Design and Biological Evaluation of Colchicine-CD44-Targeted Peptide Conjugate in an In Vitro Model of Crystal Induced Inflammation



Abstract
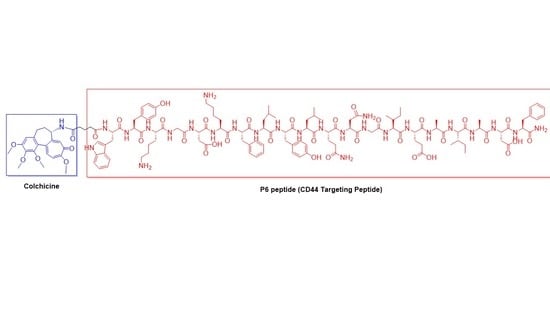
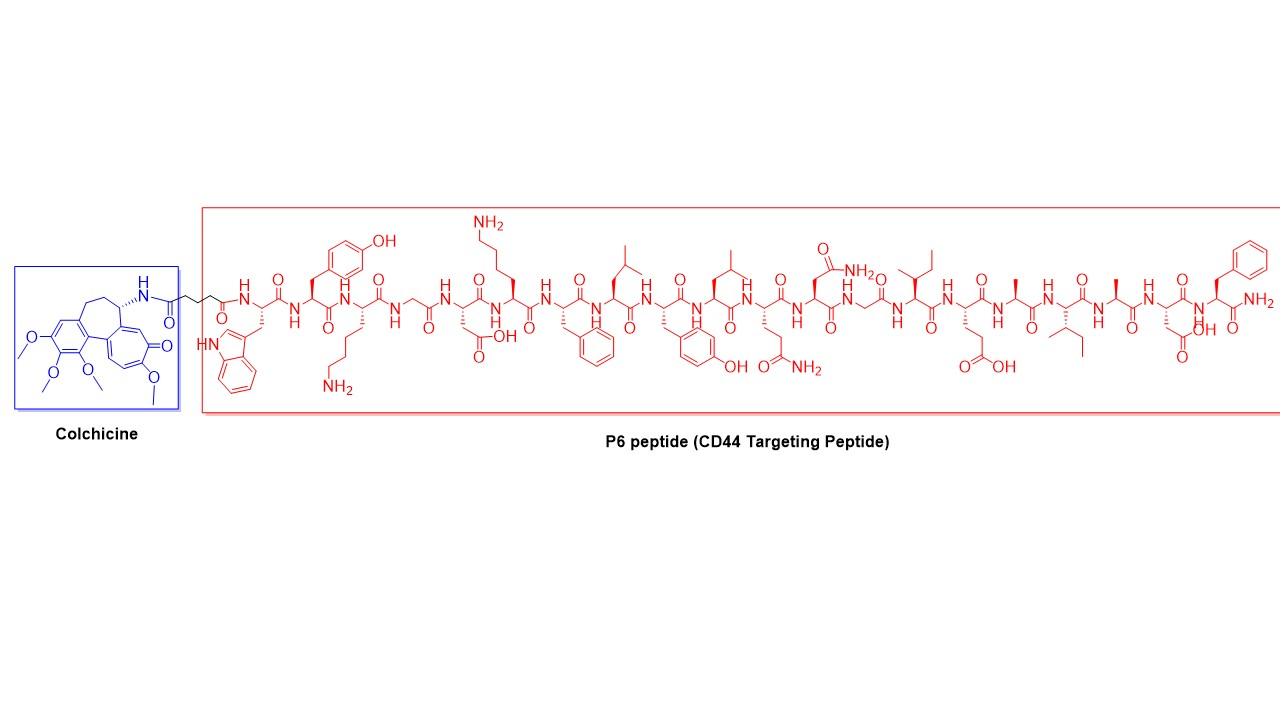
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
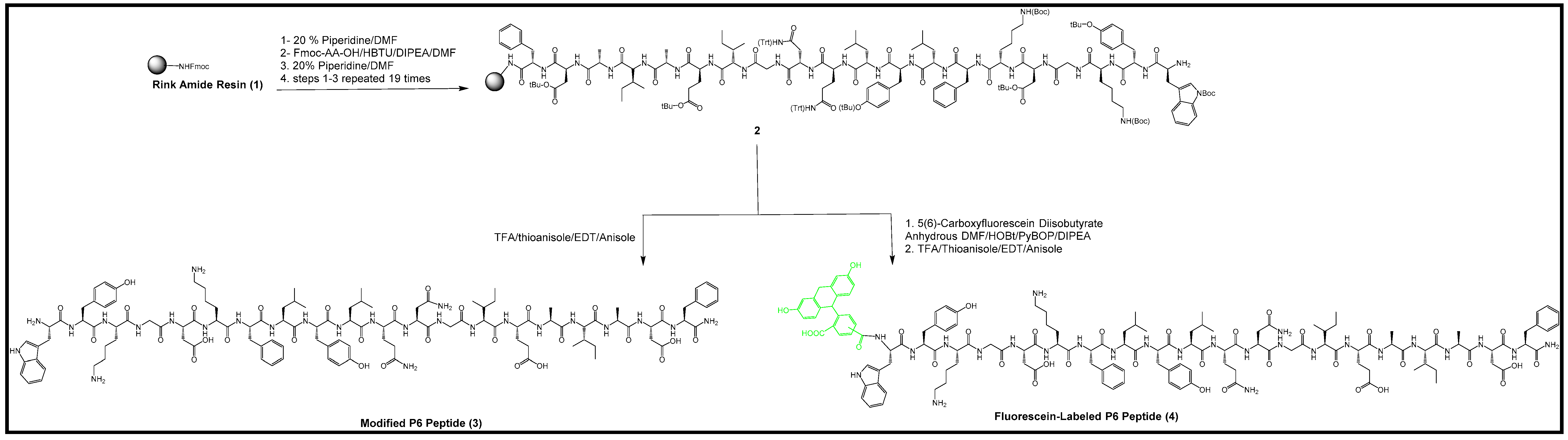
2.1. Synthesis of P6 Peptide
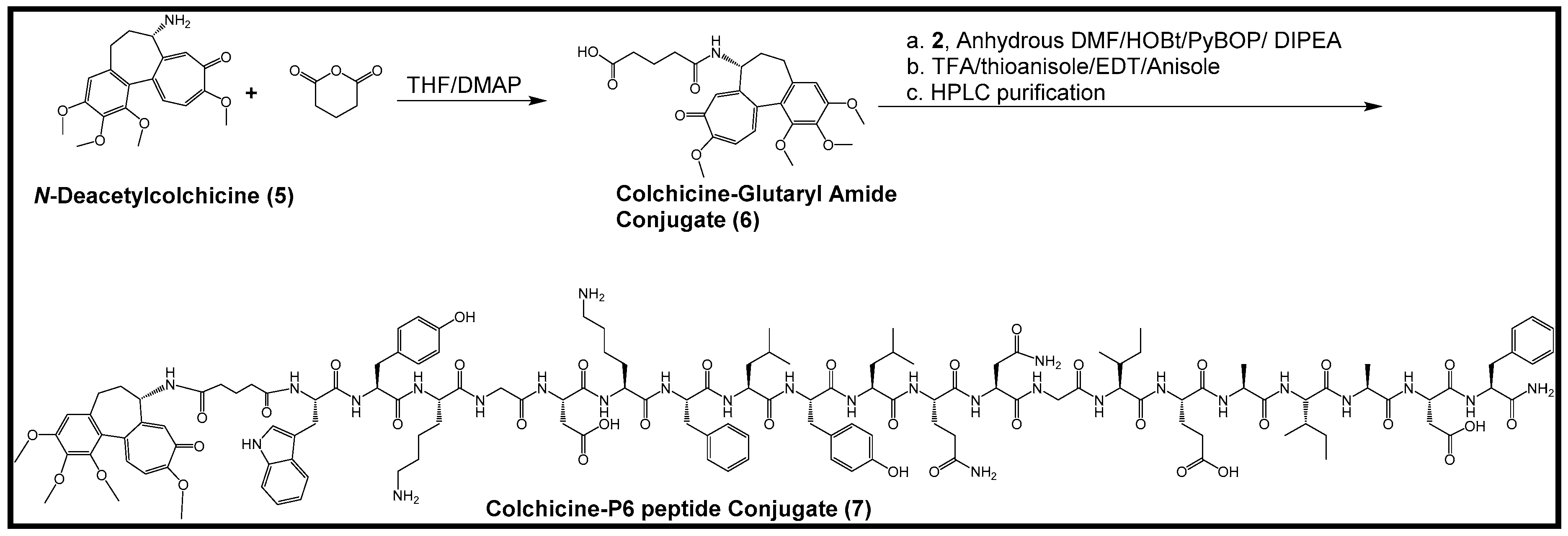
2.2. Synthesis of Colchicine-Glutaryl Amide Conjugate
2.3. Synthesis of Colchicine-P6 Peptide Conjugate
2.4. Synthesis of Fluorescein-Labeled P6 Peptide
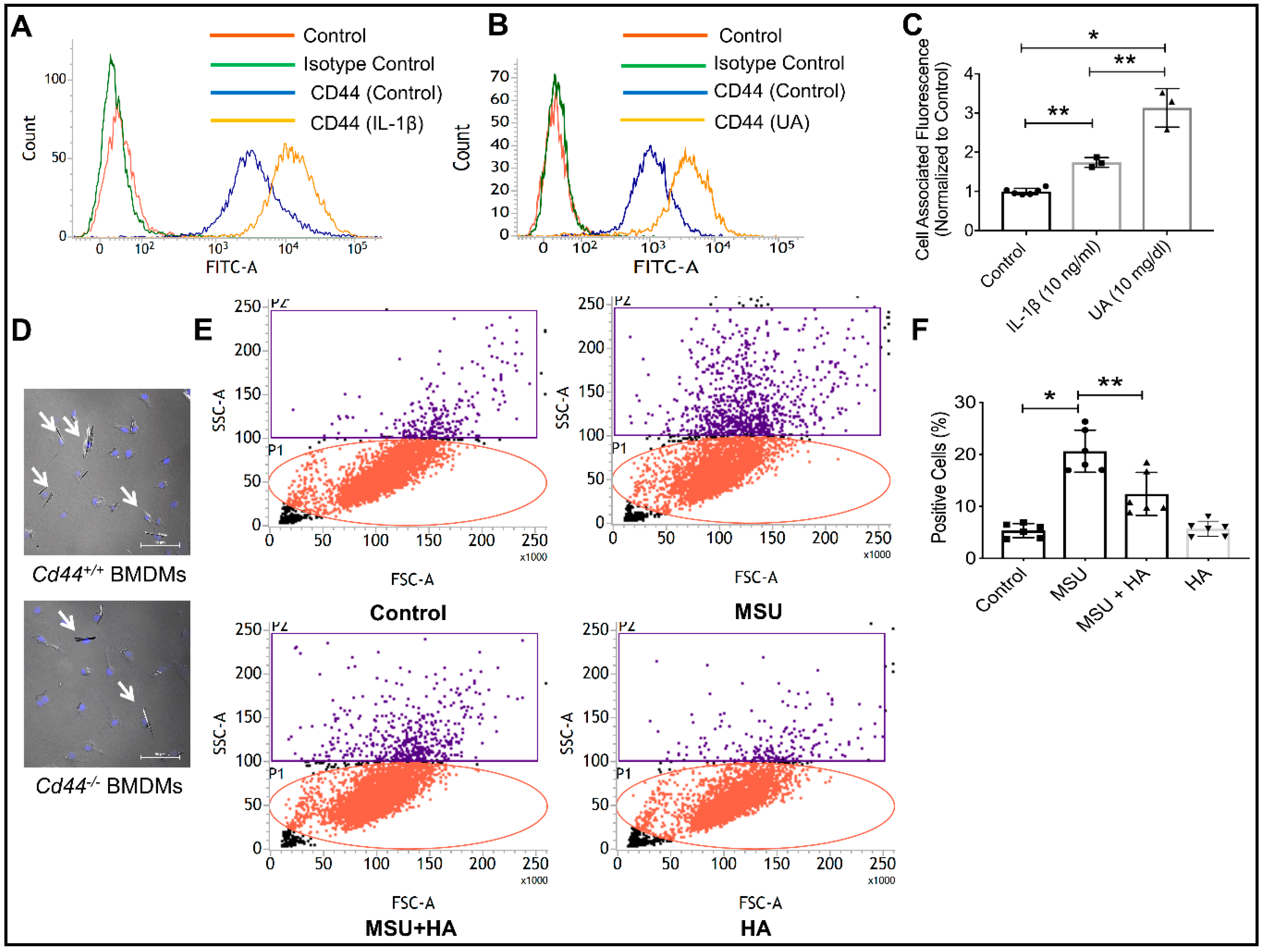
2.5. Characterization of CD44 Receptor Expression on Macrophages, Regulation under Conditions of Inflammation and Role of CD44 in Modulating Monosodium Urate (MSU) Crystal Phagocytosis by Macrophages
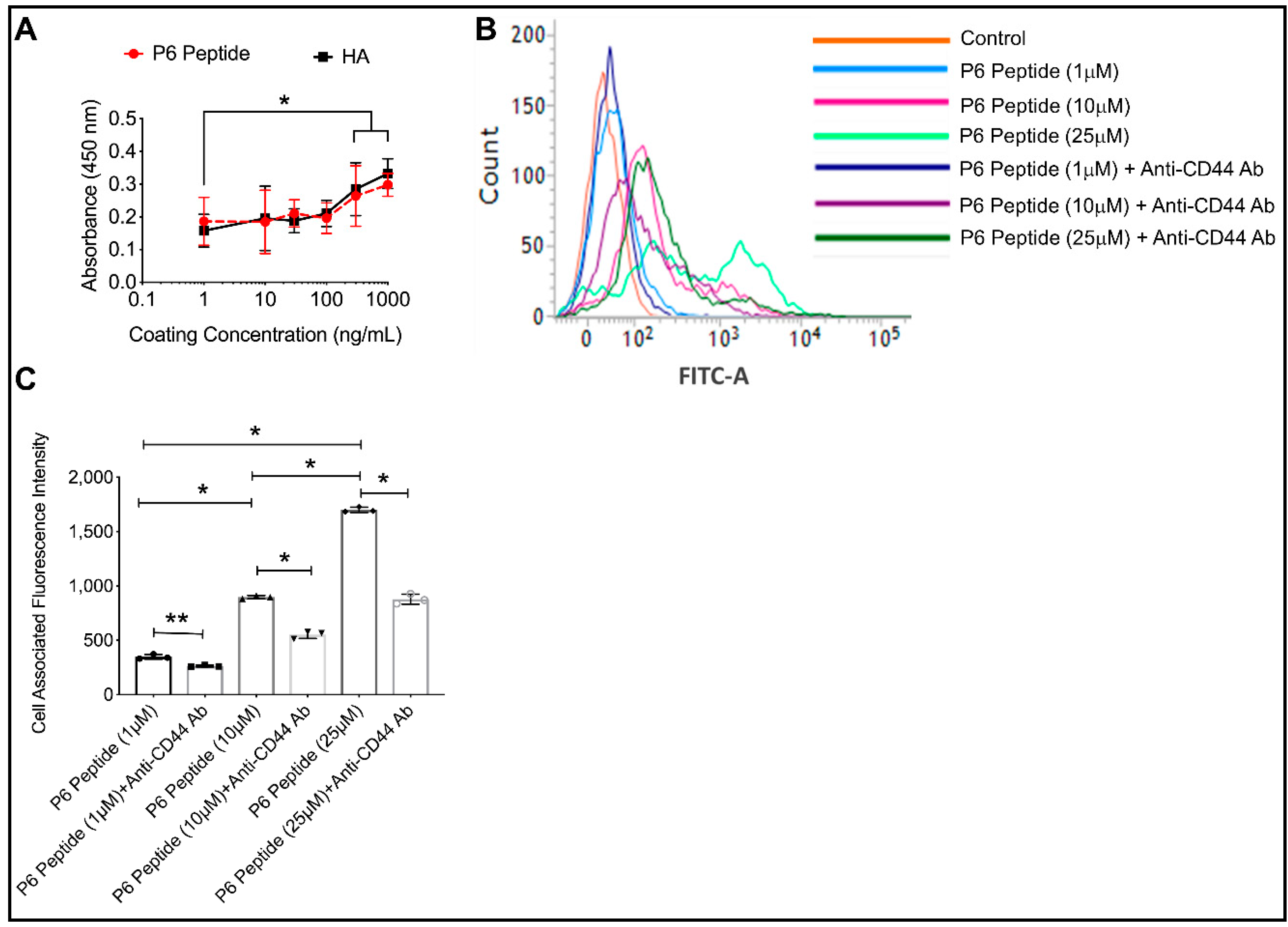
2.6. Comparative Binding of P6 Peptide and HA to CD44 Receptor and CD44 Dependent Association of P6 Peptide with THP-1 Macrophages
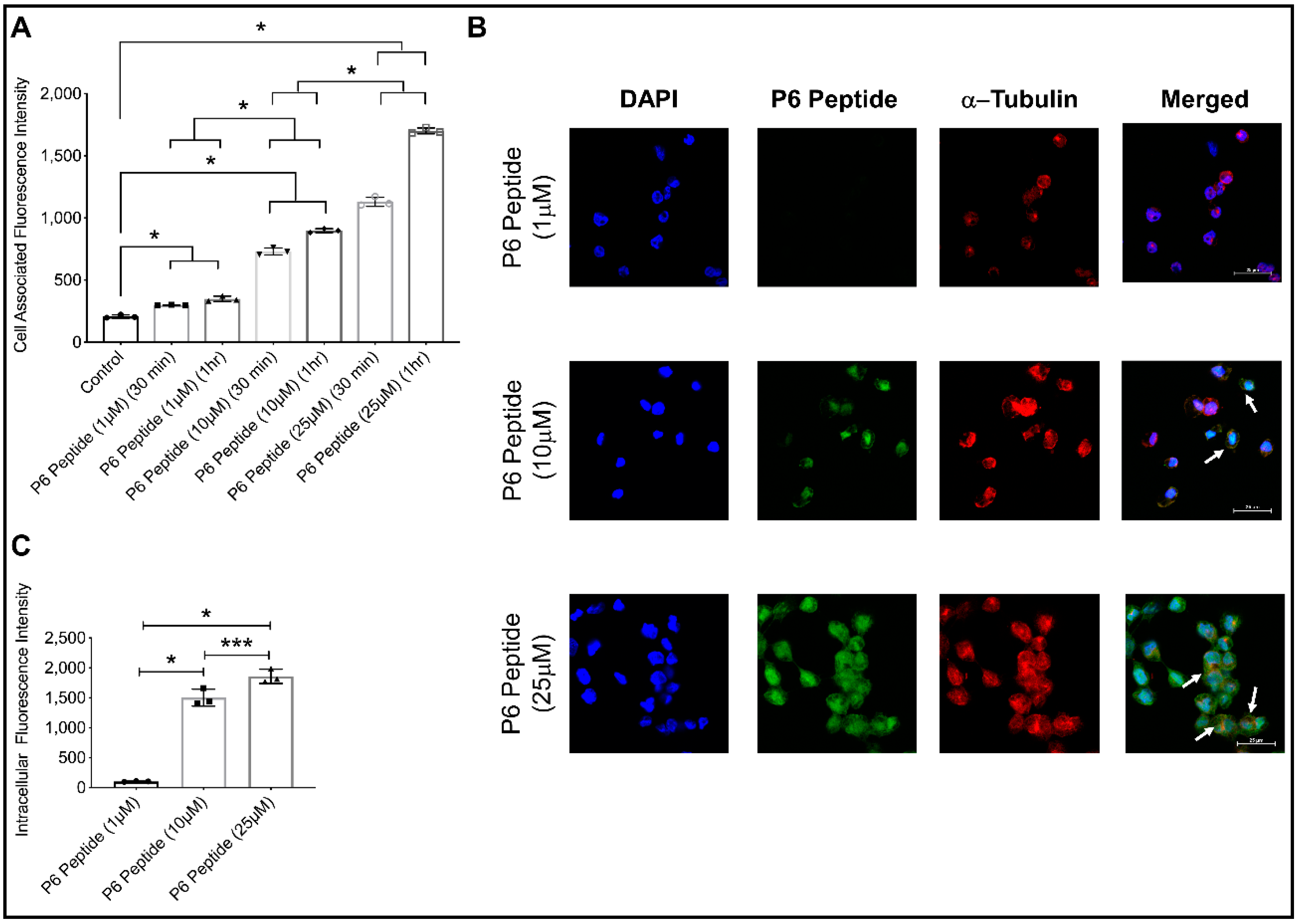
2.7. Concentration-Dependent Internalization of Fluorescein-P6 Peptide in THP-1 Macrophages Using Confocal Microscopy
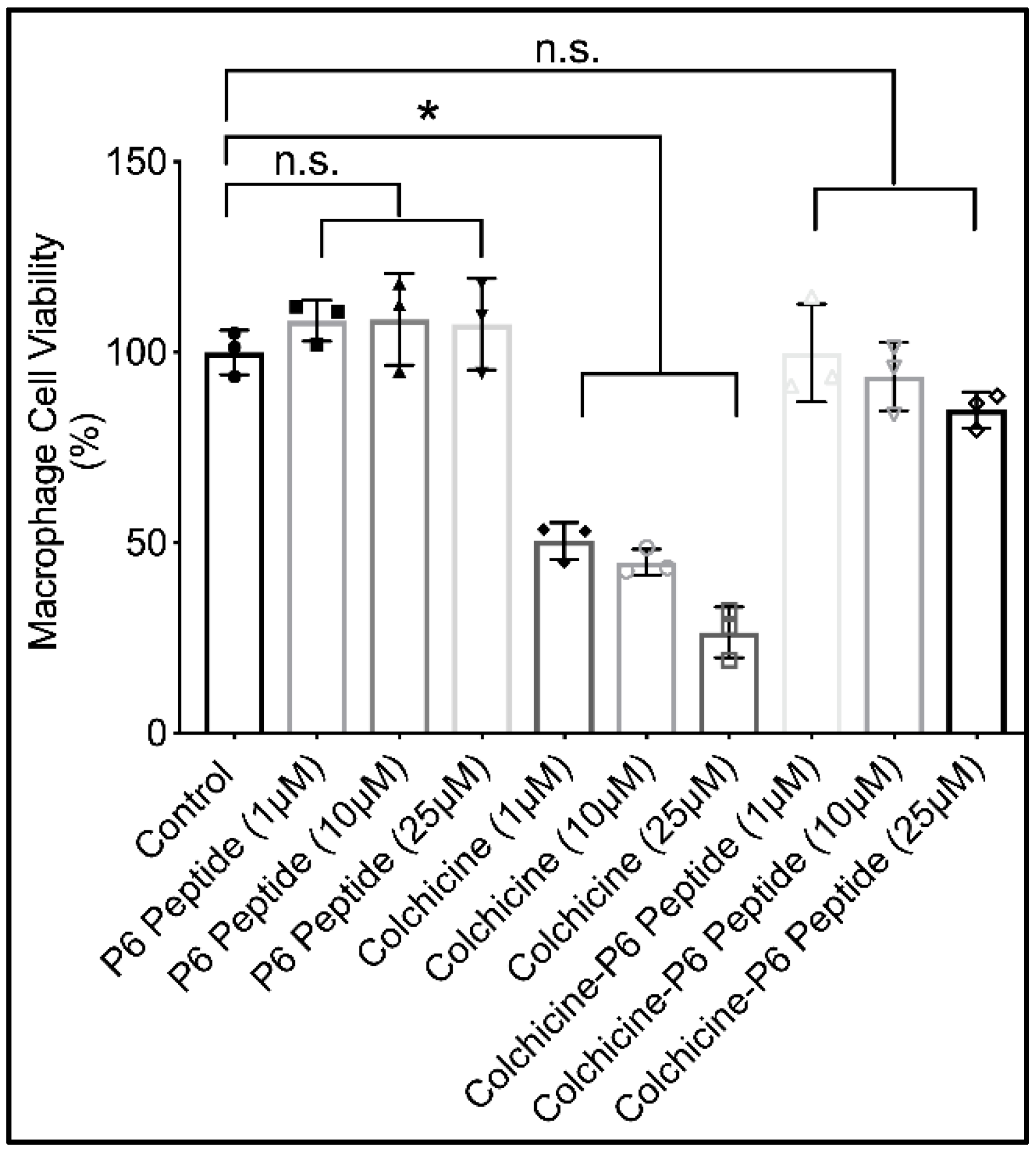
2.8. Cytotoxicity of P6 Peptide, Colchicine, and Colchicine-P6 Peptide Conjugate against Human THP-1 Macrophages
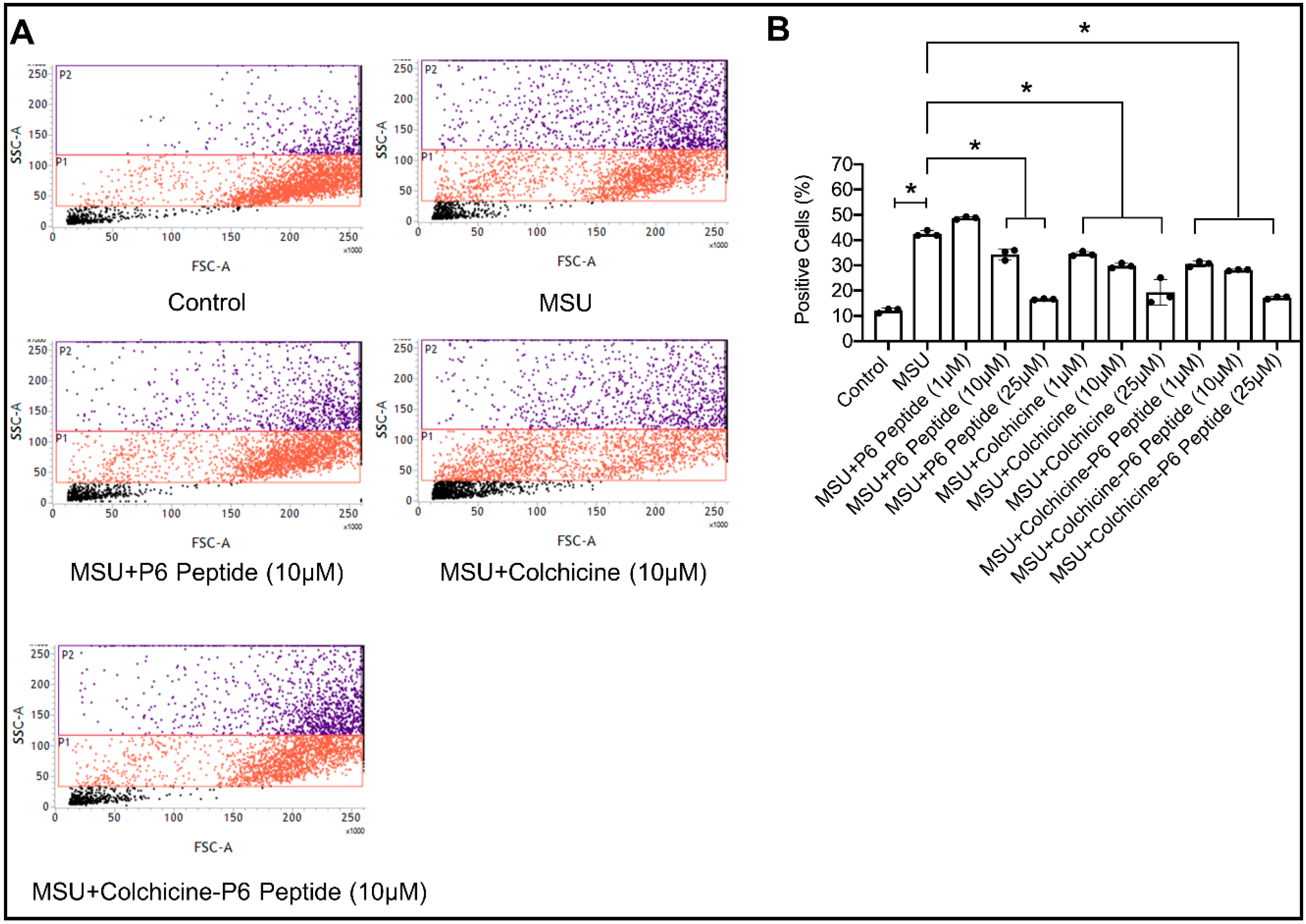
2.9. Impact of P6 Peptide, Colchicine, and Colchicine-P6 Peptide Treatments on Monosodium Urate (MSU) Crystal Phagocytosis by Human THP-1 Macrophages
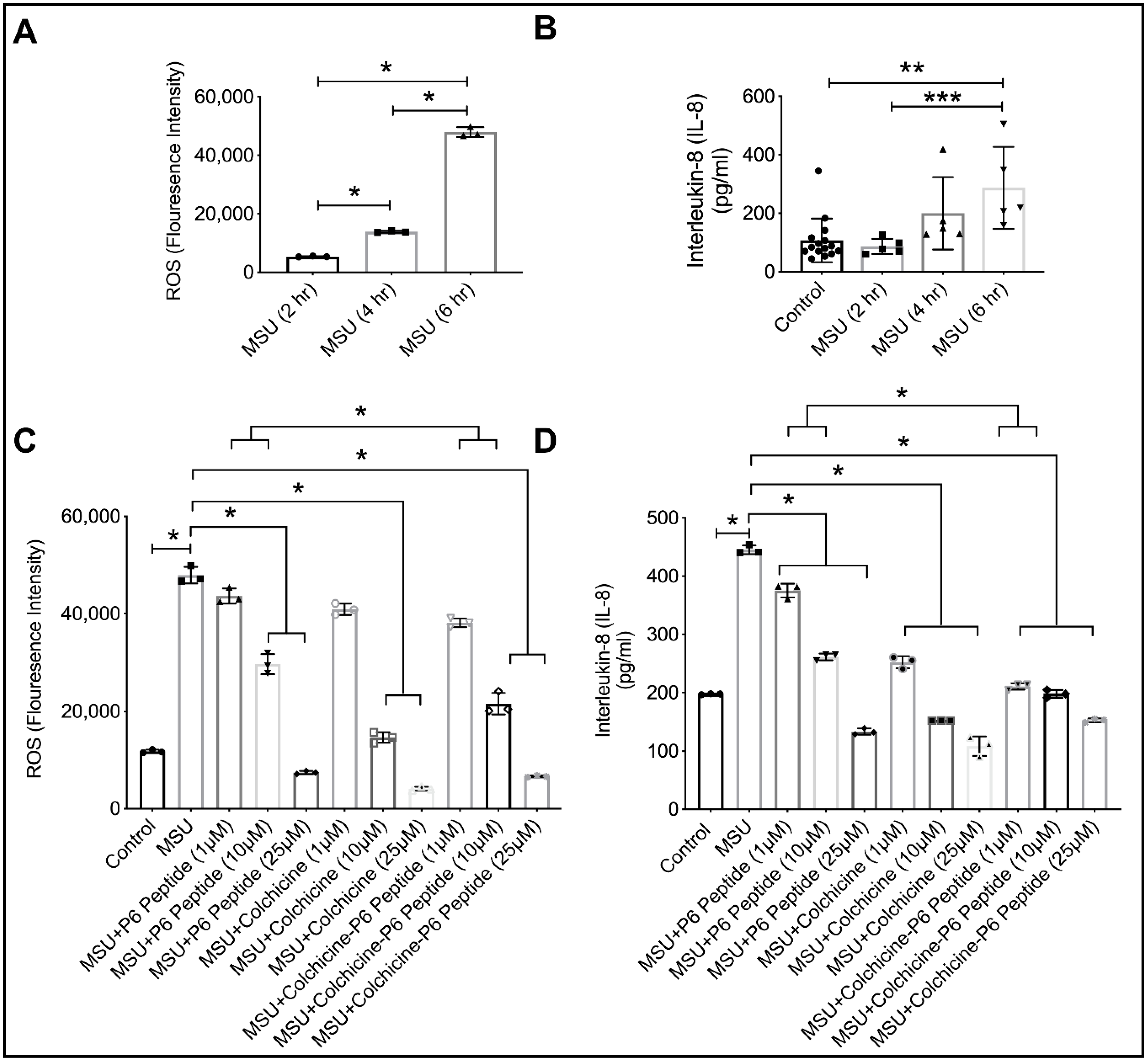
2.10. Anti-Inflammatory Efficacy of P6 Peptide, Colchicine, and Colchicine-P6 Peptide Treatments in MSU-Stimulated THP-1 Macrophages
2.11. Stability of P6 Peptide in Synovial Fluid (SF) Aspirates from Subjects with no History of Arthritis
3. Materials and Methods
3.1. Materials
3.2. Methods
3.2.1. Synthesis of P6 Peptide
3.2.2. Synthesis of Colchicine-Glutaryl Amide Conjugate
3.2.3. Synthesis of Colchicine-P6 Peptide Conjugate
3.2.4. Synthesis of Fluorescein-Labeled P6 Peptide
