
The Sensory Significance of Apocarotenoids in Wine: Importance of Carotenoid Cleavage Dioxygenase 1 (CCD1) in the Production of β-Ionone



Abstract
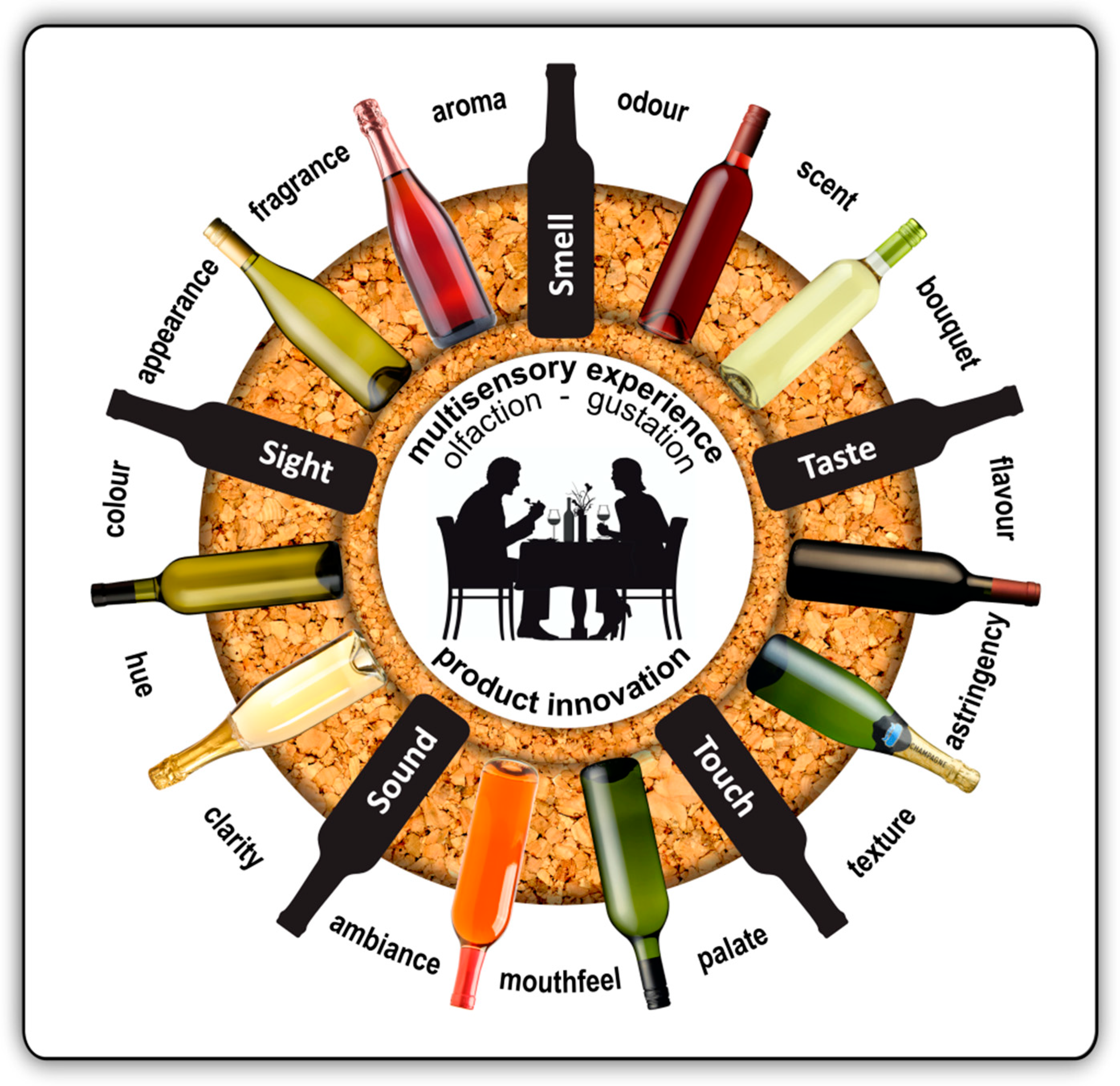
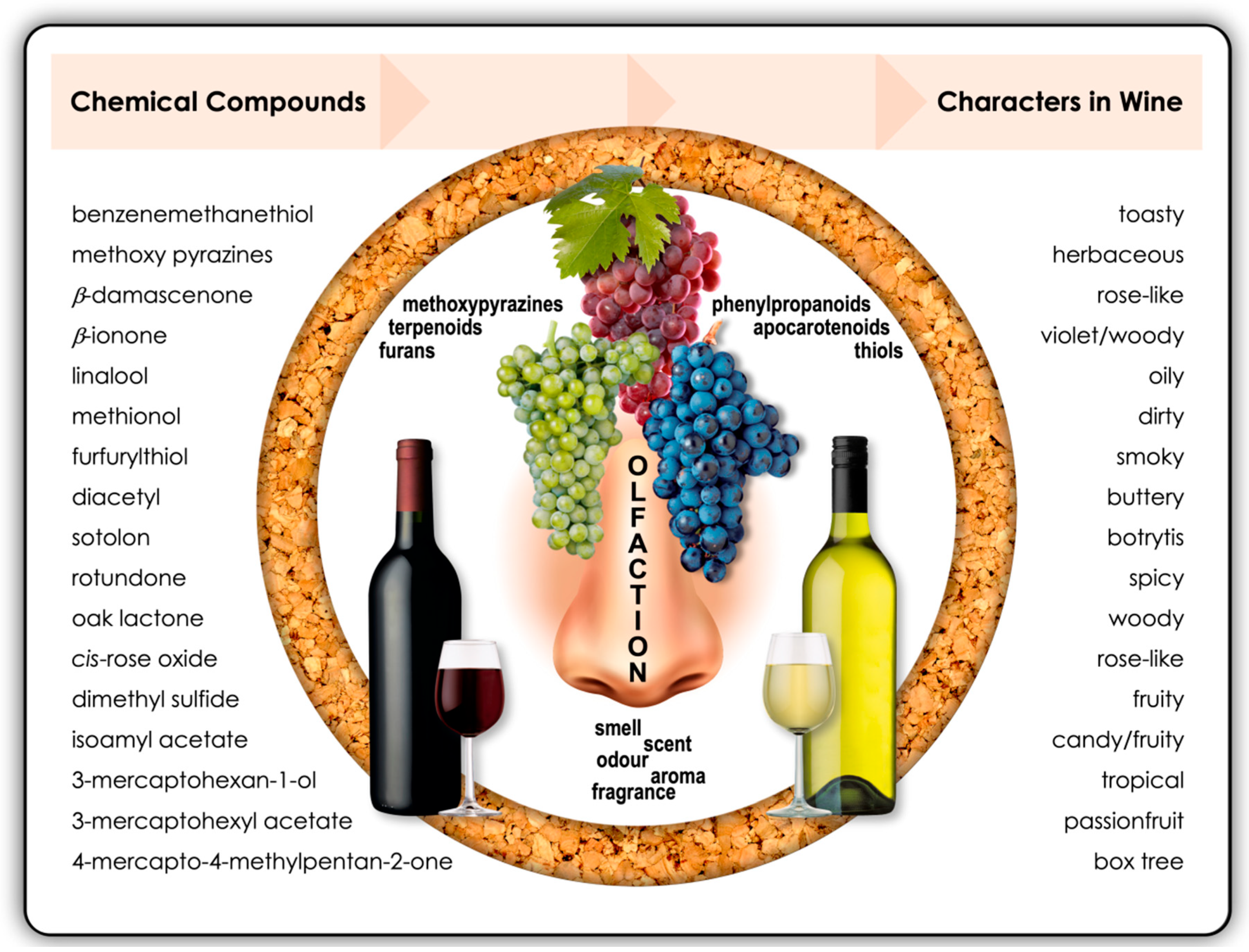
:1. Multisensory Flavour Perception
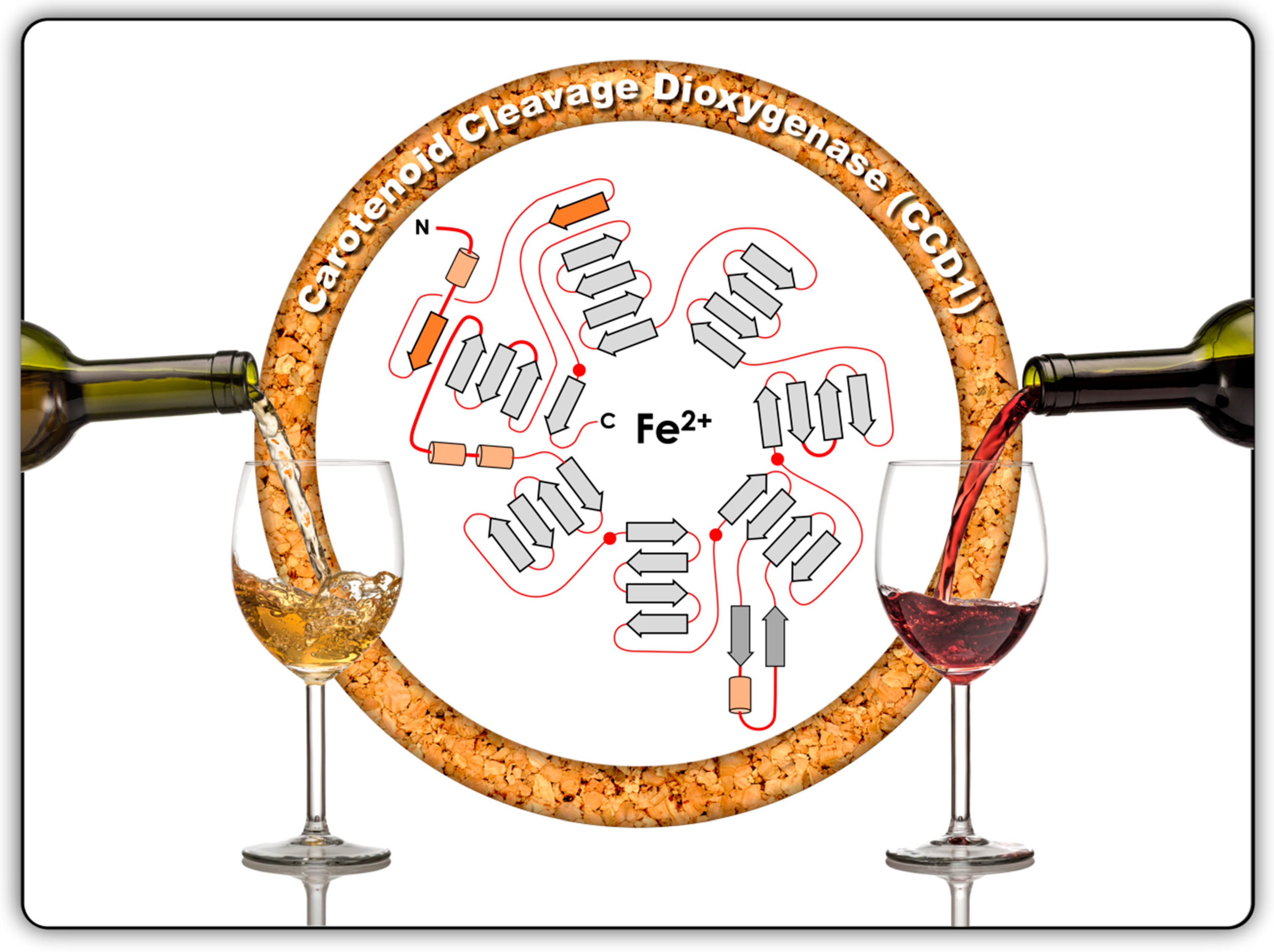
2. Classification, Structure and Sources of CCDs
3. Sources of Carotenoid Cleavage Dioxygenase 1
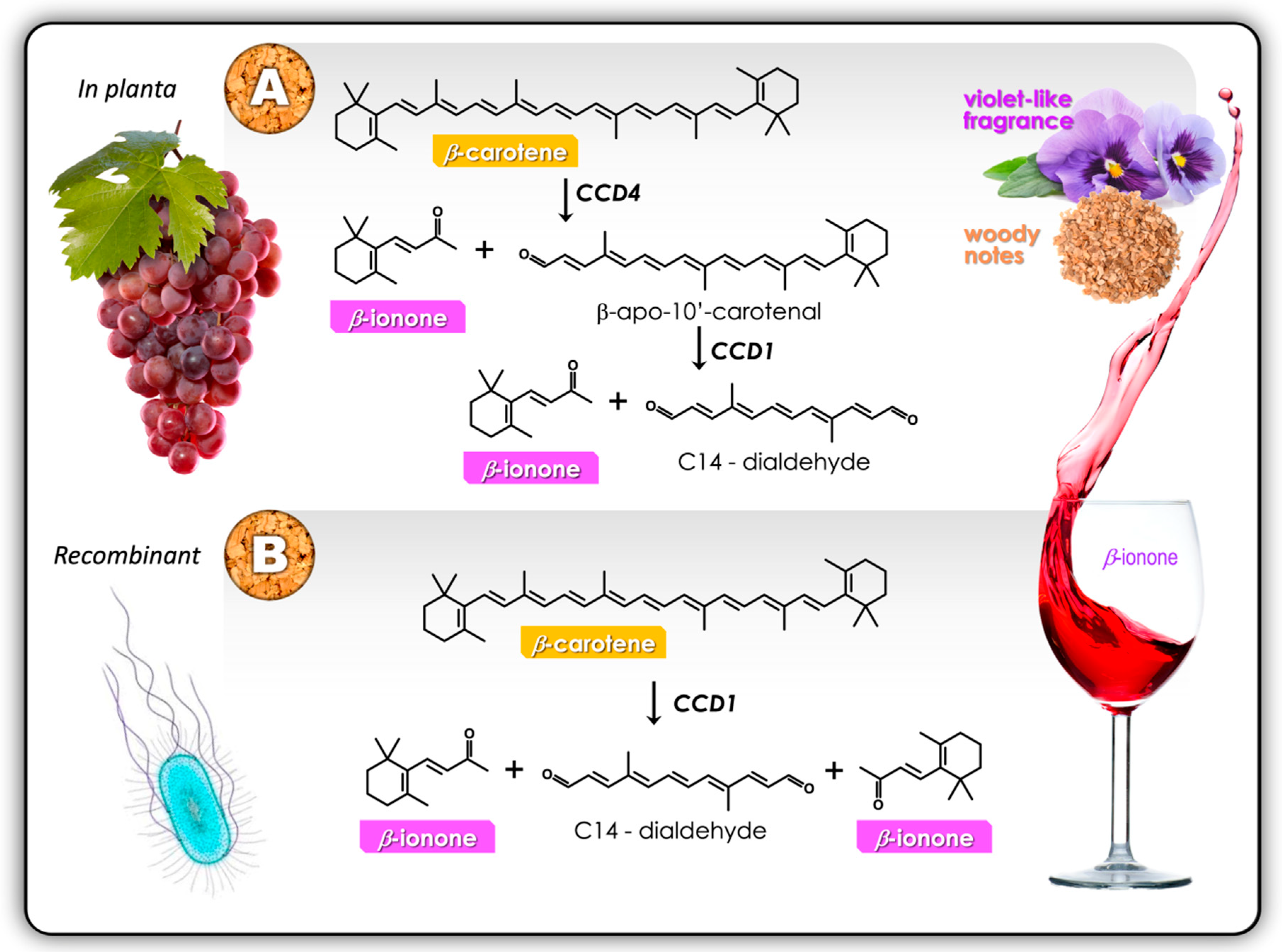
4. Biological Functions of CCDs
5. In Planta Conclusions for CCD1
6. Biotechnology of CCDs
7. A Taste of the Future
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Spence, C.; Youssef, J. Olfactory dining: Designing for the dominant sense. Flavour 2015, 4, 32–47. [Google Scholar] [CrossRef] [Green Version]
- Spence, C. Multisensory flavour perception. Cell 2015, 26, 24–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swiegers, J.H.; Chambers, P.J.; Pretorius, I.S. The genetics of olfaction and taste. Aust. J. Grape Wine Res. 2005, 11, 109–113. [Google Scholar] [CrossRef]
- Wallace, M.T. Multisensory perception: The building of flavor representations. Curr. Biol. 2015, 25, 986–988. [Google Scholar] [CrossRef] [Green Version]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and future of non-Saccharomyces yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef] [Green Version]
- Swiegers, J.H.; Capone, D.L.; Pardon, K.H.; Elsey, G.M.; Sefton, M.A.; Francis, I.L.; Pretorius, I.S. Engineering volatile thiol release in Saccharomyces cerevisiae for improved wine aroma. Yeast 2007, 24, 561–574. [Google Scholar] [CrossRef]
- Schwartz, S.H.; Qin, X.; Zeevaart, J.A.D. Characterisation of a novel carotenoid cleavage dioxygenase from plants. J. Biol. Chem. 2001, 276, 25208–25211. [Google Scholar] [CrossRef] [Green Version]
- Kloer, D.P.; Ruch, S.; Al-Babili, S.; Beyer, P.; Schulz, G.E. The structure of a retinal-forming carotenoid oxygenase. Science 2005, 308, 267–269. [Google Scholar] [CrossRef] [Green Version]
- Kloer, D.P.; Schulz, G.E. Structure and biological aspects of carotenoid cleavage. Cell. Mol. Life Sci. 2006, 63, 2291–2303. [Google Scholar] [CrossRef]
