
Tea Seed Oil Prevents Obesity, Reduces Physical Fatigue, and Improves Exercise Performance in High-Fat-Diet-Induced Obese Ovariectomized Mice

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
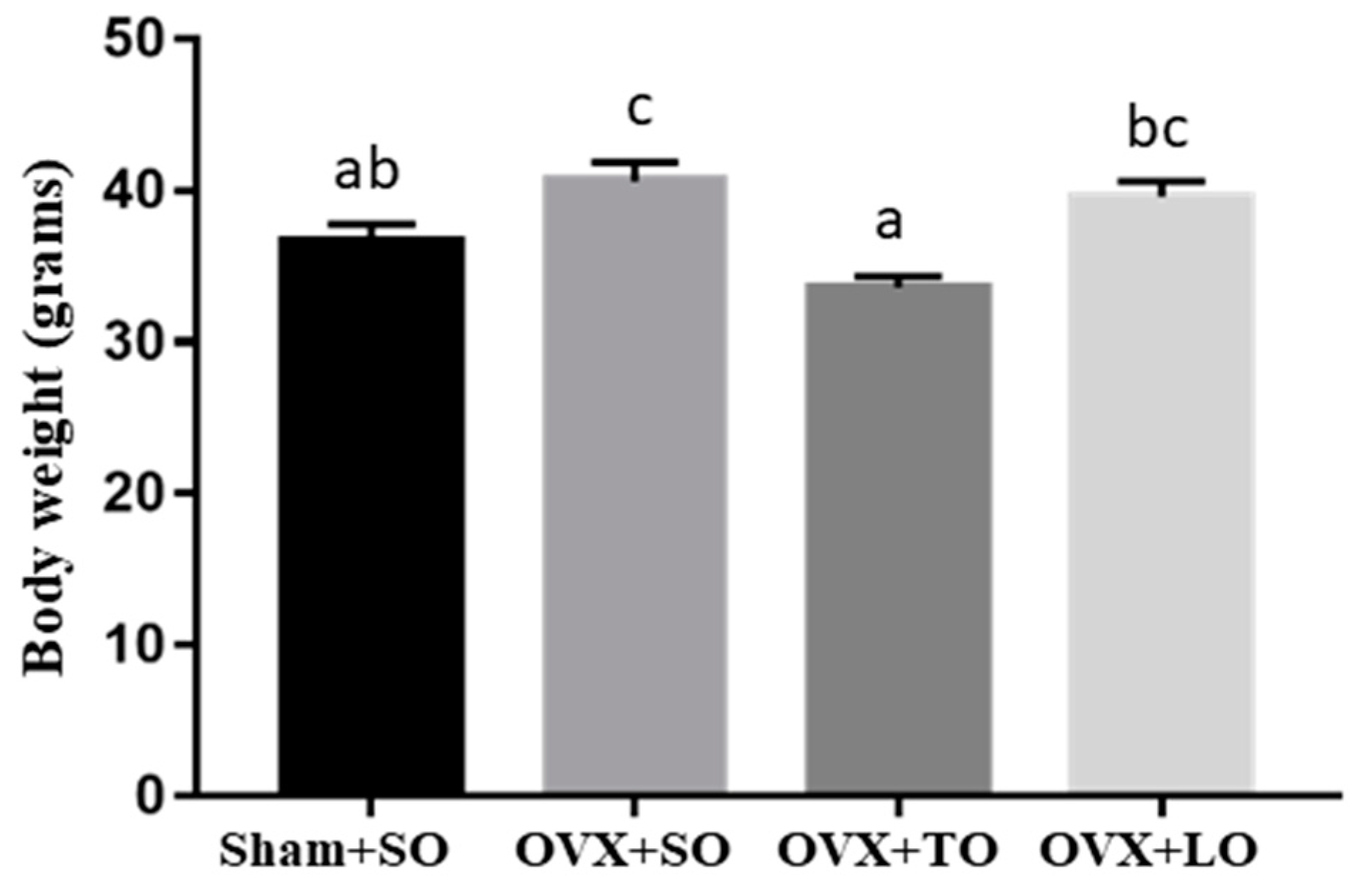
2.1. Effects of SO, TO, and LO on Food Intake, Body Weight, and Tissue Weights of OVX and HFD-Induced Obese Mice
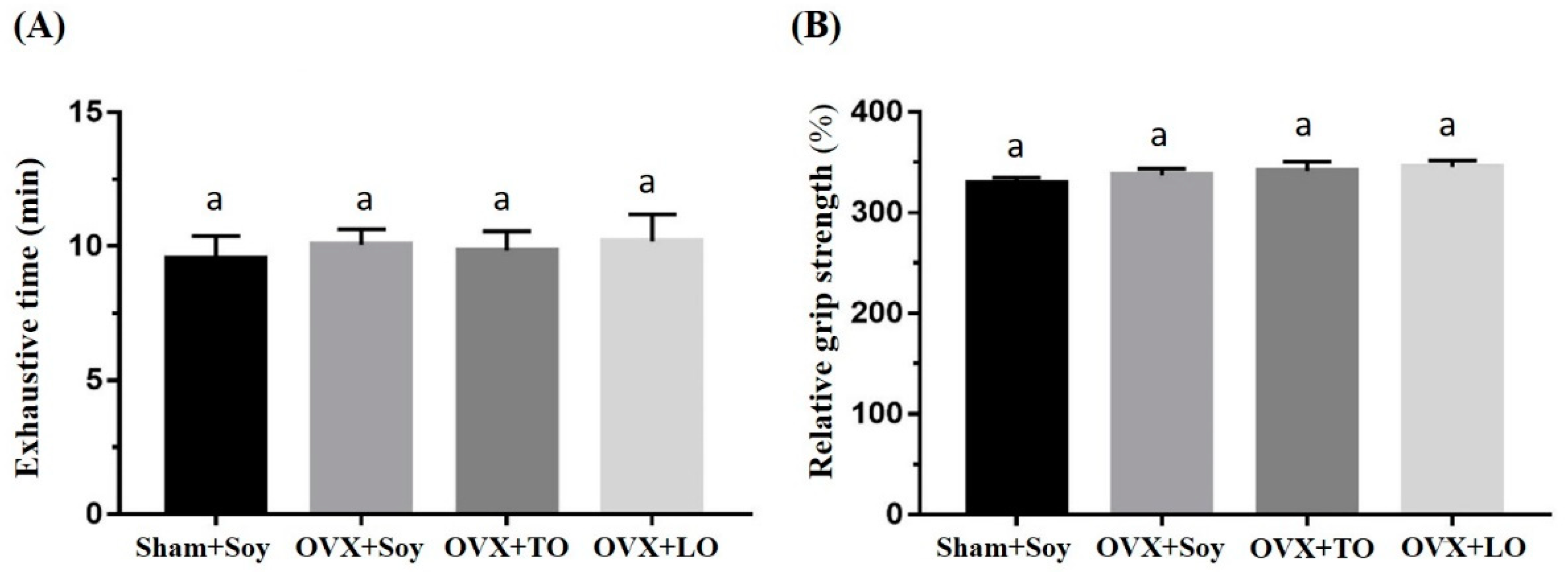
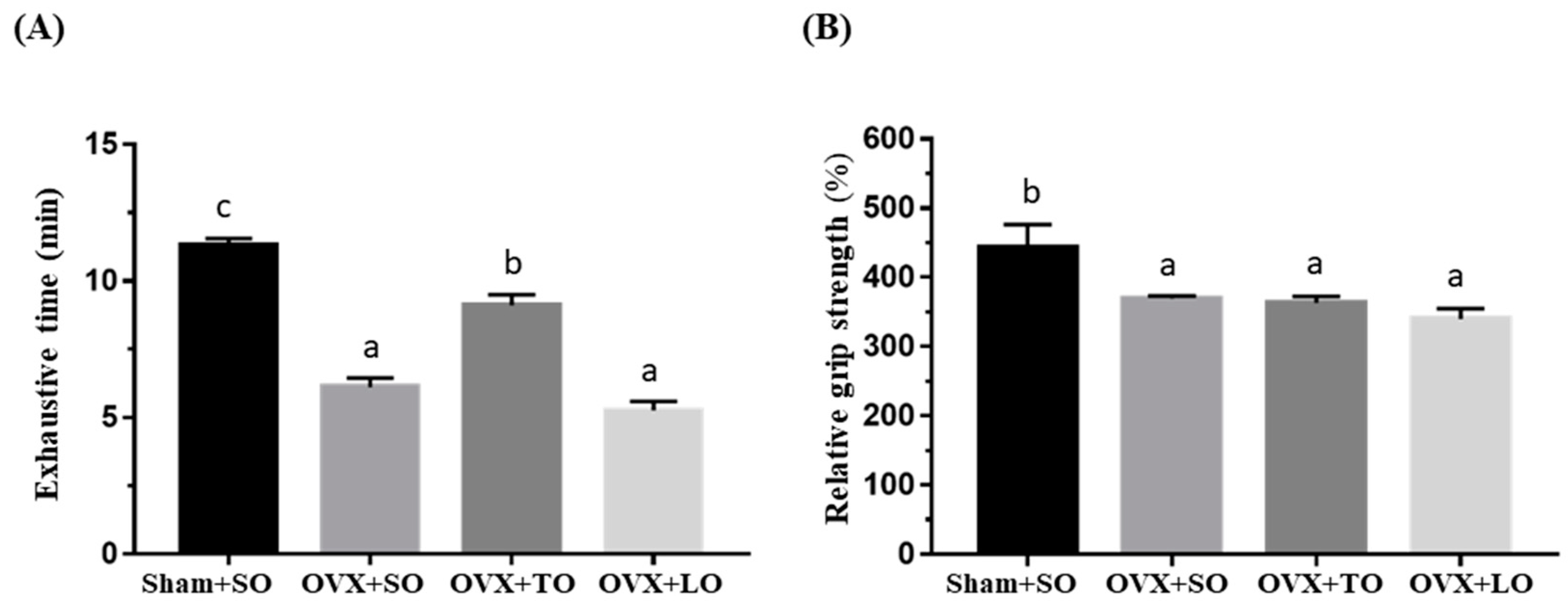
2.2. Effects of SO, TO, and LO on Exhaustive Swimming Time and Forelimb Grip Strength in OVX and HFD-Induced Obese Mice
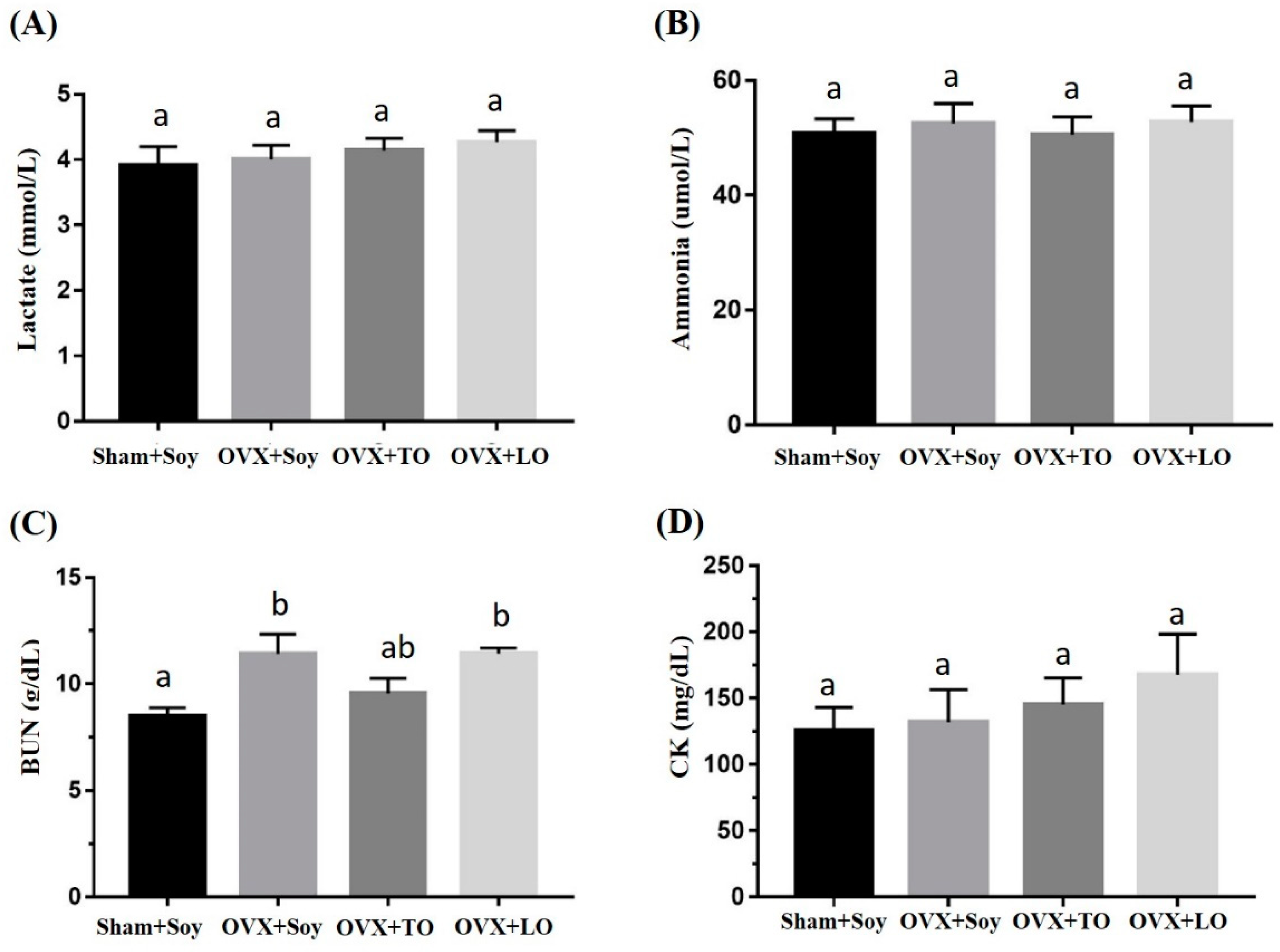
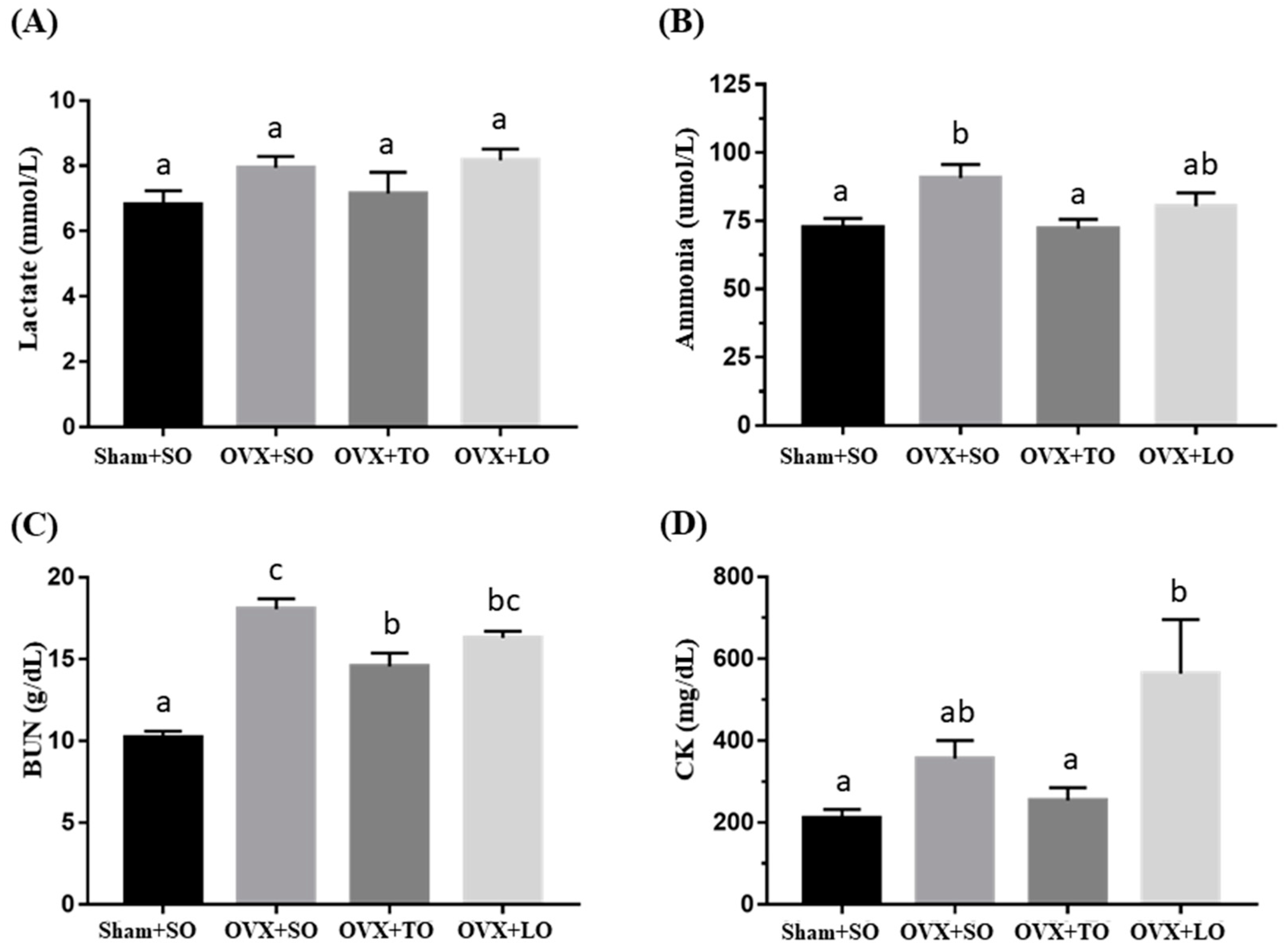
2.3. Effects of SO, TO, and LO on Lactate, Ammonia, BUN, and CK Levels After the 10 Minutes Swimming Test in OVX and HFD-Induced Obese Mice
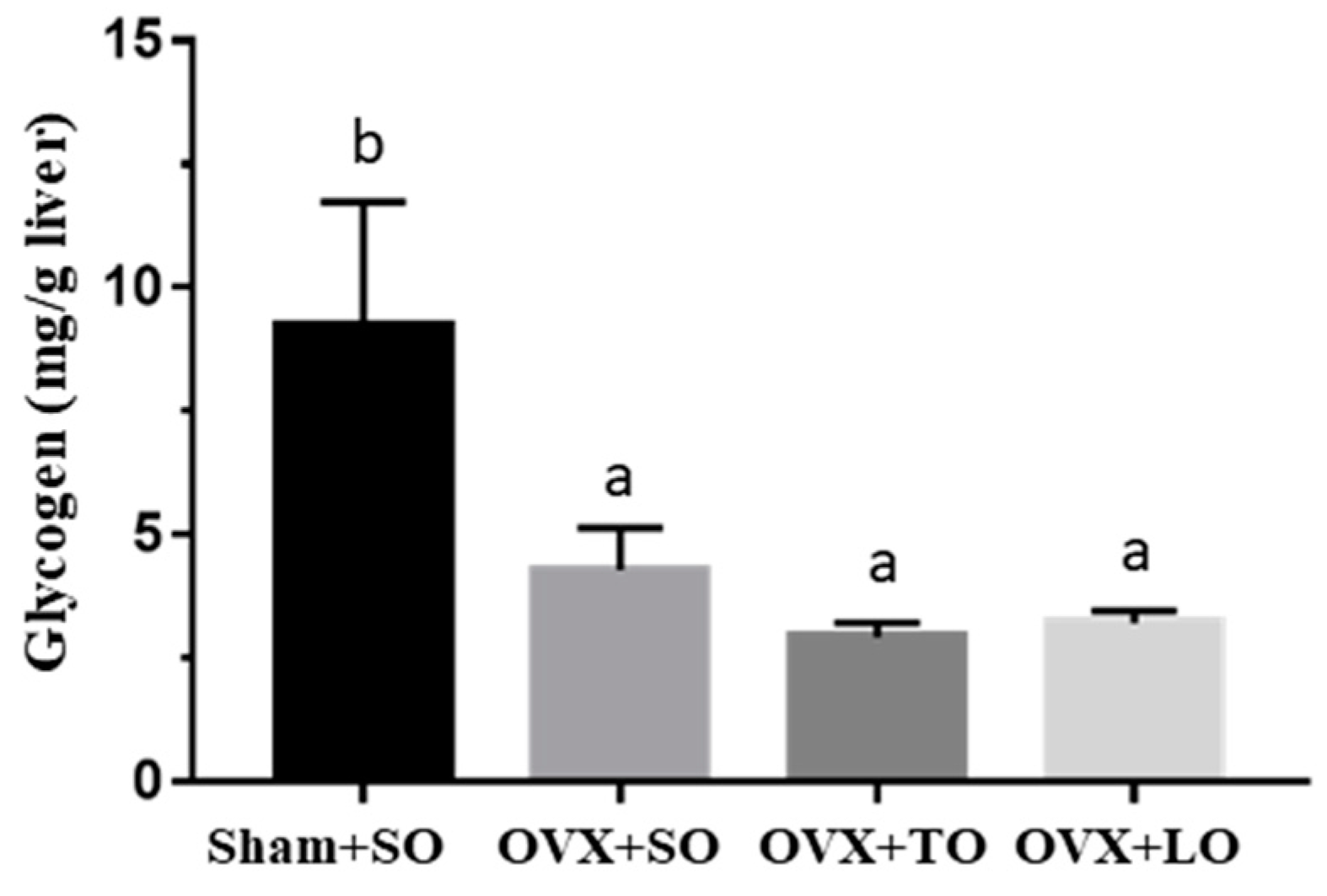
2.4. Effects of SO, TO, and LO on Liver Glycogen in OVX and HFD-Induced Obese Mice
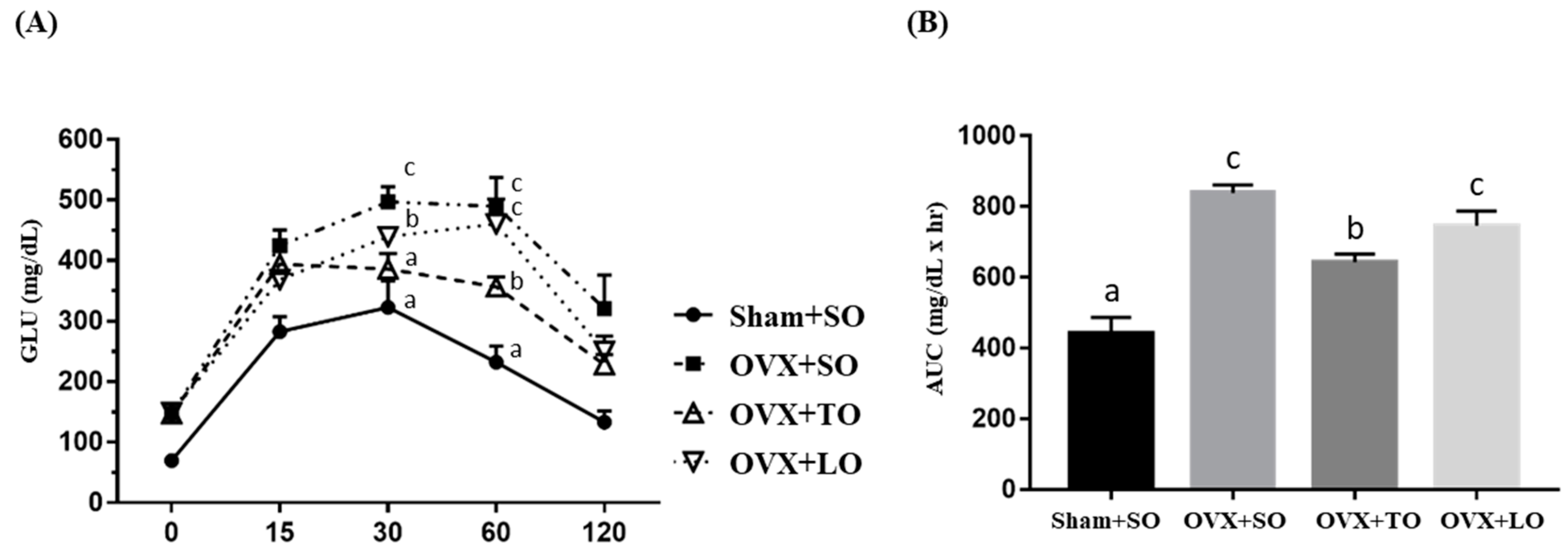
2.5. Effects of SO, TO, and LO on Oral Glucose Tolerance and AUC in OVX and HFD-Induced Obese Mice
2.6. Effects of SO, TO, and LO on Biochemical Variables in OVX and HFD-Induced Obese Mice at the End of the Experiment
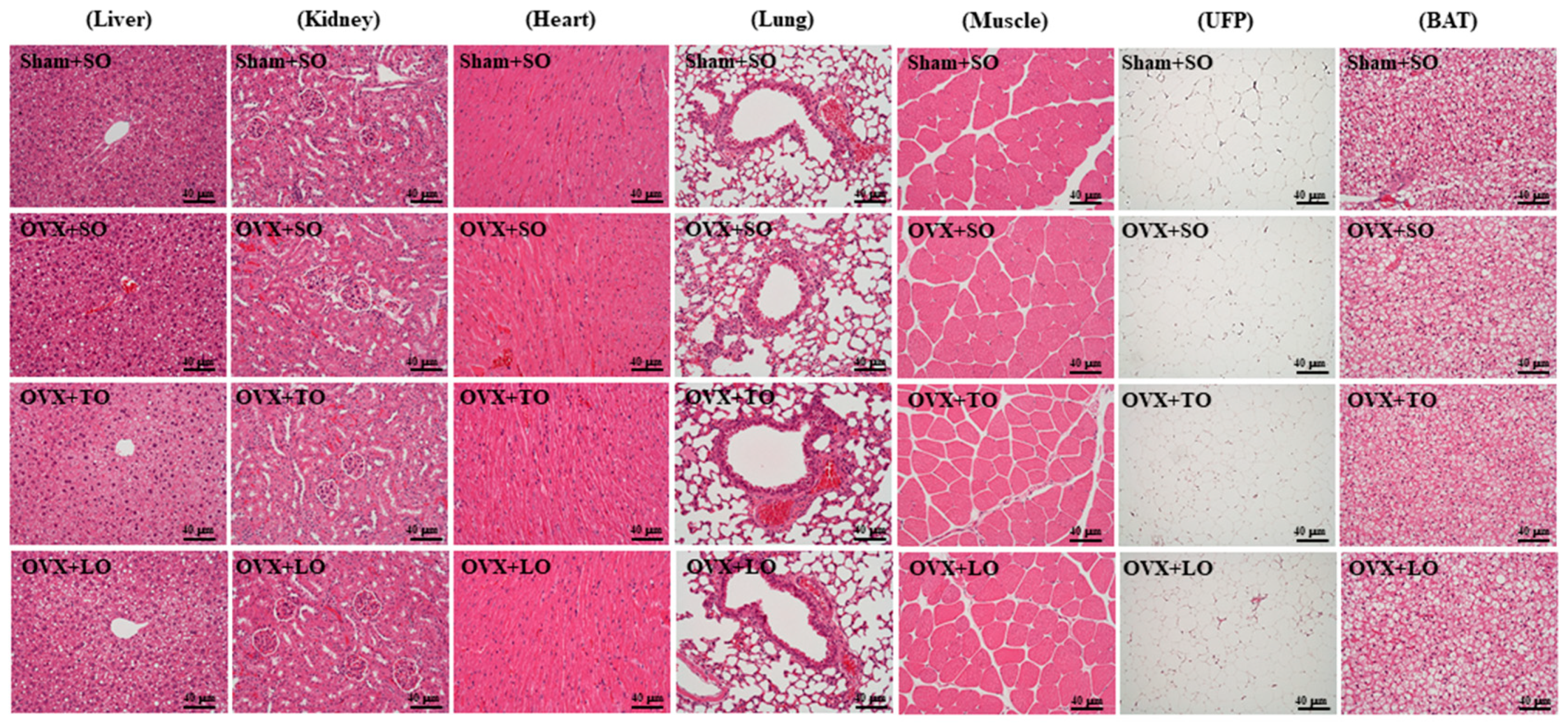
2.7. Effects of SO, TO, and LO on Histopathology of Tissues at the End of the Experiment in OVX and HFD-Induced Obese Mice
3. Discussion
4. Materials and Methods
4.1. Preparation of Oils
4.2. Animals and Experiment Design
4.3. Exhaustive Swimming Test
4.4. Forelimb Grip Strength
4.5. Fatigue-Associated Biochemical Indices
4.6. Tissue Glycogen Determination
4.7. Histological Staining of Tissues
4.8. Oral Gavage Glucose Tolerance Test
4.9. Blood Biochemical Assessments
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davis, S.R.; Castelo-Branco, C.; Chedraui, P.; Lumsden, M.A.; Nappi, R.E.; Shah, D.; Villaseca, P.; Writing Group of the International Menopause Society for World Menopause Day 2012. Understanding weight gain at menopause. Climacteric 2012, 15, 419–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaaja, R.J. Metabolic syndrome and the menopause. Menopause Int. 2008, 14, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Abdelmalek, M.F. Nonalcoholic fatty liver disease in women. Womens Health 2009, 5, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Barrett-Connor, E. Menopause, atherosclerosis, and coronary artery disease. Curr. Opin. Pharmacol. 