
Plant Natural Products for the Control of Aedes aegypti: The Main Vector of Important Arboviruses

 ,
,  ,
,  and
and
Abstract
:
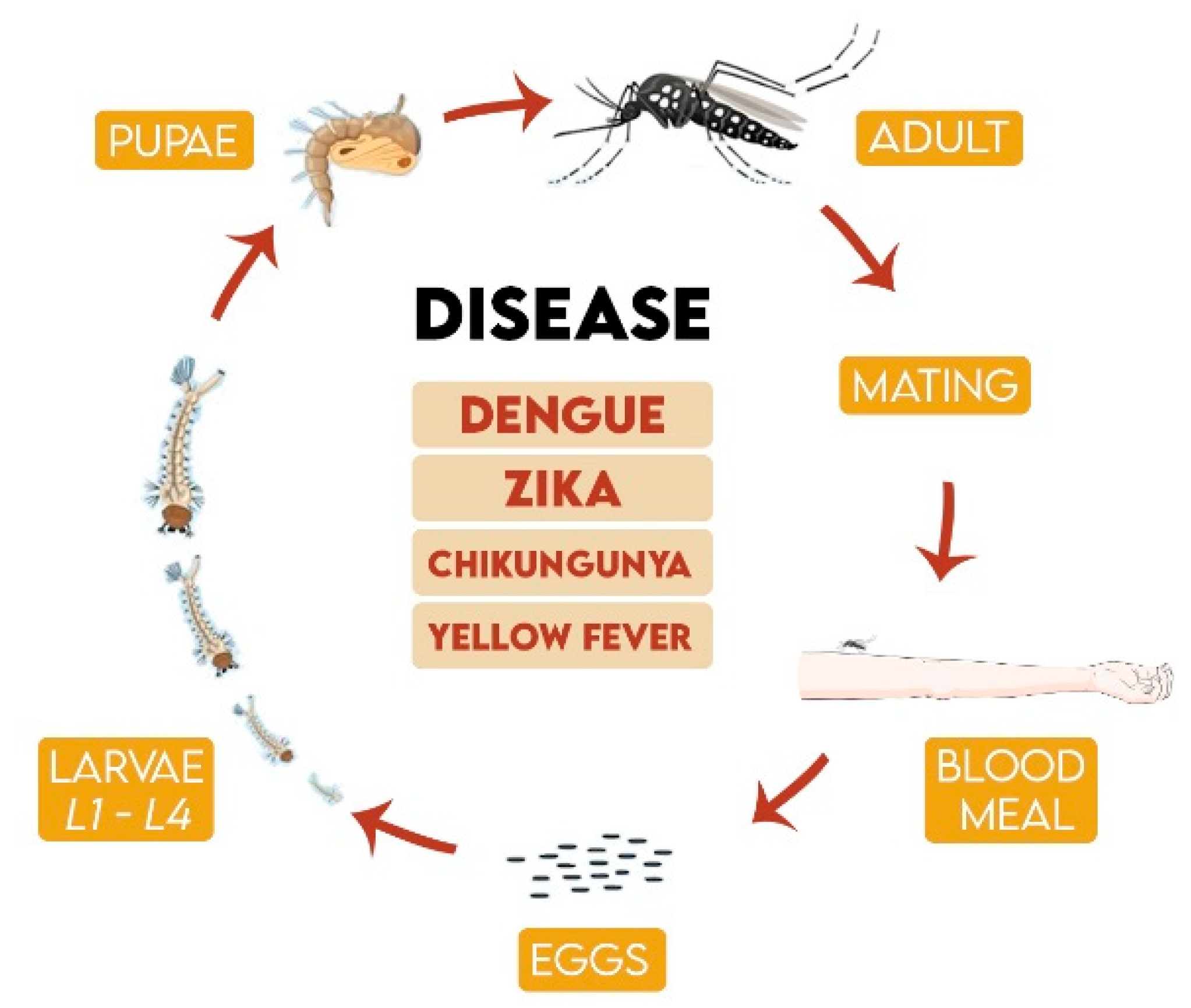
1. Introduction
2. Mosquito Control
3. Plant Natural Products to Control Mosquitoes
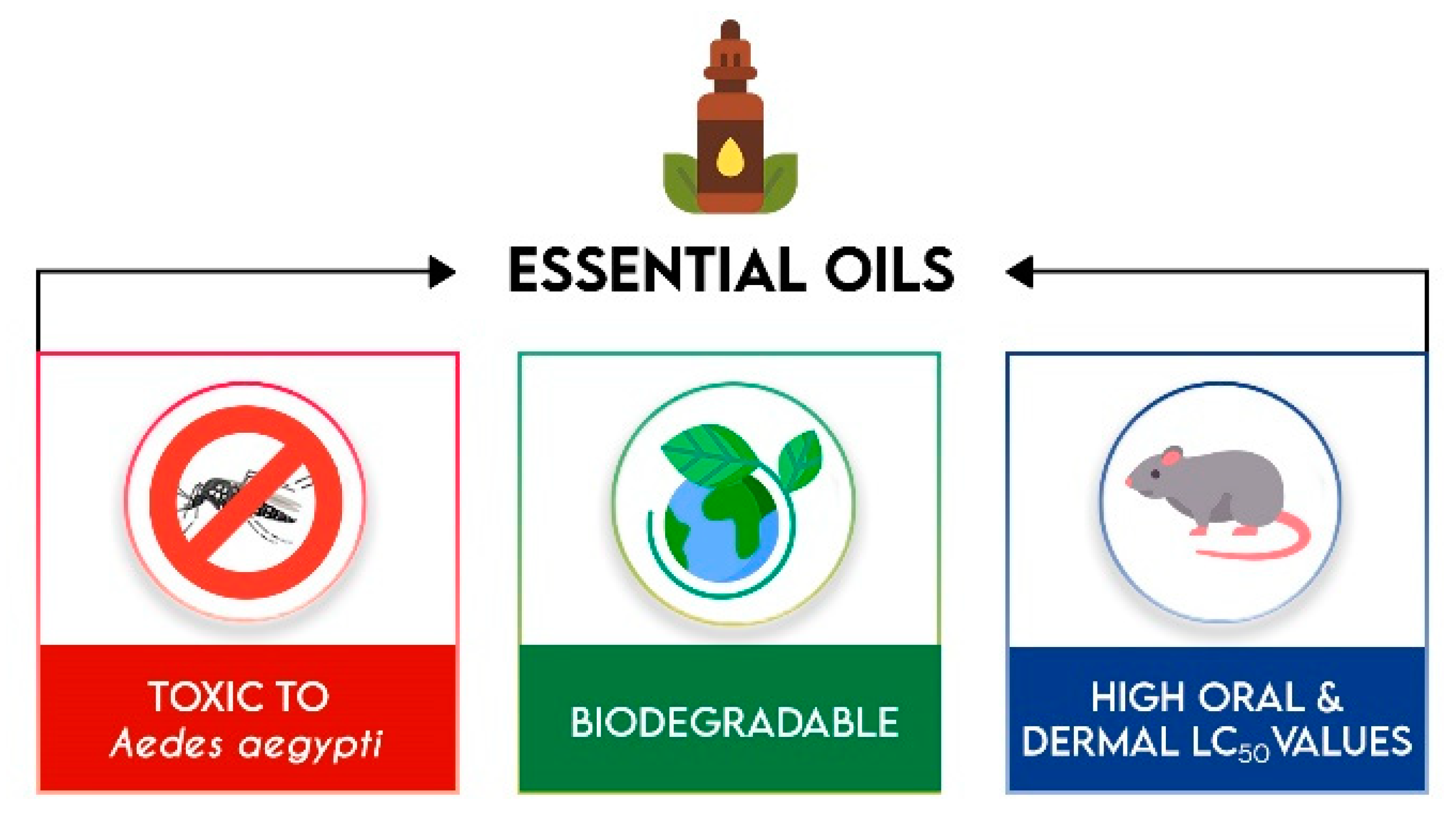
4. Essential Oils

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Family | Country | Part Used | Larval Stage | Mortality | Time of Analysis (h) | Reference | |||
|---|---|---|---|---|---|---|---|---|---|---|
| % Death | Concentration (ppm) | LC50 (ppm) | LC90 (ppm) | |||||||
| Acacia nilótica (L.) Delile | Fabaceae | India | Seeds | L4 | ND | ND | 3.17 | 11.73 | 24 | [58] |
| Alpinia purpurata (Viell.) K. Schum. | Zingiberaceae | Brazil | Red flowers | L4 | ND | ND | 80.70 | ND | 24 | [67] |
| Pink flowers | L4 | ND | ND | 71.50 | ||||||
| Baccharis reticularia DC. | Asteraceae | Brazil | Leaves | L4 | ND | ND | 221.27 | 457.47 | 24 | [68] |
| Bauhinia pulchella Benth. | Fabaceae | Brazil | Leaves | L3 | ND | ND | 105.90 | ND | 24 | [69] |
| Bauhinia ungulata L. | Fabaceae | Brazil | Leaves | L3 | ND | ND | 75.10 | ND | 24 | [69] |
| Cinnamomum osmophloeum Kaneh. | Lauraceae | Taiwan | Leaves | L4 | ND | ND | 36.0 to 177.0 | 79.0 to 296.0 | 24 | [64] |
| Croton rhamnifolioides Pax & K. Hoffm. | Euphorbiaceae | Brazil | Leaves | L4 | ND | ND | 89.0 and 122.30 | ND | 24 | [70] |
| Cunninghamia konishii Hayata | Taxodiaceae | Taiwan | Wood | L4 | ND | ND | 85.70 | 171.40 | 24 | [71] |
| Leaves | L4 | ND | ND | 91.70 | 176.50 | |||||
| Curcuma longa L. | Zingiberaceae | Thailand | Rhizome | L4 (p-s) | ND | ND | 65.51 | 110.93 | 24 | [60] |
| Eucalyptus camaldulensis Dehnh. | Myrtaceae | Taiwan | Leaves | L4 | ND | ND | 31.0 | 71.80 | 24 | [61] |
| Eucalyptus nitens (H. Deane & Maiden) Maiden | Myrtaceae | Argentina | Leaves | L3/L4 | ND | ND | 52.83 | ND | 24 | [62] |
| Eucalyptus urophylla S.T. Blake | Myrtaceae | Taiwan | Leaves | L4 | ND | ND | 95.50 | 166.30 | 24 | [61] |
| Ferula galbaniflua Boiss. & Buhse | Apiaceae | Corea | Resin | L3 | 90 | 100.0 | ND | ND | 48 | [45] |
| Foeniculum vulgare Mill. | Apiaceae | Thailand | Fruit | L4 (p-s) | ND | ND | 44.84 | 57.05 | 24 | [60] |
| Hyssopus officinalis L. | Lamiaceae | Corea | Flowers | L3 | 95 | 100.0 | ND | ND | 48 | [45] |
| Illicium verum Hook. f. | Illiciaceae | Brazil | Fruit | L3 | ND | ND | 39.80 | 53.0 | 24 | [54] |
| Larix europea Lam. & A. DC. | Pinaceae | Corea | Resin | L3 | 87 | 100.0 | ND | ND | 48 | [45] |
| Limnophila aromatica (Lamk.) Merr. | Scrophulariaceae | Thailand | Whole plant | L4 (p-s) | ND | ND | 47.94 | 65.14 | 24 | [60] |
| Mentha spicata L. | Lamiaceae | India | Leaves | L3 | ND | ND | 56.08 | 110.28 | 24 | [72] |
| Myristica fragrans Houtt. | Myristicaceae | Thailand | Flowers | L4 (p-s) | ND | ND | 47.42 | 69.28 | 24 | [60] |
| Brazil | Seeds | L3 | ND | ND | 28.20 | 41.70 | 24 | [54] | ||
| Myroxylon pereirae (Royle) Klotzsch | Fabaceae | Corea | Resin | L3 | 97.5 | 100.0 | ND | ND | 48 | [45] |
| 95 | 50.0 | ND | ND | |||||||
| Pelargonium graveolens L’Hér. ex Aiton | Geraniaceae | Corea | Leaves | L3 | 82 | 100.0 | ND | ND | 48 | [45] |
| Petroselinum crispum (Mill.) A.W. Hill | Apiaceae | Thailand | Fruit | L4 (p-s) | ND | ND | 43.22 | 66.60 | 24 | [60] |
| L4 (p-s 1) | ND | ND | 44.50 | 68.29 | 24 | |||||
| L4 (p-s 2) | ND | ND | 44.03 | 67.71 | 24 | |||||
| Pimenta dioica (L.) Merr. | Myrtaceae | Brazil | Fruit | L3 | ND | ND | 104.40 | 137.30 | 24 | [54] |
| Pinus sylvestris L. | Pinaceae | Nigeria | Needles | L4 | ND | ND | 100.39 | ND | 24 | [73] |
| Piper aduncum L. | Piperaceae | Brazil | Leaves | L3/L4 | ND | ND | 289.90 | 654.90 | 24 | [74] |
| ND | ND | 134.10 | 527.10 | 48 | ||||||
| Piper sarmentosum Roxb. ex Hunt. | Piperaceae | Thailand | Stem and Leaves | L4 (p-s) | ND | ND | 49.19 | 75.10 | 24 | [60] |
| Pogostemon patchouli Pellet. | Lamiaceae | Corea | Whole plant | L3 | 97 | 100.0 | ND | ND | 48 | [45] |
| Porophyllum ruderale (Jacq.) Cass. | Asteraceae | Brazil | Flowers and leaves | L3 | ND | ND | 60.90 | 132.48 | 24 | [75] |
| L4 | ND | ND | 72.28 | 173.65 | 24 | |||||
| Psidium guajava L. | Myrtaceae | Brazil | Leaves | L4 | ND | ND | 39.48 to 64.25 | 57.34 to 86.0 | 24 | [65] |
| Pterodon emarginatus Vogel | Fabaceae | Brazil | Fruit | L4 | ND | ND | 34.75 | ND | 48 | [66] |
| Rosmarinus officinalis L. | Lamiaceae | Brazil | Leaves | L4 | 80 | 250.0 | ND | ND | 24 | [76] |
| 90 | 250.0 | ND | ND | 48 | ||||||
