Vasodilation Elicited by Isoxsuprine, Identified by High-Throughput Virtual Screening of Compound Libraries, Involves Activation of the NO/cGMP and H2S/KATP Pathways and Blockade of α1-Adrenoceptors and Calcium Channels

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
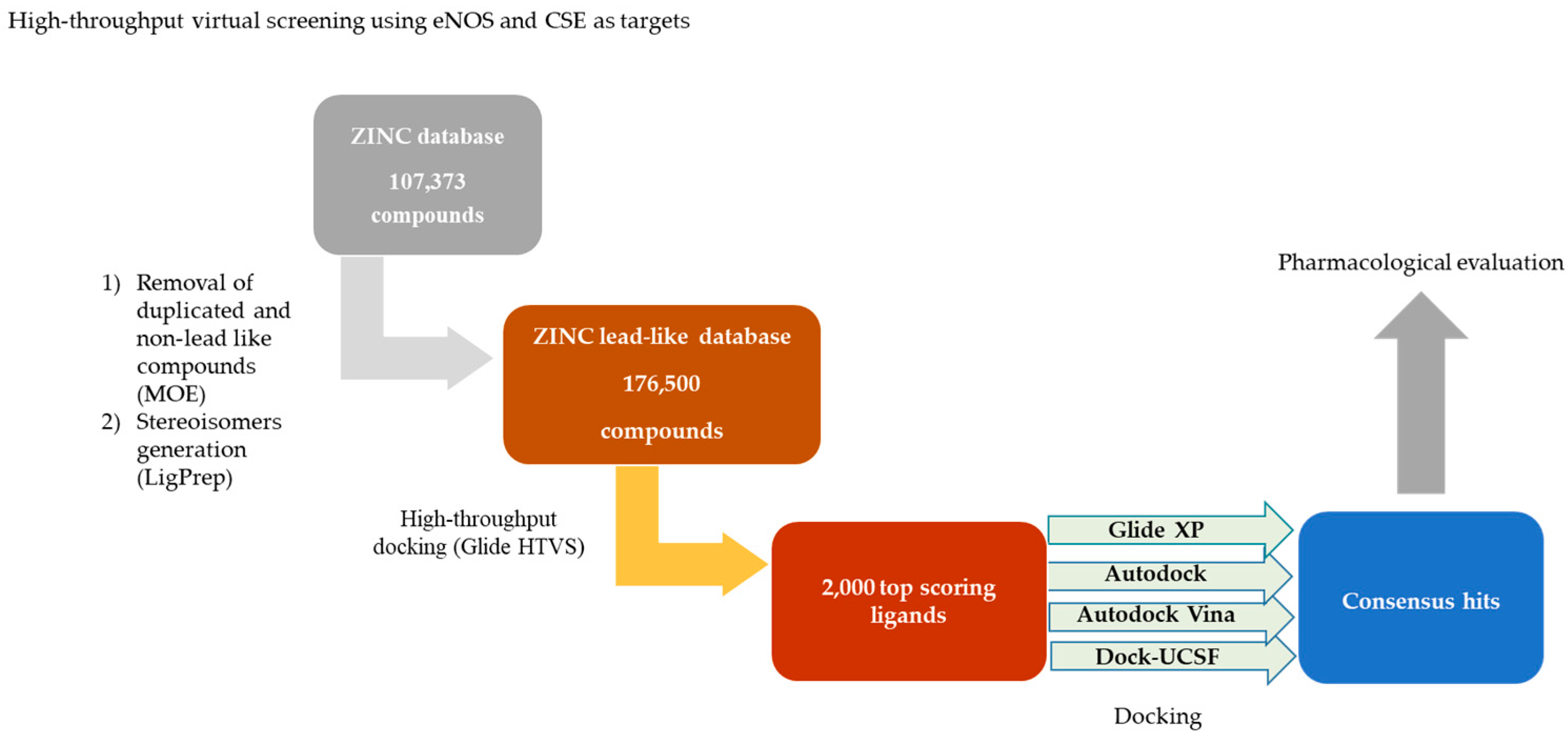
2.1. Virtual Screening
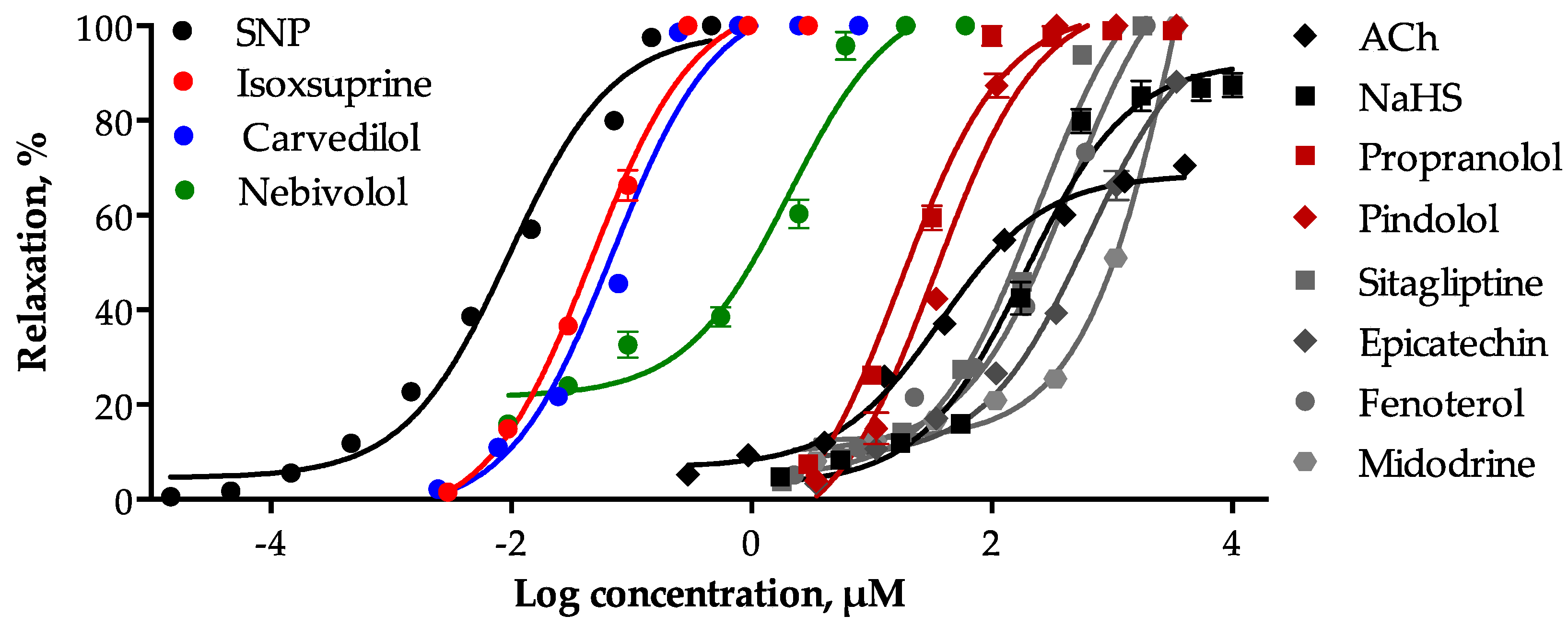
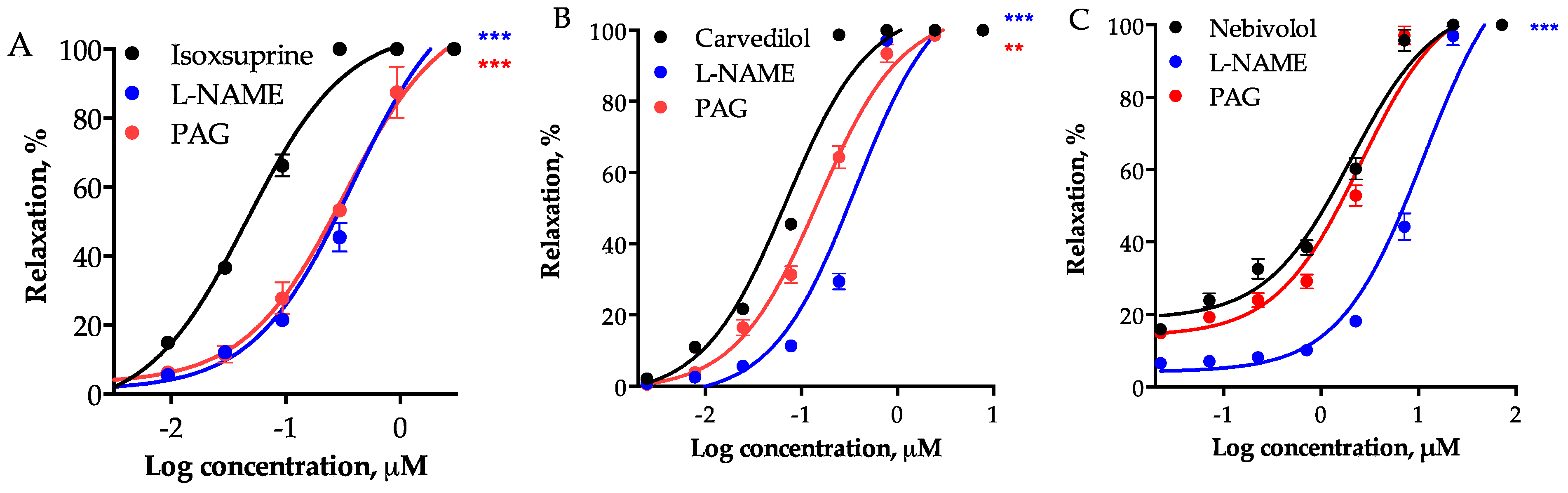
2.2. Determination of the Vasodilator Effect of the Consensus Hits and Involvement of the NO/cGMP and the H2S/KATP Pathways in Their Mechamism of Action
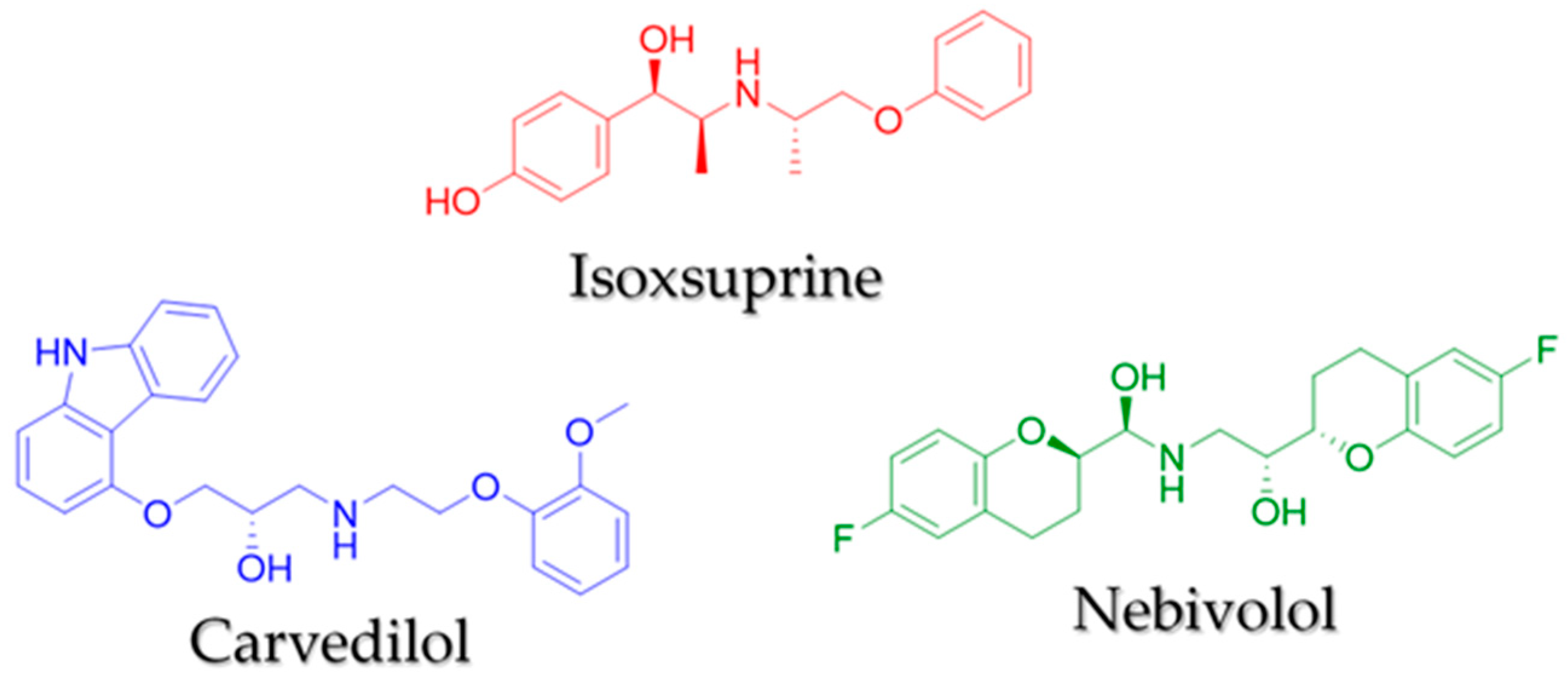
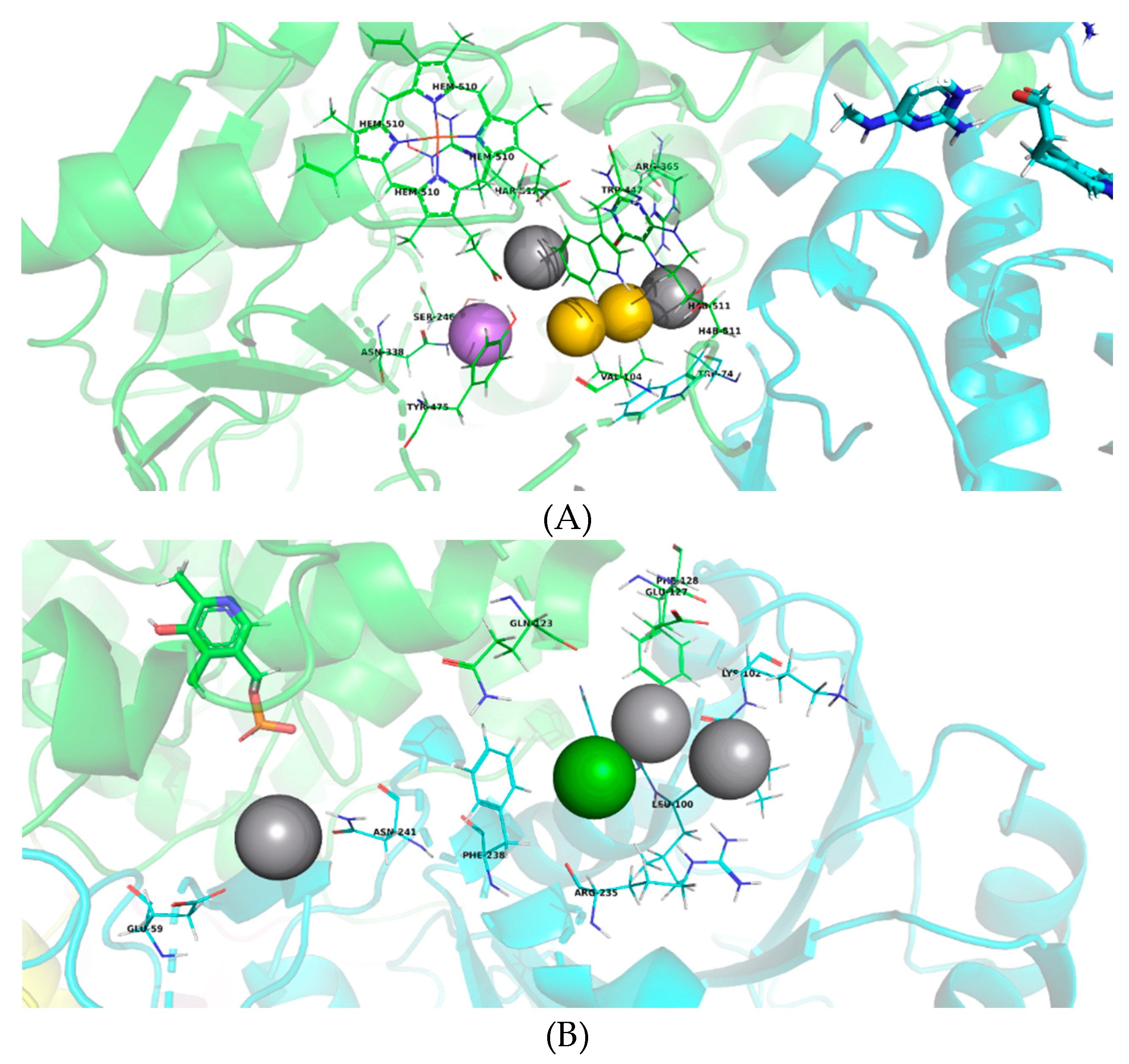
2.3. Virtual Pharmacophoric Elements Identification and General Protein-Ligand Interaction Model
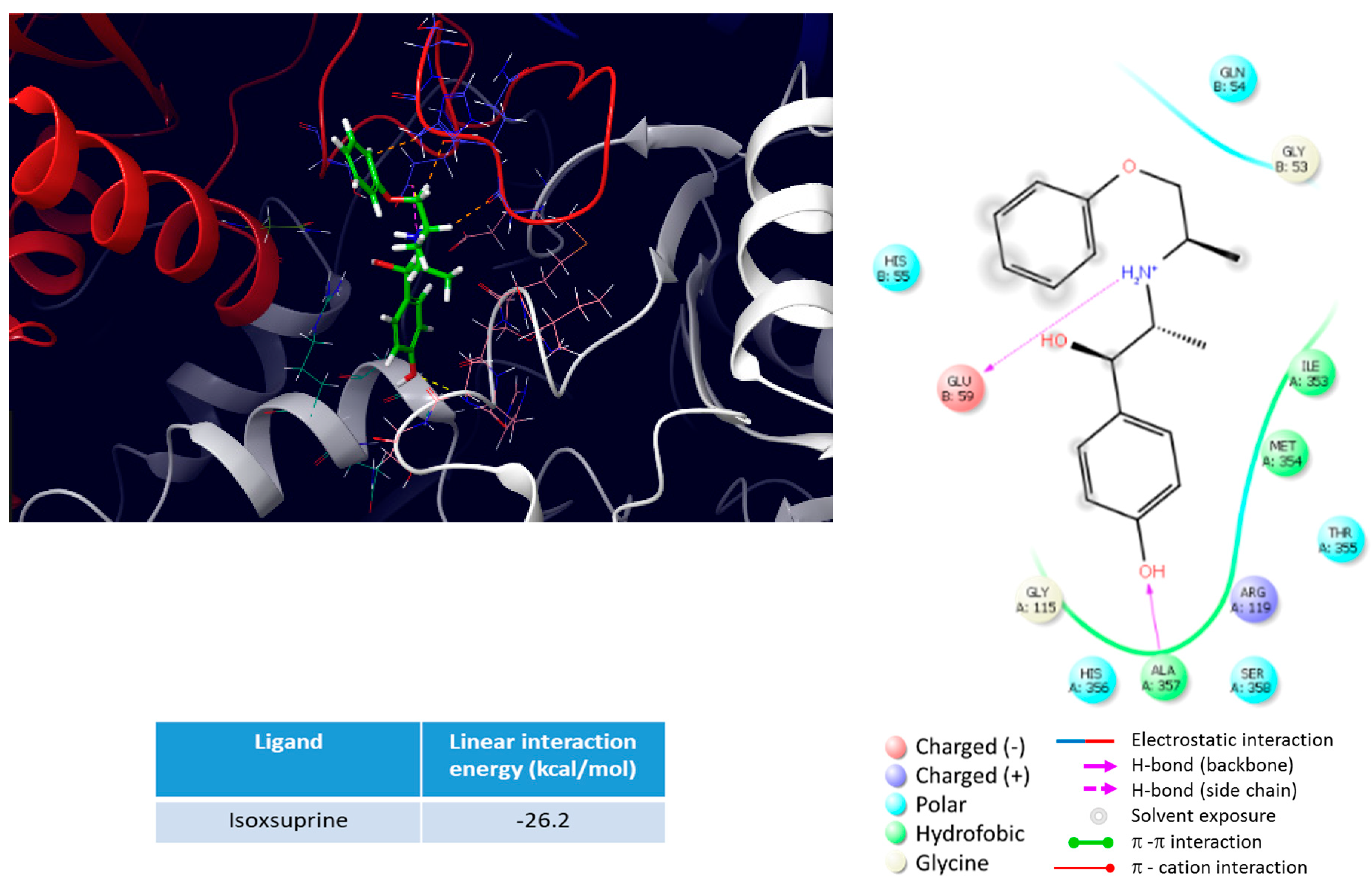
2.4. Molecular Dynamics (MD) of Isoxuprine-CSE Interaction
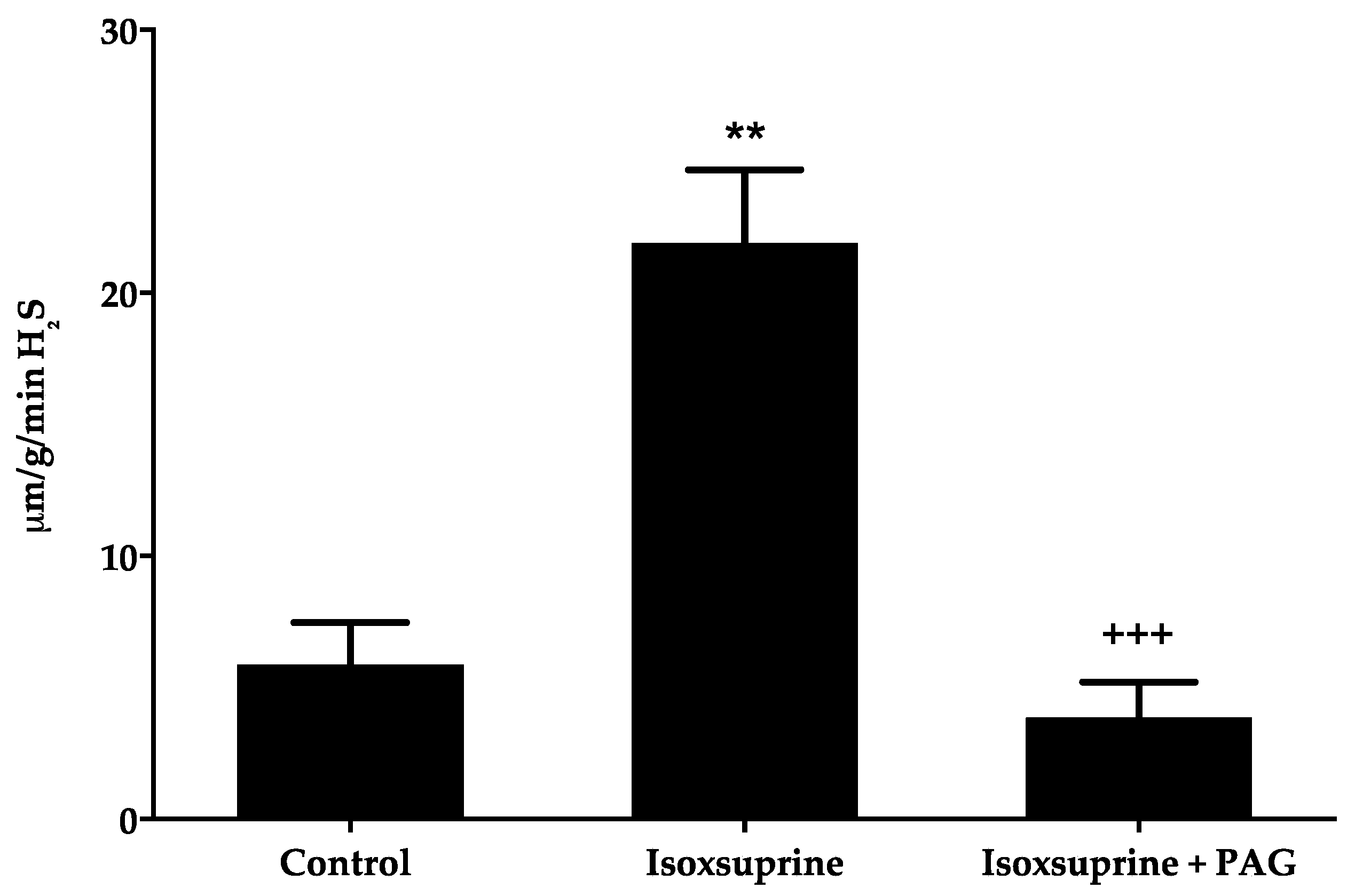
2.5. Increase in H2S Levels Elicited by Isoxsuprine
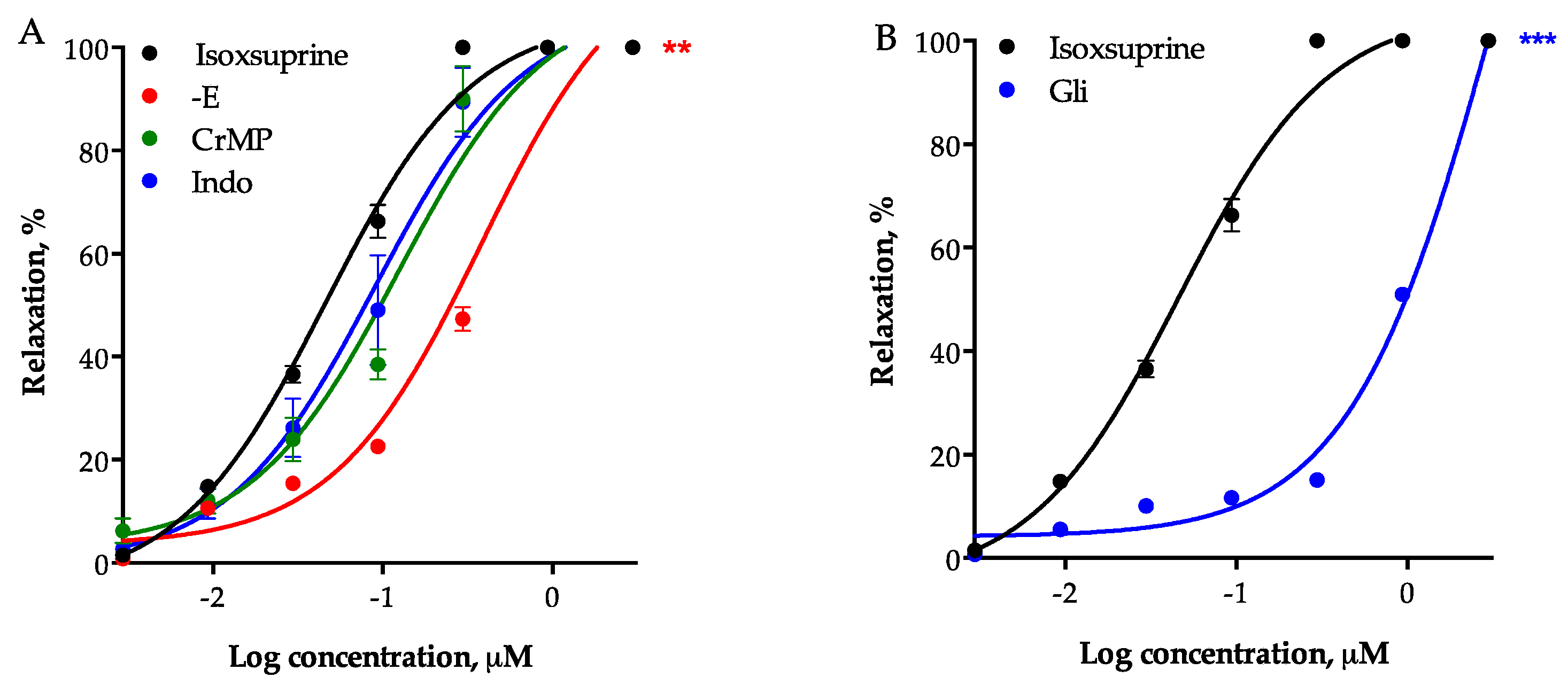
2.6. Participation of Other Endothelial-Derived Relaxing Factors in the Vasodilator Effect of Isoxsuprine
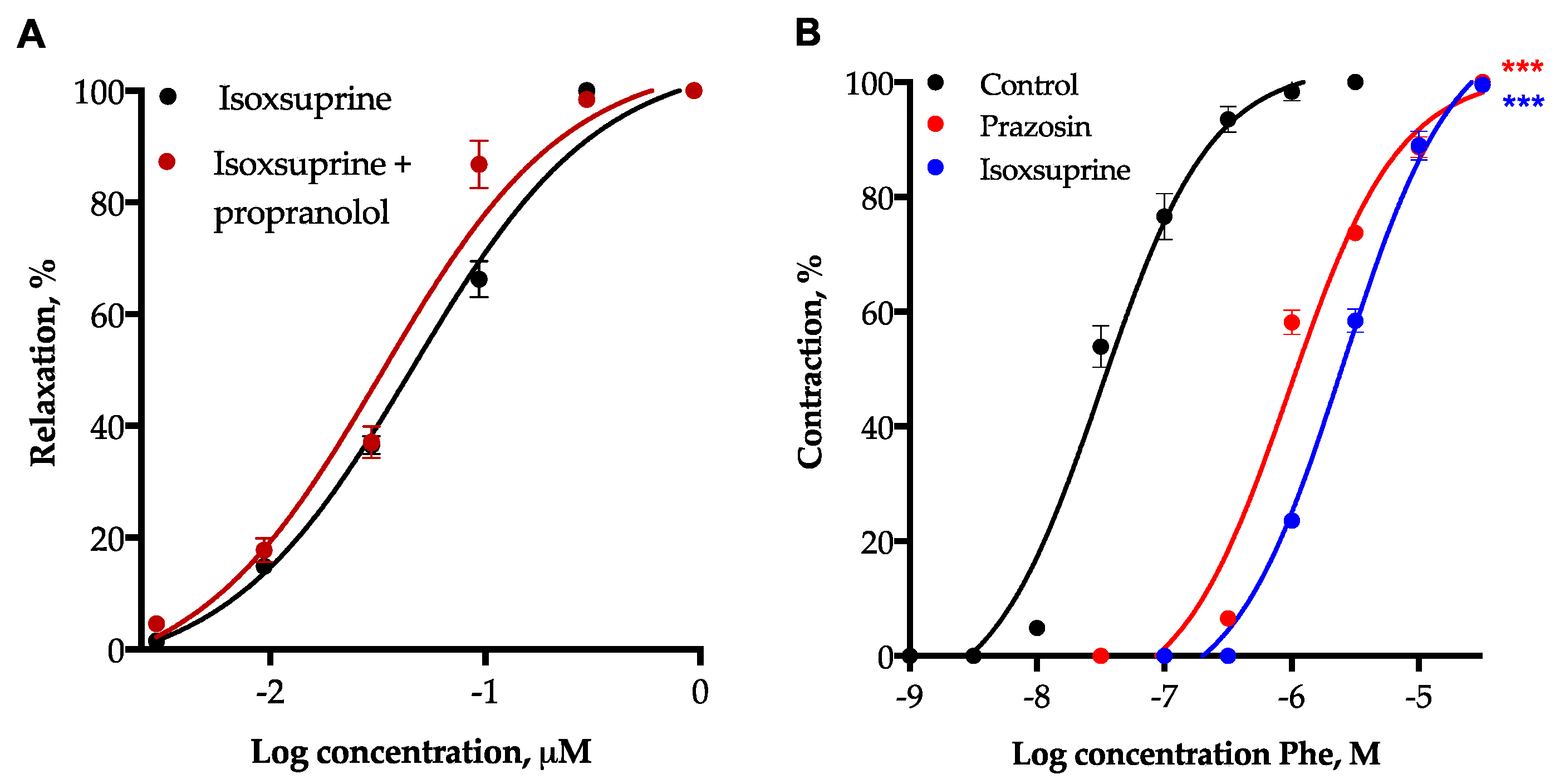
2.7. Participation of β2-Adrenoceptor Activation and α1-Adrenoceptor Blockade in Isoxsuprine-Induced Vasodilation
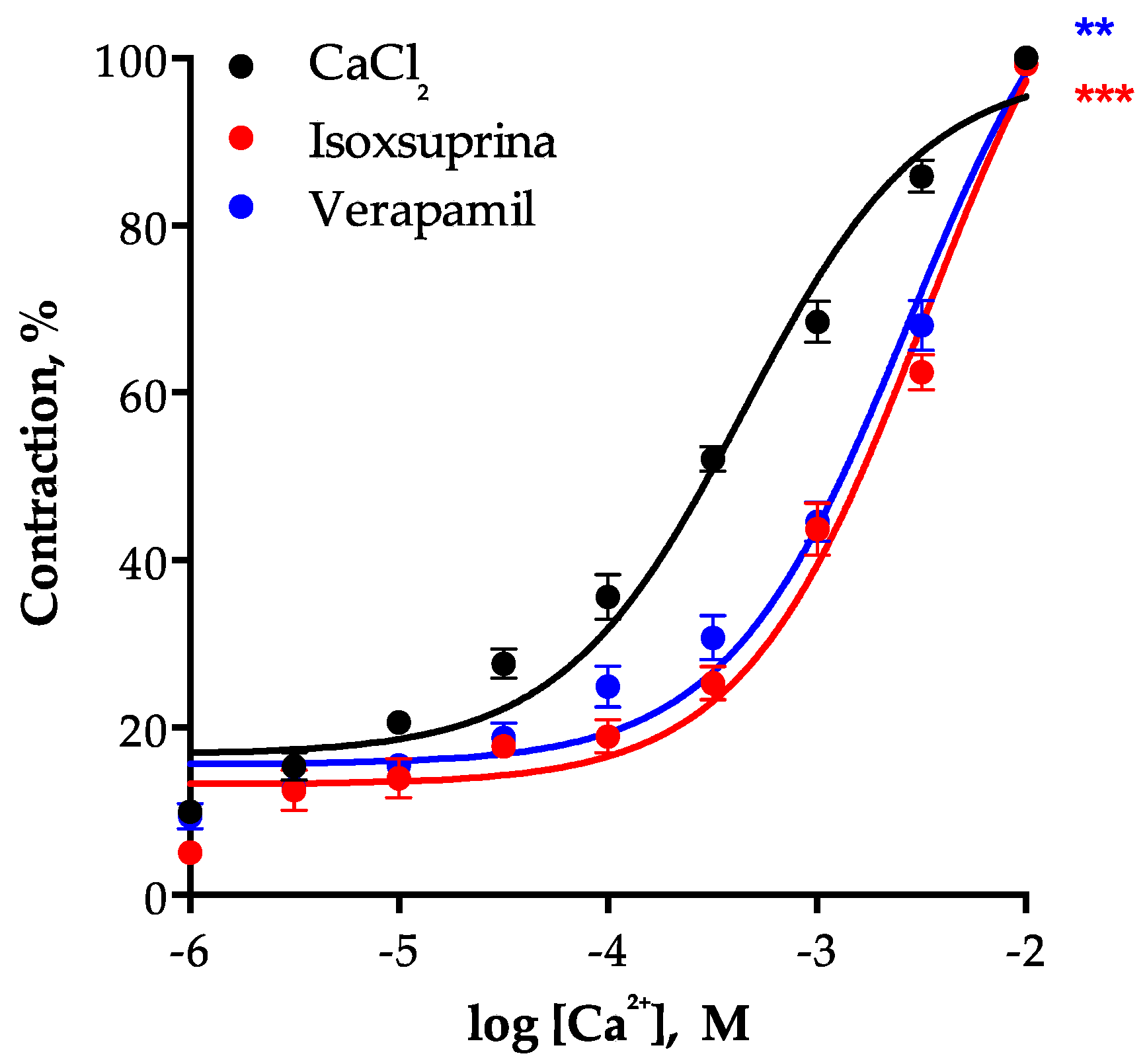
2.8. Involvement of L-Type Voltage-Dependent Calcium Channels in the Vasodilator Effect of Isoxsuprine
3. Discussion
4. Materials and Methods
4.1. Virtual Screening
4.1.1. Human eNOS and CSE Enzyme Structures Preparation
4.1.2. Lead-Like Compounds Selection from Database and Preparation
4.1.3. Virtual Docking
4.2. Pharmacological Evaluation
4.2.1. Reagents
4.2.2. Experimental Animals
4.2.3. Isolated Rat Aorta Assay and Participation of the NO/cGMP and the H2S/KATP Pathways in the Vasodilator Effect Elicited by the “Consensus Hits”
4.2.4. Participation of Endothelium, the CO/cGMP and the PGI2/cAMP Pathways, Potassium Channels and β2 Adrenoceptors in the Vasodilator Effect of Isoxsuprine
4.2.5. Participation of Blockade of α1-Adrenoceptors and Calcium Channels in the Vasodilator Effect of Isoxsuprine
4.2.6. Measurement of H2S Levels in Rat Aorta Homogenates
4.2.7. Statistical Analysis
4.3. Virtual Pharmacophoric Elements Identification and General Protein-Ligand Interaction Model
4.4. Molecular Dynamics (MD) of Isoxuprine-CSE Interaction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mukete, B.N.; Ferdinand, K.C. Polypharmacy in older adults with hypertension: A comprehensive review. J. Clin. Hypertens. 2016, 18, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Dhungana, R.R.; Pandey, A.R.; Bista, B.; Joshi, S.; Devkota, S. Prevalence and associated factors of hypertension: A community-based cross-sectional study in municipalities of Kathmandu, Nepal. Int. J. Hypertens. 2016, 2016, 1656938. [Google Scholar] [CrossRef] [PubMed]
- James, P.A.; Oparil, S.; Carter, B.L.; Cushman, W.C.; Dennison-Himmelfarb, C.; Handler, J.; Lackland, D.T.; LeFevre, M.L.; MacKenzie, T.D.; Ogedegbe, O.; et al. Evidence-based guideline for the management of high blood pressure in adults. JAMA 2014, 311, 507. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.E.; Bellamy, M.C. Antihypertensive drugs. BJA Educ. 2015, 15, 280–285. [Google Scholar] [CrossRef]
- Markovitz, A.A.; Mack, J.A.; Nallamothu, B.K.; Ayanian, J.Z.; Ryan, A.M. Incremental effects of antihypertensive drugs: Instrumental variable analysis. BMJ 2017, 359, j5542. [Google Scholar] [CrossRef] [PubMed]
- Leung, S.W.S.; Vanhoutte, P.M. Endothelium-dependent hyperpolarization: Age, gender and blood pressure, do they matter? Acta Physiol. 2017, 219, 108–123. [Google Scholar] [CrossRef] [PubMed]
