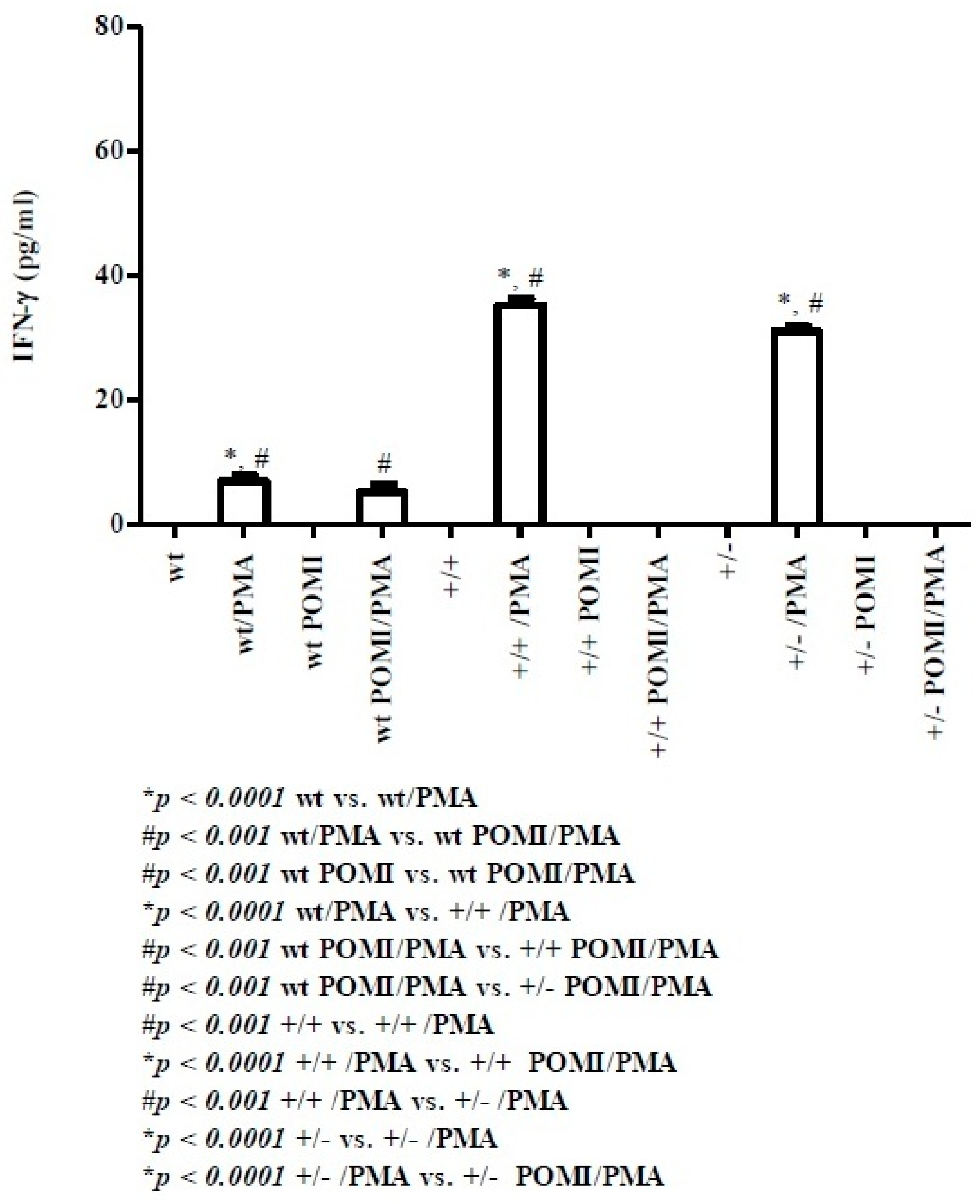
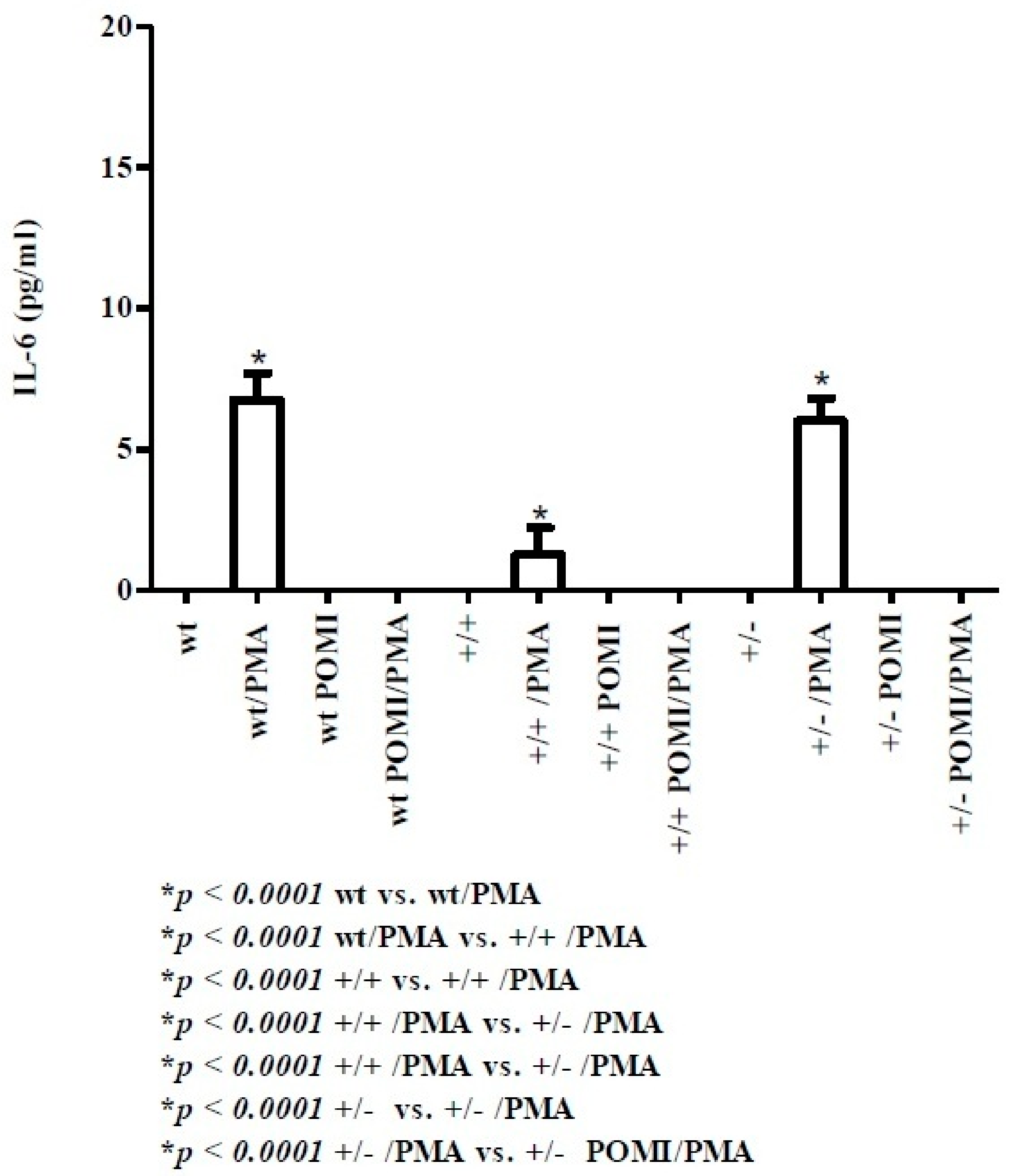
2.1. Th1-Related Cytokines
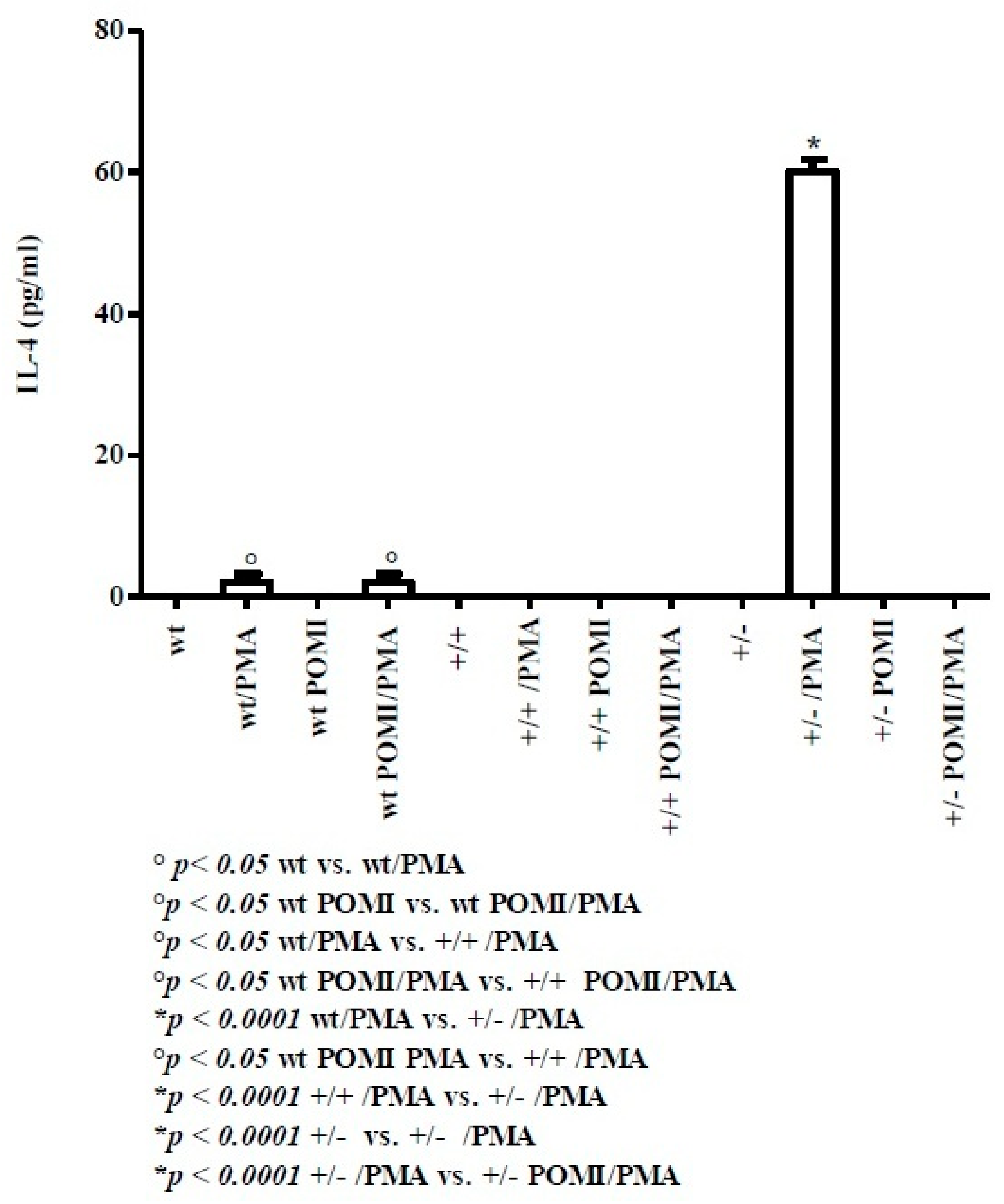
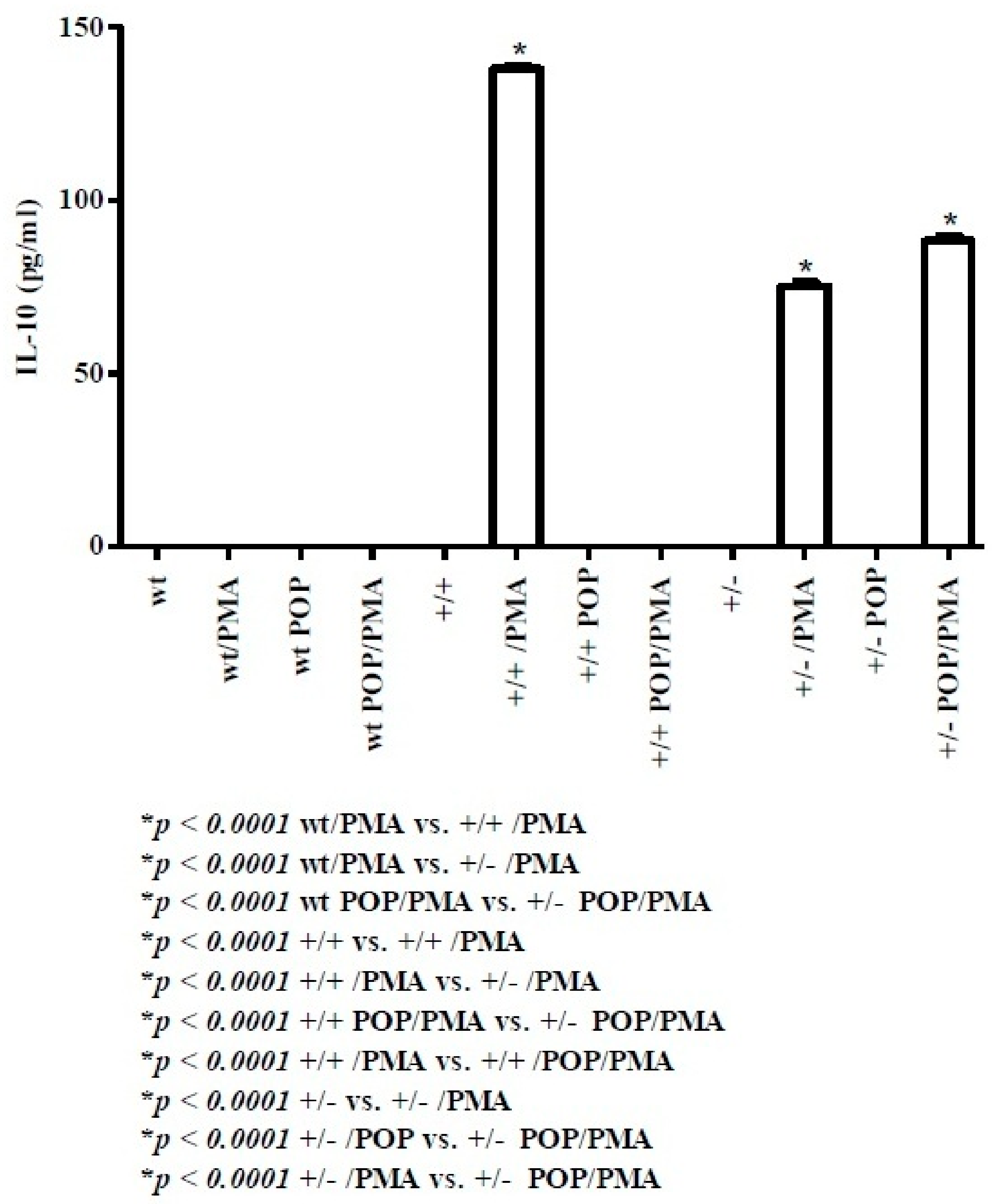
2.2. Th2-Related Cytokines
3. Discussion
4. Materials and Methods
4.1. Mice Strains
4.2. Polyphenol Source and Addition to Feed
4.3. Experimental Design
Dams during Pregnancy and Lactation
4.4. Cytokine Evaluation by Flow Cytometry
4.5. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Cacho, N.T.; Lawrence, R.M. Innate immunity and breast milk. Front. Immunol. 2017, 8, 584. [Google Scholar] [CrossRef] [PubMed]
- Zdrojewicz, Z.; Pachura, E.; Pachura, P. The Thymus: A Forgotten, But Very Important Organ. Adv. Clin. Exp. Med. 2016, 25, 369–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuper, C.F.; van Bilsen, J.; Cnossen, H.; Houben, G.; Garthoff, J.; Wolterbee, A. Development of immune organs and functioning in humans and test animals: Implications for immune intervention studies. In Reproductive Toxicology; Elsevier Science B.V.: Amsterdam, The Netherlands, 2016; Volume 64, pp. 180–190. [Google Scholar]
- Sano, S.; Takahama, Y.; Sugawara, T.; Kosaka, H.; Itami, S.; Yoshikawa, K.; Miyazaki, J.; van Ewijk, W.; Takeda, J. Stat3 in thymic epithelial cells is essential for postnatal maintenance of thymic architecture and thymocyte survival. Immunity 2001, 15, 261–273. [Google Scholar] [CrossRef]
