Anti-Inflammatory Effects of High Hydrostatic Pressure Extract of Mulberry (Morus alba) Fruit on LPS-Stimulated RAW264.7 Cells


Abstract
:1. Introduction
2. Results and Discussion
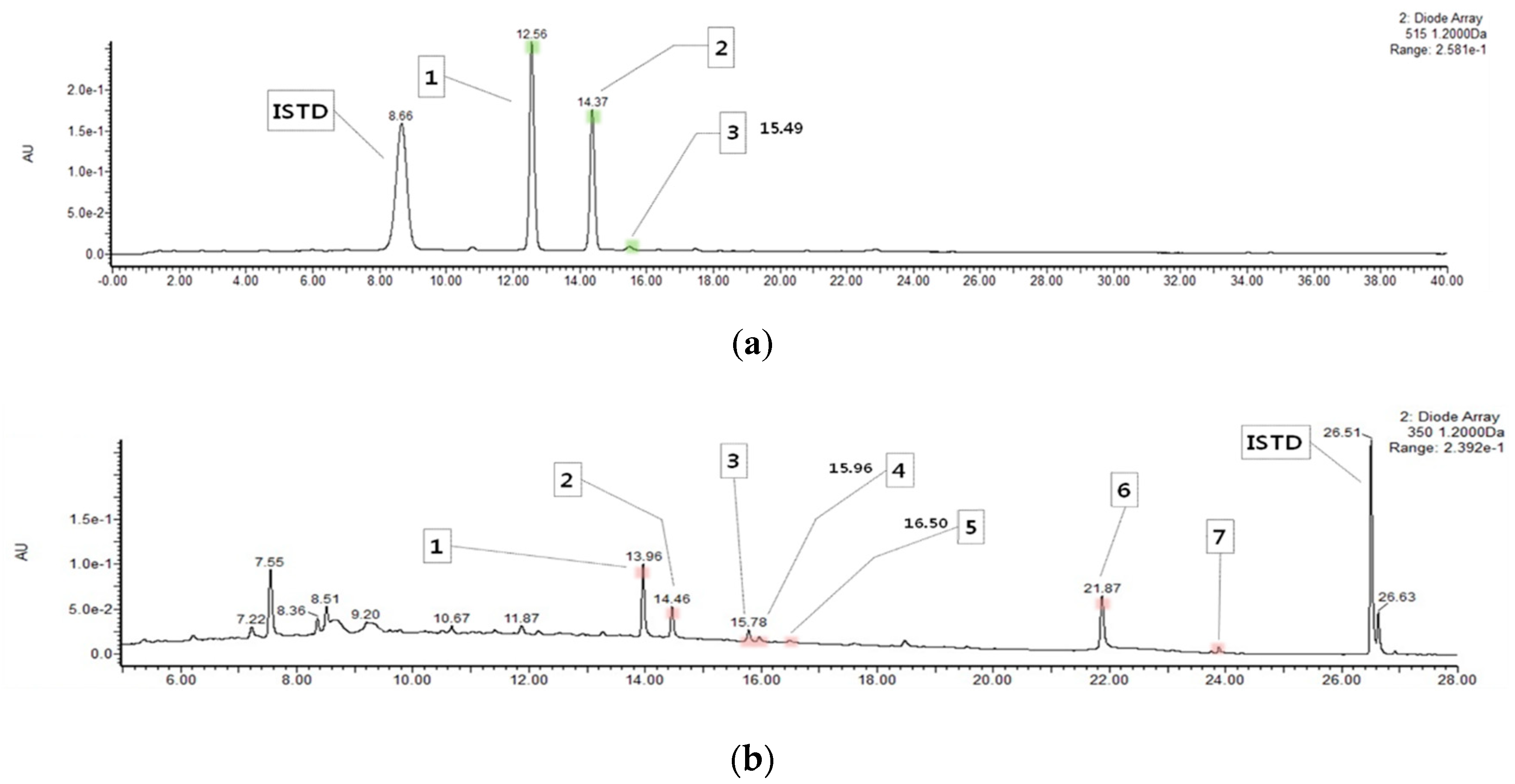
2.1. Content of Anthocyanins and Flavonols in HM
2.2. Effects of HM on Cell Viability
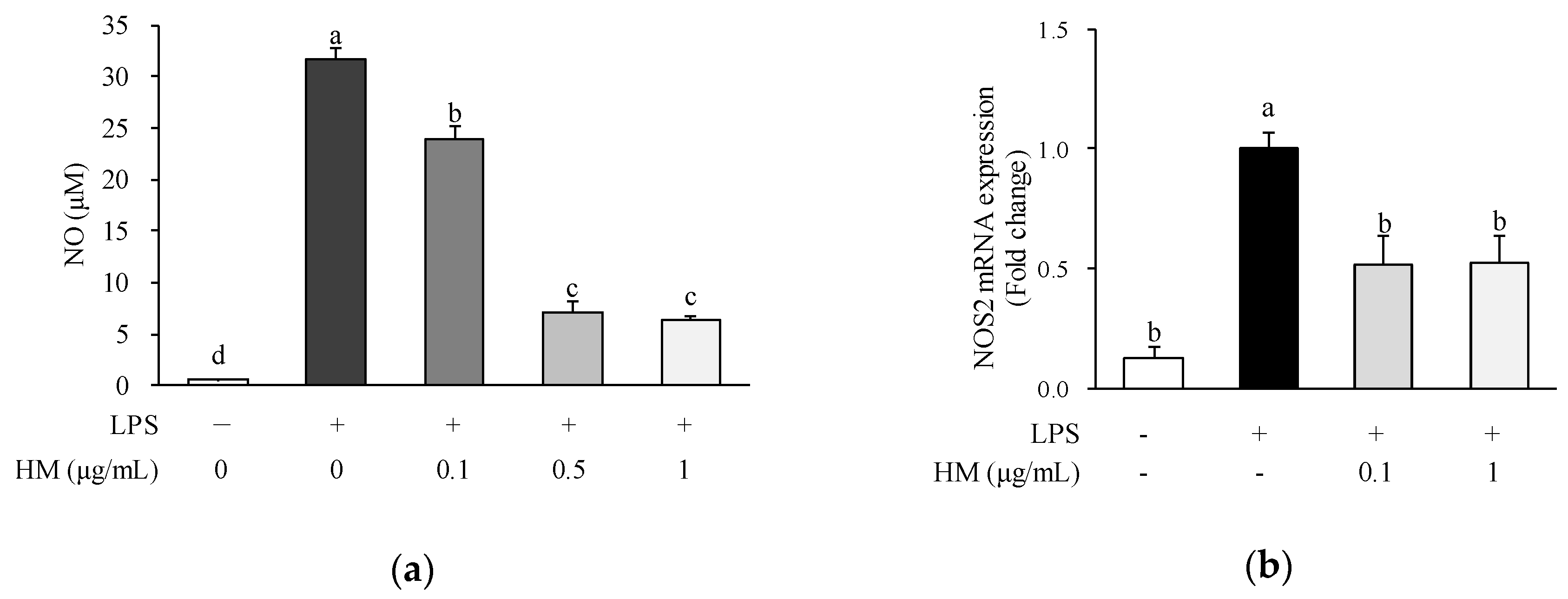
2.3. Effects of HM on NO Production and NOS2 mRNA Expression
2.4. Effects of HM on PTGS2 mRNA and Protein Expression
2.5. Effects of HM on Cytokine Production
3. Materials and Methods
3.1. Cells and Reagents
3.2. Preparation of HM
3.3. UPLC-PDA-Q/TOF-MS Analysis
3.4. Cell Culture
3.5. Cell Viability Assay
3.6. NO Assay
3.7. Enzyme-Linked Immunosorbent Assay (ELISA) for TNF-α and IL-6
3.8. Quantitative Reverse Transcriptase Polymerase Chain Reaction (qRT-PCR)
3.9. Western Blot Analysis
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Manrique-Moreno, M.; Heinbockel, L.; Suwalsky, M.; Garidel, P.; Brandenburg, K. Biophysical study of the non-steroidal anti-inflammatory drugs (NSAID) ibuprofen, naproxen and diclofenac with phosphatidylserine bilayer membranes. Biochim. Biophys. Acta-Gen. Subj. 2016, 1858, 2123–2131. [Google Scholar] [CrossRef]
- Craig, D.M.; Ashcroft, S.P.; Belew, M.Y.; Stocks, B.; Currell, K.; Baar, K.; Philp, A. Utilizing small nutrient compounds as enhancers of exercise-induced mitochondrial biogenesis. Front. Physiol. 2015, 6, 296. [Google Scholar] [CrossRef]
- Yuan, Q.; Zhao, L. The mulberry (Morus alba L.) fruit-a review of characteristic components and health benefits. J. Agric. Food Chem. 2017, 65, 10383–10394. [Google Scholar] [CrossRef]
- Ju, W.-T.; Kwon, O.-C.; Lee, M.-K.; Kim, H.-B.; Sung, G.-B.; Kim, Y.-S. Quali-quantitative analysis of flavonoids for mulberry leaf and fruit of ‘Suhyang’. Korean J. Environ. Agric. 2017, 36, 249–255. [Google Scholar] [CrossRef]
- Li, Y.; Bao, T.; Chen, W. Comparison of the protective effect of black and white mulberry against ethyl carbamate-induced cytotoxicity and oxidative damage. Food Chem. 2018, 243, 65–73. [Google Scholar] [CrossRef]
- Choi, K.H.; Lee, H.A.; Park, M.H.; Han, J.S. Mulberry (Morus alba L.) fruit extract containing anthocyanins improves glycemic control and insulin sensitivity via activation of AMP-activated protein kinase in diabetic C57BL/Ksj-db/db mice. J. Med. Food 2016, 19, 737–745. [Google Scholar] [CrossRef]