| Sphaeranthus indicus L. | Asteraceae | India | Leaves | L4 | ND | ND | 140.0 | 350.0 | 24 | [77] |
| Syzygium aromaticum (L.) Merr. & L.M. Perry | Myrtaceae | Nigeria | Bud | L4 | ND | ND | 92.56 | ND | 24 | [73] |
| Tanacetum argenteum (Lam.) Willd. subsp. argenteum (Lam.) | Asteraceae | Turkey | Aerial parts | L1 | ND | ND | 93.30 | 241.70 | 24 | [78] |
| Trachyspermum ammi (L.) Sprague | Apiaceae | Corea | Seeds | L3 | 100 | 100.0 | ND | ND | 48 | [45] |
| 80 | 50.0 | ND | ND | |||||||
| Plant Species | Family | Country | Part Used | Activity | Results | Time of Analysis (h) | Reference |
|---|---|---|---|---|---|---|---|
| Acantholippia seriphioides (A. Gray) Moldenke | Verbenaceae | Argentina | ND | Repellent | 100% of repellency at 50% | 1.2 | [63] |
| Aloysia citriodora Palau | Verbenaceae | Argentina | ND | Repellent | 100% of repellency at 12.5% | 1.5 | [63] |
| Alpinia purpurata (Viell.) K. Schum. | Zingiberaceae | Brazil | Red flowers | Oviposition | Oviposition disruptive effect | ND | [67] |
| Pink flowers | Oviposition | Oviposition disruptive effect | ND | [67] | |||
| Baccharis spartioides (Hook. & Arn.) Remy | Asteraceae | Argentina | ND | Repellent | 100% of repellency at 12.5% | 1.5 | [63] |
| Croton rhamnifolioides Pax & K. Hoffm. | Euphorbiaceae | Brazil | Leaves | Oviposition | Only 30% of oviposition at 100.0 µg/mL | 16 | [70] |
| Eucalyptus nitens (H. Deane & Maiden) Maiden | Myrtaceae | Argentina | Leaves | Repellent | 100% pure repellency | 1.5 | [62] |
| Eucalyptus saligna Sm. | Myrtaceae | Argentina | ND | Repellent | 100% of repellency at 50% | 1.5 | [63] |
| Illicium verum Hook. f. | Illiciaceae | Brazil | Fruit | Adulticide | LC50 10.30 µg/mg female LC90 17.50 µg/mg female | 24 | [54] |
| Minthostachys mollis Griseb | Lamiaceae | Argentina | ND | Repellent | 100% of repellency at 50% | 1.0 | [63] |
| Myristica fragrans Houtt. | Myristicaceae | Brazil | Seeds | Adulticide | LC50 18.50 µg/mg female LC90 31.90 µg/mg female | 24 | [54] |
| Petroselinum crispum (Mill.) A.W. Hill | Apiaceae | Thailand | Fruit | Adulticide (p-s) | LC50 6.01 µg/mg female LC90 9.39 µg/mg female | 24 | [60] |
| Adulticide (p-r) | LC50 6.15 µg/mg female LC90 9.82 µg/mg female | 24 | [60] | ||||
| Pimenta dioica (L.) Merr. | Myrtaceae | Brazil | Fruit | Adulticide | LC50 16.6 µg/mg female LC90 31.4 µg/mg female | 24 | [54] |
| Pluchea carolinensis (Jack.) G. | Asteraceae | Martinica | Leaves and flowers | Repellent | 36.6% of repellency at 1.0% | ND | [79] |
| Irritant | 66.2% of irritation at 0.1% | ND | [79] | ||||
| Rosmarinus officinalis L. | Lamiaceae | Argentina | ND | Repellent | 100% of repellency at 50% | 1.5 | [63] |
| Sphaeranthus indicus L. | Asteraceae | India | Leaves | Repellent | 100% of repellency at 200 ppm | 3.5 | [77] |
| Adulticide | 100% of mortality at 800 ppm | 24 | [77] | ||||
| Tagetes minuta L. | Asteraceae | Argentina | ND | Repellent | 100% of repellency at 25% | 1.5 | [63] |
5. Organic/Aqueous Extracts
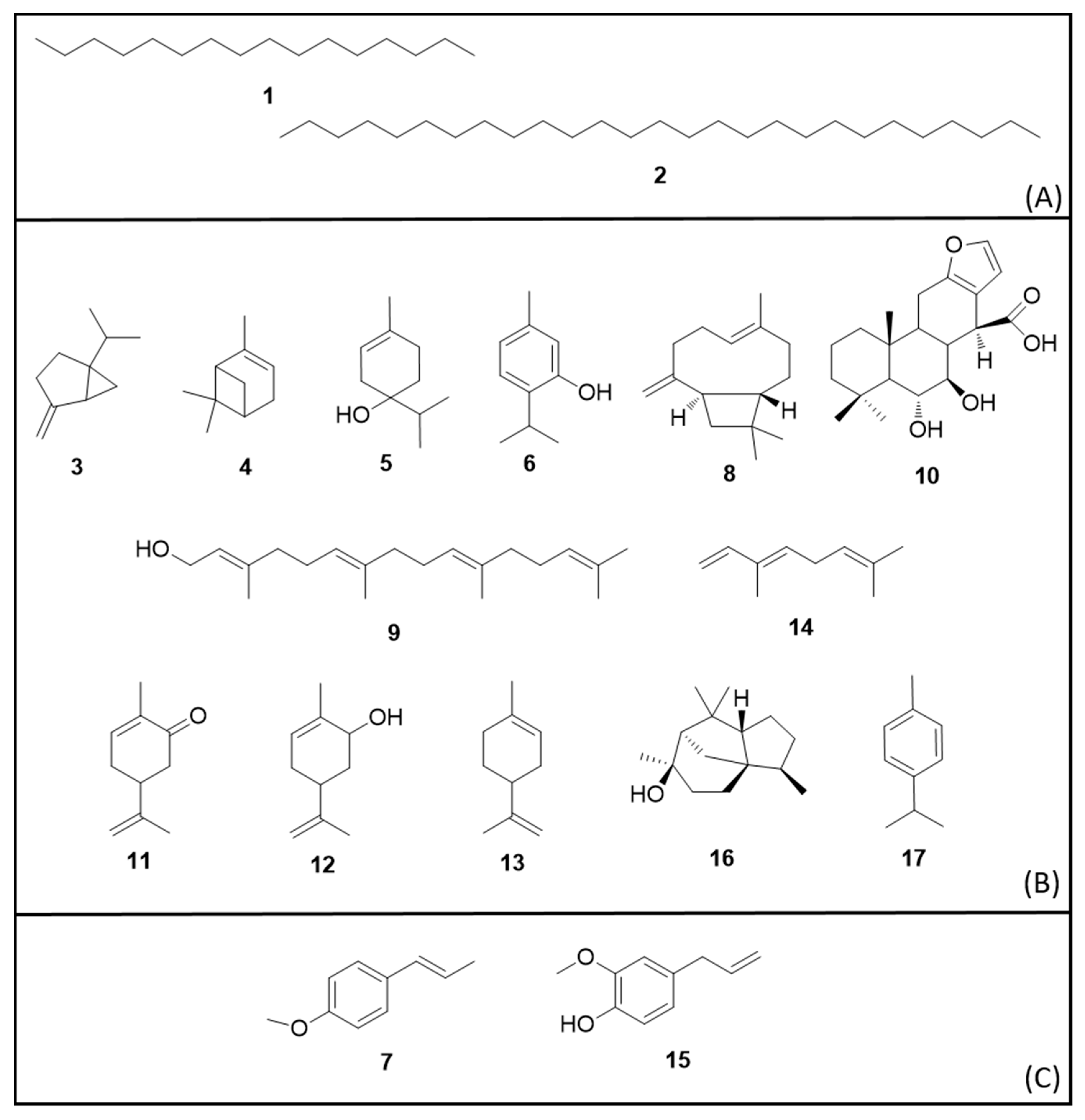
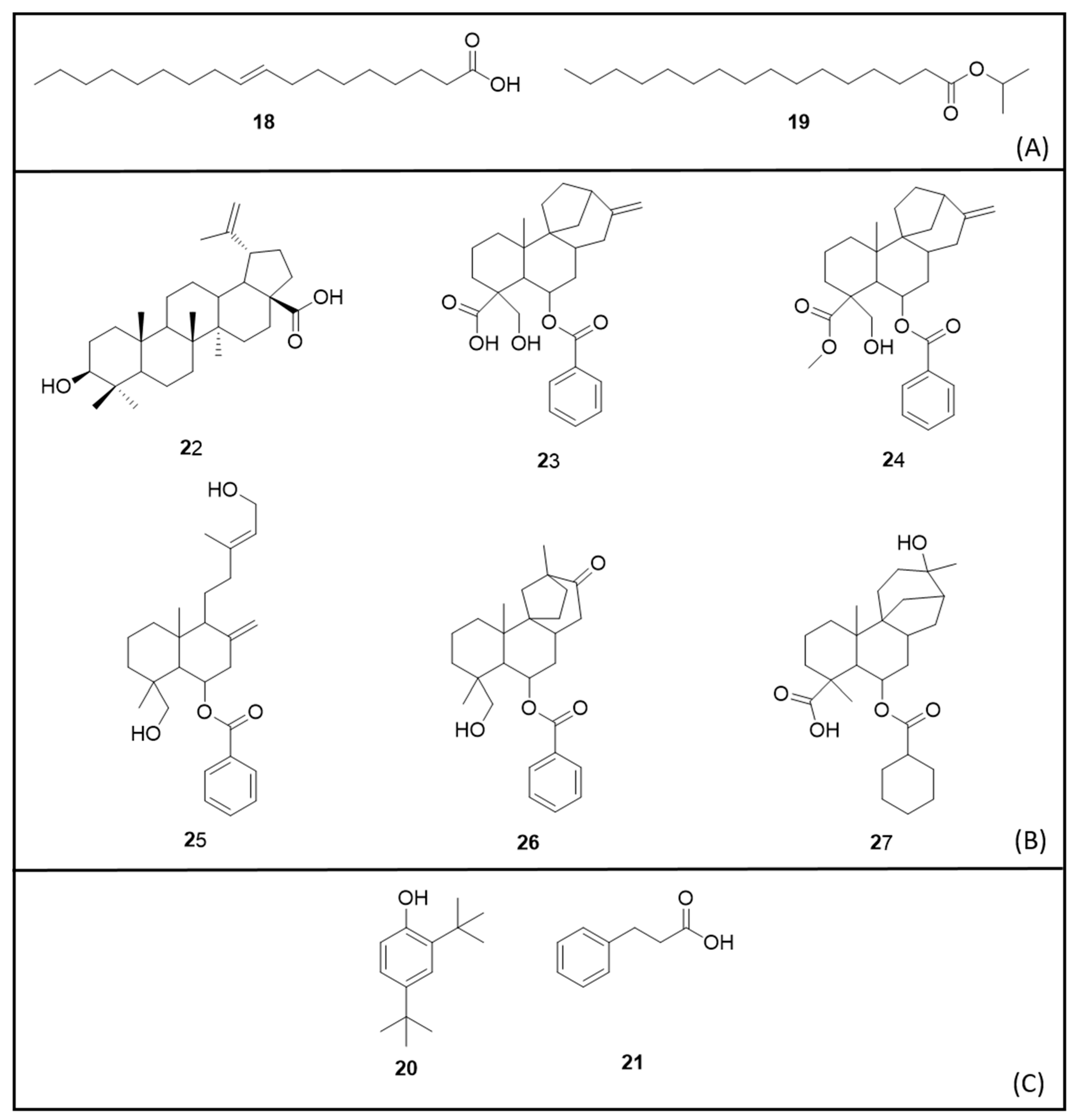
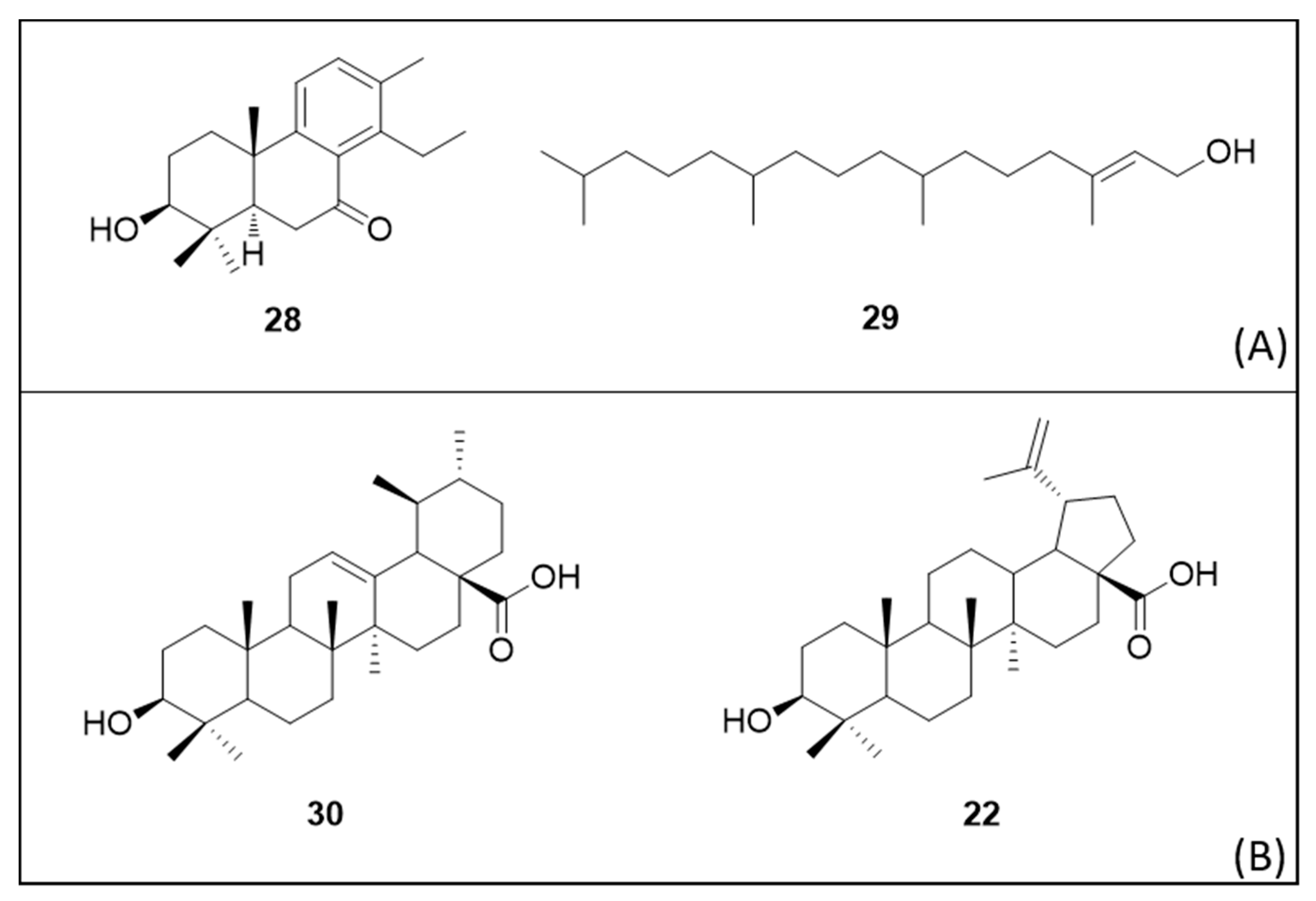
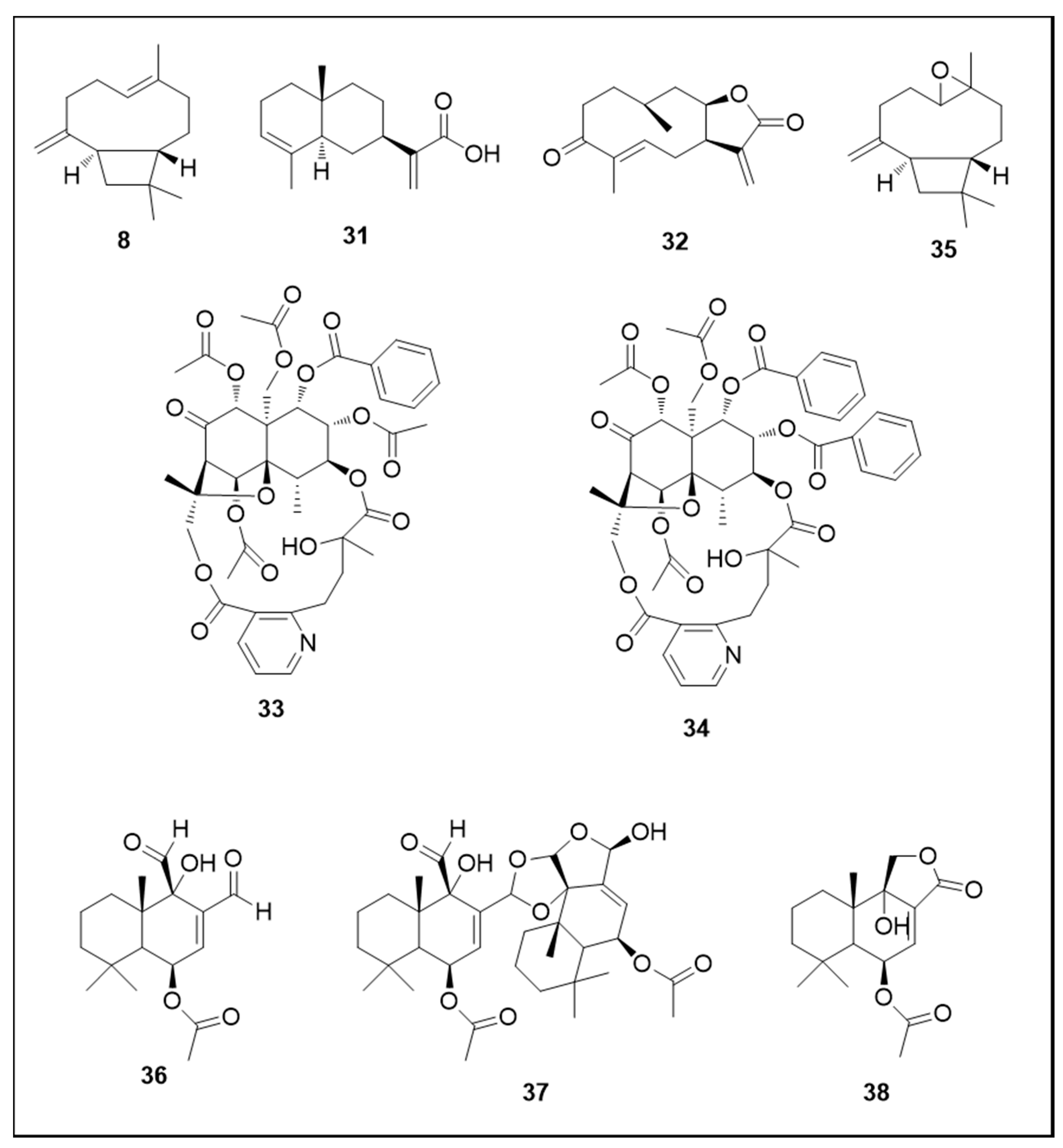
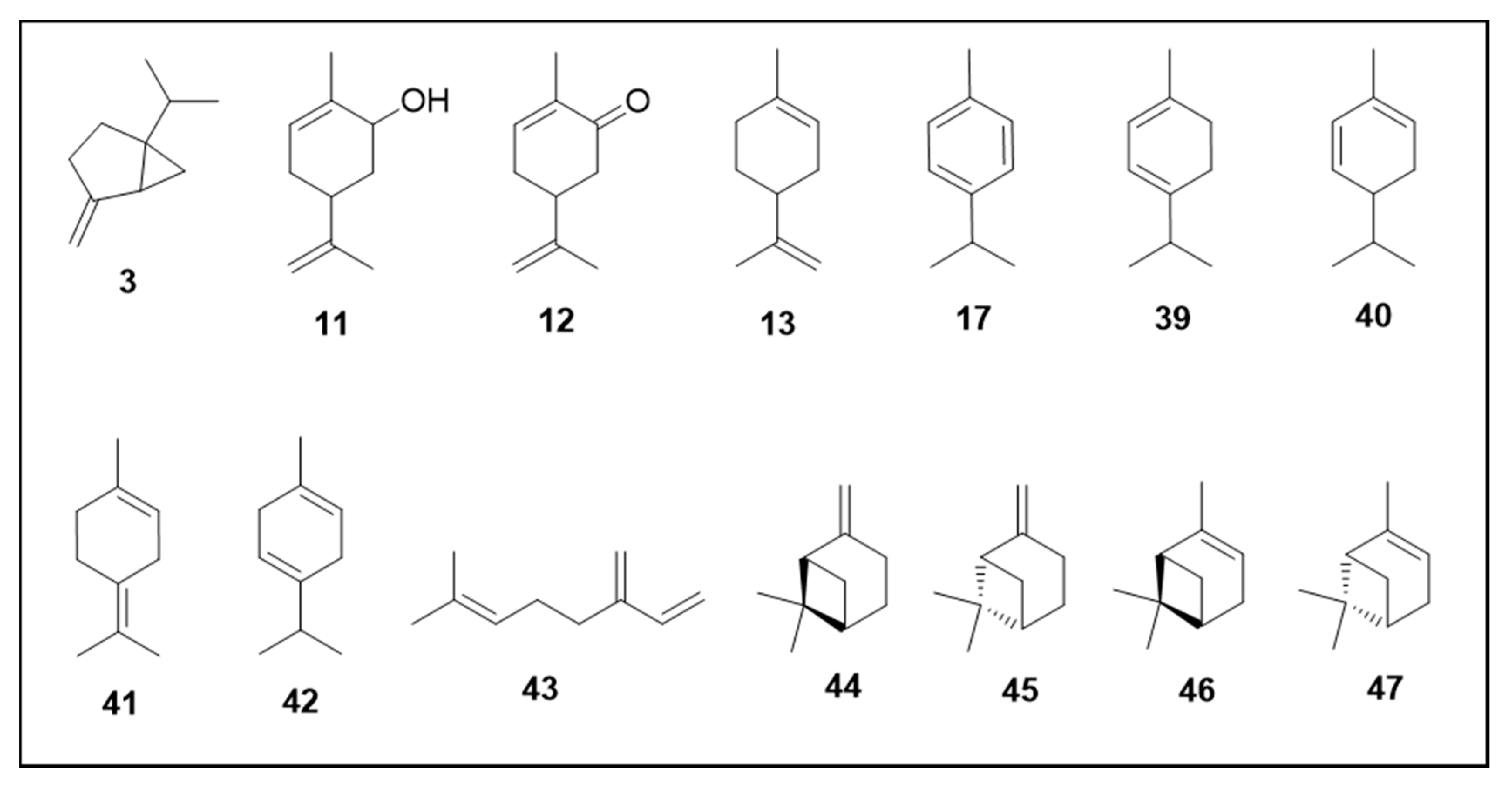
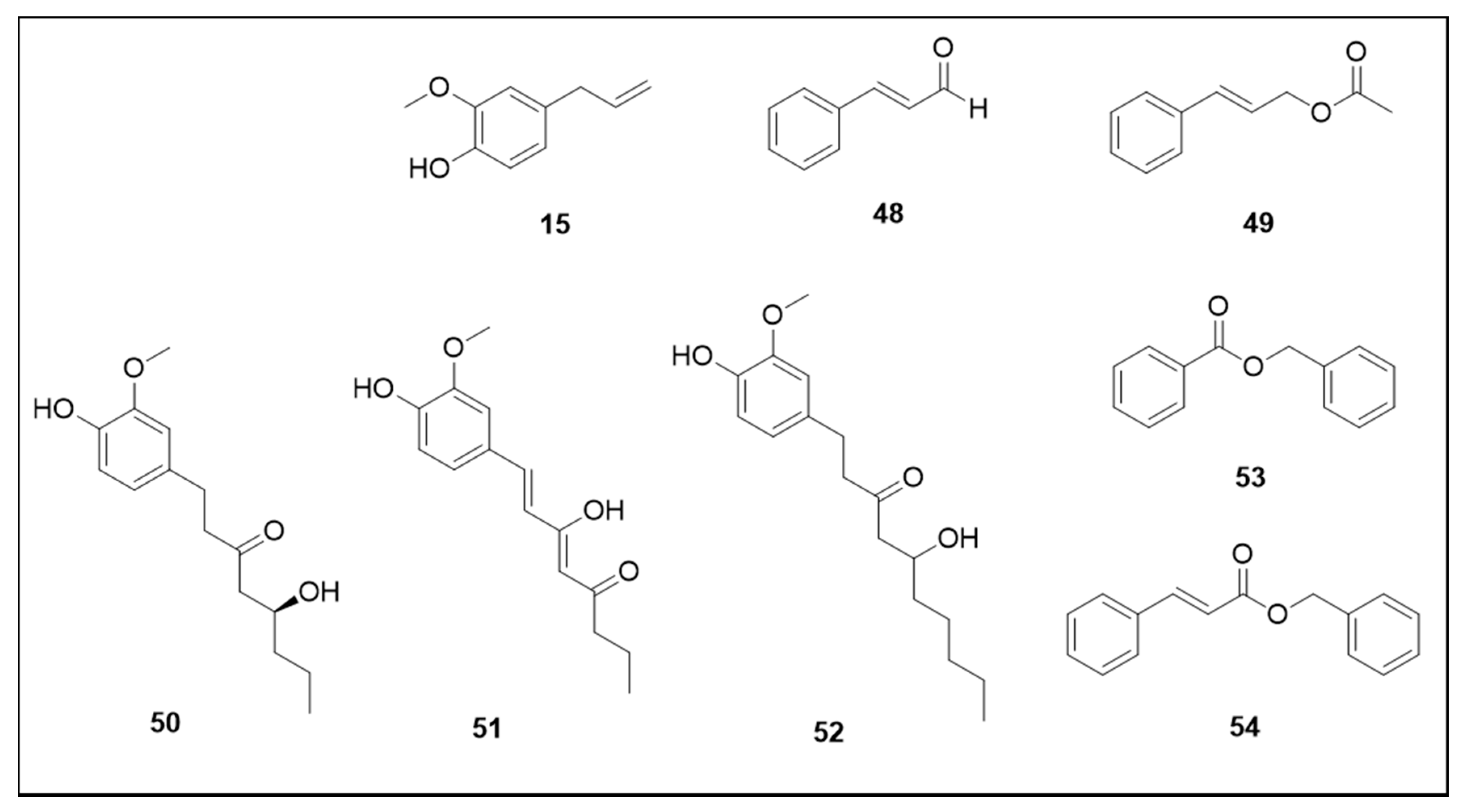
6. Secondary Metabolites
6.1. Terpenes
6.2. Phenylpropanoids and Phenolic Derivatives
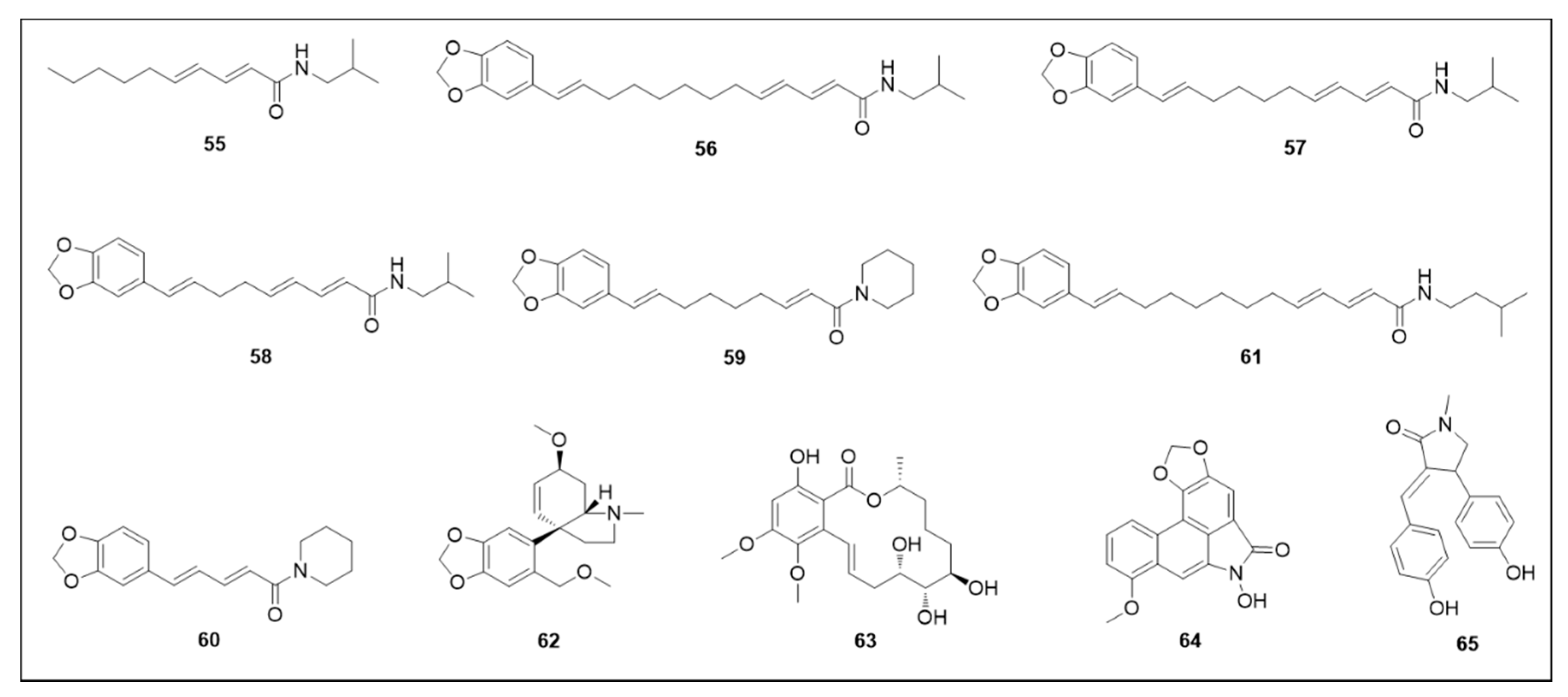
6.3. Alkaloids and Amides
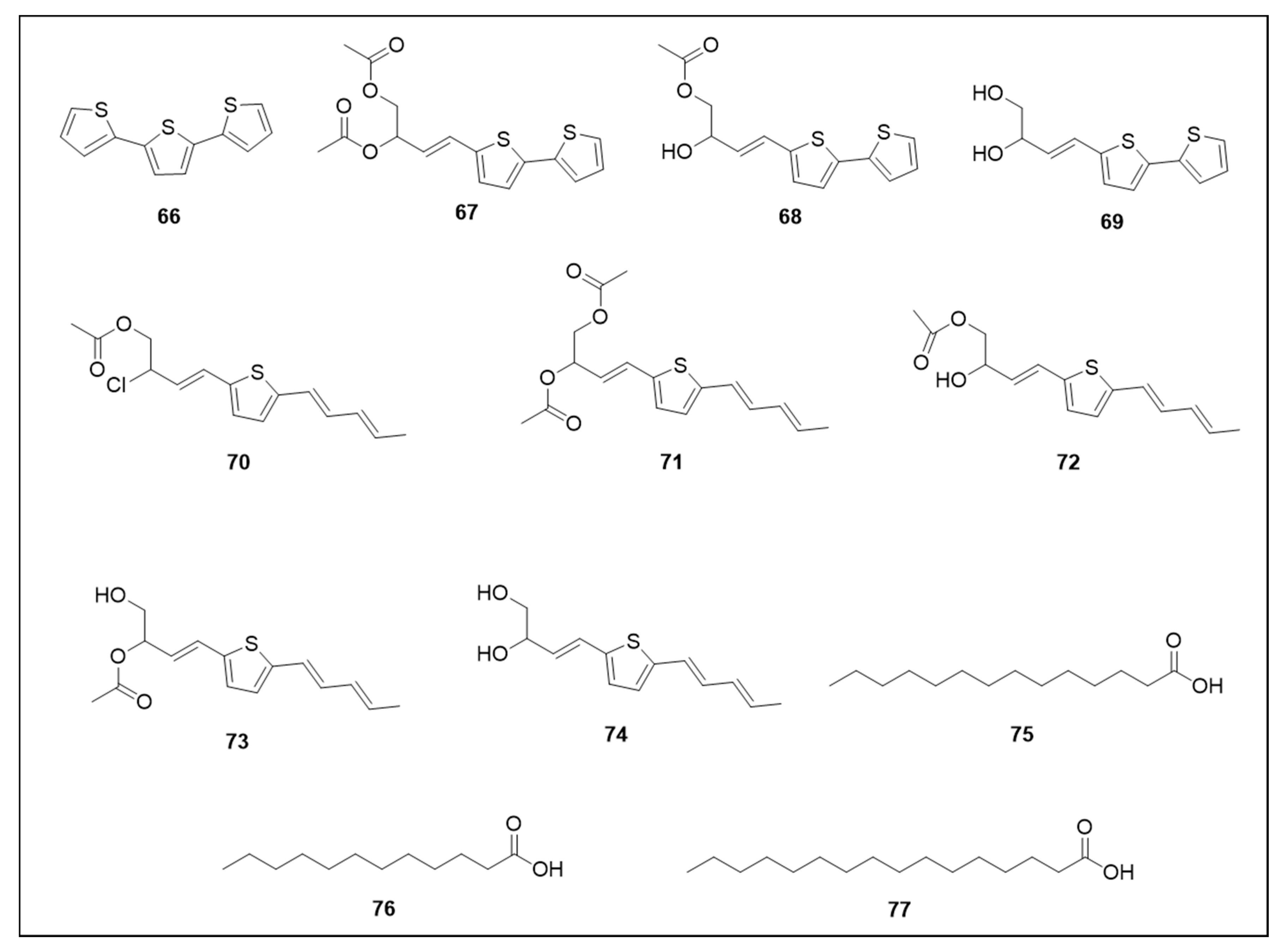
6.4. Thiophenes and Acids
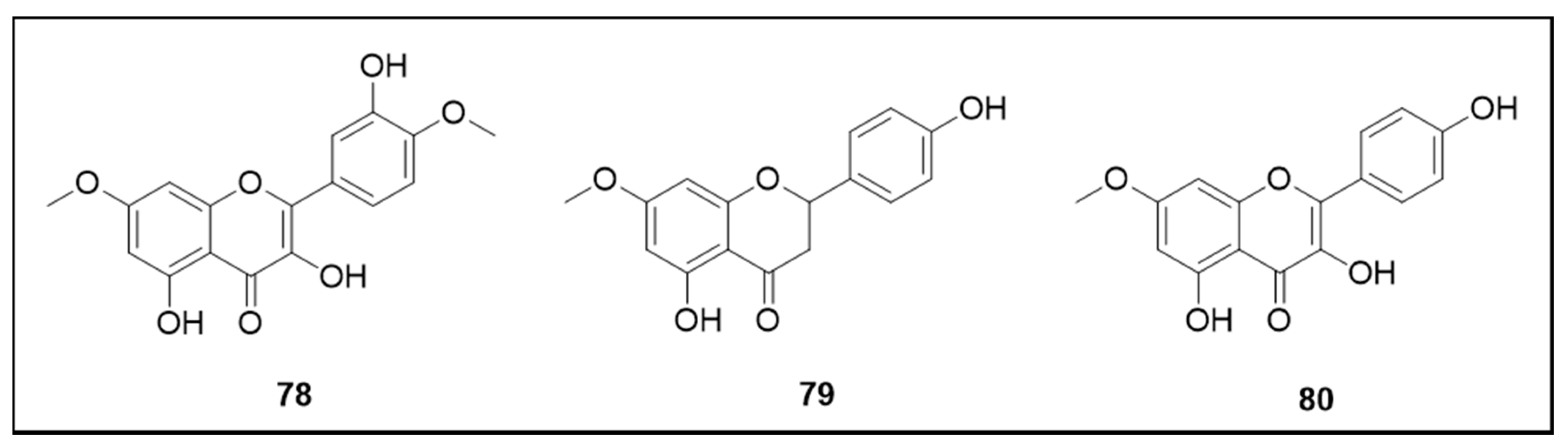
6.5. Flavonoids
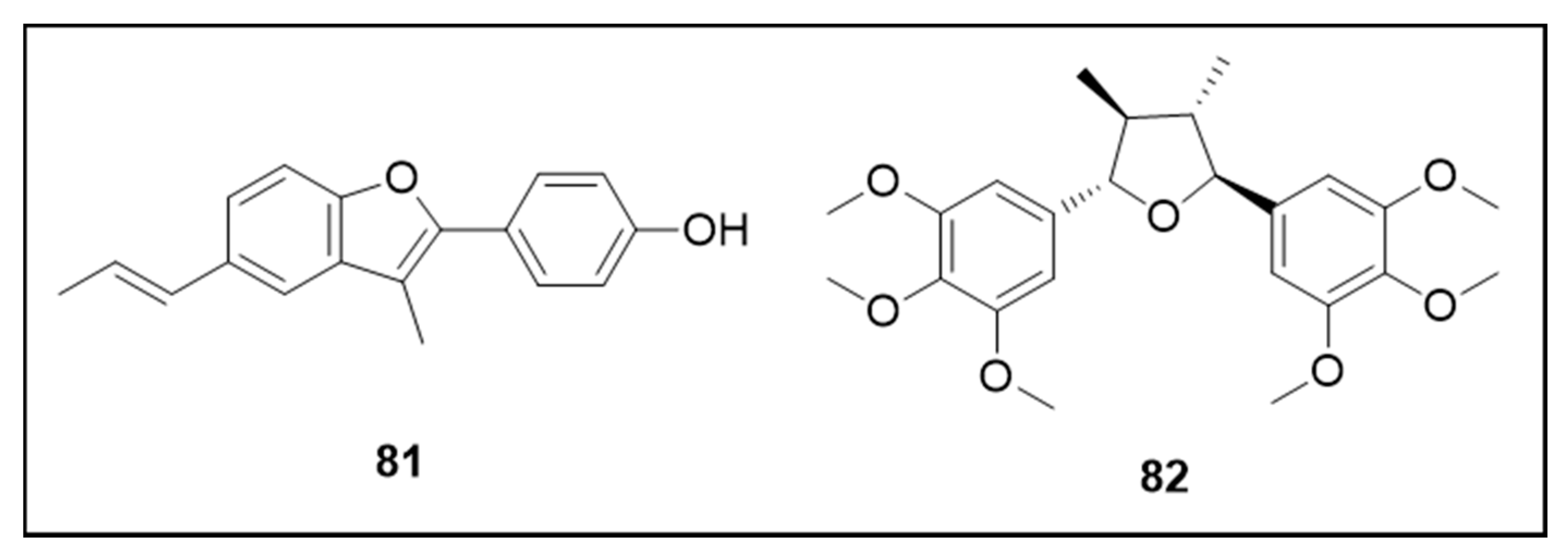
6.6. Neolignans
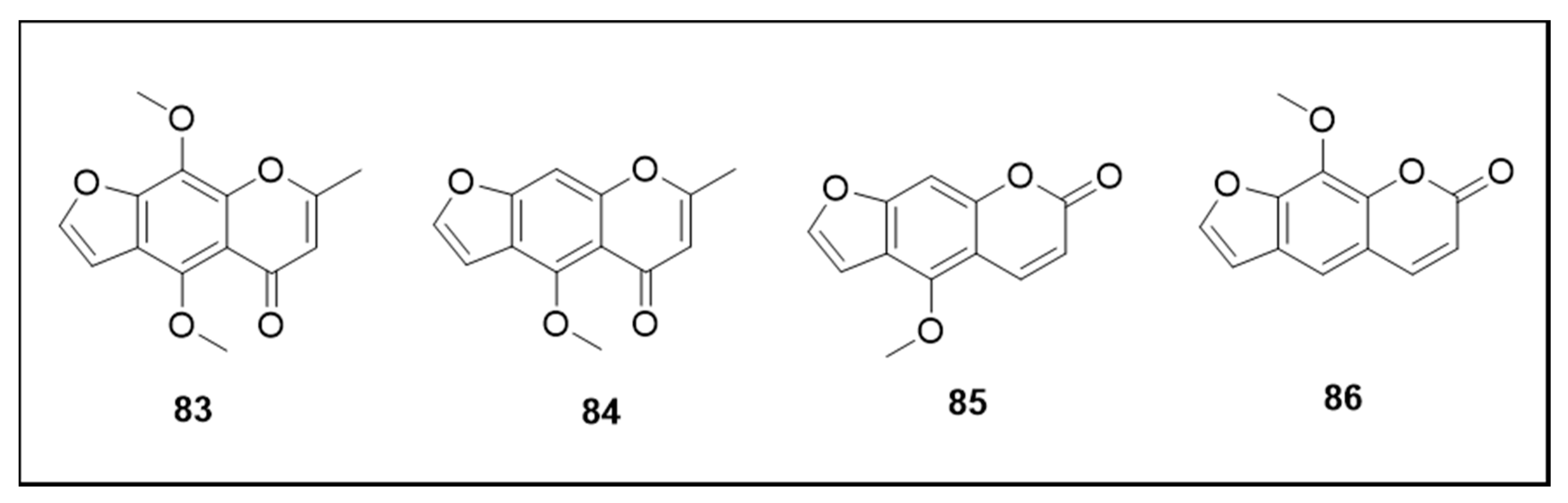
6.7. Furanochromones and Furanocoumarin
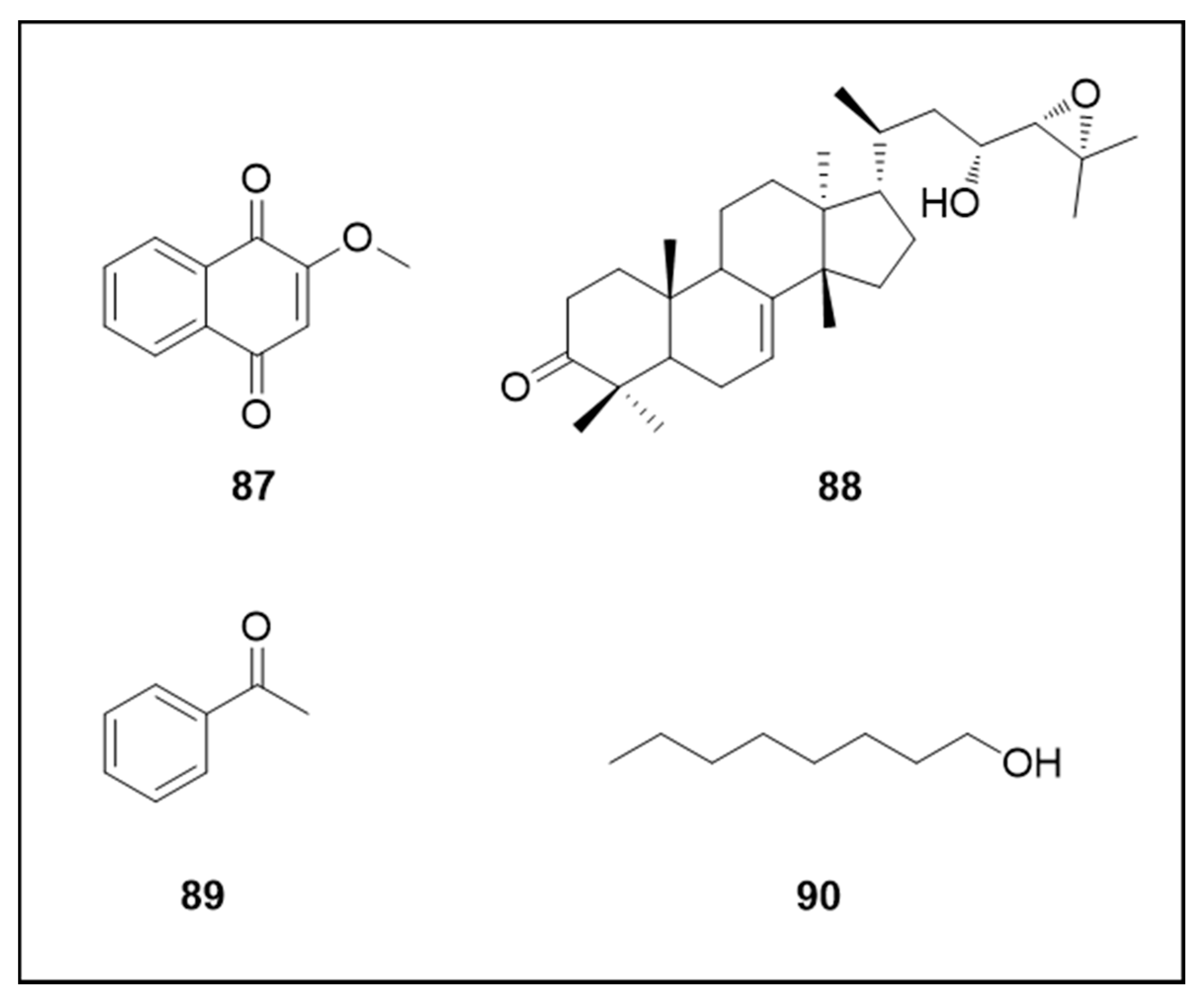
6.8. Other Secondary Metabolites
7. Mechanisms of Action
| Target Site | Mechanism of Action | Compound | Plant Species | Reference |
|---|---|---|---|---|
| Nervous system | Inhibition of acetylcholinesterase (AChE) | Essential oil | Illicium verum, Pimenta dioica and Myristica fragrans | [54] |
| Nanoemulsion with essential oil | Pterodon emarginatus | [66] | ||
| Not specified | Ethanolic extract | Piper longum, Piper ribesoides and Piper sarmentosum | [109] | |
| Gut trypsin | Inhibition of trypsin and consequent decreased absorption of nutrientes and essential aminoacids | Aqueous extract | Moringa oleifera | [147] |
| Essential oil | Croton rhamnifolioides | [70] | ||
| Peritrophic matrix | Change in internal morphology and consequent insect protection dysfuntion | Methanol extracts | Derris (Lonchocarpus) urucu | [100] |