- Golub, R.; Cumano, A. Embryonic hematopoiesis. Blood Cells Mol. Dis. 2013, 51, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.E.; Turley, S.J. Stromal infrastructure of the lymph node and coordination of immunity. Trends Immunol. 2015, 36, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Hoorweg, K.; Cupedo, T. Development of human lymph nodes and Peyer’s patches. Semin. Immunol. 2008, 20, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, H.; Horie, K.; Tanaka, K.; Nagata, H.; Aizawa, S.; Hiramoto, M.; Ryouke, T.; Aijima, H. Patterns of age-dependent changes in the numbers of lymph follicles and germinal centres in somatic and mesenteric lymph nodes in growing C57Bl/6 mice. J. Anat. 2001, 198, 189–205. [Google Scholar] [CrossRef] [PubMed]
- Cecchini, M.G.; Dominguez, M.G.; Mocci, S.; Wetterwald, A.; Felix, R.; Fleisch, H.; Chisholm, O.; Hofstetter, W.; Pollard, J.W.; Stanley, E.R. Role of colony stimulating factor-1 in the establishment and regulation of tissue macrophages during postnatal development of the mouse. Development 1994, 120, 1357–1372. [Google Scholar]
- Ridge, J.P.; Fuchs, E.J.; Matzinger, P. Neonatal tolerance revisited: Turning on newborn T cells with dendritic cells. Science 1996, 271, 1723–1726. [Google Scholar] [CrossRef]
- Garcia, A.M.; Fadel, S.A.; Cao, S.; Sarzotti, M. T cell immunity in neonates. Immunol. Res. 2000, 22, 177–190. [Google Scholar] [CrossRef]
- Zhao, J.; Kim, K.D.; Yang, X.; Auh, S.; Fu, Y.X.; Tang, H. Hyper innate responses in neonates lead to increased morbidity and mortality after infection. Proc. Natl. Acad. Sci. USA 2008, 105, 7528–7533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.D.; Zhao, J.; Auh, S.; Yang, X.; Du, P.; Tang, H.; Fu, Y.X. Adaptive immune cells temper initial innate responses. Nat. Med. 2007, 13, 1248–1252. [Google Scholar] [CrossRef] [PubMed]