- Goto, K.; Ohtsubo, T.; Kitazono, T. Endothelium-dependent hyperpolarization (EDH) in hypertension: The role of endothelial ion channels. Int. J. Mol. Sci. 2018, 19, 315. [Google Scholar] [CrossRef] [PubMed]
- John, C.M.; Mallat, R.K.; George, G.; Kim, T.; Mishra, R.C.; Braun, A.P. Pharmacologic targeting of endothelial Ca2+ -activated K+ channels: A strategy to improve cardiovascular function. Channels 2018, 12, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Montezano, A.C.; Dulak-Lis, M.; Tsiropoulou, S.; Harvey, A.; Briones, A.M.; Touyz, R.M. Oxidative stress and human hypertension: Vascular mechanisms, biomarkers, and novel therapies. Can. J. Cardiol. 2015, 31, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Stead, R.; Musa, M.G.; Bryant, C.L.; Lanham, S.A.; Johnston, D.A.; Reynolds, R.; Torrens, C.; Fraser, P.A.; Clough, G.F. Developmental conditioning of endothelium-derived hyperpolarizing factor-mediated vasorelaxation. J. Hypertens. 2016, 34, 452–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagpure, B.V.; Bian, J.-S. Interaction of hydrogen sulfide with nitric oxide in the cardiovascular system. Oxid. Med. Cell. Longev. 2016, 2016, 6904327. [Google Scholar] [CrossRef] [PubMed]
- Panthi, S.; Chung, H.J.; Jung, J.; Jeong, N.Y. Physiological importance of hydrogen sulfide: Emerging potent neuroprotector and neuromodulator. Oxid. Med. Cell. Longev. 2016, 2016, 9049782. [Google Scholar] [CrossRef] [PubMed]
- Panthi, S.; Manandhar, S.; Gautam, K. Hydrogen sulfide, nitric oxide, and neurodegenerative disorders. Transl. Neurodegener. 2018, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Cebová, M.; Košútová, M.; Pecháňová, O. Cardiovascular effects of gasotransmitter donors. Physiol. Res. 2016, 65, S291–S307. [Google Scholar] [PubMed]
- Gheibi, S.; Jeddi, S.; Kashfi, K.; Ghasemi, A. Regulation of vascular tone homeostasis by NO and H2S: Implications in hypertension. Biochem. Pharmacol. 2018, 149, 42–59. [Google Scholar] [CrossRef] [PubMed]
- Coneski, P.N.; Schoenfisch, M.H. Nitric Oxide Release Part III. Measurement and Reporting. Chem. Soc. Rev. 2012, 41, 3753–3758. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.J.; Ng, M.Y.; Lee, Z.-W.; Dai, W.; Hagen, T.; Moore, P.K.; Huang, D.; Deng, L.-W.; Tan, C.-H. Hydrogen sulfide donors in research and drug development. Med. Chem. Commun. 2014, 5, 557–570. [Google Scholar] [CrossRef]
- Luna-Vázquez, F.J.; Ibarra-Alvarado, C.; Rojas-Molina, A.; Romo-Mancillas, A.; López-Vallejo, F.H.; Solís-Gutiérrez, M.; Rojas-Molina, J.I.; Rivero-Cruz, F. Role of nitric oxide and hydrogen sulfide in the vasodilator effect of ursolic acid and uvaol from black cherry prunus serotina fruits. Molecules 2016, 21, 78. [Google Scholar] [CrossRef] [PubMed]
- Rios, M.Y.; López-Martínez, S.; López-Vallejo, F.; Medina-Franco, J.L.; Villalobos-Molina, R.; Ibarra-Barajas, M.; Navarrete-Vazquez, G.; Hidalgo-Figueroa, S.; Hernández-Abreu, O.; Estrada-Soto, S. Vasorelaxant activity of some structurally related triterpenic acids from Phoradendron reichenbachianum (Viscaceae) mainly by NO production: Ex vivo and in silico studies. Fitoterapia 2012, 83, 1023–1029. [Google Scholar] [CrossRef] [PubMed]
- López-Vallejo, F.; Caulfield, T.; Martínez-Mayorga, K.; Giulianotti, M.; Nefzi, A.; Houghten, R.; Medina-Franco, J. Integrating virtual screening and combinatorial chemistry for accelerated drug discovery. Comb. Chem. High Throughput Screen. 2011, 14, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.G.; Dos Santos, R.N.; Oliva, G.; Andricopulo, A.D. Molecular docking and structure-based drug design strategies. Molecules 2015, 20, 13384–13421. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.; Pawara, R.; Surana, S. In-silico evidences for binding of Glucokinase activators to EGFR C797S to overcome EGFR resistance obstacle with mutant-selective allosteric inhibition. Comput. Biol. Chem. 2018, 74, 167–189. [Google Scholar] [CrossRef] [PubMed]