| Nilocetin | Limonia acidissima | [99] | ||
| Midgut epithelium | Tissue destruction and cell disorganization | Methanol extracts | Derris (Lonchocarpus) urucu | [100] |
| Pellitorine | Asarum heterotropoides | [148] | ||
| Grandisin | Piper solmsianum | [139] | ||
| Anal papillae | Morphological changes, interference with the larva swin | Crude extracts | Dalbergia brasiliensis | [89] |
| Ethanolic extract | Piper longum, Piper ribesoides and Piper sarmentosum | [109] | ||
| Respiratory siphon | Morphological changes, interference with the oxygen flow | Crude extracts | Dalbergia brasiliensis | [89] |
| Nanoemulsion with limonene | Baccharis reticularia | [68] | ||
| Anal gills | Comprehensive damage; debris in hemolymph | Pellitorine | Asarum heterotropoides | [148] |
| Thorax and exoskeleton | Changes in external morphology, interfering with the molting process | Crude extracts | Dalbergia brasiliensis | [89] |
| Pellitorine | Asarum heterotropoides | [148] | ||
| Nilocetin | Limonia acidissima | [99] | ||
| Nanoemulsion with limonene | Baccharis reticularia | [68] | ||
| Digestive system | Digestive toxicity | Crude extracts | Dalbergia brasiliensis | [89] |
8. Limitations and/or Expectations of Plant Natural Product Insecticide Applications
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Consoli, R.A.G.B.; Oliveira, R.L. Principais Mosquitos de Importância Sanitária no Brasil, 1st ed.; Fiocruz: Rio de Janeiro, Brazil, 1994; pp. 115–117. [Google Scholar]
- Kraemer, M.U.F.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, 08347. [Google Scholar] [CrossRef]
- Geris, R.; Ribeiro, P.R.; Brandão, M.D.S.; Da Silva, H.H.G.; Da Silva, I.G. Bioactive Natural Products as Potential Candidates to Control Aedes aegypti, the Vector of Dengue. In Bioactive Natural Products; Elsevier BV: Amsterdam, The Netherlands, 2012; Volume 37, pp. 277–376. [Google Scholar]
- Benelli, G.; Jeffries, C.L.; Walker, T. Biological Control of Mosquito Vectors: Past, Present, and Future. Insects 2016, 7, 52. [Google Scholar] [CrossRef]
- World Health Organization. Global Strategy for Dengue Prevention and Control. Available online: https://apps.who.int/iris/bitstream/handle/10665/75303/9789241504034_eng.pdf;jsessionid=59CD5C25000FE7F9ABE29B06C33D6461?sequence=1 (accessed on 27 November 2019).
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Dengue Guidelines for Diagnosis, Treatment, Prevention and Control. Available online: https://apps.who.int/iris/bitstream/handle/10665/44188/9789241547871_eng.pdf?sequence=1&isAllowed=y (accessed on 2 December 2019).
- World Health Organization. Zika Virus: An Epidemiological Update. Available online: https://apps.who.int/iris/bitstream/handle/10665/255008/WER9215.pdf?sequence=1 (accessed on 5 December 2019).
- World Health Organization. Chikungunya Disease: Gaps and Opportunities in Public Health and Research in the Americas. Available online: https://www.who.int/wer/2015/wer9042.pdf?ua=1 (accessed on 9 December 2019).
- Shepard, D.S.; Undurraga, E.A.; Halasa, Y.A.; Stanaway, J.D. The global economic burden of dengue: A systematic analysis. Lancet Infect. Dis. 2016, 16, 935–941. [Google Scholar] [CrossRef]
- Teich, V.; Arinelli, R.; Fahham, L. Aedes aegypti e sociedade: O impacto econômico das arboviroses no Brasil. J. Bras. Econ. 2017, 9, 267–276. [Google Scholar] [CrossRef]
- Messina, J.; Brady, O.J.; Golding, N.; Kraemer, M.U.F.; Wint, G.R.W.; Ray, S.E.; Pigott, D.M.; Shearer, F.M.; Johnson, K.; Earl, L.; et al. The current and future global distribution and population at risk of dengue. Nat. Microbiol. 2019, 4, 1508–1515. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G. Research in mosquito control: Current challenges for a brighter future. Parasitol. Res. 2015, 114, 2801–2805. [Google Scholar] [CrossRef]
- Isman, M.B. A renaissance for botanical insecticides? Pest Manag. Sci. 2015, 71, 1587–1590. [Google Scholar] [CrossRef]
- Lees, R.; Gilles, J.R.; Hendrichs, J.; Vreysen, M.J.B.; Bourtzis, K. Back to the future: The sterile insect technique against mosquito disease vectors. Curr. Opin. Insect Sci. 2015, 10, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Alphey, L.; McKemey, A.; Nimmo, D.; Oviedo, M.N.; Lacroix, R.; Matzen, K.; Beech, C. Genetic control of Aedes mosquitoes. Pathog. Glob. Heal. 2013, 107, 170–179. [Google Scholar] [CrossRef] [Green Version]