2013, 13, 186–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemieux, S.; Prud’homme, D.; Bouchard, C.; Tremblay, A.; Després, J.P. Sex differences in the relation of visceral adipose tissue accumulation to total body fatness. Am. J. Clin. Nutr. 1993, 58, 463–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riant, E.; Waget, A.; Cogo, H.; Arnal, J.F.; Burcelin, R.; Gourdy, P. Estrogens protect against high-fat diet-induced insulin resistance and glucose intolerance in mice. Endocrinology 2009, 150, 2109–2117. [Google Scholar] [CrossRef] [PubMed]
- Mittendorfer, B.; Patterson, B.W.; Klein, S. Effect of sex and obesity on basal VLDL-triacylglycerol kinetics. Am. J. Clin. Nutr. 2003, 77, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Roger, V.L.; Go, A.S.; Lloyd-Jones, D.M.; Adams, R.J.; Berry, J.D.; Brown, T.M.; Carnethon, M.R.; Dai, S.; de Simone, G.; Ford, E.S.; et al. American Heart Association Statistics Committee and Stroke Statistics Subcommittee. Heart disease and stroke statistics—2011 update: A report from the American Heart Association. Circulation 2011, 123, e18–e209. [Google Scholar] [CrossRef] [PubMed]
- Keil, J.E.; Sutherland, S.E.; Knapp, R.G.; Lackland, D.T.; Gazes, P.C.; Tyroler, H.A. Mortality rates and risk factors for coronary disease in black as compared with white men and women. N. Engl. J. Med. 1993, 329, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, K.R.; Redden, D.T.; Wang, C.; Westfall, A.O.; Allison, D.B. Years of life lost due to obesity. JAMA 2003, 289, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.N.; Emfinger, C.H.; Overton, M.H.; Hill, S.; Ramaswamy, T.S.; Cappel, D.A.; Wu, K.; Fazio, S.; McDonald, W.H.; Hachey, D.L.; et al. Obesity and altered glucose metabolism impact HDL composition in CETP transgenic mice: A role for ovarian hormones. J. Lipid Res. 2012, 53, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Ribas, V.; Nguyen, M.T.; Henstridge, D.C.; Nguyen, A.K.; Beaven, S.W.; Watt, M.J.; Hevener, A.L. Impaired oxidative metabolism and inflammation are associated with insulin resistance in ERalphadeficient mice. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E304–E319. [Google Scholar] [CrossRef] [PubMed]