- Hendricks, D.W.; Fink, P.J. Recent thymic emigrants are biased against the T-helper type 1 and toward the T-helper type 2 effector lineage. Blood 2011, 117, 1239–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimoto, M.; Yoder, M.C.; Guevara, P.; Adkins, B. The murine Th2 locus undergoes epigenetic modification in the thymus during fetal and postnatal ontogeny. PLoS ONE 2013, 8, e51587. [Google Scholar] [CrossRef] [PubMed]
- Schijf, M.A.; Kruijsen, D.; Bastiaans, J.; Coenjaerts, F.E.; Garssen, J.; van Bleek, G.M.; van’t Land, B. Specific dietary oligosaccharides increase Th1 responses in a mouse respiratory syncytial virus infection model. J. Virol. 2012, 86, 11472–11482. [Google Scholar] [CrossRef] [PubMed]
- Hogenkamp, A.; Knippels, L.M.; Garssen, J.; van Esch, B.C. Supplementation of Mice with Specific Nondigestible Oligosaccharides during Pregnancy or Lactation Leads to Diminished Sensitization and Allergy in the Female Offspring. J. Nutr. 2015, 145, 996–1002. [Google Scholar] [CrossRef] [PubMed]
- Hogenkamp, A.; Thijssen, S.; van Vlies, N.; Garssen, J. Supplementing Pregnant Mice with a Specific Mixture of Nondigestible Oligosaccharides Reduces Symptoms of Allergic Asthma in Male Offspring. J. Nutr. 2015, 45, 996–1002. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, L.; Kjær, T.M.; Porsgaard, T.; Fruekilde, M.B.; Mu, H.; Frøkiær, H. Maternal intake of fish oil but not of linseed oil reduces the antibody response in neonatal mice. Lipids 2011, 46, 171–178. [Google Scholar] [CrossRef]