- Evans, H.C.; Elliot, S.L.; Barreto, R.W. Entomopathogenic fungi and their potential for the management of Aedes aegypti (Diptera: Culicidae) in the Americas. Mem. Inst. Oswaldo Cruz 2018, 113, 206–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durovni, B.; Saraceni, V.; Eppinghaus, A.; Riback, T.I.S.; Neill, S.L.O.; Moreira, L.A.; Jewell, N.P.; Dufault, S.M.; Simmons, C.P.; Tanamas, S.K.; et al. The impact of large-scale deployment of Wolbachia mosquitoes on arboviral disease incidence in Rio de Janeiro and Niterói, Brazil: Study protocol for a controlled interrupted time series analysis using routine disease surveillance data. F1000 Res. 2019, 8, 1328. [Google Scholar] [CrossRef] [Green Version]
- Sarwar, M. Control of dengue carrier Aedes mosquitoes (Diptera: Culicidae) larvae by larvivorous fishes and putting it into practice within water bodies. Int. J. Prev. Med. Res. 2015, 1, 232–237. [Google Scholar]
- Cavalcanti, L.P.D.G.; Pontes, R.J.S.; Regazzi, A.C.F.; de Paula Júnior, F.J.; Frutuoso, R.L.; Sousa, E.P.; Dantas Filho, F.D.; Lima, J.W.D.O. Efficacy of fish as predators of Aedes aegypti larvae, under laboratory conditions. Rev. Saude Pública 2007, 41, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Nam, V.S.; Yen, N.T.; Duc, H.M.; Tu, T.C.; Thang, V.T.; Le, N.H.; San, L.H.; Loan, L.L.; Huong, V.T.Q.; Khanh, L.H.K.; et al. Community-based control of Aedes aegypti by using Mesocyclops in southern Vietnam. Am. J. Trop. Med. Hyg. 2012, 86, 850–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Estrada, J.L.; Rodríguez, M.H.; Cruz-López, L.; Arredondo-Jimenez, J.I. Selective oviposition by Aedes aegypti (Diptera: Culicidae) in response to Mesocyclops longisetus (Copepoda: Cyclopoidea) under laboratory and field conditions. J. Med. Entomol. 2001, 38, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Albeny, D.S.; Martins, G.F.; Andrade, M.R.; Krüger, R.F.; Vilela, E.F. Aedes aegypti survival in the presence of Toxorhynchites violaceus (Diptera: Culicidae) fourth instar larvae. Zoologia 2011, 28, 538–540. [Google Scholar] [CrossRef] [Green Version]
- Zuharah, W.F.; Fadzly, N.; Yusof, N.A.; Dieng, H.; Ni, X. Risky behaviors: Effects of Toxorhynchites splendens (Diptera: Culicidae) predator on the behavior of three mosquito species. J. Insect Sci. 2015, 15, 128. [Google Scholar] [CrossRef] [Green Version]
- Benelli, G. Plant-borne ovicides in the fight against mosquito vectors of medical and veterinary importance: A systematic review. Parasitol. Res. 2015, 114, 3201–3212. [Google Scholar] [CrossRef]
- Pavela, R. Essential oils for the development of eco-friendly mosquito larvicides: A review. Ind. Crops Prod. 2015, 76, 174–187. [Google Scholar] [CrossRef]
- Lughadha, E.N.; Govaerts, R.; Belyaeva, I.; Black, N.; Lindon, H.; Allkin, R.; Magill, R.E.; Nicolson, N. Counting counts: Revised estimates of numbers of accepted species of flowering plants, seed plants, vascular plants and land plants with a review of other recent estimates. Phytotaxa 2016, 272, 82–88. [Google Scholar] [CrossRef] [Green Version]
- Zebitz, C.P.W. Effects of three different neem seed kernel extracts and azadirachtin on larvae of different mosquito species. J. Appl. Entomol. 1986, 102, 455–463. [Google Scholar] [CrossRef]
- Sharma, R.N.; Bhosale, A.S.; Joshi, V.N.; Hebbalkar, D.S.; Tungikar, V.B.; Gupta, A.S.; Patwardhan, S.A. Lavandula gibsonii: A plant with insectistatic potential. Phytoparasitica 1981, 9, 101–109. [Google Scholar] [CrossRef]
- Pavela, R. History, presence and perspective of using plant extracts as commercial botanical insecticides and farm products for protection against Insects—A review. Plant Prot. Sci. 2016, 52, 229–241. [Google Scholar]
- Yanola, J.; Somboon, P.; Walton, C.; Nachaiwieng, W.; Prapanthadara, L.-a. A novel F1552/C1552 point mutation in the Aedes aegypti voltage-gated sodium channel gene associated with permethrin resistance. Pestic. Biochem. Physiol. 2010, 96, 127–131. [Google Scholar] [CrossRef]
- Saavedra-Rodriguez, K.; Urdaneta-Marquez, L.; Rajatileka, S.; Moulton, M.; Flores, A.E.; Fernandez-Salas, I.; Bisset, J.; Rodriguez, M.; Mccall, P.J.; Donnelly, M.J.; et al. A mutation in the voltage-gated sodium channel gene associated with pyrethroid resistance in Latin American Aedes aegypti. Insect Mol. Biol. 2007, 16, 785–798. [Google Scholar] [CrossRef] [Green Version]
- Fundação Osvaldo Cruz. Recomendação Técnica Sobre a Interrupção do Uso de Inseticidas Piretróides no Controle do Aedes aegypti no Brasil. Available online: http://www.fiocruz.br/ioc/media/NotaTecnica%202%20IOC%20v1%20dengue%206%20set%202011.pdf (accessed on 18 December 2019).