- Cui, H.; Wang, Y.; Qin, S. Genomewide analysis of carotenoid cleavage dioxygenase in unicellular and filamentous cyanobacteria. Comp. Funct. Genom. 2012, 164690, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiefer, C.; Hessel, S.; Lampert, J.M.; Vogt, K.; Lederer, M.O.; Breithaupt, D.E.; von Lintig, J. Identification and characterisation of a mammalian enzyme catalysing the asymmetric oxidative cleavage of provitamin A. J. Biol Chem. 2001, 276, 14110–14116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, J.T.; Tan, B.-C.; McCarty, D.R.; Klee, H.J. The carotenoid cleavage dioxygenase 1 enzyme has broad substrate specificity, cleaving multiple carotenoids at two different bond positions. J. Biol. Chem. 2008, 283, 11364–11373. [Google Scholar] [CrossRef] [Green Version]
- Priya, R.; Siva, R. Phylogenic analysis and evolutionary studies of plant carotenoid cleavage dioxygenase gene. Gene 2014, 548, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Frusciante, S.; Diretto, G.; Bruno, M.; Ferrante, P.; Pietrella, M.; Prado-Cabrero, A.; Rubio-Moraga, A.; Beyer, P.; Gomez-Gomez, L.; Al-Babili, S.; et al. Novel carotenoid cleavage dioxygenase catalyzes the first dedicated step in saffron crocin biosynthesis. Proc. Natl. Acad. Sci. USA 2014, 111, 12246–12251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahrazem, O.; Rubio-Moraga, A.; Berman, J.; Capell, T.; Christou, P.; Zhu, C.; Gomez-Gomez, L. The carotenoid cleavage dioxygenase CCD2 catalysing the synthesis of crocetin in spring crocuses and saffron is a plastidial enzyme. New Phytol. 2016, 209, 650–663. [Google Scholar] [CrossRef]
- Demurtas, O.C.; Frusciante, S.; Ferrante, P.; Diretto, G.; Azad, N.H.; Pietrella, M.; Aprea, G.; Taddei, A.R.; Romano, E.; Mi, J.; et al. Candidate enzymes for saffron crocin biosynthesis are localized in multiple cellular compartments. Plant Physiol. 2018, 177, 990–1006. [Google Scholar] [CrossRef] [Green Version]