- Chang, B.Y.; Kim, S.B.; Lee, M.K.; Park, H.; Kim, S.Y. Improved chemotherapeutic activity by Morus alba fruits through immune response of toll-like receptor 4. Int. J. Mol. Sci. 2015, 16, 24139–24158. [Google Scholar] [CrossRef] [PubMed]
- Eo, H.; Lim, Y. Combined Mulberry Leaf and Fruit Extract Improved Early Stage of Cutaneous Wound Healing in High-Fat Diet-Induced Obese Mice. J. Med. Food 2016, 19, 161–169. [Google Scholar] [CrossRef]
- Liu, C.J.; Lin, J.Y. Anti-inflammatory effects of phenolic extracts from strawberry and mulberry fruits on cytokine secretion profiles using mouse primary splenocytes and peritoneal macrophages. Int. Immunopharmacol. 2013, 16, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Masi, D.; Maria, A. Anticancer activities of anthocyanin extract from genotyped Solanum tuberosum L. â Vitelotteâ. J. Funct. Foods 2015. [Google Scholar] [CrossRef]
- Huang, H.-P.; Shih, Y.-W.; Chang, Y.-C.; Hung, C.-N.; Wang, C.-J. Chemoinhibitory Effect of Mulberry Anthocyanins on Melanoma Metastasis Involved in the Ras/PI3K Pathway. J. Agric. Food Chem. 2008, 56, 9286–9293. [Google Scholar] [CrossRef] [PubMed]
- Bilancio, A.; Rinaldi, B.; Oliviero, M.A.; Donniacuo, M.; Monti, M.G.; Boscaino, A.; Marino, I.; Friedman, L.; Rossi, F.; Vanhaesebroeck, B.; et al. Inhibition of p110delta PI3K prevents inflammatory response and restenosis after artery injury. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K. Food processing by high hydrostatic pressure. Biosci. Biotechnol. Biochem. 2017, 81, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Du, B.L.; Cui, Z.W.; Xu, L.P.; Li, C.Y. Effects of high hydrostatic pressu Chemoinhibitory Effect re and thermal processing on bioactive compounds, antioxidant activity, and volatile profile of mulberry juice. Food Sci. Techno. Int. 2017, 23, 119–127. [Google Scholar] [CrossRef]
- Qian, Z.; Wu, Z.; Huang, L.; Qiu, H.; Wang, L.; Li, L.; Yao, L.; Kang, K.; Qu, J.; Wu, Y.; et al. Mulberry fruit prevents LPS-induced NF-kappaB/pERK/MAPK signals in macrophages and suppresses acute colitis and colorectal tumorigenesis in mice. Sci. Rep. 2015, 5, 17348. [Google Scholar] [CrossRef]
- Liu, C.J.; Lin, J.Y. Anti-inflammatory and anti-apoptotic effects of strawberry and mulberry fruit polysaccharides on lipopolysaccharide-stimulated macrophages through modulating pro-/anti-inflammatory cytokines secretion and Bcl-2/Bak protein ratio. Food Chem. Toxicol. 2012, 50, 3032–3039. [Google Scholar] [CrossRef]
