
Effects of Defoliation on Phenolic Concentrations, Antioxidant and Antibacterial Activity of Grape Skin Extracts of the Varieties Blaufränkisch and Merlot (Vitis vinifera L.)

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Effects of Defoliation Treatments on the Concentration of Phenolic Compounds
2.2. Effects of Defoliation Treatments on the Concentration of Total Anthocyanins
2.3. Effects of Defoliation Treatments on the Antioxidant Activity
2.4. Effects Of Defoliation Treatments on Individual Anthocyanins
2.5. Effects of Defoliation Treatments on Antibacterial Activity
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Grape Skin Extraction
4.3. Determination of Total Phenolic Concentration (TPC)
4.4. Determination of Total Anthocyanins (TAC)
4.5. 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Radical Scavenging Activity
4.6. High-Performance Liquid Chromatography (HPLC): Separation and Detection of Anthocyanins
4.7. Antibacterial Susceptibility Testing
4.7.1. Microorganisms and Growth Conditions
4.7.2. Minimum Inhibitory Concentrations
4.8. Statistical Data Processing
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Beckman, C.H. Phenolic-storing cells: Keys to programmed cell death and periderm formation in wilt disease resistance and in general defence responses in plants? Physiol. Mol. Plant Pathol. 2000, 57, 101–110. [Google Scholar] [CrossRef]
- Graf, B.A.; Milbury, P.E.; Blumberg, J.B. Flavonols, flavones, flavanones, and human health: Epidemiological evidence. J. Med. Food 2005, 8, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Arts, I.C.W.; Hollman, P.C.H. Polyphenols and disease risk in epidemiologic studies. Am. J. Clin. Nutr. 2005, 81, 317S–325S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, E.-Q.; Deng, G.-F.; Guo, Y.-J.; Li, H.-B. Biological activities of polyphenols from grapes. Int. J. Mol. Sci. 2010, 11, 622–646. [Google Scholar] [CrossRef] [PubMed]
- Raj, N.K.; Sripal, R.M.; Chaluvadi, M.R.; Krishna, D.R. Bioflavonoids classification, pharmacological, biochemical effects and therapeutic potential. Indian J. Pharmacol. 2001, 33, 2. [Google Scholar]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Han, X.; Shen, T.; Lou, H. Dietary Polyphenols and Their Biological Significance. Int. J. Mol. Sci. 2007, 8, 950–988. [Google Scholar] [CrossRef] [Green Version]
- Seeram, N.P.; Cichewicz, R.H.; Chandra, A.; Nair, M.G. Cyclooxygenase Inhibitory and Antioxidant Compounds from Crabapple Fruits. J. Agric. Food Chem. 2003, 51, 1948–1951. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.J.; Howard, L.R.; Prior, R.L.; Clark, J.R. Flavonoid glycosides and antioxidant capacity of various blackberry, blueberry and red grape genotypes determined by high-performance liquid chromatography/mass spectrometry. J. Sci. Food Agric. 2004, 84, 1771–1782. [Google Scholar] [CrossRef]