3.2.5. Differentiation of THP-1 Macrophages and CD44 Expression under Conditions of Inflammation, Isolation of BMDMs from Cd44+/+ and Cd44−/− Animals and Impact of HA Treatment on MSU Phagocytosis by THP-1 Macrophages
3.2.6. Comparative Binding of P6 Peptide and HA Using an Enzyme-Linked Immunosorbent Assay (ELISA) Format
3.2.7. CD44-Dependent Association and Internalization of Fluorescein-P6 Peptide in THP-1 Macrophages
3.2.8. Cytotoxicity of P6 Peptide, Colchicine, and Colchicine-P6 Peptide against THP-1 Macrophages
3.2.9. Impact of P6 Peptide, Colchicine, and Colchicine-P6 Peptide Treatments on MSU Phagocytosis by THP-1 Macrophages and Downstream ROS Generation and IL-8 Secretion
3.2.10. Stability of P6 Peptide in Normal Synovial Fluid (SF) Aspirates
3.2.11. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Roddy, E.; Choi, H.K. Epidemiology of gout. Rheum. Dis. Clin. 2014, 40, 155–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitik, B.; Akif Özturk, M. An old disease with new insights: Update on diagnosis and treatment of gout. Eur. J. Rheumatol. 2014, 1, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Stewart, S.; Dalbeth, N.; Vandel, A.; Rome, K. The first metatarsophalangeal joint in gout: A systematic review and meta-analysis. BMC Musculoskelet. Disord. 2016, 17, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Pandya, B.J.; Choi, H.K. Prevalence of gout and hyperuricemia in the US general population: The National Health and Nutrition Examination Survey 2007–2008. Arthritis Rheum. 2011, 63, 3136–3141. [Google Scholar] [CrossRef] [PubMed]
- Terkeltaub, R. What makes gouty inflammation so variable? BMC Med. 2017, 15, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalbeth, N.; Choi, H.K.; Terkeltaub, R. Review: A roadmap to approaches for improving global outcomes. Arthritis Rheumatol. 2017, 69, 22–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutton, I.; Gamble, G.; Gow, P.; Dalbeth, N. Factors associated with current hospital admissions for gout: A case-control study. J. Clin. Rheumatol. 2009, 15, 271–274. [Google Scholar] [CrossRef] [Green Version]