- Hemingway, J.; Ranson, H. Insecticide resistance in insect vectors of human diseases. Annu. Rev. Entomol. 2000, 45, 371–391. [Google Scholar] [CrossRef]
- Priester, T.M.; Georghiou, G.P. Penetration of permethrin and knockdown in larvae of pyrethroid-resistant and -susceptible strains of the southern house mosquito. J. Econ. Entomol. 1980, 73, 165–167. [Google Scholar] [CrossRef]
- Raymond, M.; Chevillon, C.; Guillemaud, T.; Lenormand, T.; Pasteur, N. An overview of the evolution of overproduced esterases in the mosquito Culex pipiens. Philos. Trans. R. Soc. Lond. B 1998, 353, 1707–1711. [Google Scholar] [CrossRef] [Green Version]
- Cariño, F.A.; Koener, J.F.; Plapp, F.W.; Feyereisen, R. Constitutive overexpression of the cytochrome P450 gene CYP6A1 in a house fly strain with metabolic resistance to insecticides. Insect Biochem. Mol. Biol. 1994, 24, 411–418. [Google Scholar] [CrossRef]
- Mutero, A.; Pralavorio, M.; Bride, J.M.; Fournier, D. Resistance-associated point mutations in insecticide-insensitive acetylcholinesterase. Proc. Natl. Acad. Sci. USA 1994, 91, 5922–5926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisset, J.; Rodríguez, M.M.; Fernández, D. Selection of insensitive acetylcholinesterase as a resistance mechanism in Aedes aegypti (Diptera: Culicidae) from Santiago de Cuba. J. Med. Entomol. 2006, 43, 1185–1189. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Li, M.X.; Chang, H.J.; Mao, Y.; Zhang, H.Y.; Lu, L.X.; Yan, S.G.; Lang, M.L.; Liu, L.; Qiao, C.L. Carboxylesterase-mediated insecticide resistance: Quantitative increase induces broader metabolic resistance than qualitative change. Pestic. Biochem. Physiol. 2015, 121, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Anthony, N.; Rocheleau, T.; Mocelin, G.; Lee, H.J.; Ffrench-Constant, R. Cloning, sequencing and functional expression of an acetylcholinesterase gene from the yellow fever mosquito Aedes aegypti. FEBS Lett. 1995, 368, 461–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacorte, S.; Ehresmann, N.; Barceló, D. Persistence of temephos and its transformation products in rice crop field waters. Environ. Sci. Technol. 1996, 30, 917–923. [Google Scholar] [CrossRef]
- Zhang, A.; Xie, X.; Ye, J.; Lin, C.; Hu, X. Stereoselective toxicity of malathion and its metabolites, malaoxon and isomalathion. Environ. Chem. Lett. 2011, 9, 369–373. [Google Scholar] [CrossRef]
- Park, H.-M.; Kim, J.; Chang, K.-S.; Kim, B.-S.; Yang, Y.-J.; Kim, G.-H.; Shin, S.-C.; Park, I.-K. Larvicidal activity of Myrtaceae essential oils and their components against Aedes aegypti, acute toxicity on Daphnia magna, and aqueous residue. J. Med. Entomol. 2011, 48, 405–410. [Google Scholar] [CrossRef]
- Seo, S.M.; Park, H.M.; Park, I.K. Larvicidal activity of ajowan (Trachyspermum ammi) and Peru balsam (Myroxylon pereira) oils and blends of their constituents against mosquito, Aedes aegypti, acute toxicity on water flea, Daphnia magna, and aqueous residue. J. Agric. Food Chem. 2012, 60, 5909–5914. [Google Scholar] [CrossRef]
- Williams, D.H.; Stone, M.J.; Hauck, P.R.; Rahman, S.K. Why are secondary metabolites (natural products) biosynthesized. J. Nat. Prod. 1989, 52, 1189–1208. [Google Scholar] [CrossRef]
- Isman, M.B. Bridging the gap: Moving botanical insecticides from the laboratory to the farm. Ind. Crops Prod. 2017, 110, 10–14. [Google Scholar] [CrossRef]
- Miresmailli, S.; Isman, M.B. Botanical insecticides inspired by plant–herbivore chemical interactions. Trends Plant Sci. 2014, 19, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B.; Miresmailli, S.; MacHial, C. Commercial opportunities for pesticides based on plant essential oils in agriculture, industry and consumer products. Phytochem. Rev. 2011, 10, 197–204. [Google Scholar] [CrossRef]
- Isman, M.B. Plant essential oils for pest and disease management. Crop Prot. 2000, 19, 603–608. [Google Scholar] [CrossRef]
- Pavela, R. Encapsulation—A convenient way to extend the persistence of the effect of eco-friendly mosquito larvicides. Curr. Org. Chem. 2016, 20, 2674–2680. [Google Scholar] [CrossRef] [Green Version]
- Benelli, G. Plant-mediated biosynthesis of nanoparticles as an emerging tool against mosquitoes of medical and veterinary importance: A review. Parasitol. Res. 2016, 115, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Ashokan, A.P.; Paulpandi, M.; Dinesh, D.; Murugan, K.; Vadivalagan, C.; Benelli, G. Toxicity on Dengue mosquito vectors through Myristica fragrans—synthesized zinc oxide nanorods, and their cytotoxic effects on liver cancer cells (HepG2). J. Clust. Sci. 2017, 28, 205–226. [Google Scholar] [CrossRef]
- Gomes da Rocha Voris, D.; Dos Santos Dias, L.; Alencar Lima, J.; Dos Santos Cople Lima, K.; Pereira Lima, J.B.; Dos Santos Lima, A.L. Evaluation of larvicidal, adulticidal, and anticholinesterase activities of essential oils of Illicium verum Hook. f., Pimenta dioica (L.) Merr., and Myristica fragrans Houtt. against Zika virus vectors. Environ. Sci. Pollut. Res. 2018, 25, 22541–22551. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Chang, H.; Chang, S. Bioactivity of selected plant essential oils against the yellow fever mosquito Aedes aegypti larvae. Bioresour. Technol. 2003, 89, 99–102. [Google Scholar] [CrossRef]
- Kiran, S.R.; Bhavani, K.; Devi, P.S.; Rao, B.R.R.; Reddy, K.J. Composition and larvicidal activity of leaves and stem essential oils of Chloroxylon swietenia DC against Aedes aegypti and Anopheles stephensi. Bioresour. Technol. 2006, 97, 2481–2484. [Google Scholar] [CrossRef]
- Komalamisra, N.; Trongtokit, Y.; Rongsriyam, Y.; Apiwathnasorn, C. Screening for lavicidal activity in some Thai plants against four mosquito vector species. Southeast Asian J. Trop. Med. Public Health 2005, 36, 1412–1422. [Google Scholar]
- Vivekanandhan, P.; Venkatesan, R.; Ramkumar, G.; Karthi, S.; Senthil-Nathan, S.; Shivakumar, M.S. Comparative analysis of major mosquito vectors response to seed-derived essential oil and seed pod-derived extract from Acacia nilotica. Int. J. Environ. Res. Public Health 2018, 15, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Akhtar, N.; Khan, B.A.; Khan, M.S.; Rasul, A. Acacia nilotica: A plant of multipurpose medicinal uses. J. Med. Plants Res. 2012, 6, 1492–1496. [Google Scholar] [CrossRef]
- Intirach, J.; Junkum, A.; Lumjuan, N.; Chaithong, U.; Jitpakdi, A.; Riyong, D.; Wannasan, A.; Champakaew, D.; Muangmoon, R.; Chansang, A.; et al. Antimosquito property of Petroselinum crispum (Umbellifereae) against the pyrethroid resistant and susceptible strains of Aedes aegypti (Diptera: Culicidae). Environ. Sci. Pollut. Res. 2016, 23, 23994–24008. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.S.; Huang, C.G.; Chen, Y.J.; Yu, J.J.; Chen, W.J.; Chang, S.T. Chemical compositions and larvicidal activities of leaf essential oils from two Eucalyptus species. Bioresour. Technol. 2009, 100, 452–456. [Google Scholar] [CrossRef]
- Costa, A.A.; Naspi, C.V.; Lucia, A.; Masuh, H.M.; Barrera, R. Repellent and larvicidal activity of the essential oil from Eucalyptus nitens against Aedes aegypti and Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 2017, 54, 670–676. [Google Scholar] [CrossRef]
- Gillij, Y.G.; Gleiser, R.M.; Zygadlo, J.A. Mosquito repellent activity of essential oils of aromatic plants growing in Argentina. Bioresour. Technol. 2008, 99, 2507–2515. [Google Scholar] [CrossRef]
- Cheng, S.-S.; Liu, J.-Y.; Tsai, K.-H.; Chen, W.-J.; Chang, S.-T. Chemical composition and mosquito larvicidal activity of essential oils from leaves of different Cinnamomum osmophloeum provenances. J. Agric. Food Chem. 2004, 52, 4395–4400. [Google Scholar] [CrossRef]
- Mendes, L.A.; Martins, G.F.; Valbon, W.R.; da Silva de Souza, T.; Menini, L.; Ferreira, A.; da Silva Ferreira, M.F. Larvicidal effect of essential oils from Brazilian cultivars of Guava on Aedes aegypti L. Ind. Crops Prod. 2017, 108, 684–689. [Google Scholar] [CrossRef]
- Oliveira, A.E.; Duarte, J.L.; Amado, J.R.R.; Cruz, R.A.S.; Rocha, C.F.; Souto, R.N.P.; Ferreira, R.M.A.; Santos, K.; da Conceição, E.C.; de Oliveira, L.A.R.; et al. Development of a larvicidal nanoemulsion with Pterodon emarginatus Vogel oil. PLoS ONE 2016, 11, e0145835. [Google Scholar] [CrossRef]
- Santos, G.K.N.; Dutra, K.A.; Barros, R.A.; da Câmara, C.A.G.; Lira, D.D.; Gusmão, N.B.; Navarro, D.M.A.F. Essential oils from Alpinia purpurata (Zingiberaceae): Chemical composition, oviposition deterrence, larvicidal and antibacterial activity. Ind. Crops Prod. 2012, 40, 254–260. [Google Scholar] [CrossRef]
- Botas, G.D.S.; Cruz, R.A.S.; de Almeida, F.B.; Duarte, J.L.; Araújo, R.S.; Souto, R.N.P.; Ferreira, R.; Carvalho, J.C.T.; Santos, M.G.; Rocha, L.; et al. Baccharis reticularia DC. and limonene nanoemulsions: Promising larvicidal agents for Aedes aegypti (Diptera: Culicidae) control. Molecules 2017, 22, 1990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, J.L.; Amado, J.R.R.; Oliveira, A.E.M.F.M.; Cruz, R.A.S.; Ferreira, A.M.; Souto, R.N.P.; Falcão, D.Q.; Carvalho, J.C.T.; Fernandes, C.P. Evaluation of larvicidal activity of a nanoemulsion of Rosmarinus officinalis essential oil. Braz. J. Pharmacog. 2015, 25, 189–192. [Google Scholar] [CrossRef] [Green Version]
- Santos, G.K.N.; Dutra, K.A.; Lira, C.S.; Lima, B.N.; Napoleão, T.H.; Paiva, P.M.G.; Maranhão, C.A.; Brandão, S.S.F.; Navarro, D.M.A.F. Effects of Croton rhamnifolioides essential oil on Aedes aegypti oviposition, larval toxicity and trypsin activity. Molecules 2014, 19, 16573–16587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, S.S.; Lin, C.Y.; Chung, M.J.; Liu, Y.H.; Huang, C.G.; Chang, S.T. Larvicidal activities of wood and leaf essential oils and ethanolic extracts from Cunninghamia konishii Hayata against the Dengue mosquitoes. Ind. Crops Prod. 2013, 47, 310–315. [Google Scholar] [CrossRef]
- Govindarajan, M.; Sivakumar, R.; Rajeswari, M.; Yogalakshmi, K. Chemical composition and larvicidal activity of essential oil from Mentha spicata (Linn.) against three mosquito species. Parasitol. Res. 2012, 110, 2023–2032. [Google Scholar] [CrossRef]
- Fayemiwo, K.A.; Adeleke, M.A.; Okoro, O.P.; Awojide, S.H.; Awoniyi, I.O.; Ademolu, K.O. Larvicidal efficacies and chemical composition of essential oils of Pinus sylvestris and Syzygium aromaticum against mosquitoes. Asian Pac. J. Trop. Biomed. 2014, 4, 30–34. [Google Scholar] [CrossRef]
- Oliveira, G.; Cardoso, S.; Junior, C.L. Chemical study and larvicidal activity against Aedes aegypti of essential oil of Piper aduncum L. (Piperaceae). An. Acad. Bras. Cienc. 2013, 85, 1227–1234. [Google Scholar] [CrossRef] [Green Version]
- Fontes, U.R., Jr.; Ramos, C.S.; Serafini, M.R.; Cavalcanti, S.C.H.; Alves, P.B.; Lima, G.M.; Andrade, P.H.S.; Bonjardim, L.R.; Quintans, L.J., Jr.; Araujo, A.A.S. Evaluation of the lethality of Porophyllum ruderale essential oil against Biomphalaria glabrata, Aedes aegypti and Artemia salina. Afr. J. Biotechnol. 2012, 11, 3169–3172. [Google Scholar] [CrossRef]
- Ali, A.; Tabanca, N.; Kurkcuoglu, M.; Duran, A.; Blythe, E.K.; Khan, I.A.; Baser, K.H.C. Chemical composition, larvicidal, and biting deterrent activity of essential oils of two subspecies of Tanacetum argenteum (Asterales: Asteraceae) and individual constituents against Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2014, 51, 824–830. [Google Scholar] [CrossRef] [PubMed]
- Chellappandian, M.; Thanigaivel, A.; Vasantha-Srinivasan, P.; Edwin, E.S.; Ponsankar, A.; Selin-Rani, S.; Kalaivani, K.; Senthil-Nathan, S.; Benelli, G. Toxicological effects of Sphaeranthus indicus Linn. (Asteraceae) leaf essential oil against human disease vectors, Culex quinquefasciatus Say and Aedes aegypti Linn., and impacts on a beneficial mosquito predator. Environ. Sci. Pollut. Res. 2018, 25, 10294–10306. [Google Scholar] [CrossRef]
- De Sousa, L.M.; de Carvalho, J.L.; Gois, R.W.S.; da Silva, H.C.; Santiago, G.M.P.; Lemos, T.L.G.; Arriaga, A.M.C.; Alves, P.B.; de Matos, I.L.; Militão, G.C.G.; et al. Chemical composition, larvicidal and cytotoxic activities of the essential oils from two Bauhinia species. Rec. Nat. Prod. 2016, 10, 341–348. [Google Scholar]
- Kerdudo, A.; Gonnot, V.; Ellong, E.N.; Boyer, L.; Chandre, F.; Adenet, S.; Rochefort, K.; Michel, T.; Fernandez, X. Composition and bioactivity of Pluchea carolinensis (Jack.) G. essential oil from Martinique. Ind. Crops Prod. 2016, 89, 295–302. [Google Scholar] [CrossRef]
- Kovendan, K.; Murugan, K.; Vincent, S. Evaluation of larvicidal activity of Acalypha alnifolia Klein ex Willd. (Euphorbiaceae) leaf extract against the malarial vector, Anopheles stephensi, dengue vector, Aedes aegypti and Bancroftian filariasis vector, Culex quinquefasciatus (Diptera: Culicidae). Parasitol. Res. 2012, 110, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Krishnappa, K.; Elumalai, K. Toxicity of Aristolochia bracteata methanol leaf extract against selected medically important vector mosquitoes (Diptera: Culicidae). Asian Pacific J. Trop. Dis. 2012, 2, S553–S557. [Google Scholar] [CrossRef]
- Aziz, A.T.; Alshehri, M.A.; Panneerselvam, C.; Murugan, K.; Trivedi, S.; Mahyoub, J.A.; Hassan, M.M.; Maggi, F.; Sut, S.; Dall’Acqua, S.; et al. The desert wormwood (Artemisia herba-alba)—From arabian folk medicine to a source of green and effective nanoinsecticides against mosquito vectors. J. Photochem. Photobiol. B Biol. 2018, 180, 225–234. [Google Scholar] [CrossRef]
- Alam, M.F.; Safhi, M.M.; Chopra, A.K.; Dua, V.K. Toxicological properties of several medicinal plants from the Himalayas (India) against vectors of malaria, filariasis and dengue. Trop. Biomed. 2011, 28, 343–350. [Google Scholar]
- El-Gamal, A.; Al-Massarani, S.; Fawzy, G.; Ati, H.; Al-Rehaily, A.; Basudan, O.; Abdel-Kader, M.; Tabanca, N.; Becnel, J. Chemical composition of Buddleja polystachya aerial parts and its bioactivity against Aedes aegypti. Nat. Prod. Res. 2017, 32, 2775–2782. [Google Scholar] [CrossRef]
- Govindarajan, M.; Mathivanan, T.; Elumalai, K.; Krishnappa, K.; Anandan, A. Mosquito larvicidal, ovicidal, and repellent properties of botanical extracts against Anopheles stephensi, Aedes aegypti, and Culex quinquefasciatus (Diptera: Culicidae). Parasitol. Res. 2011, 109, 353–367. [Google Scholar] [CrossRef]
- Govindarajan, M. Bioefficacy of Cassia fistula Linn. (Leguminosae) leaf extract against chikungunya vector, Aedes aegypti (Diptera: Culicidae). Eur. Rev. Med. Pharmacol. Sci. 2009, 13, 99–103. [Google Scholar]
- Inocente, E.A.; Shaya, M.; Acosta, N.; Rakotondraibe, L.H.; Piermarini, P.M. A natural agonist of mosquito TRPA1 from the medicinal plant Cinnamosma fragrans that is toxic, antifeedant, and repellent to the yellow fever mosquito Aedes aegypti. PLoS Negl. Trop. Dis. 2018, 12, e0006265. [Google Scholar] [CrossRef] [Green Version]
- Rahuman, A.A.; Venkatesan, P. Larvicidal efficacy of five Cucurbitaceous plant leaf extracts against mosquito species. Parasitol. Res. 2008, 103, 133–139. [Google Scholar] [CrossRef]
- Dalarmi, L.; Silva, C.B.d.; Ocampos, F.M.; Burci, M.L.; Nascimento, K.F.d.; Jesus, C.d.; Dias Gaspari, J.d.F.; Miguel, M.D.; Miguel, O.G.; Zanin, S.M.W. Larvicidal activity of Dalbergia brasiliensis (Fabaceae—Papilionoideae). Afr. J. Pharm. Pharmacol. 2015, 9, 881–885. [Google Scholar]
- Nakano, H.; Ali, A.; Ur Rehman, J.; Mamonov, L.K.; Cantrell, C.L.; Khan, I.A. Toxicity of thiophenes from Echinops transiliensis (Asteraceae) against Aedes aegypti (Diptera: Culicidae) larvae. Chem. Biodivers. 2014, 11, 1001–1009. [Google Scholar] [CrossRef]
- Govindarajan, M.; Karuppannan, P. Mosquito larvicidal and ovicidal properties of Eclipta alba (L.) Hassk (Asteraceae) against chikungunya vector, Aedes aegypti (Linn.) (Diptera: Culicidae). Asian Pac. J. Trop. Med. 2011, 4, 24–28. [Google Scholar] [CrossRef] [Green Version]
- Rahuman, A.A.; Gopalakrishnan, G.; Venkatesan, P.; Geetha, K. Larvicidal activity of some Euphorbiaceae plant extracts against Aedes aegypti and Culex quinquefasciatus (Diptera: Culicidae). Parasitol. Res. 2008, 102, 867–873. [Google Scholar] [CrossRef]
- Govindarajan, M. Larvicidal efficacy of Ficus benghalensis L. plant leaf extracts against Culex quinquefasciatus Say, Aedes aegypti L. and Anopheles stephensi L. (Diptera: Culicidae). Eur. Rev. Med. Pharmacol. Sci. 2010, 14, 107–111. [Google Scholar]