- Bryzgalova, G.; Gao, H.; Ahren, B.; Zierath, J.R.; Galuska, D.; Steiler, T.L.; Dahlman-Wright, K.; Nilsson, S.; Gustafsson, J.A.; Efendic, S.; et al. Evidence that oestrogen receptoralpha plays an important role in the regulation of glucose homeostasis in mice: Insulin sensitivity in the liver. Diabetologia 2006, 49, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.E.; Thorburn, A.W.; Britt, K.L.; Hewitt, K.N.; Wreford, N.G.; Proietto, J.; Oz, O.K.; Leury, B.J.; Robertson, K.M.; Yao, S.; et al. Aromatase-deficient (ArKO) mice have a phenotype of increased adiposity. Proc. Natl. Acad. Sci. USA 2000, 97, 12735–12740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshaies, Y.; Dagnault, A.; Lalonde, J.; Richard, D. Interaction of corticosterone and gonadal steroids on lipid deposition in the female rat. Am. J. Physiol. 1997, 36, E355–E363. [Google Scholar] [CrossRef] [PubMed]
- Picard, F.; Deshaies, Y.; Lalonde, J.; Samson, P.; Labrie, C.; Bélanger, A.; Labrie, F.; Richard, D. Effects of the estrogen antagonist EM-652.HCL on energy balance and lipid metabolism in ovariectomized rats. Int. J. Obes. 2000, 24, 830–840. [Google Scholar] [CrossRef]
- Rossouw, J.E.; Anderson, G.L.; Prentice, R.L.; LaCroix, A.Z.; Kooperberg, C.; Stefanick, M.L.; Jackson, R.D.; Beresford, S.A.; Howard, B.V.; Johnson, K.C.; et al. Risks and benefits of estrogen plus progestin in healthy postmenopausal women: Principal results From the Women’s Health Initiative randomized controlled trial. JAMA 2002, 288, 321–333. [Google Scholar] [PubMed]
- Terao, J. Dietary flavonoids as antioxidants. Forum Nutr. 2009, 61, 87–94. [Google Scholar] [PubMed]
- Meydani, M.; Hasan, S.T. Dietary polyphenols and obesity. Nutrients 2010, 2, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhou, G.Y.; Zhang, H.Y.; Liu, J.A. Research progress on the health function of tea oil. J. Med. Plants Res. 2011, 5, 485–489. [Google Scholar]