- Shield, G.E.; Beard, S.M. A systematic review of the economic and humanistic burden of gout. Pharmacoeconomics 2015, 33, 1029–1047. [Google Scholar] [CrossRef]
- Wechalekar, M.D.; Vinik, O.; Moi, J.H.; Sivera, F.; van Echteld, I.A.; van Dume, C.; Falzon, L.; Bombardier, C.; Carmona, L.; Aletaha, D.; et al. The efficacy and safety of treatments for acute gout: Results from a series of systematic literature reviews including cochrane reviews on intraarticular glucocorticoids, colchicine, nonsteroidal antiinflammatory drugs and interleukin-1 inhibitors. J. Rheumatol. Suppl. 2014, 92, 15–25. [Google Scholar] [CrossRef]
- Khanna, P.P.; Gladue, H.S.; Singh, M.K.; FitzGerald, J.D.; Bae, S.; Prakash, S.; Kaldas, M.; Gogia, M.; Berrocal, V.; Townsend, W.; et al. Treatment of acute gout: A systematic review. Semin. Arthritis Rheum. 2014, 44, 31–38. [Google Scholar] [CrossRef]
- van Echteld, I.; Wechalekar, M.D.; Schlesinger, N.; Buchbinder, R.; Aletaha, D. Colchicine for acute gout. Corchrane Database Syst. Rev. 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- Janssens, H.J.; Lucassen, P.L.; van de Laar, F.A.; Janssen, M.; Van de Lisonk, E.H. Systemic corticosteroids for acute gout. Cochrane Database Syst. Rev. 2008, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Durme, C.M.; Wechalekar, M.D.; Landewe, R.B. Nonsteroidal anti-inflammatory drugs for treatment of acute gout. JAMA 2015, 313, 2276–2277. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.D. The normal synovium. Open Rheumatol. J. 2011, 5, 100–106. [Google Scholar] [CrossRef]
- Kingsbury, S.R.; Conaghan, P.G.; McDermott, M.F. The role of the NLRP3 inflammasome in gout. J. Inflamm. Res. 2011, 4, 39–52. [Google Scholar]
- Busso, N.; So, A. Gout Mechanisms of inflammation in gout. Arthritis Res Ther. 2005, 44, 1090–1096. [Google Scholar]