- Stahura, F.; Bajorath, J. Virtual screening methods that complement HTS. Comb. Chem. High Throughput Screen. 2004, 7, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.M.; Labute, P.; Santavy, M. 2D structure depiction. J. Chem. Inf. Model. 2006, 46, 1107–1123. [Google Scholar] [CrossRef] [PubMed]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A new approach for rapid, accurate docking and scoring. 1. Method and assessment of docking accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef] [PubMed]
- Halgren, T.A.; Murphy, R.B.; Friesner, R.A.; Beard, H.S.; Frye, L.L.; Pollard, W.T.; Banks, J.L. Glide: A new approach fo rapid, accurate docking and scoring. II. Enrichment factors in database screening. J. Med. Chem. 2004, 2, 1750–1759. [Google Scholar] [CrossRef] [PubMed]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra precision glide: Docking and scoring incorporating a model of hydrophobic enclosure for protein-ligand complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [PubMed]
- Fischmann, T.O.; Hruza, A.; Niu, X.D.; Fossetta, J.D.; Lunn, C.A.; Dolphin, E.; Prongay, A.J.; Reichert, P.; Lundell, D.J.; Narula, S.K.; et al. Structural characterization of nitric oxide synthase isoforms reveals striking active-site conservation. Nat. Struct. Biol. 1999, 6, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Collins, R.; Huang, S.; Holmberg-Schiavone, L.; Anand, G.S.; Tan, C.-H.; Van-den-Berg, S.; Deng, L.-W.; Moore, P.K.; Karlberg, T.; et al. Structural basis for the inhibition mechanism of human cystathionine gamma-lyase, an enzyme responsible for the production of H2S. J. Biol. Chem. 2009, 284, 3076–3085. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Lang, P.T.; Brozell, S.R.; Mukherjee, S.; Pettersen, E.F.; Meng, E.C.; Thomas, V.; Rizzo, R.C.; Case, D.A.; James, T.L.; Kuntz, I.D. DOCK 6: Combining techniques to model RNA—Small molecule complexes. RNA 2009, 15, 1219–1230. [Google Scholar] [CrossRef] [PubMed]
- Houston, D.R.; Walkinshaw, M.D. Consensus docking: Improving the reliability of docking in a virtual screening context. J. Chem. Inf. Model. 2013, 53, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, K.; Nakagawa, K.; Umemura, T.; Jitsuiki, D.; Ueda, K.; Goto, C.; Chayama, K.; Yoshizumi, M.; Higashi, Y. Carvedilol improves endothelium-dependent vasodilation in patients with dilated cardiomyopathy. Heart 2007, 93, 247–248. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Polhemus, D.J.; Islam, K.N.; Torregrossa, A.C.; Li, Z.; Potts, A.; Lefer, D.J.; Bryan, N.S. Nebivolol acts as a S-nitrosoglutathione reductase inhibitor. J. Cardiovasc. Pharmacol. Ther. 2016, 21, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Priviero, F.B.M.; Teixeira, C.E.; Toque, H.A.F.; Claudino, M.A.; Webb, R.C.; De Nucci, G.; Zanesco, A.; Antunes, E. Vasorelaxing effects of propranolol in rat aorta and mesenteric artery: A role for nitric oxide and calcium entry blockade. Clin. Exp. Pharmacol. Physiol. 2006, 33, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, X.F.; Poblete, I.; Fernandez, R.; Pedemonte, C.; Cortes, V.; Huidobro-Toro, J.P. NO production and eNOS phosphorylation induced by epinephrine through the activation of -adrenoceptors. AJP Hear. Circ. Physiol. 2009, 297, H134–H143. [Google Scholar] [CrossRef] [PubMed]