- Baba, S.A.; Mohiuddin, T.; Basu, S.; Swarnkar, M.K.; Malik, A.H.; Wani, Z.A.; Abbas, N.; Singh, A.K.; Ashraf, N. Comprehensive transcriptome analysis of Crocus sativus for discovery and expression of genes involved in apocarotenoid biosynthesis. BMC Genom. 2015, 16, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Auldridge, M.E.; McCarty, D.R.; Klee, H.J. Plant carotenoid cleavage oxygenases and their apocarotenoid products. Curr. Opin. Plant Biol. 2006, 9, 315–321. [Google Scholar] [CrossRef]
- Baldermann, S.; Kato, M.; Kurosawa, M.; Kurobayashi, Y.; Fujita, A.; Fleischmann, P.; Watanabe, N. Functional characterization of a carotenoid cleavage dioxygenase 1 and its relation to the carotenoid accumulation and volatile emission during the floral development of Osmanthus fragrans Lour. J. Exp. Bot. 2010, 61, 2967–2977. [Google Scholar] [CrossRef] [Green Version]
- Baldermann, S.; Kato, M.; Fleischmann, P.; Watanabe, N. Biosynthesis of a-ionone and β-ionone, prominent scent compounds, in flowers of Osmanthus fragrans. Acta Biochim. Pol. 2012, 59, 79–81. [Google Scholar] [CrossRef]
- Lashbrooke, J.G.; Young, P.R.; Dockrall, S.J.; Vasanth, K.; Vivier, M.A. Functional characterisation of three members of the Vitis vinifera L. carotenoid cleavage dioxygenase gene family. BMC Plant Biol. 2013, 13, 156–172. [Google Scholar] [CrossRef] [Green Version]
- Rubio, A.; Rambla, J.L.; Santaella, M.; Gomez, M.D.; Orzaez, D.; Granell, A.; Gomez-Gomez, L. Cytosolic and plastoglobule-targeted carotenoid dioxygenases from Crocus sativus are both involved in β-ionone release. J. Biol. Chem. 2008, 283, 24816–24825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simkin, A.J.; Underwood, B.A.; Auldridge, M.; Loucas, H.M.; Shibuya, K.; Schmelz, E.; Clark, D.G.; Klee, H.J. Circadian regulation of the PhCCD1 carotenoid cleavage dioxygenase controls emission of β-ionone, a fragrance volatile of petunia flowers. Plant Physiol. 2004, 136, 3504–3514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, M.H.; Strack, D. Carotenoids and their cleavage products: Biosynthesis and functions. Nat. Prod. Rep. 2011, 28, 663–692. [Google Scholar] [CrossRef]
- Kaiser, R. New Volatile Constituents of the Flower Concrete of Michelia champaca L. J. Essent. Oil Res. 1991, 3, 129–146. [Google Scholar] [CrossRef]