- Alkhatib, A.; Tsang, C.; Tuomilehto, J. Olive Oil Nutraceuticals in the Prevention and Management of Diabetes: From Molecules to Lifestyle. Int. J. Mol. Sci. 2018, 19, 2024. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.T.; Tung, Y.T.; Wu, C.C.; Tu, P.S.; Yen, G.C. Camellia oil (Camellia oleifera Abel.) modifies the composition of gut microbiota and alleviates acetic acid-induced colitis in rats. J. Agric. Food Chem. 2018, 66, 7384–7392. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.H.; Huang, Y.F.; Xie, Z.F. Health functions and prospective of camellia oil. Food Sci. Technol. 2005, 5, 94–96. [Google Scholar]
- Rogers, N.H.; Perfield, J.W., 2nd.; Strissel, K.J.; Obin, M.S.; Greenberg, A.S. Reduced energy expenditure and increased inflammation are early events in the development of ovariectomy-induced obesity. Endocrinology 2009, 150, 2161–2168. [Google Scholar] [CrossRef] [PubMed]
- Vieira Potter, V.J.; Strissel, K.J.; Xie, C.; Chang, E.; Bennett, G.; Defuria, J.; Obin, M.S.; Greenberg, A.S. Adipose tissue inflammation and reduced insulin sensitivity in ovariectomized mice occurs in the absence of increased adiposity. Endocrinology 2012, 153, 4266–4277. [Google Scholar] [CrossRef] [PubMed]
- Terauchi, M.; Hiramitsu, S.; Akiyoshi, M.; Owa, Y.; Kato, K.; Obayashi, S.; Matsushima, E.; Kubota, T. Effects of the kampo formula tokishakuyakusan on headaches and concomitant depression in middle-aged women. Evid. Based Complement. Altern. Med. 2014, 2014, 593560. [Google Scholar] [CrossRef] [PubMed]
- Lurati, A.R. Menopause and Exercise Intolerance. Nurs. Womens Health. 2017, 21, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Bondarev, D.; Laakkonen, E.K.; Finni, T.; Kokko, K.; Kujala, U.M.; Aukee, P.; Kovanen, V.; Sipilä, S. Physical performance in relation to menopause status and physical activity. Menopause 2018, 25, 1432–1441. [Google Scholar] [CrossRef] [PubMed]