- Van Vlies, N.; Hogenkamp, A.; Fear, A.L.; van Esch, B.C.; Oosting, A.; van de Heijning, B.; van der Beek, E.; Calder, P.C.; Garssen, J. Perinatal programming of murine immune responses by polyunsaturated fatty acids. J. Dev. Orig. Health Dis. 2011, 2, 112–123. [Google Scholar] [CrossRef]
- Toomer, O.T.; Ferguson, M.; Pereira, M.; Do, A.; Bigley, E.; Gaines, D.; Williams, K. Maternal and postnatal dietary probiotic supplementation enhances splenic regulatory T helper cell population and reduces ovalbumin allergen-induced hypersensitivity responses in mice. Immunobiology 2014, 219, 367–376. [Google Scholar] [CrossRef]
- Nallagatla, H.; Hemalatha, R.; Kondapalli, N.B.; Mohammed, S. Impact of a novel synbiotic supplementation during gestation and lactation on immune responses in the Swiss albino mice offspring. J. Sci. Food Agric. 2017, 97, 2453–2459. [Google Scholar] [CrossRef] [PubMed]
- Magrone, T.; Jirillo, E. Effects of Polyphenols on Inflammatory-Allergic Conditions: Experimental and Clinical Evidences. In Polyphenols: Prevention and Treatment of Human Disease, 2nd ed.; Elsevier: London, UK, 2018; Volume 2, pp. 253–259. [Google Scholar]
- Magrone, T.; Kumazawa, Y.; Jirillo, E. Polyphenol-mediated beneficial effects in healthy status and disease with special references to immune-based mechanisms. In Polyphenols in Human Health and Disease, 1st ed.; Elsevier: London, UK, 2014; Volume 1, pp. 467–479. [Google Scholar]