- Aggio, A.; Grassi, D.; Onori, E.; D’Alessandro, A.; Masedu, F.; Valenti, M.; Ferri, C. Endothelium/nitric oxide mechanism mediates vasorelaxation and counteracts vasoconstriction induced by low concentration of flavanols. Eur. J. Nutr. 2013, 52, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Alegría-González, C.L. Estudios de Acoplamiento Molecular y Simulaciones de Dinámica Molecular de Triterpenos Selectos en las Enzimas Óxido Nítrico Sintasa Endotelial y Cistationina Gamma-Liasa; Universidad Autónoma de Querétaro: Santiago de Querétaro, Mexico, 2017. [Google Scholar]
- Lu, S.; Huang, W.; Zhang, J. Recent computational advances in the identification of allosteric sites in proteins. Drug Discov. Today 2014, 19, 1595–1600. [Google Scholar] [CrossRef] [PubMed]
- Krimm, I. Identifying protein allosteric transitions for drug discovery with 1D NMR. ChemMedChem 2017, 12, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Marzo, A.; Zava, D.; Coa, K.; Dal Bo, L.; Ismaili, S.; Tavazzi, S.; Cantoni, V. Pharmacokinetics of isoxsuprine hydrochloride administered orally and intramuscularly to female healthy volunteers. Arzneimittelforschung 2009, 59, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Jozwiak, K.; Woo, A.; Tanga, M.; Toll, L.; Jimenez, L.; Kozocas, J.; Plazinska, A.; Xiao, R.; Wainer, I. Comparative molecular field analysis of fenoterol derivatives: A platform towards highly selective and effective beta(2)-adrenergic receptor agonists. Bioorg. Med. Chem. 2011, 18, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Cekic, E.G.; Soydan, G.; Guler, S.; Babaoglu, M.O.; Tuncer, M. Propranolol-induced relaxation in the rat basilar artery. Vascul. Pharmacol. 2013, 58, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, P.M.; Gao, Y. Beta blockers, nitric oxide, and cardiovascular disease. Curr. Opin. Pharmacol. 2013, 13, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Poveromo, L.B.; Pharmd, E.L.M.; Sutherland, S.E. Midodrine for the weaning of vasopressor infusions. J. Clin. Pharm. Ther. 2016, 41, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.T.; Su, H.; Zhang, Q.; Tang, H.Q.; Wang, C.J.; Zhou, Q.; Wei, W.; Zhu, H.Q.; Wang, Y. Sitagliptin inhibits endothelin-1 expression in the aortic endothelium of rats with streptozotocin-induced diabetes by suppressing the nuclear factor-B/IB system through the activation of AMP-activated protein kinase. Int. J. Mol. Med. 2016, 37, 1558–1566. [Google Scholar] [CrossRef] [PubMed]
- Galleano, M.; Bernatova, I.; Puzserova, A.; Balis, P.; Sestakova, N.; Pechanova, O.; Fraga, C.G. (–) -Epicatechin reduces blood pressure and improves vasorelaxation in spontaneously hypertensive rats by NO-mediated mechanism. IUBMB Life 2013, 65, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Eliseeva, E.V.; Kulakova, N.V.; Nevzorova, V.A. Nitric oxide synthase in bronchial epithelium and nitric oxide metabolites in the lungs of rats with bronchial asthma after fenoterol inhalation. Bull. Exp. Biol. Med. 2000, 130, 763–765. [Google Scholar] [CrossRef] [PubMed]
- Odnoshivkina, U.G.; Sytchev, V.I.; Nurullin, L.F.; Giniatullin, A.R.; Zefirov, A.L.; Petrov, A.M. β2-adrenoceptor agonist-evoked reactive oxygen species generation in mouse atria: Implication in delayed inotropic effect. Eur. J. Pharmacol. 2015, 765, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Nade, V.S.; Kawale, L.A.; Pateland, K.M. Protective effect of sitagliptin and rosuvastatin combination on vascular endothelial dysfunction in type-2 diabetes. Indian J. Pharm. Sci. 2015, 77, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Szabo, C. Hydrogen sulfide, an enhancer of vascular nitric oxide signaling: Mechanisms and implications. Am. J. Physiol. Cell Physiol. 