- Cabelka, C.A.; Baumann, C.W.; Collins, B.C.; Nash, N.; Le, G.; Lindsay, A.; Spangenburg, E.E.; Lowe, D.A. Effects of ovarian hormones and estrogen receptor α on physical activity and skeletal muscle fatigue in female mice. Exp. Gerontol. 2019, 115, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Hirose, A.; Terauchi, M.; Osaka, Y.; Akiyoshi, M.; Kato, K.; Miyasaka, N. Effect of soy lecithin on fatigue and menopausal symptoms in middle-aged women: A randomized, double-blind, placebo-controlled study. Nutr. J. 2018, 17, 4. [Google Scholar] [CrossRef] [PubMed]
- Panotopoulos, G.; Ruiz, J.C.; Raison, J.; Guy-Grand, B.; Basdevant, A. Menopause, fat and lean distribution in obese women. Maturitas 1996, 25, 11–19. [Google Scholar] [CrossRef]
- Kotani, K.; Tokunaga, K.; Fujioka, S.; Kobatake, T.; Keno, Y.; Yoshida, S.; Shimomura, I.; Tarui, S.; Matsuzawa, Y. Sexual dimorphism of age-related changes in whole-body fat distribution in the obese. Int. J. Obes. Relat. Metab. Disord. 1994, 18, 207–212. [Google Scholar] [PubMed]
- Lovejoy, J.C.; Champagne, C.M.; de Jonge, L.; Xie, H.; Smith, S.R. Increased visceral fat and decreased energy expenditure during the menopausal transition. Int. J. Obes. (Lond.) 2008, 32, 949–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Risérus, U.; Willett, W.C.; Hu, F.B. Dietary fats and prevention of type 2 diabetes. Prog. Lipid Res. 2009, 48, 44–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, M.; McInerney, D.; Owens, D.; Collins, P.; Johnson, A.; Tomkin, G.H. Diabetes and the Mediterranean diet: A beneficial effect of oleic acid on insulin sensitivity, adipocyte glucose transport and endothelium-dependent vasoreactivity. QJM 2000, 93, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Mata, P.; Garrido, J.A.; Ordovas, J.M.; Blazquez, E.; Alvarez-Sala, L.A.; Rubio, M.J.; Alonso, R.; de Oya, M. Effect of dietary monounsaturated fatty acids on plasma lipoproteins and apolipoproteins in women. Am. J. Clin. Nutr. 1992, 56, 77–83. [Google Scholar] [CrossRef] [PubMed]
- McElroy, J.F.; Wade, G.N. Short- and long-term effects of ovariectomy on food intake, body weight, carcass composition, and brown adipose tissue in rats. Physiol. Behav. 1987, 39, 361–365. [Google Scholar] [CrossRef]