- Zhang, H.; Tchabo, W.; Ma, Y. Quality of extracts from blueberry pomace by high hydrostatic pressure, ultrasonic, microwave and heating extraction: A comparison study. Emir. J. Food Agric. 2017, 29, 815–819. [Google Scholar] [CrossRef]
- MacMicking, J.; Xie, Q.W.; Nathan, C. Nitric oxide and macrophage function. Annu. Rev. Immunol. 1997, 15, 323–350. [Google Scholar] [CrossRef]
- Tang, C.C.; Huang, H.P.; Lee, Y.J.; Tang, Y.H.; Wang, C.J. Hepatoprotective effect of mulberry water extracts on ethanol-induced liver injury via anti-inflammation and inhibition of lipogenesis in C57BL/6J mice. Food Chem. Toxicol. 2013, 62, 786–796. [Google Scholar] [CrossRef]
- Ou, T.T.; Kuo, C.Y.; Chyau, C.C.; Lee, H.J.; Peng, J.S.; Wang, C.J. Improvement of lipopolysaccharide-induced hepatic injuries and inflammation with mulberry extracts. J. Sci. Food Agric. 2013, 93, 1880–1886. [Google Scholar] [CrossRef]
- Nakanishi, M.; Rosenberg, D.W. Multifaceted roles of PGE2 in inflammation and cancer. Semin. Immunopathol. 2013, 35, 123–137. [Google Scholar] [CrossRef]
- Horng, C.T.; Liu, Z.H.; Huang, Y.T.; Lee, H.J.; Wang, C.J. Extract from mulberry (Morus australis) leaf decelerate acetaminophen induced hepatic inflammation involving downregulation of myeloid differentiation factor 88 (MyD88) signals. J. Food Drug Anal. 2017, 25, 862–871. [Google Scholar] [CrossRef]
- Vassalli, P. The pathophysiology of tumor necrosis factors. Annu. Rev. Immunol. 1992, 10, 411–452. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef]
- de Padua Lucio, K.; Rabelo, A.C.S.; Araujo, C.M.; Brandao, G.C.; de Souza, G.H.B.; da Silva, R.G.; de Souza, D.M.S.; Talvani, A.; Bezerra, F.S.; Cruz Calsavara, A.J.; et al. Anti-inflammatory and antioxidant properties of black mulberry (Morus nigra L.) in a model of LPS-induced sepsis. Oxid. Med. Cell Longev. 2018, 2018, 5048031. [Google Scholar] [CrossRef]
- Lim, H.H.; Lee, S.O.; Kim, S.Y.; Yang, S.J.; Lim, Y. Anti-inflammatory and antiobesity effects of mulberry leaf and fruit extract on high fat diet-induced obesity. Exp. Biol. Med. (Maywood) 2013, 238, 1160–1169. [Google Scholar] [CrossRef]
- Lim, H.H.; Yang, S.J.; Kim, Y.; Lee, M.; Lim, Y. Combined treatment of mulberry leaf and fruit extract ameliorates obesity-related inflammation and oxidative stress in high fat diet-induced obese mice. J. Med. Food 2013, 16, 673–680. [Google Scholar] [CrossRef]