- MacTavish, H.; Davies, N.W.; Menary, R.C. Carotenoid-Derived Aroma Compounds; Winterhalter, P., Rouseff, R.L., Eds.; ACS Publications: Washington, DC, USA, 2001; Volume 802, Chapter 13; pp. 183–193. [Google Scholar]
- Ahrazem, O.; Diretto, G.; Argandona, J.; Rubio-Moraga, A.; Julve, J.M.; Orzaez, D.; Granell, A.; Gomez-Gomez, L. Evolutionarily distinct carotenoid cleavage dioxygenases are responsible for crocetin production in Buddleja davidii. J. Exp. Bot. 2017, 68, 4663–4677. [Google Scholar] [CrossRef]
- Marasco, E.K.; Vay, K.; Schmidt-Dannert, C. Identification of carotenoid cleavage dioxygenases from Nostoc sp. PCC7120 with different cleavage activities. J. Biol. Chem. 2006, 281, 31583–31593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Floss, D.S.; Schliemann, W.; Schmidt, J.; Strack, D.; Walter, M.H. RNA interference-mediated repression of MtCCD1 in mycorrhizal roots of Medicago truncatula causes accumulation of C27 apocarotenoids, shedding light on the functional role of CCD1. Plant Physiol. 2008, 148, 1267–1282. [Google Scholar] [CrossRef] [Green Version]
- Czajka, J.J.; Nathenson, J.A.; Benites, V.T.; Baidoo, E.E.K.; Cheng, Q.; Wang, Y.; Tang, Y.J. Engineering the oleaginous yeast Yarrowia lipolytica to produce the aroma compound β-ionone. Microb. Cell Fact. 2018, 17, 1–13. [Google Scholar] [CrossRef]
- Lin, J.; Massonnet, M.; Cantu, D. The genetic basis of grape and wine aroma. Hortic. Res. 2019, 6, 81. [Google Scholar] [CrossRef] [Green Version]
- Ahrazem, O.; Trapero, A.; Gomez, M.D.; Rubio-Moraga, A.; Gomez-Gomez, L. Genomic analysis and gene structure of the plant carotenoid dioxygenase 4 family: A deeper study in Crocus sativus and its allies. Genomics 2010, 96, 239–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, S.H.; Qin, X.; Loewen, M.C. The biochemical characterisation of two carotenoid cleavage enzymes from Arabidopsis indicates that a carotenoid-derived compound inhibits lateral branching. J. Biol. Chem. 2004, 279, 46940–46945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, M.H.; Floss, D.S.; Hans, J.; Fester, T.; Strack, D. Apocarotenoid biosynthesis in arbuscular mycorrhizal roots: Contributions from methylerythritol phosphate pathway isogenes and tools for its manipulation. Phytochemistry 2007, 68, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Floss, D.S.; Walter, M.H. Role of carotenoid cleavage dioxygenase 1 (CCD1) in apocarotenoid biogenesis revisited. Plant Signal. Behav. 