2017, 312, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Coletta, C.; Papapetropoulos, A.; Erdelyi, K.; Olah, G.; Módis, K.; Panopoulos, P. Hydrogen sulfide and nitric oxide are mutually dependent in the regulation of angiogenesis and endothelium-dependent vasorelaxation. Proc. Natl. Acad. Sci. USA 2012, 109, 9161–9166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Dong, X. Nebivolol ameliorates asymmetric dimethylarginine-induced vascular response in rat aorta via β3 adrenoceptor-mediated mechanism. Clin. Exp. Hypertens. 2016, 38, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Broeders, M.A.; Doevendans, P.A.; Bekkers, B.; Bronsaer, R.; van Gorsel, E.; Heemskerk, J.; Egbrink, M.G.; van Breda, E.; Reneman, R.; van Der Zee, R. Nebivolol: A third-generation beta-blocker that augments vascular nitric oxide release: Endothelial beta(2)-adrenergic receptor-mediated nitric oxide production. Circulation 2000, 102, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Dessy, C.; Saliez, J.; Ghisdal, P.; Daneau, G.; Lobysheva, I.I.; Frérart, F.; Belge, C.; Jnaoui, K.; Noirhomme, P.; Feron, O.; et al. Endothelial β3-adrenoreceptors mediate nitric oxide-dependent vasorelaxation of coronary microvessels in response to the third-generation β-blocker nebivolol. Circulation 2005, 112, 1198–1205. [Google Scholar] [CrossRef] [PubMed]
- Quang, T.T.; Rozec, B.; Audigane, L.; Gauthier, C. Investigation of the different adrenoceptor targets of nebivolol enantiomers in rat thoracic aorta. Br. J. Pharmacol. 2009, 156, 601–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Huang, C.; Zhang, B.; Huang, Q.; Chen, J.; Xu, L. The effects of carvedilol on cardiac structural remodeling: The role of endogenous nitric oxide in the activity of carvedilol. Mol. Med. Rep. 2013, 7, 1155–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlovski, V.I.; Lomnicka, M.; Bartus, M.; Sternak, M.; Chlopicki, S. Anti-thrombotic effects of nebivolol and carvedilol: Involvement of β2 receptors and COX-2/PGI2 pathways. Pharmacol. Rep. 2015, 67, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.; Zhang, W.W.; Yao, L.; Yang, S.; Nie, W.; Huang, F. Carvedilol promotes mitochondrial biogenesis by regulating the PGC-1/TFAM pathway in human umbilical vein endothelial cells (HUVECs). Biochem. Biophys. Res. Commun. 2016, 470, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, P.J.; Gonçalves, L.; Monteiro, P.; Providencia, L.A.; Moreno, A.J. Are the antioxidant properties of carvedilol important for the protection of cardiac mitochondria? Curr. Vasc. Pharmacol. 2005, 3, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, R.; Jin, M.; Huang, Y.; Liu, A.; Qin, J.; Chen, M.; Wen, S.; Pi, R.; Shen, W. Carvedilol attenuates 6-hydroxydopamine-induced cell death in PC12 cells: Involvement of Akt and Nrf2/ARE pathways. Neurochem. Res. 2014, 39, 1733–1740. [Google Scholar] [CrossRef] [PubMed]
- Garland, C.J.; Hiley, C.R.; Dora, K.A. EDHF: Spreading the influence of the endothelium. Br. J. Pharmacol. 2011, 164, 839–852. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhang, J.; Lu, Y.; Wang, R. The vasorelaxant effect of H2S as a novel endogenous gaseous KATP channel opener. EMBO J. 2001, 20, 6008–6016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peers, C.; Bauer, C.C.; Boyle, J.P.; Scragg, J.L.; Dallas, M.L. Modulation of ion channels by hydrogen sulfide. Antioxid. Redox Signal. 2012, 17, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Kim-Egloff, C.; Hässig, M.; Bruckmaier, R.; Bleul, U. Doppler sonographic examination of uterine and placental perfusion in cows in the last month of gestation and effects of epidural anesthesia and isoxsuprine. Theriogenology 2016, 85, 986–998. [Google Scholar] [CrossRef] [PubMed]