- Amaral, F.; Costa, V.V.; Tavares, L.D.; Sachs, D.; Coelho, F.M.; Fagundes, C.T.; Soriani, F.M.; Silveira, T.N.; Cunha, L.D.; Zamboni, D.S.; et al. NLRP3 inflammasome-mediated neutrophil recruitment and Hypernociception depend on leukotriene B(4) in a murine model of gout. Arthritis Rheum. 2012, 64, 474–484. [Google Scholar] [CrossRef]
- Tramontini, N.; Huber, C.; Liu-Bryan, R.; Terkeltaub, R.; Kilgore, K.S. Central role of complement membrane attack complex in monosodium urate crystal-induced neutrophilic rabbit knee synovitis. Arthritis Rheum. 2004, 50, 2633–2639. [Google Scholar] [CrossRef]
- Martin, W.J.; Walton, M.; Harper, J. Resident macrophages initiating and driving inflammation in a monosodium urate monohydrate crystal-induced murine peritoneal model. Arthritis Rheum. 2009, 60, 281–289. [Google Scholar] [CrossRef]
- So, A.K.; Martinon, F. Inflammation in gout: Mechanisms and therapeutic targets. Nat. Rev. Rheumatol. 2017, 13, 639–647. [Google Scholar] [CrossRef]
- Liu-Bryan, R.; Scott, P.; Sydalske, A.; Rose, D.M.; Terkeltaub, R. Innate immunity conferred by toll-like receptors 2 and 4 and myeloid differentiation factor 88 expression is pivotal to monosodium urate monohydrate crystal-induced inflammation. Arthritis Rheum. 2005, 52, 2936–2946. [Google Scholar] [CrossRef] [PubMed]
- Scott, P.; Mia, H.; Viriyakosol, S.; Terkeltaub, R.; Liu-Bryan, R. Engagement of CD14 mediates the inflammatory potential of monosodium urate crystals. J. Immunol. 2006, 177, 6370–6378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brovold, H.; Lund, T.; Svistounov, D.; Solbu, M.D.; Jenssen, T.G.; Ytrehus, K.; Zykova, S.N. Crystallized but not soluble uric acid elicits pro-inflammatory response in short-term whole blood cultures from healthy men. Sci. Rep. 2019, 9, 10513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abias, J.M.; Xia, M.; Zhang, Y.; Boini, K.M.; Li, P.L. Redox regulation of NLRP3 inflammasomes: ROS as Trigger of Effector? Antioxid. Redox Signal. 2015, 22, 1111–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponta, H.; Sherman, L.; Herrlich, P. CD44: From adhesion molecules to signaling regulators. Nat. Rev. Mol. Cell Biol. 2003, 4, 33–45. [Google Scholar] [CrossRef]