- Wang, X.; Cheng, M.; Zhao, M.; Ge, A.; Guo, F.; Zhang, M.; Yang, Y.; Liu, L.; Yang, N. Differential effects of high-fat-diet rich in lard oil or soybean oil on osteopontin expression and inflammation of adipose tissue in diet-induced obese rats. Eur. J. Nutr. 2013, 52, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Piers, L.S.; Walker, K.Z.; Stoney, R.M.; Soares, M.J.; O’Dea, K. The influence of the type of dietary fat on postprandial fat oxidation rates: Monounsaturated (olive oil) vs. saturated fat (cream). Int. J. Obes. Relat. Metab. Disord. 2002, 26, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, J.M.; Pighon, A. NAFLD, estrogens, and physical exercise: The animal model. J. Nutr. Metab. 2012, 2012, 914938. [Google Scholar] [CrossRef] [PubMed]
- Maric, C.; Sandberg, K.; Hinojosa-Laborde, C. Glomerulosclerosis and tubulointerstitial fibrosis are attenuated with 17beta-estradiol in the aging Dahl salt sensitive rat. J. Am. Soc. Nephrol. 2004, 15, 1546–1556. [Google Scholar] [CrossRef] [PubMed]
- Ke, J.Y.; Kliewer, K.L.; Hamad, E.M.; Cole, R.M.; Powell, K.A.; Andridge, R.R.; Straka, S.R.; Yee, L.D.; Belury, M.A. The flavonoid, naringenin, decreases adipose tissue mass and attenuates ovariectomy-associated metabolic disturbances in mice. Nutr. Metab. (Lond.) 2015, 12, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, P.J.; Zhang, Y.H.; Huang, J.X. A study of the effect of tea seed oil on serum lipid in normal adults. Acta Nutr. Sin. 1993, 3, 289–292. [Google Scholar]
- Deng, X.L.; Xie, G.S.; Huang, S.G. Study on the health care oil tea and just the function of lipid. China Oils Fats 2002, 5, 96–98. [Google Scholar]
- Carr, M.C. The emergence of the metabolic syndrome with menopause. J. Clin. Endocrinol. Metab. 2003, 88, 2404–2411. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.D.; Sun, Z. Research progress on natural resources and application of the bioactive substance—Squalene. Chem. Ind. For. Prod. 2004, 24, 107–112. [Google Scholar]
- Wang, Y.; Sun, D.; Chen, H.; Qian, L.; Xu, P. Fatty acid composition and antioxidant activity of tea (Camellia sinensis L.) seed oil extracted by optimized supercritical carbon dioxide. Int. J. Mol. Sci. 2011, 12, 7708–7719. [Google Scholar] [CrossRef] [PubMed]