2009, 4, 172–175. [Google Scholar] [CrossRef] [Green Version]
- Walter, M.H.; Floss, D.S.; Strack, D. Apocarotenoids: Hormones, mycorrhizal metabolites and aroma volatiles. Planta 2010, 232, 1–17. [Google Scholar] [CrossRef]
- Ohloff, G. Importance of minor components in flavors and fragrances. Perfum. Flavorist 1978, 3, 11–22. [Google Scholar]
- Kotseridis, Y.; Baumes, R.; Skouroumounis, G.K. Synthesis of labelled [2H4]β-damascenone, [2H2]2-methoxy-3-isobutylpyrazine, [2H3]α-ionone and [2H3]β-ionone, for quantification in grapes, juices and wines. J. Chromatogr. A 1998, 824, 71–78. [Google Scholar] [CrossRef]
- Baldwin, E.A.; Scott, J.W.; Shewmaker, C.K.; Schuch, W. Flavour trivia and tomato aroma: Biochemistry and possible mechanisms for control of important aroma components. HortScience 2000, 35, 1013–1021. [Google Scholar] [CrossRef] [Green Version]
- Beekwilder, J.; van Rossum, H.M.; Koopman, F.; Sonntag, F.; Buchhaupt, M.; Schrader, J.; Hall, R.D.; Bosch, D.; Pronk, J.T.; van Maris, A.J.A.; et al. Polycistronic expression of a β-carotene biosynthetic pathway in Saccharomyces cerevisiae coupled to β-ionone production. J. Biotechnol. 2014, 192, 383–392. [Google Scholar] [CrossRef]
- Mathieu, S.; Terrier, N.; Procureur, J.; Bigey, F.; Gunata, Z. A carotenoid cleavage dioxygenase from Vitis vinifera L.: Functional characterisation and expression during grape berry development in relation to C13-norisoprenoid accumulation. J. Exp. Bot. 2005, 56, 2721–2731. [Google Scholar] [CrossRef]
- Mendes-Pinto, M.M. Carotenoid breakdown products—The norisoprenoids—In wine aroma. Arch. Biochem. Biophys. 2009, 483, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, F.; Suire, C.; Mutterer, J.; Camara, B. Oxidative remodelling of chromoplast carotenoids: Identification of the carotenoid dioxygenase CsCCD and CsZCD genes involved in crocus secondary metabolite biogenesis. Plant Cell 2003, 15, 47–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eugster, C.H.; Märki-Fischer, E. The chemistry of rose pigments. Angew. Chem. 1991, 30, 654–672, (International Edition in English). [Google Scholar] [CrossRef]