- Letenneur, L.; Proust-Lima, C.; Le Gouge, A.; Dartigues, J.F.; Barberger-Gateau, P. Flavonoid intake and cognitive decline over a 10-year period. Am. J. Epidemiol. 2007, 165, 1364–1371. [Google Scholar] [CrossRef]
- Scarmeas, N.; Luchsinger, J.A.; Mayeux, R.; Stern, Y. Mediterranean diet and Alzheimer disease mortality. Neurology 2007, 69, 1084–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, K.; Maltese, F.; Choi, Y.H.; Verpoorte, R. Metabolic constituents of grapevine and grape-derived products. Phytochem. Rev. Proc. Phytochem. Soc. Eur. 2010, 9, 357–378. [Google Scholar] [CrossRef] [PubMed]
- Perumalla, A.V.S.; Hettiarachchy, N.S. Green tea and grape seed extracts — Potential applications in food safety and quality. Food Res. Int. 2011, 44, 827–839. [Google Scholar] [CrossRef]
- Jayaprakasha, G.K.; Selvi, T.; Sakariah, K.K. Antibacterial and antioxidant activities of grape (Vitis vinifera) seed extracts. Food Res. Int. 2002, 36, 117–122. [Google Scholar] [CrossRef]
- Rhodes, P.L.; Mitchell, J.W.; Wilson, M.W.; Melton, L.D. Antilisterial activity of grape juice and grape extracts derived from Vitis vinifera variety Ribier. Int. J. Food Microbiol. 2006, 107, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.C.; Huang, G.; Haley-Zitlin, V.; Jiang, X. Antibacterial Effects of Grape Extracts on Helicobacter pylori. Appl. Environ. Microbiol. 2009, 75, 848–852. [Google Scholar] [CrossRef] [Green Version]
- Sternad Lemut, M.; Trost, K.; Sivilotti, P.; Vrhovsek, U. Pinot Noir grape colour related phenolics as affected by leaf removal treatments in the Vipava Valley. J. Food Compos. Anal. 2011, 24, 777–784. [Google Scholar] [CrossRef]
- Reynolds, A.G.; Wardle, D.A.; Hall, J.W.; Dever, M. Fruit Maturation of Four Vitis vinifera Cultivars in Response to Vineyard Location and Basal Leaf Removal. Am. J. Enol. Vitic. 1995, 46, 542–558. [Google Scholar]
- Matus, J.T.; Poupin, M.J.; Cañón, P.; Bordeu, E.; Alcalde, J.A.; Arce-Johnson, P. Isolation of WDR and bHLH genes related to flavonoid synthesis in grapevine (Vitis vinifera L.). Plant Mol. Biol. 2010, 72, 607–620. [Google Scholar] [CrossRef]
- Pereira, G.E.; Gaudillere, J.-P.; Pieri, P.; Hilbert, G.; Maucourt, M.; Deborde, C.; Moing, A.; Rolin, D. Microclimate influence on mineral and metabolic profiles of grape berries. J. Agric. Food Chem. 2006, 54, 6765–6775. [Google Scholar] [CrossRef]
- Guidoni, S.; Ferrandino, A.; Novello, V. Effects of Seasonal and Agronomical Practices on Skin Anthocyanin Profile of Nebbiolo Grapes. Am. J. Enol. Vitic. 2008, 59, 22–29. [Google Scholar]
- Teixeira, A.; Eiras-Dias, J.; Castellarin, S.D.; Gerós, H. Berry phenolics of grapevine under challenging environments. Int. J. Mol. Sci. 2013, 14, 18711–18739. [Google Scholar] [CrossRef] [PubMed]