- Ochieng, C.O.; Mid, J.O.; Owu, P.O. Anti-plasmodial and larvicidal effects of surface exudates of Gardenia ternifolia aerial parts. Res. J. Pharmacol. 2010, 4, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, E.A.; de Carvalho, C.M.D.; Costa, A.L.S.; Conceição, A.S.; Moura, F.D.B.P.; Santana, A.E.G. Bioactivity evaluation of plant extracts used in indigenous medicine against the snail, Biomphalaria glabrata, and the larvae of Aedes aegypti. Evid. Based Complement. Altern. Med. 2012, 2012, 846583. [Google Scholar] [CrossRef] [Green Version]
- Kumuda, S.S.; Mohankumar, T.K.; Prathibha, K.P.; Vijayan, V.A. Efficacy of plant extracts against the larvae of filariasis vector, Culex quinquefasciatus Say and the Dengue vector Aedes aegypti Linn at Mysore. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 242–249. [Google Scholar]
- Puthur, S.; Anoopkumar, A.N.; Rebello, S.; Aneesh, E.M. Hypericum japonicum: A double-headed sword to combat vector control and cancer. Appl. Biochem. Biotechnol. 2018, 186, 1–11. [Google Scholar] [CrossRef]
- Komansilan, A.; Suriani, N.W.; Lawalata, H. Test toxic tuba root extract as a natural insecticide on larvae of Aedes aegypti mosquito vector of dengue fever. Int. J. ChemTech Res. 2017, 10, 522–528. [Google Scholar]
- Reegan, A.D.; Gandhi, M.R.; Paulraj, M.G.; Balakrishna, K.; Ignacimuthu, S. Effect of niloticin, a protolimonoid isolated from Limonia acidissima L. (Rutaceae) on the immature stages of dengue vector Aedes aegypti L. (Diptera: Culicidae). Acta Trop. 2014, 139, 67–76. [Google Scholar] [CrossRef]
- Gusmão, D.S.; Páscoa, V.; Mathias, L.; Vieira, I.J.C.; Braz-Filho, R.; Lemos, F.J.A. Derris (Lonchocarpus) urucu (Leguminosae) extract modifies the peritrophic matrix structure of Aedes aegypti (Diptera: Culicidae). Mem. Inst. Oswaldo Cruz 2002, 97, 371–375. [Google Scholar] [CrossRef] [Green Version]
- Touré, S.; Nirma, C.; Falkowski, M.; Dusfour, I.; Boulogne, I.; Jahn-Oyac, A.; Coke, M.; Azam, D.; Girod, R.; Moriou, C.; et al. Aedes aegypti larvicidal sesquiterpene alkaloids from Maytenus oblongata. J. Nat. Prod. 2017, 80, 384–390. [Google Scholar] [CrossRef]
- Lalchhandama, K. Mosquitocidal activity of Millettia pachycarpa on the larvae and eggs of Aedes aegypti. Ann. Biol. Res. 2011, 2, 217–222. [Google Scholar]
- Govindarajan, M.; Ramya, A.; Sivakumar, R. Mosquito larvicidal properties of Mirabilis jalapa (Nyctaginaceae) against Anopheles stephensi, Aedes aegypti & Culex quinquefasciatus (Diptera: Culicidae). Indian J. Med. Res. 2014, 140, 438–440. [Google Scholar]
- Masi, M.; van der Westhuyzen, A.E.; Tabanca, N.; Evidente, M.; Cimmino, A.; Green, I.R.; Bernier, U.R.; Becnel, J.J.; Bloomquist, J.R.; van Otterlo, W.A.L.; et al. Sarniensine, a mesembrine-type alkaloid isolated from Nerine sarniensis, an indigenous South African Amaryllidaceae, with larvicidal and adulticidal activities against Aedes aegypti. Fitoterapia 2017, 116, 34–38. [Google Scholar] [CrossRef] [Green Version]
- Anees, A.M. Larvicidal activity of Ocimum sanctum Linn. (Labiatae) against Aedes aegypti (L.) and Culex quinquefasciatus (Say). Parasitol. Res. 2008, 103, 1451–1453. [Google Scholar] [CrossRef]
- Porto, K.R.d.A.; Motti, P.R.; Yano, M.; Roel, A.R.; Cardoso, C.A.L.; Matias, R. Screening of plant extracts and fractions on Aedes aegypti larvae found in the state of Mato Grosso do Sul (Linnaeus, 1762) (Culicidae). An. Acad. Bras. Cienc. 2017, 89, 895–906. [Google Scholar] [CrossRef] [Green Version]
- Kovendan, K.; Murugan, K.; Vincent, S.; Barnard, D.R. Mosquito larvicidal properties of Orthosiphon thymiflorus (Roth) Sleesen. (Family: Labiatae) against mosquito vectors, Anopheles stephensi, Culex quinquefasciatus and Aedes aegypti (Diptera: Culicidae). Asian Pac. J. Trop. Med. 2012, 5, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Samidurai, K.; Jebanesan, A.; Saravanakumar, A.; Govindarajan, M.; Pushpanathan, T. Larvicidal, ovicidal and repellent activities of Pemphis acidula Forst. (Lythraceae) against filarial and dengue vector mosquitoes. Acad. J. Entomol. 2009, 2, 62–66. [Google Scholar]
- Chaitong, U.; Choochote, W.; Kamsuk, K.; Jitpakdi, A.; Tippawangkosol, P.; Chaiyasit, D.; Champakaew, D.; Tuetun, B.; Pitasawat, B. Larvicidal effect of pepper plants on Aedes aegypti (L.) (Diptera: Culicidae). J. Vector Ecol. 2006, 31, 138–144. [Google Scholar] [CrossRef]
- Santiago, V.S.; Alvero, R.G.; Villaseñor, I.M. Aedes aegypti larvicide from the ethanolic extract of Piper nigrum black peppercorns. Nat. Prod. Res. 2015, 29, 441–443. [Google Scholar] [CrossRef]
- Govindarajan, M.; Rajeswary, M.; Sivakumar, R. Larvicidal & ovicidal efficacy of Pithecellobium dulce (Roxb.) Benth. (Fabaceae) against Anopheles stephensi Liston & Aedes aegypti Linn. (Diptera: Culicidae). Indian J. Med. Res. 2013, 138, 129–134. [Google Scholar]
- Azarudeen, R.M.S.T.; Govindarajan, M.; AlShebly, M.M.; AlQahtani, F.S.; Amsath, A.; Benelli, G. One pot green synthesis of colloidal silver nanocrystals using the Ventilago maderaspatana leaf extract: Acute toxicity on malaria, zika virus and filariasis mosquito vectors. J. Clust. Sci. 2017, 28, 369–392. [Google Scholar] [CrossRef] [Green Version]
- Raghavendra, K.; Singh, S.P.; Subbarao, S.K.; Dash, A.P. Laboratory studies on mosquito larvicidal efficacy of aqueous & hexane extracts of dried fruit of Solanum nigrum Linn. Indian J. Med. Res. 2009, 130, 74–77. [Google Scholar]
- Kumar, P.M.; Murugan, K.; Kovendan, K.; Panneerselvam, C.; Kumar, K.P.; Amerasan, D.; Subramaniam, J.; Kalimuthu, K.; Nataraj, T. Mosquitocidal activity of Solanum xanthocarpum fruit extract and copepod Mesocyclops thermocyclopoides for the control of dengue vector Aedes aegypti. Parasitol. Res. 2012, 111, 609–618. [Google Scholar] [CrossRef]
- Munhoz, V.M.; Longhini, R.; Souza, J.R.P.; Zequi, J.A.C.; Mello, E.V.S.L.; Lopes, G.C.; Mello, J.C.P. Extraction of flavonoids from Tagetes patula: Process optimization and screening for biological activity. Braz. J. Pharmacog. 2014, 24, 576–583. [Google Scholar] [CrossRef] [Green Version]
- Kovendan, K.; Chandramohan, B.; Govindarajan, M.; Jebanesan, A.; Kamalakannan, S.; Vincent, S.; Benelli, G. Orchids as sources of novel nanoinsecticides? Efficacy of Bacillus sphaericus and Zeuxine gracilis—fabricated silver nanoparticles against dengue, malaria and filariasis mosquito vectors. J. Clust. Sci. 2018, 29, 345–357. [Google Scholar] [CrossRef]
- Govindarajan, M.; Mathivanan, T.; Elumalai, K.; Krishnappa, K.; Anandan, A. Ovicidal and repellent activities of botanical extracts against Culex quinquefasciatus, Aedes aegypti and Anopheles stephensi (Diptera: Culicidae). Asian Pac. J. Trop. Biomed. 2011, 1, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Govindarajan, M.; Sivakumar, R. Repellent properties of Cardiospermum halicacabum Linn. (Family: Sapindaceae) plant leaf extracts against three important vector mosquitoes. Asian Pac. J. Trop. Med. 2012, 2, 602–607. [Google Scholar] [CrossRef] [Green Version]
- Govindarajan, M. Ovicidal and repellent properties of Coccinia indica Wight and Arn. (Family: Cucurbitaceae) against three important vector mosquitoes. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 1010–1019. [Google Scholar] [PubMed]
- Bhargava, S. Mosquito repellency potential of the essential oil from Mentha spp. Int. J. Pharm. Pharm. Res. 2016, 6, 509–522. [Google Scholar]
- Kumar, S.; Singh, A.P.; Nair, G.; Batra, S.; Seth, A.; Wahab, N.; Warikoo, R. Impact of Parthenium hysterophorus leaf extracts on the fecundity, fertility and behavioural response of Aedes aegypti L. Parasitol. Res. 2011, 108, 853–859. [Google Scholar] [CrossRef]
- Dick, F.D. Solvent Neurotoxicity. Occup. Environ. Med. 2006, 63, 221–226. [Google Scholar] [CrossRef]
- Vincent, J.F.V.; Wegst, U.G.K. Design and mechanical properties of insect cuticle. Arthropod Struct. Dev. 2004, 33, 187–199. [Google Scholar] [CrossRef]
- Fahmy, M.A.H.; Mallipudi, N.M.; Fukuto, T.R. Selective toxicity of N,N′-thiodicarbamates. J. Agric. Food Chem. 1978, 26, 550–557. [Google Scholar] [CrossRef]
- Ferreira, M.C.; Cantrell, C.L.; Duke, S.O.; Ali, A.; Rosa, L.H. New pesticidal diterpenoids from Vellozia gigantea (Velloziaceae), an endemic neotropical plant living in the endangered Brazilian biome rupestrian grasslands. Molecules 2017, 22, 175. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, G.N.; Trindade, F.T.; Dos Santos, F.; Gosmann, G.; E Silva, A.A.; Gnoatto, S.C. Larvicidal activity of natural and modified triterpenoids against Aedes aegypti (Diptera: Culicidae). Pest Manag. Sci. 2016, 72, 1883–1887. [Google Scholar] [CrossRef]
- Masi, M.; Cimmino, A.; Tabanca, N.; Becnel, J.J.; Bloomquist, J.R.; Evidente, A. A survey of bacterial, fungal and plant metabolites against Aedes aegypti (Diptera: Culicidae), the vector of yellow and dengue fevers and zika virus. Open Chem. 2017, 15, 156–166. [Google Scholar] [CrossRef]
- Rahuman, A.A.; Gopalakrishnan, G.; Venkatesan, P.; Geetha, K.; Bagavan, A. Mosquito larvicidal activity of isolated compounds from the rhizome of Zingiber officinale. Phyther. Res. 2008, 22, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Park, I.K.; Lee, S.G.; Shin, S.C.; Park, J.D.; Ahn, Y.J. Larvicidal activity of isobutylamides identified in Piper nigrum fruits against three mosquito species. J. Agric. Food Chem. 2002, 50, 1866–1870. [Google Scholar] [CrossRef] [PubMed]
- Park, I.K. Insecticidal activity of isobutylamides derived from Piper nigrum against adult of two mosquito species, Culex pipiens pallens and Aedes aegypti. Nat. Prod. Res. 2012, 26, 2129–2131. [Google Scholar] [CrossRef]
- Yang, Y.C.; Lee, S.G.; Lee, H.K.; Kim, M.K.; Lee, S.H.; Lee, H.S. A Piperidine amide extracted from Piper longum L. fruit shows activity against Aedes aegypti mosquito larvae. J. Agric. Food Chem. 2002, 50, 3765–3767. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, B.S.; Gulzar, T.; Begum, S.; Afshan, F.; Sattar, F.A. Insecticidal amides from fruits of Piper nigrum Linn. Nat. Prod. Res. 2005, 19, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Cala, A.; Tabanca, N.; Cimmino, A.; Green, I.R.; Bloomquist, J.R.; Van Otterlo, W.A.L.; Macias, F.A.; Evidente, A. Alkaloids with activity against the Zika virus vector Aedes aegypti (L.)-crinsarnine and sarniensinol, two new crinine and mesembrine type alkaloids isolated from the South African plant Nerine sarniensis. Molecules 2016, 21, 1432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.-W.; Dong, C.-Z.; Zhou, X.-M.; Bu, M.-M.; Yu, S.-Q. Aristololactam derivatives from the fruits of Aristolochia contorta Bunge. Nat. Prod. Res. 2018, 32, 2505–2509. [Google Scholar] [CrossRef]
- Liu, X.C.; Lai, D.; Liu, Q.Z.; Zhou, L.; Liu, Q.; Liu, Z.L. Bioactivities of a new pyrrolidine alkaloid from the root barks of Orixa japonica. Molecules 2016, 21, 1665. [Google Scholar] [CrossRef] [Green Version]
- Sivakumar, R.; Jebanesan, A.; Govindarajan, M.; Rajasekar, P. Larvicidal and repellent activity of tetradecanoic acid against Aedes aegypti (Linn.) and Culex quinquefasciatus (Say.) (Diptera: Culicidae). Asian Pac. J. Trop. Med. 2011, 4, 706–710. [Google Scholar] [CrossRef] [Green Version]
- Sivakumar, R.; Jebanesan, A.; Govindarajan, M.; Rajasekar, P. Oviposition attractancy of dodecanoic, hexadecanoic and tetradecanoic acids against Aedes aegypti and Culex quinquefasciatus (Diptera: Culicidae). Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 1172–1175. [Google Scholar]
- Macedo, A.L.; Duprat, R.C.; Moreira, D.d.L.; Kaplan, M.A.C.; Vasconcelos, T.R.A.; Pinto, L.C.; Montenegro, R.C.; Ratcliffe, N.A.; Mello, C.B.; Valverde, A.L. Isolation of a larvicidal compound from Piper solmsianum C.DC. (Piperaceae). Nat. Prod. Res. 2017, 32, 2701–2704. [Google Scholar] [CrossRef]
- Leite, A.C.C.F.; Kato, M.J.; Soares, R.O.A.; Guimarães, A.E.; Santos-Mallet, J.R.; Cabral, M.M.O. Grandisin caused morphological changes larval and toxicity on Aedes aegypti. Braz. J. Pharmacog. 2012, 22, 517–521. [Google Scholar] [CrossRef] [Green Version]
- Maleck, M.; Dos Santos, F.C.; Serdeiro, M.; Ferreira, B.; Gunaydin, K.; De Almeida, A. Khellin: A furanochromone with toxicity against Oncopeltus fasciatus (Hemiptera) and Aedes aegypti (Diptera). J. Nat. Pharm. 2013, 4, 32–36. [Google Scholar] [CrossRef] [Green Version]
- Meepagala, K.M.; Estep, A.S.; Becnel, J.J. Mosquitocidal activity of extracts from Ammi visnaga (Apiaceae) seeds. J. Agric. Chem. Environ. 2016, 5, 170–178. [Google Scholar] [CrossRef] [Green Version]
- von Oppen, S.; Masuh, H.; Licastro, S.; Zerba, E.; Gonzalez-Audino, P. A Floral-derived attractant for Aedes aegypti mosquitoes. Entomol. Exp. Appl. 2015, 155, 184–192. [Google Scholar]
- Braga, I.A.; Valle, D. Aedes aegypti: Inseticidas, mecanismos de ação e resistência. Epidemiol. Serv. Saúde 2007, 16, 279–293. [Google Scholar] [CrossRef] [Green Version]
- Kostyukovsky, M.; Rafaeli, A.; Gileadi, C.; Demchenko, N.; Shaaya, E. Activation of octopaminergic receptors by essential oil constituents isolated from aromatic plants: Possible mode of action against insect pests. Pest Manag. Sci. 2002, 58, 1101–1106. [Google Scholar] [CrossRef]
- Rattan, R.S. Mechanism of action of insecticidal secondary metabolites of plant origin. Crop Prot. 2010, 29, 913–920. [Google Scholar] [CrossRef]
- Khanikor, B.; Parida, P.; Yadav, R.N.S.; Bora, D. Comparative mode of action of some terpene compounds against octopamine receptor and acetylcholinesterase of mosquito and human system by the help of homology modeling and docking studies. J. Appl. Pharm. Sci. 2013, 3, 6–12. [Google Scholar]
- Pontual, E.V.; Napoleão, T.H.; Dias de Assis, C.R.; de Souza Bezerra, R.; Xavier, H.S.; Navarro, D.M.d.A.F.; Coelho, L.C.B.B.; Paiva, P.M.G. Effect of Moringa oleifera flower extract on larval trypsin and acethylcholinesterase activities in Aedes aegypti. Arch. Insect Biochem. Physiol. 2012, 79, 135–152. [Google Scholar] [CrossRef]
- Perumalsamy, H.; Kim, J.R.; Oh, S.M.; Jung, J.W.; Ahn, Y.J.; Kwon, H.W. Novel histopathological and molecular effects of natural compound pellitorine on larval midgut epithelium and anal gills of Aedes aegypti. PLoS ONE 2013, 8, e80226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohlit, A.M.; Lopes, N.P.; Gama, R.A.; Tadei, W.P.; Neto, V.F. Patent literature on mosquito repellent inventions which contain plant essential oils—A review. Planta Med. 2011, 77, 598–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, J.L.; Campos, E.V.R.; Fraceto, L.F. Recent developments and challenges for nanoscale formulation of botanical pesticides for use in sustainable agriculture. J. Agric. Food Chem. 2018, 66, 8898–8913. [Google Scholar] [CrossRef] [PubMed]
- Valdiani, A.; Hansen, O.K.; Nielsen, U.B.; Johannsen, V.K.; Shariat, M.; Georgiev, M.I.; Omidvar, V.; Ebrahimi, M.; Dinanai, E.T.; Abiri, R. Bioreactor-based advances in plant tissue and cell culture: Challenges and prospects. Crit. Ver. Biotechnol. 2018, 39, 20–34. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, P.; Zhou, M.; Wang, T.; Fang, S.; Shang, X.; Fu, X. Geographic variation in the chemical composition and antioxidant properties of phenolic compounds from Cyclocarya paliurus (Batal) Iljinskaja leaves. Molecules 2018, 23, 2440. [Google Scholar] [CrossRef] [Green Version]
- Zouari, N.; Ayadi, I.; Fakhfakh, N.; Rebai, A.; Zouari, S. Variation of chemical composition of essential oils in wild populations of Thymus algeriensis Boiss. et Reut., a North African endemic species. Lipids Health Dis. 2012, 11, 28. [Google Scholar] [CrossRef] [Green Version]
- Al-Hamwi, M.; Bakkour, Y.; Abou-Ela, M.; El-Lakany, A.; Tabcheh, M.; El-Omar, F. Chemical composition and seasonal variation of the essential oil of Micromeria fruticosa. J. Nat. Prod. 2011, 4, 147–150. [Google Scholar]
- Gobbo-Neto, L.; Lopes, N.P. Plantas medicinais: Fatores de influência no conteúdo de metabólitos secundários. Quim. Nova 2007, 30, 374–381. [Google Scholar] [CrossRef]
- Workman, M.J.; Gomes, B.; Weng, J.-L.; Ista, L.K.; Jesus, C.P.; David, M.R.; Ramalho-Ortigao, M.; Genta, F.A.; Matthews, S.K.; Durvasula, R.; et al. Yeast-encapsulated essential oils: A new perspective as an environmentally friendly larvicide. Parasite Vector 2020, 13, 1–19. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for Laboratory and Field Testing of Mosquito Larvicides. 2005. Available online: http://apps.who.int/iris/bitstream/handle/10665/69101/WHO_CDS_WHOPES_GCDPP_2005.13.pdf?sequence=1 (accessed on 21 February 2020).