- Belloli, C.; Carcano, R.; Arioli, F.; Beretta, C. Affinity of isoxsuprine for adrenoreceptors in equine digital artery and implications for vasodilatory action. Equine Vet. J. 2000, 32, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.W.; Thompson, J.F.; Carter, M.B.; Edwards, B.S.; Sklar, L.A.; Rosenberg, G.A. Identification of isoxsuprine hydrochloride as a neuroprotectant in ischemic stroke through cell-based high-throughput screening. PLoS ONE 2014, 9, e96761. [Google Scholar] [CrossRef] [PubMed]
- Amer, M.; Dungan, K.; McKinney, G. The effects of isoxsuprine hydrochloride on the cyclic nucleotide system in the rat uterus. J. Pharmacol. Exp. Ther. 1974, 190, 243–248. [Google Scholar] [PubMed]
- Cohen, M.; Wiley, K. Beta1 and beta2 receptor mechanisms in rat jugular veins: Differences between norepinephrine and isoproterenol-induced relaxation. Life Sci. 1978, 23, 1997–2006. [Google Scholar] [CrossRef]
- Calixto, J.; Simas, C. Mechanism of action of isoprenaline, isoxuprine, terbutaline and orciprenaline on gravid human isolated myometrium. Influence of the neuronal uptake process. Biol. Reprod. 1984, 30, 1117–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abel, P.; Fox, A.; Minneman, K. Effects of isoxuprine and nylidrin on adrenoreceptors in rat vas deferens. J. Aut. Pharmacol. 1985, 5, 213–219. [Google Scholar] [CrossRef]
- Elliott, J.; Soydan, J. Characterization of b-adrenoceptors in equine digital veins: Implications of the modes of vasodilatory action of isoxsuprine. Equine Vet. J. 1995, 27 (Suppl. 19), 101–107. [Google Scholar] [CrossRef]
- Brousseau, D.; Desjardins, J.; Jasmin, G.; Dumont, L. Altered coronary and cardiac adrenergic response in the failing hamster heart: Role of cyclooxygenase derivatives. Can. J. Physiol. Pharmacol. 2002, 80, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Limberg, J.; Johansson, R.; Peltonen, G.; Harrell, J.; Kellawan, J.; Eldridge, M.; Sebranek, J.; Schrage, W. β-Adrenergic-mediated vasodilation in young men and women: Cyclooxygenase restrains nitric oxide synthase. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H756–H764. [Google Scholar] [CrossRef] [PubMed]
- Toda, N.; Okamura, T. Beta adrenoceptor subtype in isolated human, monkey and dog epicardial coronary arteries. J. Pharmacol. Exp. Ther. 1990, 253, 518–524. [Google Scholar] [PubMed]
- Raymajhi, R.; Pratap, K. A comparative study between nifedipine and isoxsuprine in the suppression of preterm labour. Kathmandu Univ. Med. J. 2003, 1, 85–90. [Google Scholar]
- Zahir, F.; Nath, K.K.; Choudhury, H.H. A clinical study of feto-maternal outcome of tocolytic agent nifedipine as compared to isoxsuprine hydrochloride in preterm labor. MedPulse-Int. Med. J. 2015, 2, 483–487. [Google Scholar]
- Elliott, W.J.; Ram, C.V.S. Calcium channel blockers. J. Clin. Hypertens. 2011, 13, 687–689. [Google Scholar] [CrossRef] [PubMed]
- Godfraind, T. Calcium channel blockers in cardiovascular pharmacotherapy. J. Cardiovasc. Pharmacol. Ther. 2014, 19, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Salomone, S.; Silva, C.L.M.; Morel, N.; Godfraind, T. Facilitation of the vasorelaxant action of calcium antagonists by basal nitric oxide in depolarized artery. Naunyn. Schmiedebergs. Arch. Pharmacol. 1996, 354, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Gamal El-Din, T.; Swanson, T.; Pryde, D.; Scheuer, T.; Zheng, N.; Catterall, W. Structural basis for inhibition of a voltage-gated Ca2+ channel by Ca2+ antagonist drugs. Nature 2016, 537, 117–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sud, M. MayaChemTools: An Open Source Package for Computational Drug Discovery. J. Chem. Inf. Model. 2016, 56, 2292–2297. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Jalily Hasani, H.; Ganesan, A.; Houghton, M.; Barakat, K. Modeling the human Nav1.5 sodium channel: Structural and mechanistic insights of ion permeation and drug blockade. Drug Des. Devel. Ther. 2017, 11, 2301–2324. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berman, H.M.; Henrick, K.; Nakamura, H. Announcing the worldwide Protein Data Bank. Nat. Struct. Biol. 2003, 10, 980. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.W.; Prlic, A.; Bi, C.; Bluhm, W.F.; Christie, C.H.; Dutta, S.; Green, R.K.; Goodsell, D.S.; Westbrook, J.D.; Woo, J.; et al. The RCSB Protein Data Bank: Views of structural biology for basic and applied research and education. Nucleic Acids Res. 2015, 43, D345–D356. [Google Scholar] [CrossRef] [PubMed]
- Sastry, G.M.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput. Aid Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Sterling, T.; Irwin, J.J. ZINC 15—Ligand discovery for everyone. J. Chem. Inf. Model. 2015, 55, 2324–2337. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.J.; Sterling, T.; Mysinger, M.M.; Bolstad, E.S.; Coleman, R.G. ZINC: A free tool to discover chemistry for biology. J. Chem. Inf. Model. 2012, 52, 1757–1768. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.J.; Shoichet, B.K. ZINC—A free database of commercially available compounds for virtual screening. J. Chem. Inf. Model. 2005, 45, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Ibarra-Alvarado, C.; García, J.A.; Aguilar, M.B.; Rojas, A.; Falcón, A.; Heimer de la Cotera, E.P. Biochemical and pharmacological characterization of toxins obtained from the fire coral Millepora complanata. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Castro-Ruiz, J.E.; Rojas-Molina, A.; Luna-Vázquez, F.J.; Rivero-Cruz, F.; García-Gasca, T.; Ibarra-Alvarado, C. Affinin (Spilanthol), isolated from heliopsis longipes, induces vasodilation via activation of gasotransmitters and prostacyclin signaling pathways. Int. J. Mol. Sci. 2017, 18, 218. [Google Scholar] [CrossRef] [PubMed]
- Huo, L.; Zhang, J.; Qu, Z.; Chen, H.; Li, Y.; Gao, W. Vasorelaxant effects of Shunaoxin pill are mediated by NO/cGMP pathway, HO/CO pathway and calcium channel blockade in isolated rat thoracic aorta. J. Ethnopharmacol. 2015, 173, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Stipanuk, M.H.; Beck, P.W. Characterization of the enzymic capacity for cysteine desulphhydration in liver and kidney of the rat. Biochem. J. 1982, 206, 267–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koes, D.R.; Camacho, C.J. Pharmer: Efficient and exact pharmacophore search. J. Chem. Inf. Model. 2011, 51, 1307–1314. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.P.; Richards, G.; De La Iglesia, B.; Rayward-Smith, V.J. Clustering rules: A comparison of partitioning and hierarchical clustering algorithms. J. Math. Model. Algorithms 2006, 5, 475–504. [Google Scholar] [CrossRef]
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K. Cluster: Cluster Analysis Basics and Extensions. R Packag. Version 2.0.7-1. 2018. Available online: https://cran.r-project.org/package=cluster (accessed on 10 March 2019).
- R-Core-Team R: A Language and Environment for Statistical Computing. 2013. Available online: https://www.r-project.org/ (accessed on 10 March 2019).
- Hess, B.; Kutzner, C.; Van Der Spoel, D.; Lindahl, E. GROMACS 4: Algorithms for highly efficient, load-balanced, and scalable molecular simulation. J. Chem. Theory Comput. 2008, 4, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, W.; Kollman, P.A.; Case, D.A. Automatic atom type and bond type perception in molecular mechanical calculations. J. Mol. Graph. Model. 2006, 25, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Sousa Da Silva, A.W.; Vranken, W.F. ACPYPE—AnteChamber PYthon Parser interfacE. BMC Res. Notes 2012, 5, 367. [Google Scholar] [CrossRef] [PubMed]