- Verzera, A.; Tripodi, G.; Dima, G.; Condurso, C.; Scacco, A.; Cincotta, F.; Giglio, D.M.L.; Santangelo, T.; Sparacio, A. Leaf removal and wine composition of Vitis vinifera L. cv. Nero d’Avola: The volatile aroma constituents. J. Sci. Food Agric. 2016, 96, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Buesa, I.; Caccavello, G.; Basile, B.; Merli, M.C.; Poni, S.; Chirivella, C.; Intrigliolo, D.S. Delaying berry ripening of Bobal and Tempranillo grapevines by late leaf removal in a semi-arid and temperate-warm climate under different water regimes. Aust. J. Grape Wine Res. 2019, 25, 70–82. [Google Scholar] [CrossRef]
- Lanari, V.; Lattanzi, T.; Borghesi, L.; Silvestroni, O.; Palliotti, A. Post-Veraison Mechanical Leaf Removal Delays Berry Ripening on ‘Sangiovese’ and ‘Montepulciano’ Grapevines. In Proceedings of the Proceedings of the 1st International Workshop on Vineyard Mechanization and Grape and Wine Quality, Piacenza, Italy, 27–29 June 2012; pp. 327–333. [Google Scholar]
- Zhang, Z.-Z.; Che, X.-N.; Pan, Q.-H.; Li, X.-X.; Duan, C.-Q. Transcriptional activation of flavan-3-ols biosynthesis in grape berries by UV irradiation depending on developmental stage. Plant Sci. Int. J. Exp. Plant Biol. 2013, 208, 64–74. [Google Scholar] [CrossRef]
- Kennedy, J.A.; Matthews, M.A.; Waterhouse, A.L. Effect of Maturity and Vine Water Status on Grape Skin and Wine Flavonoids. Am. J. Enol. Vitic. 2002, 53, 268–274. [Google Scholar]
- Zhu, L.; Zhang, Y.; Lu, J. Phenolic contents and compositions in skins of red wine grape cultivars among various genetic backgrounds and originations. Int. J. Mol. Sci. 2012, 13, 3492–3510. [Google Scholar] [CrossRef]
- Yin, H.-T.; Tian, Q.-Z.; Guan, L.; Zhou, Y.; Huang, X.-E.; Zhang, H. In vitro and in vivo evaluation of the antitumor efficiency of resveratrol against lung cancer. Asian Pac. J. Cancer Prev. APJCP 2013, 14, 1703–1706. [Google Scholar] [CrossRef]
- Fan, G.-J.; Liu, X.-D.; Qian, Y.-P.; Shang, Y.-J.; Li, X.-Z.; Dai, F.; Fang, J.-G.; Jin, X.-L.; Zhou, B. 4,4′-Dihydroxy-trans-stilbene, a resveratrol analogue, exhibited enhanced antioxidant activity and cytotoxicity. Bioorg. Med. Chem. 2009, 17, 2360–2365. [Google Scholar] [CrossRef]
- Bonnefont-Rousselot, D. Resveratrol and Cardiovascular Diseases. Nutrients 2016, 8. [Google Scholar] [CrossRef]
- Drenjančević, M.; Jukić, V.; Zmaić, K.; Kujundžić, T.; Rastija, V. Effects of early leaf removal on grape yield, chemical characteristics, and antioxidant activity of grape variety Cabernet Sauvignon and wine from eastern Croatia. Acta Agric. Scand. Sect. B Soil Plant Sci. 2017, 67, 705–711. [Google Scholar]
- Poni, S.; Casalini, L.; Bernizzoni, F.; Civardi, S.; Intrieri, C. Effects of Early Defoliation on Shoot Photosynthesis, Yield Components, and Grape Composition. Am. J. Enol. Vitic. 2006, 57, 397–407. [Google Scholar]
- Risco, D.; Pérez, D.; Yeves, A.; Castel, J.R.; Intrigliolo, D.S. Early defoliation in a temperate warm and semi-arid Tempranillo vineyard: Vine performance and grape composition. Aust. J. Grape Wine Res. 2014, 20, 111–122. [Google Scholar] [CrossRef]