- Tibesku, C.O.; Szuwart, T.; Ocken, S.A.; Skwara, A.; Fuchs, S. Expression of the matrix receptor CD44v5 on chondrocytes changes with osteoarthritis: An experimental investigation in the rabbit. Ann. Rheum. Dis. 2006, 65, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, S.; Dankbar, B.; Wildenau, G.; Goetz, W.; Lohmann, CH.; Tibesku, C.O. Expression of the CD44 variant isoform 5 in the human osteoarthritic knee joint: Correlation with radiological, histomorphological, and biochemical parameters. J. Orthop. Res. 2004, 22, 774–780. [Google Scholar] [CrossRef]
- Zhang, F.J.; Luo, W.; Gao, S.G.; Su, D.Z.; Li, Y.S.; Zeng, C.; Lei, G.H. Expression of CD44 in articular cartilage is associated with disease severity in knee osteoarthritis. Mod. Rheumatol. 2013, 23, 1186–1191. [Google Scholar] [CrossRef]
- Amash, A.; Wang, L.; Wang, Y.; Bhakta, V.; Faim, G.D.; Hou, M.; Peng, J.; Sheffield, W.P.; Lazarus, A.H. CD44 antibody inhibition of macrophage phagocytosis targets Fcγ receptor- and complement receptor 3-dependent mechanisms. J. Immunol. 2016, 196, 3331–3340. [Google Scholar] [CrossRef] [Green Version]
- Al-Sharif, A.; Jamal, M.; Zhang, L.X.; Larson, K.; Schmidt, T.A.; Jay, G.D.; Elsaid, K.A. Lubricin/proteoglycan 4 binding to CD44 receptor: A mechanism of the suppression of proinflammatory cytokine-induced synoviocyte proliferation by lubricin. Arthritis Rheumatol. 2015, 67, 1503–1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knudson, W.; Chow, G.; Knudson, C.B. CD44-mediated uptake and degradation of hyaluronan. Matrix Biol. 2002, 21, 15–23. [Google Scholar] [CrossRef]