- Lätari, K.; Wüst, F.; Hübner, M.; Schaub, P.; Beisel, K.G.; Matsubara, S.; Beyer, P.; Welsch, R. Tissue-specific apocarotenoid glycosylation contributes to carotenoid homeostasis in arabidopsis leaves. Plant Physiol. 2015, 168, 1550–1562. [Google Scholar] [CrossRef] [PubMed]
- Ahrazem, O.; Argandona, J.; Fiore, A.; Aguado, C.; Lujan, R.; Rubio-Moraga, A.; Marro, M.; Araujo-Andrade, C.; Loza-Alvarez, P.; Diretto, G.; et al. Transcriptome analysis in tissue sectors with contrasting crocins accumulation provides novel insights into apocarotenoid biosynthesis and regulation during chromoplast biogenesis. Sci. Rep. 2018, 8, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Pedroza, M.A.; Zalacain, A.; Lara, J.F.; Salinas, M.R. Global grape aroma potential and its individual analysis by SBSE-GC-MS. Food Res. Int. 2010, 43, 1003–1008. [Google Scholar] [CrossRef]
- Demurtas, O.C.; de Brito Francisco, R.; Martinoia, E.; Giuliano, G. Plant and Food Carotenoids: Methods and Protocols; Rodriguez-Concepcion, M., Welsch, R., Eds.; Springer Science+Business Media LLC: New York, NY, USA, 2020; Volume 2083, Chapter 7; pp. 89–99. [Google Scholar]
- Ilg, A.; Yu, Q.; Schaub, P.; Beyer, P.; Al-Babili, S. Overexpression of the rice carotenoid cleavage dioxygenase 1 gene in Golden Rice endosperm suggests apocarotenoids as substrates in planta. Planta 2010, 232, 691–699. [Google Scholar] [CrossRef]
- Dunlevy, J.D.; Kalua, C.M.; Keyzers, R.A.; Boss, P.K. Grapevine Molecular Physiology and Biotechnology, 2nd ed.; Roubelakis-Angelakis, K.A., Ed.; Springer Science+Business Media BV: Dordrecht, The Netherlands, 2009; Chapter 11; pp. 293–340. [Google Scholar]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [Green Version]
- Swiegers, J.H.; Pretorius, I.S. Yeast modulation of wine flavour. Adv. Appl. Microbiol. 2005, 57, 131–175. [Google Scholar]