- Mijowska, K.; Ochmian, I.; Oszmiański, J. Impact of Cluster Zone Leaf Removal on Grapes cv. Regent Polyphenol Content by the UPLC-PDA/MS Method. Molecules 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Jerman, T.; Lemut, M.S.; Trôst, K.; Pospišil, M. The impact of early leaf removal on polyphenol/anthocyanin content and in vitro antioxidant potential of “Pinot Noir” grapes from Vipava Valley. Proceedings of 46th Croatian and 6th International Symposium on Agriculture, Opatija, Croatia, 14–18 February 2011; p. 940. [Google Scholar]
- Jones, G.V.; Davis, R.E. Climate Influences on Grapevine Phenology, Grape Composition, and Wine Production and Quality for Bordeaux, France. Am. J. Enol. Vitic. 2000, 51, 249–261. [Google Scholar]
- Ferrer, M.; Echeverría, G.; Carbonneau, A. Effect of berry weight and its components on the contents of sugars and anthocyanins of three varieties of Vitis vinifera L. under different water supply conditions. South Afr. J. Enol. Vitic. 2014, 35, 103–113. [Google Scholar] [CrossRef]
- Barbagallo, M.G.; Guidoni, S.; Hunter, J.J. Berry Size and Qualitative Characteristics of Vitis vinifera L. cv. Syrah. South Afr. J. Enol. Vitic. 2011, 32, 129–136. [Google Scholar] [CrossRef]
- Roby, G.; Harbertson, J.F.; Adams, D.A.; Matthews, M.A. Berry size and vine water deficits as factors in winegrape composition: Anthocyanins and tannins. Aust. J. Grape Wine Res. 2004, 10, 100–107. [Google Scholar] [CrossRef]
- Nirmala, J.G.; Narendhirakannan, R.T. In Vitro Antioxidant And Antimicrobial Activities Of Grapes (Vitis Vinifera. L) Seed And Skin Extracts - Muscat Variety. Int. J. Pharm. Pharm. Sci. 2011, 3, 8. [Google Scholar]
- Intrigliolo, D.S.; Llacer, E.; Revert, J.; Esteve, M.D.; Climent, M.D.; Palau, D.; Gómez, I. Early defoliation reduces cluster compactness and improves grape composition in Mandó, an autochthonous cultivar of Vitis vinifera from southeastern Spain. Sci. Hortic. 2014, 167, 71–75. [Google Scholar] [CrossRef]
- Ribereau-Gayon, P. The anthocyanins of the genus Vitis. Application to the differentiation of wines. C. r. Hebd. Seances Acad. Sci. 1960, 250, 591–593. [Google Scholar]
- Mira de Orduña, R. Climate change associated effects on grape and wine quality and production. Food Res. Int. 2010, 43, 1844–1855. [Google Scholar] [CrossRef]
- Castellarin, S.D.; Bavaresco, L.; Falginella, L.; Goncalves, M.I.V.Z.; Di Gaspero, G. Phenolics in Grape Berry and Key Antioxidants; Bentham Science Publishers: Sharjah, United Arab Emirates, 2012; ISBN 978-1-60805-360-5. [Google Scholar]
- Katalinić, V.; Možina, S.S.; Skroza, D.; Generalić, I.; Abramovič, H.; Miloš, M.; Ljubenkov, I.; Piskernik, S.; Pezo, I.; Terpinc, P.; et al. Polyphenolic profile, antioxidant properties and antimicrobial activity of grape skin extracts of 14 Vitis vinifera varieties grown in Dalmatia (Croatia). Food Chem. 2010, 119, 715–723. [Google Scholar] [CrossRef]