- Bumrungpert, A.; Pavadhgul, P.; Kalpravidh, R.W. Camellia oil-enriched diet attenuates oxidative stress and inflammatory markers in hypercholesterolemic subjects. J. Med. Food 2016, 19, 895–898. [Google Scholar] [CrossRef] [PubMed]
- Tu, P.S.; Tung, Y.T.; Lee, W.T.; Yen, G.C. Protective effect of camellia oil (Camellia oleifera Abel.) against ethanol-induced acute oxidative injury of the gastric mucosa in mice. J. Agric. Food Chem. 2017, 21, 4932–4941. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.T.; Wu, S.L.; Ho, C.Y.; Huang, S.M.; Cheng, C.L.; Yen, G.C. Beneficial effects of camellia oil (Camellia oleifera Abel.) on ketoprofen-induced gastrointestinal mucosal damage through upregulation of HO-1 and VEGF. J. Agric. Food Chem. 2014, 62, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Kan, N.W.; Ho, C.S.; Chiu, Y.S.; Huang, W.C.; Chen, P.Y.; Tung, Y.T.; Huang, C.C. Effects of resveratrol supplementation and exercise training on the exercise performance in middle-aged mice. Molecules 2016, 21, 661. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C.; Hsu, M.C.; Huang, W.C.; Yang, H.R.; Hou, C.C. Triterpenoid-rich extract from Antrodia camphorata improves physical fatigue and exercise performance in mice. Evid. Based Complement. Altern. Med. 2012, 2012, 364741. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Lin, C.I.; Chiu, C.C.; Lin, Y.T.; Huang, W.K.; Huang, H.Y.; Huang, C.C. Chicken essence improves exercise performance and ameliorates physical fatigue. Nutrients 2014, 6, 2681–2696. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not Available. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Sham+SO | OVX+SO | OVX+TO | OVX+LO |
|---|---|---|---|---|
| Relative liver weight (%) | 4.02 ± 0.16 b | 3.59 ± 0.07 a | 4.09 ± 0.16 b | 3.54 ± 0.11 a |
| Relative kidney weight (%) | 1.03 ± 0.06 a | 0.84 ± 0.02 a | 0.93 ± 0.05 a | 0.86 ± 0.06 a |
| Relative heart weight (%) | 0.44 ± 0.02 b | 0.40 ± 0.01 a | 0.47 ± 0.01 b | 0.42 ± 0.02 a |
| Relative lung weight (%) | 0.56 ± 0.03 b | 0.51 ± 0.03 a | 0.64 ± 0.02 b | 0.54 ± 0.02 a |