- Kim, H.K.; Cheon, B.S.; Kim, Y.H.; Kim, S.Y.; Kim, H.P. Effects of naturally occurring flavonoids on nitric oxide production in the macrophage cell line RAW 264.7 and their structure-activity relationships. Biochem. Pharmacol. 1999, 58, 759–765. [Google Scholar] [CrossRef]
- Hassimotto, N.M.; Moreira, V.; do Nascimento, N.G.; Souto, P.C.; Teixeira, C.; Lajolo, F.M. Inhibition of carrageenan-induced acute inflammation in mice by oral administration of anthocyanin mixture from wild mulberry and cyanidin-3-glucoside. BioMed Res. Int. 2013, 2013, 146716. [Google Scholar] [CrossRef]
- Ma, M.M.; Li, Y.; Liu, X.Y.; Zhu, W.W.; Ren, X.; Kong, G.Q.; Huang, X.; Wang, L.P.; Luo, L.Q.; Wang, X.Z. Cyanidin-3-O-glucoside ameliorates lipopolysaccharide-induced injury both in vivo and in vitro suppression of NF-kappaB and MAPK pathways. Inflammation 2015, 38, 1669–1682. [Google Scholar] [CrossRef]
- Gao, M.; Ma, Y.; Liu, D. Rutin suppresses palmitic acids-triggered inflammation in macrophages and blocks high fat diet-induced obesity and fatty liver in mice. Pharm. Res. 2013, 30, 2940–2950. [Google Scholar] [CrossRef]
- Kim, M.S.; Kim, S.H. Inhibitory effect of astragalin on expression of lipopolysaccharide-induced inflammatory mediators through NF-kappaB in macrophages. Arch. Pharm. Res. 2011, 34, 2101–2107. [Google Scholar] [CrossRef]
- Li, T.; Li, F.; Liu, X.; Liu, J.; Li, D. Synergistic anti-inflammatory effects of quercetin and catechin via inhibiting activation of TLR4-MyD88-mediated NF-kappaB and MAPK signaling pathways. Phytother. Res. 2019, 33, 756–767. [Google Scholar] [CrossRef]
- Lee, H.N.; Shin, S.A.; Choo, G.S.; Kim, H.J.; Park, Y.S.; Kim, B.S.; Kim, S.K.; Cho, S.D.; Nam, J.S.; Choi, C.S.; et al. Antiinflammatory effect of quercetin and galangin in LPSstimulated RAW264.7 macrophages and DNCB induced atopic dermatitis animal models. Int. J. Mol. Med. 2018, 41, 888–898. [Google Scholar] [CrossRef]
- Hamalainen, M.; Nieminen, R.; Vuorela, P.; Heinonen, M.; Moilanen, E. Anti-inflammatory effects of flavonoids: Genistein, kaempferol, quercetin, and daidzein inhibit STAT-1 and NF-kappaB activations, whereas flavone, isorhamnetin, naringenin, and pelargonidin inhibit only NF-kappaB activation along with their inhibitory effect on iNOS expression and NO production in activated macrophages. Mediators Inflamm. 2007, 2007, 45673. [Google Scholar] [CrossRef]