- Magrone, T.; Panaro, M.A.; Jirillo, E.; Covelli, V. Molecular effects elicited in vitro by red wine on human healthy peripheral blood mononuclear cells: Potential therapeutical application of polyphenols to diet-related chronic diseases. Curr. Pharm. Des. 2008, 14, 2758–2766. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, K.; Matsumoto, T.; Kumazawa, Y. Effects of antioxidant polyphenols on TNF-alpha-related diseases. Curr. Top. Med. Chem. 2011, 11, 1767–1779. [Google Scholar] [CrossRef]
- Tominaga, T.; Kawaguchi, K.; Kanesaka, M.; Kawauchi, H.; Jirillo, E.; Kumazawa, Y. Suppression of type-I allergic responses by oral administration of grape marc fermented with Lactobacillus plantarum. Immunopharmacol. Immunotoxicol. 2010, 32, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Marzulli, G.; Magrone, T.; Vonghia, L.; Kaneko, M.; Takimoto, H.; Kumazawa, Y.; Jirillo, E. Immunomodulating and anti-allergic effects of Negroamaro and Koshu Vitis vinifera fermented grape marc (FGM). Curr. Pharm. Des. 2014, 20, 864–868. [Google Scholar] [CrossRef]
- Bizzoca, A.; Virgintino, D.; Lorusso, L.; Buttiglione, M.; Yoshida, L.; Polizzi, A.; Tattoli, M.; Cagiano, R.; Rossi, F.; Kozlov, S.; et al. Transgenic mice expressing F3/contactin from the TAG-1 promoter exhibit developmentally regulated changes in the differentiation of cerebellar neurons. Development 2003, 130, 