| Relative muscle weight (%) | 0.89 ± 0.02 b,c | 0.87 ± 0.03 a,b | 0.98 ± 0.02 c | 0.83 ± 0.03 a |
| Relative BAT weight (%) | 0.23 ± 0.01 c | 0.18 ± 0.00 a | 0.22 ± 0.01 b | 0.19 ± 0.01 a |
| Relative UPF weight (%) | 4.31 ± 0.62 a | 6.07 ± 0.65 b | 4.76 ± 0.50 a,b | 5.72 ± 0.23 a,b |
| Relative TF weight (%) | 7.96 ± 0.28 a,b | 9.41 ± 0.64 b | 7.19 ± 0.32 a | 9.17 ± 0.12 b |
| Parameter | Sham+SO | OVX+SO | OVX+TO | OVX+LO |
|---|---|---|---|---|
| Insulin (μIU/mL) | 1.69 ± 0.11 a | 1.11 ± 0.10 a | 1.63 ± 0.24 a | 1.26 ± 0.17 a |
| AST (U/L) | 116 ± 4 a | 131 ± 8 a | 112 ± 7 a | 138 ± 11 a |
| ALT (U/L) | 39 ± 1 a | 50 ± 2 a,b | 47 ± 4 a,b | 54 ± 4 b |
| TC (mg/dL) | 57 ± 2 a | 60 ± 3 a,b | 59 ± 3 a,b | 68 ± 3 b |
| TG (mg/dL) | 81 ± 3 a | 92 ± 2 b | 89 ± 3 a,b | 104 ± 3 c |
| BUN (g/dL) | 8.5 ± 0.3 a | 11.4 ± 0.8 b | 9.6 ± 0.6 a,b | 11.4 ± 0.2 b |
| Creatinine (mg/dL) | 0.23 ± 0.00 a | 0.28 ± 0.01 b | 0.25 ± 0.01 a,b | 0.24 ± 0.01 a |
| ALB (g/dL) | 3.3 ± 0.0 b | 3.0 ± 0.1 a | 3.1 ± 0.0 a | 3.1 ± 0.0 a |
| TP (g/dL) | 5.3 ± 0.1 a | 5.1 ± 0.0 a | 5.2 ± 0.1 a | 5.3 ± 0.0 a |
| LDH (mg/dL) | 516 ± 20 a | 576 ± 22 a | 540 ± 27 a | 672 ± 87 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tung, Y.-T.; Hsu, Y.-J.; Chien, Y.-W.; Huang, C.-C.; Huang, W.-C.; Chiu, W.-C. Tea Seed Oil Prevents Obesity, Reduces Physical Fatigue, and Improves Exercise Performance in High-Fat-Diet-Induced Obese Ovariectomized Mice. Molecules 2019, 24, 980. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24050980
Tung Y-T, Hsu Y-J, Chien Y-W, Huang C-C, Huang W-C, Chiu W-C. Tea Seed Oil Prevents Obesity, Reduces Physical Fatigue, and Improves Exercise Performance in High-Fat-Diet-Induced Obese Ovariectomized Mice. Molecules. 2019; 24(5):980. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24050980
Chicago/Turabian StyleTung, Yu-Tang, Yi-Ju Hsu, Yi-Wen Chien, Chi-Chang Huang, Wen-Ching Huang, and Wan-Chun Chiu. 2019. "Tea Seed Oil Prevents Obesity, Reduces Physical Fatigue, and Improves Exercise Performance in High-Fat-Diet-Induced Obese Ovariectomized Mice" Molecules 24, no. 5: 980. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24050980