- Palacz-Wrobel, M.; Borkowska, P.; Paul-Samojedny, M.; Kowalczyk, M.; Fila-Danilow, A.; Suchanek-Raif, R.; Kowalski, J. Effect of apigenin, kaempferol and resveratrol on the gene expression and protein secretion of tumor necrosis factor alpha (TNF-alpha) and interleukin-10 (IL-10) in RAW-264.7 macrophages. Biomed. Pharmacother. 2017, 93, 1205–1212. [Google Scholar] [CrossRef]
- Lee, M.S.; Kim, Y. Effects of Isorhamnetin on Adipocyte Mitochondrial Biogenesis and AMPK Activation. Molecules 2018, 23, 1853. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
Sample Availability: Not available. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Content (mg/100 g) |
|---|---|
| Cyanidin 3-O-glucoside | 75.85 ± 2.79 |
| Cyanidin 3-O-rutinoside | 49.08 ± 1.92 |
| Pelargonidin 3-O-glucoside | 2.22 ± 0.06 |
| Total anthocyanins | 127.15 ± 4.73 |
| Compound | Content (mg/100 g) |
|---|---|
| Quercetin 3-O-rutinoside (rutin) | 8.05 ± 0.19 |
| Quercetin 3-O-glucoside (isoquercitrin) | 3.14 ± 0.11 |
| Quercetin 3-O-(6″-O-malonyl)glucoside | 1.22 ± 0.03 |
| Kaempferol 3-O-rutinoside (nicotiflorin) | 0.47 ± 0.03 |
| Kaempferol 3-O-glucoside (astragalin) | 0.37 ± 0.20 |
| Quercetin | 12.99 ± 0.45 |
| Kaempferol | 0.54 ± 0.00 |
| Total flavonols | 26.79 ± 0.94 |
| Gene 1 | GenBank Number | Primer Sequences (5′-3′) | Product Size (bp) |
|---|---|---|---|
| β-actin | NM_007393 | F: GGACCTGACAGACTACCTCA | 208 |
| R: GTTGCCAATAGTGATGACCT | |||
| NOS2 | BC062378.1 | F: GCTACTGGGTCAAAGACAAG | 191 |
| R: GCTGAACTTCCAGTCATTGT | |||
| PTGS2 | NM_011198.4 | F: GAACCTGCAGTTTGCTGTGG | 93 |
| R: ACTCTGTTGTGCTCCCGAAG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, S.; Lee, M.-S.; Choi, A.-J.; Kim, C.-T.; Kim, Y. Anti-Inflammatory Effects of High Hydrostatic Pressure Extract of Mulberry (Morus alba) Fruit on LPS-Stimulated RAW264.7 Cells. Molecules 2019, 24, 1425. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24071425
Jung S, Lee M-S, Choi A-J, Kim C-T, Kim Y. Anti-Inflammatory Effects of High Hydrostatic Pressure Extract of Mulberry (Morus alba) Fruit on LPS-Stimulated RAW264.7 Cells. Molecules. 2019; 24(7):1425. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24071425
Chicago/Turabian StyleJung, Sunyoon, Mak-Soon Lee, Ae-Jin Choi, Chong-Tai Kim, and Yangha Kim. 2019. "Anti-Inflammatory Effects of High Hydrostatic Pressure Extract of Mulberry (Morus alba) Fruit on LPS-Stimulated RAW264.7 Cells" Molecules 24, no. 7: 1425. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24071425