- Lorenz, D.H.; Eichhorn, K.W.; Bleiholder, H.; Klose, R.; Meier, U.; Weber, E. Growth Stages of the Grapevine: Phenological growth stages of the grapevine (Vitis vinifera L. ssp. vinifera)—Codes and descriptions according to the extended BBCH scale†. Aust. J. Grape Wine Res. 1995, 1, 100–103. [Google Scholar] [CrossRef]
- Rustioni, L.; Maghradze, D.; Popescu, C.F.; Cola, G.; Abashidze, E.; Aroutiounian, R.; Brazao, J.; Coletti, S.; Cornea, V.; Dejeu, L.; et al. First results of the European grapevine collections’ collaborative network: Validation of a standard eno-carpological phenotyping method. Vitis 2014, 53, 219–226. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Nagel, C.W.; Wulf, L.W. Changes in the Anthocyanins, Flavonoids and Hydroxycinnamic Acid Esters during Fermentation and Aging of Merlot and Cabernet Sauvignon. Am. J. Enol. Vitic. 1979, 30, 111–116. [Google Scholar]
- Shih, M.-H.; Su, Y.-S.; Wu, C.-L. Syntheses of Aromatic Substituted Hydrazino-thiazole Derivatives to Clarify Structural Characterization and Antioxidant Activity between 3-Arylsydnonyl and Aryl Substituted Hydrazino-thiazoles. Chem. Pharm. Bull. (Tokyo) 2007, 55, 1126–1135. [Google Scholar] [CrossRef] [Green Version]
- OIV. Compendium of International Methods of Analysis of Wines and Musts (2 vol.); OIV: Paris, France, 2017. [Google Scholar]
- Burns, J.; Mullen, W.; Landrault, N.; Teissedre, P.-L.; Lean, M.E.J.; Crozier, A. Variations in the Profile and Content of Anthocyanins in Wines Made from Cabernet Sauvignon and Hybrid Grapes. J. Agric. Food Chem. 2002, 50, 4096–4102. [Google Scholar] [CrossRef]
- Ryan, J.-M.; Revilla, E. Anthocyanin composition of Cabernet Sauvignon and Tempranillo grapes at different stages of ripening. J. Agric. Food Chem. 2003, 51, 3372–3378. [Google Scholar] [CrossRef]
- Radovanović, B.; Radovanović, A. Free radical scavenging activity and anthocyanin profile of Cabernet Sauvignon wines from the Balkan region. Molecules 2010, 15, 4213–4226. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, R.; Xu, Z.; Wang, J.; Chen, T.; Chen, F.; Wang, Z. Identification and quantification of anthocyanins in Kyoho grape juice-making pomace, Cabernet Sauvignon grape winemaking pomace and their fresh skin. J. Sci. Food Agric. 2013, 93, 1404–1411. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Wang, S. Synthesis and antimicrobial activities of novel 1H-dibenzo[a,c]carbazoles from dehydroabietic acid. Eur. J. Med. Chem. 2010, 45, 4692–4696. [Google Scholar] [CrossRef] [PubMed]
- Pavić, V.; Flačer, D.; Jakovljević, M.; Molnar, M.; Jokić, S. Assessment of Total Phenolic Content, In Vitro Antioxidant and Antibacterial Activity of Ruta graveolens L. Extracts Obtained by Choline Chloride Based Natural Deep Eutectic Solvents. Plants 2019, 8. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the grape skin extracts are available from the authors, except for Blaufränkisch variety from 2015. |


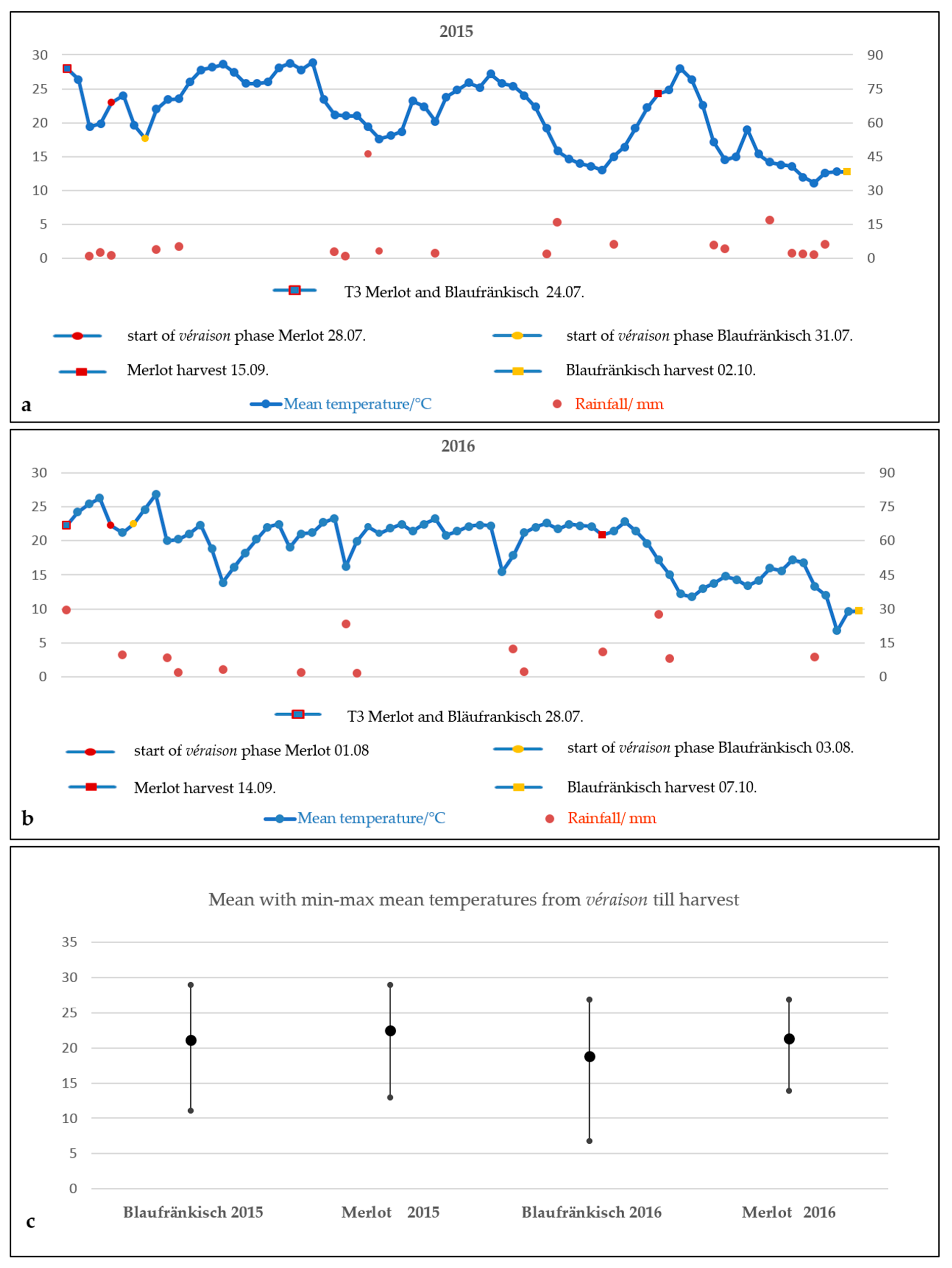{kind=link}