- Qadri, M.; Jay, G.D.; Zhang, L.X.; Wong, W.; Reginato, A.M.; Sun, C.; Schmidt, T.A.; Elsaid, K.A. Recombinant human proteoglycan-4 reduces phagocytosis of urate crystals and downstream nuclear factor kappa B and inflammasome activation and production of cytokines and chemokines in human and murine macrophages. Arthritis Res Ther. 2018, 20, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rios de la Rosa, J.M.; Tirella, A.; Gennari, A.; Stratford, I.J.; Tirelli, N. The CD44-mediate uptake of hyaluronic acid-based carriers in macrophages. Adv. Healthc. Mater. 2017, 6, 1601012. [Google Scholar] [CrossRef]
- Farajzadeh, R.; Zarghami, N.; Serati-Nouri, H.; Momeni-Javid, Z.; Farajzadeh, T.; Jalilzadeh-Tabrizi, S.; Sadeghi-Soureh, S.; Naseri, N.; Pilehvar-Soltanahmadi, Y. Macrophage repolarization using CD44-targeting hyaluronic acid-polylactide nanoparticles containing curcumin. Artif. Cells Nanomed. Biotechnol. 2018, 46, 2013–2021. [Google Scholar] [CrossRef] [Green Version]
- Dalbeth, N.; Lauterio, T.J.; Wolfe, H.R. Mechanism of action of colchicine in the treatment of gout. Clin. Ther. 2014, 36, 1465–1479. [Google Scholar] [CrossRef] [Green Version]
- Leung, Y.Y.; Yao Hui, L.L.; Kraus, V.B. Colchicine—update on mechanisms of action and therapeutic used. Semin. Arthritis Rheum. 2015, 45, 341–350. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Sharma, P.R.; Mondhe, D.M. Potential anticancer role of colchicine-based derivatives. Anti Cancer Drugs 2017, 28, 250–262. [Google Scholar] [CrossRef]
- Marques-da Silva, C.; Chaves, M.M.; Castro, N.G.; Coutinho-Silva, R.; Guimaraes, M.Z.P. Colchicine inhibits cationic dye uptake induced by ATP in P2X2 and P2X7 receptor-expressing cells: Implication for its therapeutic action. Br. J. Pharmacol. 2011, 163, 912–926. [Google Scholar] [CrossRef] [Green Version]
- Finkelstein, Y.; Aks, S.E.; Hutson, J.R.; Juurlink, D.N.; Nguyen, P.; Dubnov-Raz, G.; Pollak, U.; Koren, G.; Bentur, Y. Colchicine poisoning: The dark side of an ancient drug. Clin. Toxicol. 2010, 48, 407–414. [Google Scholar] [CrossRef]