- Isman, M.B.; Grieneisen, M.L. Botanical insecticide research: Many publications, limited useful data. Trends Plant Sci. 2014, 19, 140–145. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F. Insecticides mode of action in relation to their toxicity to non-target organisms. J. Environ. Anal. Toxicol. 2011, S4, S4-002. [Google Scholar] [CrossRef] [Green Version]
- Borrero-Landazabal, M.A.; Duque, J.E.; Mendez-Sanchez, S.C. Model to design insecticides against Aedes aegypti using in silico and in vivo analysis of different pharmacological targets. Comp. Biochem. Phys. C 2020, 229, 108664. [Google Scholar] [CrossRef] [PubMed]
- Rueda, A.G.; Otero, A.L.C.; Duque, J.E.; Kouznetsov, V.V. Synthesis of new α-amino nitriles with insecticidal action on Aedes aegypti (Diptera: Culicidae). Rev. Bras. Entomol. 2018, 62, 112–118. [Google Scholar] [CrossRef]
- Devillers, J.; Lagneau, C.; Lattes, A.; Garrigues, J.C.; Clémenté, M.M.; Yébakima, A. In silico models for predicting vector control chemicals targeting Aedes aegypti. SAR QSAR Environ. Res. 2014, 25, 805–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Y.-J.; Jia, Y.-F.; Chen, N.; Bian, W.-P.; Li, Q.-K.; Ma, Y.-B.; Chen, Y.-L.; Pei, D.-S. Zebrafish as a model system to study toxicology. Environ. Toxicol. Chem. 2014, 33, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Organisation for Economic Co-operation and Development. OECD Guidelines for the Testing of Chemicals. 2013. Available online: https://0-www-oecd--ilibrary-org.brum.beds.ac.uk/docserver/9789264203709-en.pdf?expires=1591301187&id=id&accname=guest&checksum=30910D5C63FC44FE9C43DDAB70F56739 (accessed on 23 March 2020).
- Maia, J.D.; Corte, R.L..; Martinez, J.; Ubbink, J.; Prata, A.S. Improved activity of thyme essential oil (Thymus vulgaris) against Aedes aegypti larvae using a biodegradable controlled release system. Ind. Crops. Prod. 2019, 136, 110–120. [Google Scholar] [CrossRef]
- Benelli, G.; Mehlhorn, H. Declining malaria, rising of dengue and Zika virus: Insights for mosquito vector control. Parasitol. Res. 2016, 115, 1747–1754. [Google Scholar] [CrossRef]
- Ferreira, T.P.; Haddi, K.; Corrêa, R.F.T.; Zapata, V.L.B.; Piau, T.B.; Souza, L.F.N.; Santos, S.-M.G.; Oliveira, E.E.; Jumbo, L.O.V.; Ribeiro, B.M.; et al. Prolonged mosquitocidal activity of Siparuna guianensis essential oil encapsulated in chitosan nanoparticles. PLoS Negl. Trop. Dis. 2019, 13, e0007624. [Google Scholar] [CrossRef] [PubMed] [Green Version]














| Plant Species | Family | Country | Part Used | Extraction Solvent | Larval Stage | Mortality | Time (h) | Reference | |
|---|---|---|---|---|---|---|---|---|---|
| LC50 (ppm) | LC90 (ppm) | ||||||||
| Acacia nilótica (L.) Delile | Fabaceae | India | Seed pod | n-Hexane | L4 | 169.25 | 201.62 | 24 | [58] |
| Benzene | L4 | 45.32 | 99.32 | 24 | [58] | ||||
| Chloroform | L4 | 158.13 | 198.24 | 24 | [58] | ||||
| Ethyl acetate | L4 | 59.12 | 75.82 | 24 | [58] | ||||
| Acetone | L4 | 103.68 | 162.03 | 24 | [58] | ||||
| Acalypha alnifolia Klein ex Willd. | Euphorbiaceae | India | Leaves | n-Hexane | L4 | 202.15 | 476.57 | 24 | [80] |
| Chloroform | L4 | 182.58 | 460.83 | 24 | [80] | ||||
| Ethyl acetate | L4 | 160.35 | 440.78 | 24 | [80] | ||||
| Acetone | L4 | 146.07 | 415.38 | 24 | [80] | ||||
| Methanol | L4 | 128.55 | 381.67 | 24 | [80] | ||||
| Aristolochia bracteata Retz. | Aristolochiaceae | India | Leaves | Methanol | L3 | 114.89 | 216.24 | 24 | [81] |
| Artemisia herba-alba Asso | Asteraceae | Saudi Arabia | Leaves | Water | L4—India | 117.18 | 227.63 | 24 | [82] |
| Water | L4—Saudi Arabia | 614.52 | 1273.33 | 24 | [82] | ||||
| Water + AgNP | L4—India | 10.70 | 21.24 | 24 | [82] | ||||
| Water + AgNP | L4—Saudi Arabia | 33.58 | 57.0 | 24 | [82] | ||||
| Boenninghausenia albiflora (Hook.) Rchb. ex Meisn. | Rutaceae | India | Leaves | Petroleum ether | L4 | 125.0 | 190.0 | ND | [83] |
| Buddleja polystachya Fresen. | Buddlejaceae | Saudi Arabia | Flowers | n-Butanol | L1 | ND | ND | ND | [84] |
| Caesalpinia pulcherrima (L.) Sw. | Fabaceae | India | Leaves | Ethyl acetate | L3 | 144.67 | 276.99 | 24 | [85] |
| Benzene | L3 | 136.36 | 272.15 | 24 | [85] | ||||
| Cassia fistula L. | Fabaceae | India | Leaves | Methanol | L3 | 10.69 | 20.47 | 24 | [86] |
| Benzene | L3 | 18.27 | 35.67 | 24 | [86] | ||||
| Acetone | L3 | 23.95 | 47.13 | 24 | [86] | ||||
| Catharanthus roseus (L.) G. Don | Apocynaceae | India | Leaves | Petroleum ether | L4 | 145.0 | 255.0 | 24 | [83] |
| Cinnamosma fragrans Baill. | Canellaceae | Madagascar | Root Bark | Methanol | L1 | 52.5 | ND | 24 | [87] |
| Citrullus colocynthis (L.) Schrad. | Cucurbitaceae | India | Leaves | Petroleum ether | L4 | 74.57 | 538.30 | 24 | [88] |
| Cunninghamia konishii Hayata | Taxodiaceae | Taiwan | Wood | Ethanol | L4 | 240.0 | >400.0 | 24 | [73] |
| Leaves | Ethanol | L4 | >400.0 | >400.0 | 24 | [73] | |||
| Dalbergia brasiliensis Vogel | Fabaceae | Brazil | Leaves | Ethanol | L3 | 30.0 | 91.0 | 24 | [89] |
| n-Hexane Fraction | L3 | 44.0 | 81.0 | 24 | [89] | ||||
| Chloroform Fraction | L3 | 33.0 | 75.0 | 24 | [89] | ||||
| Ethyl acetate Fraction | L3 | 24.0 | 66.0 | 24 | [89] | ||||
| Bark | Ethanol | L3 | 32.0 | 71.0 | 24 | [89] | |||
| n-Hexane Fraction | L3 | 31.0 | 72.0 | 24 | [89] | ||||
| Chloroform Fraction | L3 | 25.0 | 50.0 | 24 | [89] | ||||
| Ethyl acetate Fraction | L3 | 28.0 | 93.0 | 24 | [89] | ||||
| Echinops transiliensis Golosk. | Asteraceae | Kazakhstan | Root | Dichlorometane | ND | 3.21 | 6.81 | 24 | [90] |
| Eclipta alba (L.) Hassk | Asteraceae | India | Leaves | Benzene | L3 | 151.38 | 274.34 | 24 | [91] |
| n-Hexane | L3 | 165.10 | 297.70 | 24 | [91] | ||||
| Ethyl acetate | L3 | 154.88 | 288.61 | 24 | [91] | ||||
| Methanol | L3 | 127.64 | 245.73 | 24 | [91] | ||||
| Chloroform | L3 | 146.28 | 274.42 | 24 | [91] | ||||
| Ervatamia coronaria (Jacq.) Stapf. | Apocynaceae | India | Leaves | Ethyl acetate | L3 | 97.53 | 179.37 | 24 | [85] |
| Benzene | L3 | 89.59 | 166.04 | 24 | [85] | ||||
| Eupatorium odoratum L. | Asteraceae | India | Leaves | Petroleum ether | L4 | 155.0 | 290.0 | ND | [83] |
| Euphorbia hirta L. | Euphorbiaceae | India | Leaves | Petroleum ether | L4 | 272.36 | 703.76 | 24 | [92] |
| Euphorbia tirucalli L. | Euphorbiaceae | India | Stem bark | Petroleum ether | L4 | 4.25 | 13.14 | 24 | [92] |
| Ficus benghalensis L. | Moraceae | India | Leaves | Methanol | L2 | 56.54 | 109.29 | 24 | [93] |
| Methanol | L3 | 70.29 | 137.23 | 24 | [93] | ||||
| Methanol | L4 | 80.85 | 169.58 | 24 | [93] | ||||
| Benzene | L2 | 108.95 | 227.13 | 24 | [93] | ||||
| Benzene | L3 | 116.09 | 235.54 | 24 | [93] | ||||
| Benzene | L4 | 159.15 | 430.91 | 24 | [93] | ||||
| Acetone | L2 | 189.11 | 444.42 | 24 | [93] | ||||
| Acetone | L3 | 244.41 | 573.52 | 24 | [93] | ||||
| Acetone | L4 | 288.10 | 668.71 | 24 | [93] | ||||
| Gardenia ternifolia Schumach. & Thonn. | Rubiaceae | Kenya | Leaves | Acetone | L2 | 83.31 | ND | 24 | [94] |
| Methanol | L2 | 32.01 | ND | 24 | [94] | ||||
| Helicteres velutina K. Schum. | Malvaceae | Brazil | Root | Ethanol 90% | L4 | 171.68 | 403.61 | 48 | [95] |
| Stem | Ethanol 90% | L4 | 138.90 | 319.37 | 48 | [95] | |||
| Heracleum rigens Wall. | Apiaceae | India | Seed | Petroleum ether | L2 | 40.64 | 65.49 | 24 | [96] |
| Chloroform | L2 | 69.22 | 132.95 | 24 | [96] | ||||
| Ethyl acetate | L2 | 70.65 | 93.11 | 24 | [96] | ||||
| Methanol | L2 | 74.70 | 135.07 | 24 | [96] | ||||
| Acetone | L2 | 97.07 | 198.99 | 24 | [96] | ||||
| Petroleum ether | L3 | 91.55 | 162.09 | 24 | [96] | ||||
| Chloroform | L3 | 114.25 | 179.99 | 24 | [96] | ||||
| Ethyl acetate | L3 | 143.48 | 207.45 | 24 | [96] | ||||
| Methanol | L3 | 195.57 | 348.56 | 24 | [96] | ||||
| Acetone | L3 | 234.77 | ND | 24 | [96] | ||||
| Petroleum ether | L4 | 113.69 | 171.12 | 24 | [96] | ||||
| Chloroform | L4 | 144.64 | 209.16 | 24 | [96] | ||||
| Ethyl acetate | L4 | 165.43 | 230.21 | 24 | [96] | ||||
| Methanol | L4 | 231.26 | 361.68 | 24 | [96] | ||||
| Acetone | L4 | 308.65 | 577.14 | 24 | [96] | ||||
| Hypericum japonicum Thunb. | Hypericaceae | India | Whole plant | Acetone | L4 | 13.15 | 18.54 | 24 | [97] |
| n-Hexane | L4 | 9.63 | 19.53 | 24 | [97] | ||||
| Petroleum ether | L4 | 8.27 | 15.2 | 24 | [97] | ||||
| Methanol | L4 | 7.37 | 11.59 | 24 | [97] | ||||
| Jatropha curcas L. | Euphorbiaceae | India | Leaves | Petroleum ether | L4 | 8.79 | 35.39 | 24 | [92] |
| Indonesia | Root | Ethanol | ND | 44.75 | ND | 24 | [98] | ||
| Limonia acidíssima L. | Rutaceae | India | Leaves | n-Hexane—Purified fractions | L3 | 4.11 to 23.53 | ND | 24 | [99] |
| Lonchocarpus urucu Killip & A.C. Sm. | Fabaceae | Brazil | Medulla Root | Methanol | L4 | 33.32 | 83.69 | 24 | [100] |
| Bark Root | Methanol | L4 | 17.60 | 55.40 | 24 | [100] | |||
| Maytenus oblongata Reissek | Celasteraceae | French Guiana | Bark | Ethyl acetate | L3/L4 | 74.40 | ND | 24 | [101] |
| Millettia pachycarpa Benth. | Fabaceae | India | Root | Ethanol | L3 | 98.47 | ND | 24 | [102] |
| Mirabilis jalapa L. | Nyctaginaceae | India | Leaves | Benzene | L3 | 97.03 | 172.15 | 24 | [103] |
| Chloroform | L3 | 88.20 | 162.16 | 24 | [103] | ||||
| Ethyl acetate | L3 | 72.77 | 127.91 | 24 | [103] | ||||
| Methanol | L3 | 64.58 | 120.28 | 24 | [103] | ||||
| Momordica charantia L. | Cucurbitaceae | India | Leaves | Methanol | L4 | 199.14 | 780.10 | 24 | [88] |
| Myristica fragans Houtt. | Myristicaceae | India | Leaves | Methanol | L1 | 162.03 | 502.04 | 24 | [53] |
| Methanol | L2 | 194.11 | 542.56 | 24 | [53] | ||||
| Methanol | L3 | 240.10 | 604.78 | 24 | [53] | ||||
| Methanol | L4 | 273.90 | 660.96 | 24 | [53] | ||||
| Methanol + ZnO NP | L1 | 3.44 | 18.35 | 24 | [53] | ||||
| Methanol + ZnO NP | L2 | 5.25 | 30.37 | 24 | [53] | ||||
| Methanol + ZnO NP | L3 | 8.02 | 39.14 | 24 | [53] | ||||
| Methanol + ZnO NP | L4 | 10.28 | 44.07 | 24 | [53] | ||||
| Myristica fragrans Houtt. | Myristicaceae | Thailand | Flowers | Ethanol | L4 (p-s) | 75.45 | 123.60 | 24 | [60] |
| Nerine sarniensis (L.) Herb. | Amaryllidaceae | South Africa | Bulb | Ethyl acetate | L1 | 8.0 | ND | 24 | [104] |
| Nyctanthes arbor-tristis L. | Oleaceae | India | Leaves | Petroleum ether | L4 | 180.0 | 340.0 | [83] | |
| Ocimum sanctum L. | Labiatae | India | Leaves and Flowers | Acetone | L4 | 425.94 | ND | 24 | [105] |
| Chloroform | L4 | 150.40 | ND | 24 | [105] | ||||
| Ethyl acetate | L4 | 350.78 | ND | 24 | [105] | ||||
| n-Hexane | L4 | 575.26 | ND | 24 | [105] | ||||
| Methanol | L4 | 175.67 | ND | 24 | [105] | ||||
| Ormosia arborea Vell | Fabaceae | Brazil | Leaves | Ethanol | L3 | 238.0 | 347.0 | 24 | [106] |
| Seeds | Ethanol | L3 | 111.0 | 194.0 | 24 | [106] | |||
| Orthosiphon thymiflorus (Roth) Sleesen | Labiatae | India | Leaves | n-Hexane | L3 | 228.13 | 526.12 | 24 | [107] |
| Chloroform | L3 | 209.72 | 502.84 | 24 | [107] | ||||
| Ethyl acetate | L3 | 183.35 | 463.35 | 24 | [107] | ||||
| Acetone | L3 | 163.55 | 442.32 | 24 | [107] | ||||
| Methanol | L3 | 149.96 | 426.16 | 24 | [107] | ||||
| Pedilanthus tithymaloides (L.) Poit. | Euphorbiaceae | India | Leaves | Petroleum ether | L4 | 55.26 | 256.77 | 24 | [92] |
| Pemphis acidula J.R. Forst. & G. Forst. | Lythraceae | India | Leaves | Methanol | L3 | 22.10 | 43.71 | 24 | [108] |
| Benzene | L3 | 43.99 | 84.87 | 24 | [108] | ||||
| Acetone | L3 | 57.66 | 106.51 | 24 | [108] | ||||
| Phyllanthus amarus Schumach. & Thonn. | Euphorbiaceae | India | Leaves | Petroleum ether | L4 | 90.92 | 384.19 | 24 | [92] |