| Vitis vinifera L. variety | Year | Treatment | TPC (mgGAE gskin−1) | TAC (mgMAE gskin−1) | DPPH Radical Scavenging Activity (%) |
|---|---|---|---|---|---|
| Blaufränkisch | 2015 | T1 | 12.79 ± 0.59aAc | 2.06 ± 0.16aAb | 54.03 ± 0.53bAb |
| T2 | 18.2 ± 0.06aAa | 1.93 ± 0.11aAb | 56.52 ± 0.91bAa | ||
| T3 | 17.26 ± 0.53aAb | 2.37 ± 0.09aAa | 55.86 ± 0.31bAa | ||
| 2016 | T1 | 12.86 ± 0.26aAc | 1.98 ± 0.09bAb | 56.34 ± 0.84aAb | |
| T2 | 17.12 ± 1.02aAa | 1.72 ± 0.03bAc | 57.69 ± 0.85aAa | ||
| T3 | 15.61 ± 0.18aAb | 2.35 ± 0.11aAa | 57.06 ± 0.08aAa | ||
| Merlot | 2015 | T1 | 12.85 ± 0.22aAb | 1.60 ± 0.06aBb | 51.97 ± 0.34bBc |
| T2 | 14.97 ± 0.31aBa | 1.84 ± 0.04aAa | 57.18 ± 0.49aAa | ||
| T3 | 14.96 ± 0.63aBa | 1.62 ± 0.05aBb | 54.04 ± 0.97bBb | ||
| 2016 | T1 | 7.73 ± 0.33bBb | 1.02 ± 0.07bAb | 52.86 ± 1.55aBc | |
| T2 | 9.29 ± 0.27bBa | 1.32 ± 0.04bAa | 57.50 ± 0.63aAa | ||
| T3 | 8.36 ± 0.13bBa | 1.09 ± 0.12bAb | 56.59 ± 0.30aBb |
| Skin anthocyanins (mgMAE g skin−1) | Blaufränkisch | |||||
|---|---|---|---|---|---|---|
| Year | 2015 | 2016 | ||||
| Treatment | T1 | T2 | T3 | T1 | T2 | T3 |
| Delphinidin-3-glucoside | n.d. | n.d. | n.d. | 0.0665 ± 0.0102 a | 0.0498 ± 0.0058 b | 0.059 ± 0.0095 a,b |
| Cyanidin-3-glucoside | n.d. | n.d. | n.d. | 0.0312 ± 0.0107 a | 0.0337 ± 0.0095 a | 0.0403 ± 0.0101 a |
| Petunidin-3-glucoside | n.d. | n.d. | n.d. | 0.0867 ± 0.0069 a | 0.0605 ± 0.0048 b | 0.0762 ± 0.0071 c |
| Peonidin-3-glucoside | n.d. | n.d. | n.d. | 0.3372 ± 0.0585 a | 0.3098 ± 0.0656 a | 0.3961 ± 0.0587 a |
| Malvidin-3-glucoside | n.d. | n.d. | n.d. | 0.9323 ± 0.0547 a | 0.5611 ± 0.0459 b | 0.8167 ± 0.1464 a |
| Petunidin-3-(acetyl)-glucoside | n.d. | n.d. | n.d. | 0.0005 ± 0.0004 b | 0.0005 ± 0.000 b | 0.0009 ± 0.0004 a |
| Delphinidin-3-O-(6-p-(coumaroyl)-glucoside | n.d. | n.d. | n.d. | 0.0028 ± 0.0013 b | 0.0022 ± 0.0001 b | 0.0031 ± 0.0005 a |
| Peonidin-3-(acetyl)-glucoside | n.d. | n.d. | n.d. | 0.0021 ± 0.0002 a | 0.0016 ± 0.0003 b | 0.0023 ± 0.0005 a |
| Malvidin-3-(acetyl)-glucoside | n.d. | n.d. | n.d. | 0.0120 ± 0.0013 a | 0.0059 ± 0.0012 b | 0.0110 ± 0.0036 a |
| Cyanidin-3-O-(6″-p-(coumaroyl)-glucoside | n.d. | n.d. | n.d. | 0.0035 ± 0.0003 a | 0.0025 ± 0.0004 b | 0.0039 ± 0.0006 a |
| Petunidin-3-O-(6-p-(coumaroyl)-glucoside | n.d. | n.d. | n.d. | 0.0041 ± 0.0023 b | 0.0034 ± 0.0001 b | 0.0047 ± 0.0006 a |
| Malvidin-3-O-(6-p-(coumaroyl)-glucoside (cis isomer) | n.d. | n.d. | n.d. | 0.0029 ± 0.0018 b | 0.0016 ± 0.0001 b | 0.0027 ± 0.0008 a |
| Peonidin-3-O-(6-p-(coumaroyl)-glucoside | n.d. | n.d. | n.d. | 0.0252 ± 0.0005 a | 0.0165 ± 0.001 b | 0.0221 ± 0.0039 a |
| Malvidin-3-O-(6-p-(coumaroyl)-glucoside (trans isomer) | n.d. | n.d. | n.d. | 0.073 ± 0.0108 a | 0.0377 ± 0.0104 b | 0.0638 ± 0.0229 a |
| ∑ c Anthocyanins (with unknown) | n.d. | n.d. | n.d. | 1.5908 ± 0.0237 a | 1.0944 ± 0.0357 b | 1.5144 ± 0.109 a |
| Skin anthocyanins (mgMAE g skin−1) | Merlot | |||||
|---|---|---|---|---|---|---|