- Owen, S.G.; Francis, H.W.; Roberts, M.S. Disappearance kinetics of solutes from synovial fluid after intra-articular injections. Br. J. Clin. Pharmacol. 1994, 38, 349–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerwin, N.; Hops, C.; Lucke, A. Intraarticular drug delivery in osteoarthritis. Adv. Drug Deliv. Rev. 2006, 58, 226–242. [Google Scholar] [CrossRef] [PubMed]
- Rudnik-Jansen, I.; Schrijver, K.; Woike, N.; Tellegen, A.; Versteeg, S.; Emans, P.; Mihov, G.; Thies, J.; Eijkelkamp, N.; Tryfoniduo, M.; et al. Intra-articular injection of triamcinolone acetonide releasing biomaterial microspheres inhibits pain and inflammation in an acute arthritis model. Drug Deliv. 2019, 26, 226–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsaid, K.A.; Ubhe, A.; Shaman, Z.; D’Souza, G. Intra-articular interleukin-1 receptor antagonist (IL1-ra) microspheres for posttraumatic osteoarthritis: In vitro biological activity and in vivo disease modifying effect. J. Exp. Orthop. 2016, 3, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Cheetham, A.G.; Angacian, G.; Su, H.; Xie, L.; Cui, H. Peptide–drug conjugates as effective prodrug strategies for targeted delivery. Adv. Drug Deliv. Rev. 2017, 110, 112–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ugarte-Berzal, E.; Bailón, E.; Amigo-Jiménez, I.; Albar, J.P.; García-Marco, J.A.; García-Pardo, A. A novel CD44-binding peptide from the pro-matrix metalloproteinase-9 hemopexin domain impairs adhesion and migration of chronic lymphocytic leukemia (CLL) cells. J. Biol. Chem. 2014, 289, 15340–15349. [Google Scholar] [CrossRef] [Green Version]
- Scanu, A.; Oliviero, F.; Ramonda, R.; Frallonardo, P.; Dayer, J.M.; Punzi, L. Cytokine levels in human synovial fluid during the different stages of acute gout: Role of transforming growth factor β1 in the resolution phase. Ann. Rheum. Dis. 2012, 71, 621–624. [Google Scholar] [CrossRef]
- Powell, M.F.; Grey, H.; Gaeta, F.; Sette, A.; Colon, S. Peptide stability in drug development: A comparison of peptide reactivity in different biological media. J. Pharm. Sci. 1992, 81, 731–735. [Google Scholar] [CrossRef]