| Piper aduncum L. | Piperaceae | Brazil | Leaves | n-Hexane | L3 | 342.0 | 473.0 | 24 | [106] |
| Chloroform | L3 | 192.0 | 346.0 | 24 | [106] | ||||
| Piper hispidum Sw. | Piperaceae | Brazil | Leaves | Ethanol | L3 | 169.0 | 474.0 | 24 | [106] |
| Chloroform | L3 | 567.0 | 1003.0 | 24 | [106] | ||||
| Piper longum L. | Piperaceae | Thailand | Fruits | Ethanol | L4 | 2.23 | ND | 24 | [109] |
| Piper nigrum L. | Piperaceae | Philippines | Peppercorns | Ethanol | L3/L4 | 71.25 | 9.37 | 24 | [110] |
| Ethanol—Fraction 1A | L3/L4 | 17.10 | 3.84 | 24 | [110] | ||||
| Ethanol—Fraction 1B | L3/L4 | 18.10 | 3.84 | 24 | [110] | ||||
| Piper ribesoides Wall. | Piperaceae | Thailand | Wood | Ethanol | L4 | 8.13 | ND | 24 | [109] |
| Piper sarmentosum Roxb. ex Hunt. | Piperaceae | Thailand | Whole plant | Ethanol | L4 | 4.06 | ND | 24 | [109] |
| Pithecellobium Dulce (Roxb.) Benth. | Fabaceae | India | Leaves | Methanol | L3 | 155.78 | 279.73 | 24 | [111] |
| Ethyl acetate | L3 | 162.36 | 283.43 | 24 | [111] | ||||
| Chloroform | L3 | 169.08 | 293.17 | 24 | [111] | ||||
| Benzene | L3 | 176.02 | 308.88 | 24 | [111] | ||||
| n-Hexane | L3 | 185.14 | 316.46 | 24 | [111] | ||||
| Seeds | Methanol | L3 | 193.66 | 377.39 | 24 | [111] | |||
| Ethyl acetate | L3 | 215.63 | 416.51 | 24 | [111] | ||||
| Chloroform | L3 | 240.39 | 461.28 | 24 | [111] | ||||
| Benzene | L3 | 259.42 | 489.41 | 24 | [111] | ||||
| n-Hexane | L3 | 281.18 | 516.33 | 24 | [111] | ||||
| Scoparia dulcis L. | Plantaginaceae | Brazil | Leaves | Ethanol 90% | L4 | 83.43 | 158.83 | 48 | [112] |
| Solanum nigrum L. | Solanaceae | India | Fruit | Water | L3/L4 | 359.0 | 931.0 | 24 | [113] |
| n-Hexane | L3/L4 | 17.63 | 65.22 | 24 | [113] | ||||
| Solanum variabile Mart. | Solanaceae | Brazil | Leaves | Ethanol | L3 | 188.0 | 284.0 | 24 | [106] |
| Solanum xanthocarpum Schrad. & J.C. Wendl. | Solanaceae | India | Fruit | Methanol | L1 | 170.91 | 320.62 | 24 | [114] |
| Methanol | L2 | 195.07 | 366.48 | 24 | [114] | ||||
| Methanol | L3 | 221.45 | 410.20 | 24 | [114] | ||||
| Methanol | L4 | 253.18 | 435.16 | 24 | [114] | ||||
| Spermacoce latifólia Aubl. | Rubiaceae | Brazil | Leaves | n-Hexane | L3 | 415.0 | 901.0 | 24 | [106] |
| Methanol | L3 | 625.0 | 1122.0 | 24 | [106] | ||||
| Tagetes patula L. | Asteraceae | Brazil | Seeds | Acetone | L4 | 15.74 | ND | 48 | [115] |
| Ethanol 50% | L4 | 25.46 | ND | 48 | [115] | ||||
| Turnera ulmifolia L. | Turneracea | Brazil | Leaves | Ethanol | L3 | 242.0 | 899.0 | 24 | [106] |
| Valeriana hardwickii Wall. | Valerianaceae | India | Leaves | Petroleum ether | L4 | 235.0 | 415.0 | ND | [83] |
| Ventilago madraspatana Gaertn. | Rhammnaceae | India | Leaves | Water + AgNP | L3 | 26.92 | ND | 24 | [112] |
| Water | L3 | 267.27 | ND | 24 | [112] | ||||
| Zeuxine gracilis (Berda) Bl. | Orchidaceae | India | Leaves | Water + AgNP | L3 | 10.39 | 23.58 | 24 | [116] |
| Plant Species | Family | Country | Part Used | Extraction Solvent | Activity | Results | Time (h) | Reference |
|---|---|---|---|---|---|---|---|---|
| Alpinia purpurata (Viell.) K. Schum. | Zingiberaceae | Brazil | Red Flowers | Water | Oviposition | Oviposition disruptive effect | 24 | [67] |
| Pink Flowers | Water | Oviposition | Oviposition disruptive effect | 24 | [67] | |||
| Aristolochia bracteata Retz. | Aristolochiaceae | India | Leaves | Methanol | Ovicide | Zero hatchability at 240 ppm | 48 | [81] |
| Methanol | Repellent | 100% of repellency at 6 mg/cm2 | 3 | [81] | ||||
| Artemisia herba-alba Asso | Asteraceae | Saudi Arabia | Leaves | Water | Adulticide—strain from India | LC50 327.15 µg/mL LC90 = 779.98 µg/mL | 24 | [82] |
| Water | Adulticide—strain from Saudi Arabia | LC50 450.21 µg/mL LC90 1153.18 µg/mL | 24 | [82] | ||||
| Water + AgNP | Adulticide—strain from India | LC50 8.71 µg/mL LC90 39.88 µg/mL | 24 | [82] | ||||
| Water + AgNP | Adulticide—strain from Saudi Arabia | LC50 25.62 µg/mL LC90 48.88 µg/mL | 24 | [82] | ||||
| Buddleja polystachya Fresen. | Buddlejaceae | Saudi Arabia | Flowers | n-Hexane | Adulticide | 96.7% mortality at 5 µg/mg female | ND | [84] |
| Ethanol | Adulticide | 83.3% mortality at 5 µg/mg female | ND | [84] | ||||
| Aerial parts | n-Hexane | Adulticide | 100% mortality at 5 µg/mg female | ND | [84] | |||
| Ethanol | Adulticide | 90% mortality at 5 µg/mg female | ND | [84] | ||||
| Caesalpinia pulcherrima (L.) Sw. | Fabaceae | India | Leaves | Methanol | Ovicide | Zero hatchability at 300 ppm | 48 | [85,117] |
| Methanol | Repellent | 100% of repellency at 5 mg/cm2 | 3 | [85,114] | ||||
| Ethyl acetate | Ovicide | Zero hatchability at 450 ppm | 48 | [85,117] | ||||
| Ethyl acetate | Repellent | 100% of repellency at 5 mg/cm2 | 1.5 | [85,117] | ||||
| Benzene | Ovicide | Zero hatchability at 375 ppm | 48 | [85,117] | ||||
| Benzene | Repellent | 100% of repellency at 5 mg/cm2 | 2 | [85,117] | ||||
| Cardiospermum halicacabum L. | Sapindaceae | India | Leaves | Methanol | Repellent | 100% of repellency at 5 mg/cm2 | 3 | [118] |
| n-Hexane | Repellent | 100% of repellency at 5 mg/cm2 | 3 | [118] | ||||
| Ethyl acetate | Repellent | 100% of repellency at 5 mg/cm2 | 3 | [118] | ||||
| Chloroform | Repellent | 100% of repellency at 5 mg/cm2 | 3 | [118] | ||||
| Benzene | Repellent | 100% of repellency at 5 mg/cm2 | 3 | [118] | ||||
| Cassia fistula L. | Fabaceae | India | Leaves | Methanol | Ovicide | Zero hatchability at 120 ppm | 48 | [86] |
| Methanol | Repellent | 100% of repellency at 5 mg/cm2 | 6 | [86] | ||||
| Benzene | Ovicide | Zero hatchability at 140 ppm | 48 | [86] | ||||
| Benzene | Repellent | 100% of repellency at 5 mg/cm2 | 5 | [86] | ||||
| Acetone | Ovicide | Zero hatchability at 160 ppm | 48 | [86] | ||||
| Acetone | Repellent | 100% of repellency at 5 mg/cm2 | 4.3 | [86] | ||||
| Cinnamosma fragrans Baill. | Canellaceae | Madagascar | Root barks | Methanol | Adulticide | LC50 0.17 µg/mg female | 24 | [87] |
| Methanol | Repellent | 80% of repellency at 20.8 µg/cm2 | 3 | [87] | ||||
| Coccinia indica Wight & Arn. | Cucurbitaceae | India | Leaves | Benzene | Ovicide | Zero hatchability at 250 ppm | 48 | [119] |
| Benzene | Repellent | 100% of repellency at 5 mg/cm2 | 3 | [119] | ||||
| n-Hexane | Ovicide | Zero hatchability at 300 ppm | 48 | [119] | ||||
| n-Hexane | Repellent | 100% of repellency at 1 mg/cm2 | 2.5 | [119] | ||||
| Ethyl acetate | Ovicide | Zero hatchability at 250 ppm | 48 | [119] | ||||
| Ethyl acetate | Repellent | 100% of repellency at 2.5 mg/cm2 | 2.5 | [119] | ||||
| Methanol | Ovicide | Zero hatchability at 200 ppm | 48 | [119] | ||||
| Methanol | Repellent | 100% of repellency at 5 mg/cm2 | 3.5 | [119] | ||||
| Chloroform | Ovicide | Zero hatchability at 250 ppm | 48 | [119] | ||||
| Chloroform | Repellent | 100% of repellency at 2.5 mg/cm2 | 2.5 | [119] | ||||
| Eclipta alba (L.) Hassk | Asteraceae | India | Leaves | Benzene | Ovicide | Zero hatchability at 350 ppm | 48 | [119] |
| n-Hexane | Ovicide | 21% hatchability at 350 ppm | 48 | [119] | ||||
| Ethyl acetate | Ovicide | Zero hatchability at 350 ppm | 48 | [119] | ||||
| Methanol | Ovicide | Zero hatchability at 300 ppm | 48 | [91] | ||||
| Chloroform | Ovicide | Zero hatchability at 350 ppm | 48 | [119] | ||||
| Ervatamia coronaria (Jacq.) Stapf. | Apocynaceae | India | Leaves | Methanol | Ovicide | Zero hatchability at 200 ppm | 48 | [117] |
| Methanol | Repellent | 100% of repellency at 5 mg/cm2 | 3 | [117] | ||||
| Ethyl acetate | Ovicide | Zero hatchability at 300 ppm | 48 | [117] | ||||
| Ethyl acetate | Repellent | 100% of repellency at 5 mg/cm2 | 2 | [117] | ||||
| Benzene | Ovicide | Zero hatchability at 250 ppm | 48 | [117] | ||||
| Benzene | Repellent | 100% of repellency at 5 mg/cm2 | 2.5 | [117] | ||||
| Limonia acidíssima L. | Rutaceae | India | Leaves | n-Hexane—Purified fractions | Ovicide | 78.4 hatchability at 10 ppm | 120 | [99] |
| n-Hexane—Purified fractions | Pupicide | LC50 4.19—39.48 µg/mL | 24 | [99] | ||||
| Mentha piperita L. | Lamiaceae | India | Whole plant | Methanol | Repellent | Repellency | ND | [120] |
| Millettia pachycarpa Benth. | Fabaceae | India | Root | Ethanol | Ovicide | Zero hatchability at 200 ppm | 24 | [102] |
| Myristica fragans Houtt. | Myristicaceae | India | Leaves | Methanol | Pupicide | LC50 359.08 µg/mL LC90 803.52 µg/mL | 24 | [53] |
| Methanol + ZnONP | Pupicide | LC50 14.63 µg/mL LC90 51.22 µg/mL | 24 | [53] | ||||
| Methanol | Adulticide | LC50 180.26 µg/mL LC90 368.93 µg/mL | 24 | [53] | ||||
| Methanol + ZnONP | Adulticide | LC50 15.0 µg/mL LC90 34.2 µg/mL | 24 | [53] | ||||
| Nerine sarniensis (L.) Herb. | Amaryllidaceae | South Africa | Bulbs | Ehtyl acetate | Adulticide | LC50 4.6 µg/mg female | 24 | [104] |
| Parthenium hysterophorus | Asteraceae | India | Leaves | Ether | Repellent | 99.6% of repellency at 1000 ppm | 48 | [121] |
| Ether | Ovicide | Zero hatchability at 1000 ppm | 48 | [121] | ||||
| Benzene | Repellent | 93.8% of repellency at 1000 ppm | 48 | [121] | ||||
| Benzene | Ovicide | Zero hatchability at 1000 ppm | 48 | [121] | ||||
| Pemphis acidula J.R. Forst. & G. Forst. | Lythraceae | India | Leaves | Methanol | Ovicide | Zero hatchability at 450 ppm | 48 | [108] |
| Acetone | Ovicide | Zero hatchability at 500 ppm | 48 | [108] | ||||
| Pithecellobium Dulce (Roxb.) Benth. | Fabaceae | India | Leaves | Methanol | Ovicide | Zero hatchability at 400 ppm | 48 | [111] |
| Ehtyl acetate | Ovicide | Zero hatchability at 500 ppm | 48 | [111] | ||||
| Chloroform | Ovicide | Zero hatchability at 500 ppm | 48 | [111] | ||||
| Benzene | Ovicide | Zero hatchability at 600 ppm | 48 | [111] | ||||
| n-Hexane | Ovicide | Zero hatchability at 600 ppm | 48 | [111] | ||||
| Seeds | Methanol | Ovicide | Zero hatchability at 625 ppm | 48 | [111] | |||
| Ethyl acetate | Ovicide | Zero hatchability at 750 ppm | 48 | [111] | ||||
| Chloroform | Ovicide | Zero hatchability at 750 ppm | 48 | [111] | ||||
| Benzene | Ovicide | Zero hatchability at 750 ppm | 48 | [111] | ||||
| Solanum xanthocarpum Schrad. & J.C. Wendl. | Solanaceae | India | Fruit | Methanol | Pupicide | LC50 279.52 µg/mL LC90 462.10 µg/mL | 24 | [114] |
| Ventilago madraspatana Gaertn. | Rhammnaceae | India | Leaves | Water + AgNP | Ovicide | Zero hatchability at 120 ppm | 48 | [112] |
| Water | Ovicide | Zero hatchability at 400 ppm | 48 | [112] | ||||
| Water + AgNP | Adulticide | LC50 44.85 µg/mL | 24 | [112] | ||||
| Water | Adulticide | LC50 334.46 µg/mL | 24 | [112] | ||||
| Zeuxine gracilis (Berda) Bl. | Orchidaceae | India | Leaves | Water + AgNP | Ovicide | Zero hatchability at 12 ppm | 48 | [116] |
| Adulticide | LC50 27.90 µg/mL LC90 = 59.20 µg/mL | 24 | [116] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silvério, M.R.S.; Espindola, L.S.; Lopes, N.P.; Vieira, P.C. Plant Natural Products for the Control of Aedes aegypti: The Main Vector of Important Arboviruses. Molecules 2020, 25, 3484. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25153484
Silvério MRS, Espindola LS, Lopes NP, Vieira PC. Plant Natural Products for the Control of Aedes aegypti: The Main Vector of Important Arboviruses. Molecules. 2020; 25(15):3484. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25153484
Chicago/Turabian StyleSilvério, Maíra Rosato Silveiral, Laila Salmen Espindola, Norberto Peporine Lopes, and Paulo Cézar Vieira. 2020. "Plant Natural Products for the Control of Aedes aegypti: The Main Vector of Important Arboviruses" Molecules 25, no. 15: 3484. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25153484