| Year | 2015 | 2016 | ||||
| Treatment | T1 | T2 | T3 | T1 | T2 | T3 |
| Delphinidin-3-glucoside | 0.1384 ± 0.0186 a | 0.1382 ± 0.0022 a | 0.1247 ± 0.0004 b | 0.0424 ± 0.0023 b | 0.0633 ± 0.0014 a | 0.0397 ± 0.0114 b |
| Cyanidin-3-glucoside | 0.0207 ± 0.0032 a | 0.0202 ± 0.0048 a | 0.0201 ± 0.0043 a | 0.0115 ± 0.0014 b | 0.0184 ± 0.0035 a | 0.0140 ± 0.0026 b |
| Petunidin-3-glucoside | 0.1347 ± 0.0134 a | 0.1353 ± 0.0032 a | 0.1168 ± 0.0067 b | 0.043 ± 0.0016 b | 0.0674 ± 0.0093 a | 0.0422 ± 0.008 b |
| Peonidin-3-glucoside | 0.0687 ± 0.0138 a | 0.072 ± 0.0143 a | 0.0611 ± 0.0031 a | 0.0457 ± 0.008 b | 0.0753 ± 0.0087 a | 0.0534 ± 0.0066 b |
| Malvidin-3-glucoside | 0.5212 ± 0.0436 a | 0.5293 ± 0.0143 a | 0.4169 ± 0.0507 b | 0.2048 ± 0.0113 b | 0.3318 ± 0.0482 a | 0.1948 ± 0.0197 b |
| Delphinidin-3-(acetyl)-glucoside | 0.0201 ± 0.0029 a | 0.021 ± 0.0023 a | 0.0186 ± 0.0026 a | 0.0051 ± 0.0013 b | 0.0079 ± 0.0014 a | 0.0048 ± 0.0013 b |
| Cyanidin-3-(acetyl)-glucoside | 0.003 ± 0.0004 a | 0.0036 ± 0.0009 a | 0.0032 ± 0.001 a | 0.0008 ± 0.0001 b | 0.0015 ± 0.0004 a | 0.0009 ± 0.0005 b |
| Petunidin-3-(acetyl)-glucoside | 0.0207 ± 0.0026 a | 0.0207 ± 0.0018a,b | 0.0178 ± 0.003 a | 0.0051 ± 0.0019 b | 0.0086 ± 0.0008 a | 0.0052 ± 0.0008 b |
| Delphinidin-3-O-(6-p-(coumaroyl)-glucoside | 0.0305 ± 0.0024 a | 0.0301 ± 0.0015 a | 0.0251 ± 0.004 b | 0.0067 ± 0.0002 b | 0.0104 ± 0.001 a | 0.0062 ± 0.0008 b |
| Peonidin-3-(acetyl)-glucoside | 0.0085 ± 0.0022 a | 0.0084 ± 0.0031 a | 0.0048 ± 0.0031 b | 0.0047 ± 0.0006 b | 0.0065 ± 0.0025 a | 0.0041 ± 0.0016 b |
| Malvidin-3-(acetyl)-glucoside | 0.1077 ± 0.0123 a | 0.093 ± 0.0322 a | 0.0513 ± 0.033 b | 0.0338 ± 0.0028 b | 0.0558 ± 0.0072 a | 0.0298 ± 0.0035 b |
| Cyanidin-3-O-(6″-p-(coumaroyl)-glucoside | 0.0095 ± 0.0013 b | 0.015 ± 0.0053 a | 0.0127 ± 0.0073 a | 0.0052 ± 0.0008 b | 0.0081 ± 0.0009 a | 0.0052 ± 0.0006 b |
| Petunidin-3-O-(6-p-(coumaroyl)-glucoside | 0.0298 ± 0.0022 a | 0.029 ± 0.0012 a | 0.0184 ± 0.0089 b | 0.0068 ± 0.0005 b | 0.0107 ± 0.0012 a | 0.006 ± 0.0006 b |
| Malvidin-3-O-(6-p-(coumaroyl)-glucoside (cis isomer) | 0.0037 ± 0.0004 a | 0.0041 ± 0.0002 a | 0.0022 ± 0.0013 b | 0.0016 ± 0.0003 b | 0.0025 ± 0.0003 a | 0.0013 ± 0.0004 b |
| Peonidin-3-O-(6-p-(coumaroyl)-glucoside | 0.0203 ± 0.0019 a | 0.0226 ± 0.0032 a | 0.0479 ± 0.061 a | 0.0116 ± 0.0015 b | 0.0181 ± 0.0004 a | 0.0098 ± 0.0011 c |
| Malvidin-3-O-(6-p-(coumaroyl)-glucoside (trans isomer) | 0.1753 ± 0.0157 a | 0.166 ± 0.0069 a | 0.097 ± 0.057 b | 0.0455 ± 0.0013 b | 0.0746 ± 0.0047 a | 0.0365 ± 0.0062 c |
| ∑ c Anthocyanins (with unknown) | 1.3222 ± 0.1247 a | 1.318 ± 0.0299 a | 1.0471 ± 0.0597 b | 0.4783 ± 0.0241 b | 0.7667 ± 0.0806 a | 0.4572 ± 0.0475 b |
| Vitis vinifera L. variety | Year | Treatment | MIC (mg mL−1) | |||
|---|---|---|---|---|---|---|
| E. coli | P. aeruginosa | B. subtilis | S. aureus | |||