- Yamagiwa, H.; Sarkar, G.; Charlesworth, M.C.; McCormick, D.J.; Bolander, M.E. Two dimensional gel electrophoresis of synovial fluid: Method for detecting candidate protein markers for osteoarthritis. J. Orhop. Sci. 2003, 8, 482–490. [Google Scholar] [CrossRef]
- Bennike, T.; Ayturk, U.; Haslauer, C.M.; Froehlich, J.W.; Proffen, B.L.; Barnaby, O.; Birkelund, S.; Murray, M.M.; Warman, M.L.; Stensballe, A.; et al. A normative study of synovial fluid proteome from healthy porcine knee joints. J. Proteome Res. 2014, 13, 4377–4387. [Google Scholar] [CrossRef]
- Bottiger, R.; Hoffmann, R.; Knappe, D. Differential stability of therapeutic peptides with different proteolytic cleavage sites in blood, plasma and serum. PLoS ONE. 2017, 12, e0178943. [Google Scholar] [CrossRef] [PubMed]
- Qadri, M.; Almadani, S.; Jay, G.D.; Elsaid, K.A. Role of CD44 in regulating TLR2 activation of human macrophages and downstream expression of proinflammatory cytokines. J. Immunol. 2018, 200, 758–767. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zoghebi, K.A.; Bousoik, E.; Parang, K.; Elsaid, K.A. Design and Biological Evaluation of Colchicine-CD44-Targeted Peptide Conjugate in an In Vitro Model of Crystal Induced Inflammation. Molecules 2020, 25, 46. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25010046
Zoghebi KA, Bousoik E, Parang K, Elsaid KA. Design and Biological Evaluation of Colchicine-CD44-Targeted Peptide Conjugate in an In Vitro Model of Crystal Induced Inflammation. Molecules. 2020; 25(1):46. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25010046
Chicago/Turabian StyleZoghebi, Khalid A., Emira Bousoik, Keykavous Parang, and Khaled A. Elsaid. 2020. "Design and Biological Evaluation of Colchicine-CD44-Targeted Peptide Conjugate in an In Vitro Model of Crystal Induced Inflammation" Molecules 25, no. 1: 46. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25010046