- Bussi, G.; Donadio, D.; Parrinello, M. Canonical sampling through velocity rescaling. J. Chem. Phys. 2007, 126, 014101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrinello, M.; Rahman, A. Polymorphic transitions in single crystals: A new molecular dynamics method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Aqvist, J.; Medina, C.; Samuelsson, J.E. A new method for predicting binding affinity in computer-aided drug design. Protein Eng. 1994, 7, 385–391. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of isoxsuprine, carvedilol, and nebivolol are available from the authors. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | EC50 (µM) ± SEM | Emax (%) ± SEM |
|---|---|---|
| Controls | ||
| SNP | 0.0099 ± 0.001 | 100.7 ± 0.747 |
| ACh | 50.30 ± 5.126 | 68.46 ± 1.122 |
| NaHS | 191.0 ± 9.841 | 92.17 ± 2.721 |
| Highly Potent Compounds | ||
| Isoxsuprine | 0.046 ± 0.004 | 106.1 ± 0.343 |
| Carvedilol | 0.069 ± 0.003 | 106.6 ± 0.115 |
| Nebivolol | 2.014 ± 0.215 | 106.7 ± 0.543 |
| Lowly Potent Compounds | ||
| Propranolol | 18.120 ± 1.419 | 103.6 ± 1.663 |
| Pindolol | 39.490 ± 2.603 | 106 ± 0.463 |
| Sitagliptin | 252.30 ± 8.058 | 117.6 ± 0.497 |
| Fenoterol | 608.60 ± 43.225 | 120.3 ± 1.548 |
| Epicatechin | 626.40 ± 67.372 | 101.5 ± 2.400 |
| Midodrine | 4698 ± 324.691 | 219.5 ± 8.219 |
| Compound | Control | L-NAME | PAG |
|---|---|---|---|
| EC50 (µM) ± SEM | EC50 (µM) ± SEM | EC50 (µM) ± SEM | |
| Most Potent Vasodilators | |||
| Isoxsuprine | 0.0461 ± 0.004 | 0.3846 ± 0.040 *** | 0.3255 ± 0.026 *** |
| Carvedilol | 0.0695 ± 0.003 | 0.3643 ± 0.011 *** | 0.1500 ± 0.019 ** |
| Nebivolol | 2.0135 ± 0.215 | 11.290 ± 1.501 *** | 2.5280 ± 0.229 |
| Less Potent Vasodilators | |||
| Propranolol | 18.120 ± 1.419 | 40.430 ± 5.696 ** | 27.900 ± 3.651 |
| Pindolol | 39.490 ± 2.603 | 166.1 ± 3.610 *** | 89.47 ± 7.940 *** |
| Sitagliptin | 252.30 ± 8.058 | 864.3 ± 8.692 *** | 436.2 ± 12.63 *** |
| Fenoterol | 608.6 ± 43.225 | 882.1 ± 27.513 *** | 1038 ± 22.241 *** |
| Epicatechin | 626.4 ± 67.372 | 1789 ± 389.469 * | 3087 ± 552.718 ** |
| Midodrine | 4698 ± 324.691 | 8618 ± 2060.477 * | 10076 ± 1955.342 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medina-Ruiz, D.; Erreguin-Luna, B.; Luna-Vázquez, F.J.; Romo-Mancillas, A.; Rojas-Molina, A.; Ibarra-Alvarado, C. Vasodilation Elicited by Isoxsuprine, Identified by High-Throughput Virtual Screening of Compound Libraries, Involves Activation of the NO/cGMP and H2S/KATP Pathways and Blockade of α1-Adrenoceptors and Calcium Channels. Molecules 2019, 24, 987. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24050987
Medina-Ruiz D, Erreguin-Luna B, Luna-Vázquez FJ, Romo-Mancillas A, Rojas-Molina A, Ibarra-Alvarado C. Vasodilation Elicited by Isoxsuprine, Identified by High-Throughput Virtual Screening of Compound Libraries, Involves Activation of the NO/cGMP and H2S/KATP Pathways and Blockade of α1-Adrenoceptors and Calcium Channels. Molecules. 2019; 24(5):987. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24050987
Chicago/Turabian StyleMedina-Ruiz, Daniella, Berenice Erreguin-Luna, Francisco J. Luna-Vázquez, Antonio Romo-Mancillas, Alejandra Rojas-Molina, and César Ibarra-Alvarado. 2019. "Vasodilation Elicited by Isoxsuprine, Identified by High-Throughput Virtual Screening of Compound Libraries, Involves Activation of the NO/cGMP and H2S/KATP Pathways and Blockade of α1-Adrenoceptors and Calcium Channels" Molecules 24, no. 5: 987. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24050987