| Blaufränkisch | 2015 | T1 | 12.79 ± 0.52aAa | 6.23 ± 0.49bAa | 12.46 ± 0.99aAa | 12.46 ± 0.79bAa |
| T2 | 9.15 ± 0.16aAc | 4.71 ± 0.24bAc | 9.37 ± 0.45aAc | 9.61 ± 0.79bAc | ||
| T3 | 9.96 ± 0.34aAb | 5.05 ± 0.11bAb | 10.06 ± 0.24aAb | 9.96 ± 0.34bAb | ||
| 2016 | T1 | 12.79 ± 0.53aAa | 12.79 ± 0.54 aAa | 12.46 ± 0.10aAa | 25.93 ± 0.65aAa | |
| T2 | 9.15 ± 0.17aAc | 9.15 ± 0.18aAc | 9.37 ± 0.46aAc | 18.22 ± 0.29aAc | ||
| T3 | 9.96 ± 0.35aAb | 9.96 ± 0.36aAb | 10.06 ± 0.25aAb | 19.93 ± 0.69aAb | ||
| Merlot | 2015 | T1 | 7.72 ± 0.66aBa | 4.03 ± 0.36aBa | 7.73 ± 0.67aBa | 7.61 ± 0.58bBa |
| T2 | 6.88 ± 0.68aBb | 3.27 ± 0.39aBb | 6.84 ± 0.51aBb | 7.14 ± 0.12bBb | ||
| T3 | 7.31 ± 0.63aBa,b | 3.82 ± 0.49aBa | 7.31 ± 0.63aBa,b | 7.28 ± 0.25bBa,b | ||
| 2016 | T1 | 7.74 ± 0.67aBa | 4.13 ± 0.37aBa | 7.83± 0.68aBa | 15.78 ± 1.29aBa | |
| T2 | 6.98 ± 0.69aBa | 3.47 ± 0.40aBb | 6.94 ± 0.52aBa | 13.75 ± 1.26aBb | ||
| T3 | 7.41 ± 0.64aBa,b | 3.92 ± 0.50aBa | 7.41 ± 0.64aBa,b | 15.18 ± 0.37aBa | ||
| Amikacine sulfate | 0.000016 | 0.00003 | 0.000016 | 0.000008 | ||
| Month | Mean Monthly Temperature, °C | Mean Min-Max Temperature, °C | Rainfall, mm | |||
|---|---|---|---|---|---|---|
| 2015 | 2016 | 2015 | 2016 | 2015 | 2016 | |
| March | 7.8 | 7.9 | 2.7–15.0 | 3.3–17.6 | 48.5 | 82.7 |
| April | 12.7 | 13.7 | 5.9–19.7 | 5.8–19.2 | 18.2 | 46.5 |
| May | 18.2 | 16.6 | 12.0–25.4 | 9.4–23.8 | 130.8 | 72.1 |
| June | 21.3 | 21.4 | 13.5–27.0 | 17.1–28.4 | 16.8 | 92.7 |
| July | 25.0 | 23.3 | 17.6–30.0 | 14.0–28.4 | 12.4 | 125.2 |
| August | 24.2 | 21.0 | 17.6–28.9 | 16.9–26.8 | 64.9 | 51.0 |
| September | 18.2 | 18.4 | 11.1–28.0 | 11.8–22.8 | 63.1 | 61.5 |
| October | 11.3 | 10.4 | 11.1–17.7 | 5.6–17.2 | 114.6 | 66.0 |
| Mean temp., (°C) | 17.33 | 16.58 | ||||
| Cumulative rainfall, mm | 469.3 | 597.7 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavić, V.; Kujundžić, T.; Kopić, M.; Jukić, V.; Braun, U.; Schwander, F.; Drenjančević, M. Effects of Defoliation on Phenolic Concentrations, Antioxidant and Antibacterial Activity of Grape Skin Extracts of the Varieties Blaufränkisch and Merlot (Vitis vinifera L.). Molecules 2019, 24, 2444. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24132444
Pavić V, Kujundžić T, Kopić M, Jukić V, Braun U, Schwander F, Drenjančević M. Effects of Defoliation on Phenolic Concentrations, Antioxidant and Antibacterial Activity of Grape Skin Extracts of the Varieties Blaufränkisch and Merlot (Vitis vinifera L.). Molecules. 2019; 24(13):2444. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24132444
Chicago/Turabian StylePavić, Valentina, Toni Kujundžić, Marina Kopić, Vladimir Jukić, Ulrike Braun, Florian Schwander, and Mato Drenjančević. 2019. "Effects of Defoliation on Phenolic Concentrations, Antioxidant and Antibacterial Activity of Grape Skin Extracts of the Varieties Blaufränkisch and Merlot (Vitis vinifera L.)" Molecules 24, no. 13: 2444. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24132444