29–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.D.; Ang, B.T.; Karsak, M.; Hu, W.P.; Cui, X.Y.; Duka, T.; Takeda, Y.; Chia, W.; Sankar, N.; Ng, Y.K.; et al. F3/contactin acts as a functional ligand for Notch during oligodendrocyte maturation. Cell 2003, 115, 163–175. [Google Scholar] [CrossRef]
- Hu, Q.D.; Ma, Q.H.; Gennarini, G.; Xiao, Z.C. Cross-talk between F3/contactin and Notch at axoglial interface: A role in oligodendrocyte development. Dev. Neurosci. 2006, 28, 25–33. [Google Scholar] [CrossRef]
- Bizzoca, A.; Corsi, P.; Polizzi, A.; Pinto, M.F.; Xenaki, D.; Furley, A.J.; Gennarini, G. F3/Contactin acts as a modulator of neurogenesis during cerebral cortex development. Dev. Biol. 2012, 365, 133–151. [Google Scholar] [CrossRef] [Green Version]
- Coluccia, A.; Tattoli, M.; Bizzoca, A.; Arbia, S.; Lorusso, L.; De Benedictis, L.; Buttiglione, M.; Cuomo, V.; Furley, A.; Gennarini, G.; et al. Transgenic mice expressing F3/contactin from the transient axonal glycoprotein promoter undergo developmentally regulated deficits of the cerebellar function. Neuroscience 2004, 123, 155–166. [Google Scholar] [CrossRef]
- Abdi, K.; Singh, N.J. Making many from few: IL-12p40 as a model for the combinatorial assembly of heterodimeric cytokines. Cytokine 2015, 76, 53–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Justiz Vaillant, A.A.; Qurie, A. Interleukin. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2019. Available online: https://www.ncbi.nlm.nih.gov/books/NBK499840/ (accessed on 6 June 2019).