- Pretorius, I.S. Tasting the terroir of wine yeast innovation. FEMS Yeast Res. 2020, 2, foz084. [Google Scholar] [CrossRef] [Green Version]
- Van Wyk, N.; Kroukamp, H.; Pretorius, I.S. The smell of synthetic biology: Engineering strategies for aroma compound production in yeast. Fermentation 2018, 4, 54. [Google Scholar] [CrossRef] [Green Version]
- Lopez, J.; Essus, K.; Kim, I.-k.; Pereira, R.; Herzog, J.; Siewers, V.; Nielsen, J.; Agosin, E. Production of β-ionone by combined expression of carotenogenic and plant CCD1 genes in Saccharomyces cerevisiae. Microb. Cell Factories 2015, 14, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolia, A.; Manzo, A.; Veronesi, F.; Rosellini, D. An overview of the last 10 years of genetically engineered crop safety research. Crit. Rev. Biotechnol. 2013, 34, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Sinemus, K.; Egelhofer, M. Transparent communication strategy on GMOs: Will it change public opinion? Biotechnol. J. 2007, 2, 1141–1146. [Google Scholar] [CrossRef]
- Petronilho, S.; Lopez, R.; Ferreira, V.; Coimbra, M.A.; Rocha, S.M. Revealing the usefulness of aroma networks to explain wine aroma properties: A case study of Portuguese wines. Molecules 2020, 25, 272. [Google Scholar] [CrossRef] [Green Version]
- Van Wyk, N.; Grossmann, M.; Wendland, J.; Von Wallbrunn, C.; Pretorius, I.S. The whiff of wine yeast innovation: Strategies for enhancing aroma production by yeast during wine fermentation. J. Agric. Chem. 2019, 67, 13496–13505. [Google Scholar] [CrossRef]
- Van Wyk, N.; Kroukamp, H.; Espinosa, M.I.; von Wallbrunn, C.; Wendland, J.; Pretorius, I.S. Blending wine yeast phenotypes with the aid of CRISPR DNA editing technologies. Int. J. Food Microbiol. 2020, 324, 108615. [Google Scholar] [CrossRef]
- Pretorius, I.S. Tailoring wine yeast for the new millennium: Novel approaches to the ancient art of winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Gene | Common Name | Cited Authors |
|---|---|---|---|
| Arabidopsis thaliana | AtCCD1 | Thale cress | [12,13,18,19,20,21,22,23,24] |
| Averrhoa carambola | AcCCD1 | Star fruit | [18,19,20] |