- Kak, G.; Raza, M.; Tiwari, B.K. Interferon-gamma (IFN-γ): Exploring its implications in infectious diseases. Biomol. Concepts 2018, 9, 64–79. [Google Scholar] [CrossRef] [PubMed]
- May, R.D.; Fung, M. Strategies targeting the IL-4/IL-13 axes in disease. Cytokine 2015, 75, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.M.S.; Gomes, S.T.M.; Ishak, R.; Vallinoto, A.C.R. Regulatory T Cell and Forkhead Box Protein 3 as Modulators of Immune Homeostasis. Front. Immunol. 2017, 8, 605. [Google Scholar] [CrossRef] [Green Version]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef] [PubMed]
- Hunter, C.A.; Jones, S.A. Corrigendum: IL-6 as a keystone cytokine in health and disease. Nat. Immunol. 2017, 18, 1271. [Google Scholar] [CrossRef] [PubMed]
- Watts, T.H. TNF/TNFR family members in costimulation of T cell responses. Annu. Rev. Immunol. 2005, 23, 23–68. [Google Scholar] [CrossRef] [PubMed]
- Magrone, T.; Jirillo, E. The interplay between the gut immune system and microbiota in health and disease: Nutraceutical intervention for restoring intestinal homeostasis. Curr. Pharm. Des. 2013, 19, 1329–1342. [Google Scholar] [CrossRef] [PubMed]
- Puzzo, D.; Bizzoca, A.; Privitera, L.; Furnari, D.; Giunta, S.; Girolamo, F.; Pinto, M.; Gennarini, G.; Palmeri, A. F3/Contactin promotes hippocampal neurogenesis, synaptic plasticity, and memory in adult mice. Hippocampus 2013, 23, 1367–1382. [Google Scholar] [CrossRef] [PubMed]
- Massaro, A.; Bizzoca, A.; Corsi, P.; Pinto, M.F.; Carratù, M.R.; Gennarini, G. Significance of F3/Contactin gene expression in cerebral cortex and nigrostriatal development. Mol. Cell Neurosci. 2012, 50, 221–237. [Google Scholar] [CrossRef] [PubMed]
- Edelman, G.M. Mediation and inhibition of cell adhesion by morphoregulatory molecules. Cold Spring Harb. Symp. Quant. Biol. 1992, 57, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Roselli, M.; Lovece, A.; Bruno, C.; Cavalluzzi, M.M.; Laghezza, A.; Mercurio, A.; Lentini, G.; Corbo, F.; La Forgia, F.; Fontana, S.; et al. Antioxidant activity of Uva di Troia Canosina: Comparison of two extraction methods. Clin. Immunol. Endocr. Metab. Drugs 2015, 2, 1–5. [Google Scholar] [CrossRef]
Sample Availability: Samples of polyphenols are available from the authors. |








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magrone, T.; Spagnoletta, A.; Bizzoca, A.; Russo, M.A.; Jirillo, E.; Gennarini, G. Polyphenol Effects on Splenic Cytokine Response in Post-Weaning Contactin 1-Overexpressing Transgenic Mice. Molecules 2019, 24, 2205. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24122205
Magrone T, Spagnoletta A, Bizzoca A, Russo MA, Jirillo E, Gennarini G. Polyphenol Effects on Splenic Cytokine Response in Post-Weaning Contactin 1-Overexpressing Transgenic Mice. Molecules. 2019; 24(12):2205. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24122205
Chicago/Turabian StyleMagrone, Thea, Anna Spagnoletta, Antonella Bizzoca, Matteo Antonio Russo, Emilio Jirillo, and Gianfranco Gennarini. 2019. "Polyphenol Effects on Splenic Cytokine Response in Post-Weaning Contactin 1-Overexpressing Transgenic Mice" Molecules 24, no. 12: 2205. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24122205