| Boronia megastigma | BmCCD1 | Brown boronia | [23,24,25,26] * |
| Buddleja davidii | BdCCD1 | Summer lilac | [27] |
| Citrus limon | CitCCD1 | Lemon | [12,13,19,20] |
| Coffea arabica | CaCCD1 | Coffee | [12,13,19,20,24] |
| Crocus sativus | CsCCD1 | Crocus | [12,13,18,22,23,24] |
| Cucumis melo | CmCCD1 | Melon | [12,13,19,20,21,22,24] |
| Fragaria x ananassa | FaCCD1 | Strawberry | [13,19,20,21,24] |
| Mus musculus | MmBCO2 | Mouse | [11,28] |
| Mustela putorius furo | MpCMO2/BCO2 | Ferret | [11,24] |
| Nostoc commune/spp. | NosCCD/NosNSC1 | Cyanobacterium | [24,28,29] |
| Osmanthus fragrans | OfCCD1 | Sweet olive | [13,19,20,21,23,24,25,30] |
| Petunia hybrid | PhCCD1 | Petunia | [13,18,19,20,21,22,23,24,30] |
| Phaseolus vulgaris | PvCCD1 | Common bean | [13,23] |
| Prunus persica | PpCCD1 | Nectarine | [18,22] |
| Rubus idaeus | RiCCD1 | Raspberry | [19,20,24] |
| Rosa x damascena | RdCCD1 | Damask rose | [13,19,20,21,24] |
| Scutellaria baicalensis | SbCCD1 | Skullcap | [27] |
| Solanum lycopersicum | SlCCD1 | Tomato | [12,13,18,19,20,21,22,23,24] |
| Vitis vinifera | VviCCD1 | Grape | [12,13,18,19,20,21,22,24,27,30] |
| Zea mays | ZmCCD1 | Corn | [12,13,21,24] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Timmins, J.J.B.; Kroukamp, H.; Paulsen, I.T.; Pretorius, I.S. The Sensory Significance of Apocarotenoids in Wine: Importance of Carotenoid Cleavage Dioxygenase 1 (CCD1) in the Production of β-Ionone. Molecules 2020, 25, 2779. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25122779
Timmins JJB, Kroukamp H, Paulsen IT, Pretorius IS. The Sensory Significance of Apocarotenoids in Wine: Importance of Carotenoid Cleavage Dioxygenase 1 (CCD1) in the Production of β-Ionone. Molecules. 2020; 25(12):2779. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25122779
Chicago/Turabian StyleTimmins, John J. B., Heinrich Kroukamp, Ian T. Paulsen, and Isak S. Pretorius. 2020. "The Sensory Significance of Apocarotenoids in Wine: Importance of Carotenoid Cleavage Dioxygenase 1 (CCD1) in the Production of β-Ionone" Molecules 25, no. 12: 2779. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25122779