
Composition of Primary and Secondary Metabolite Compounds in Seeds and Pods of Asparagus Bean (Vigna unguiculata (L.) Walp.) from China

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Total Protein Content
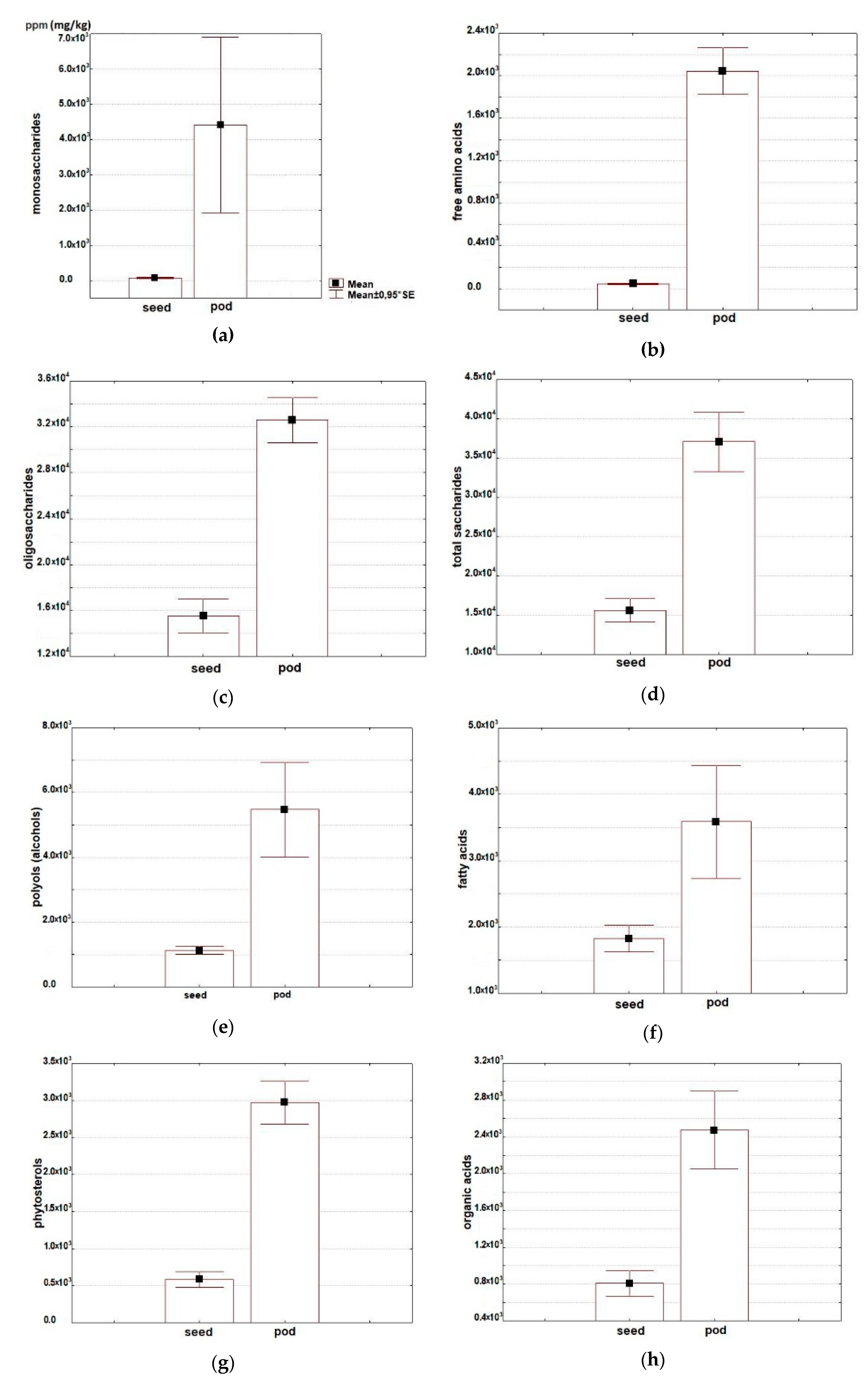
2.2. Analysis of Metabolomic Profiles in Seeds and Pods
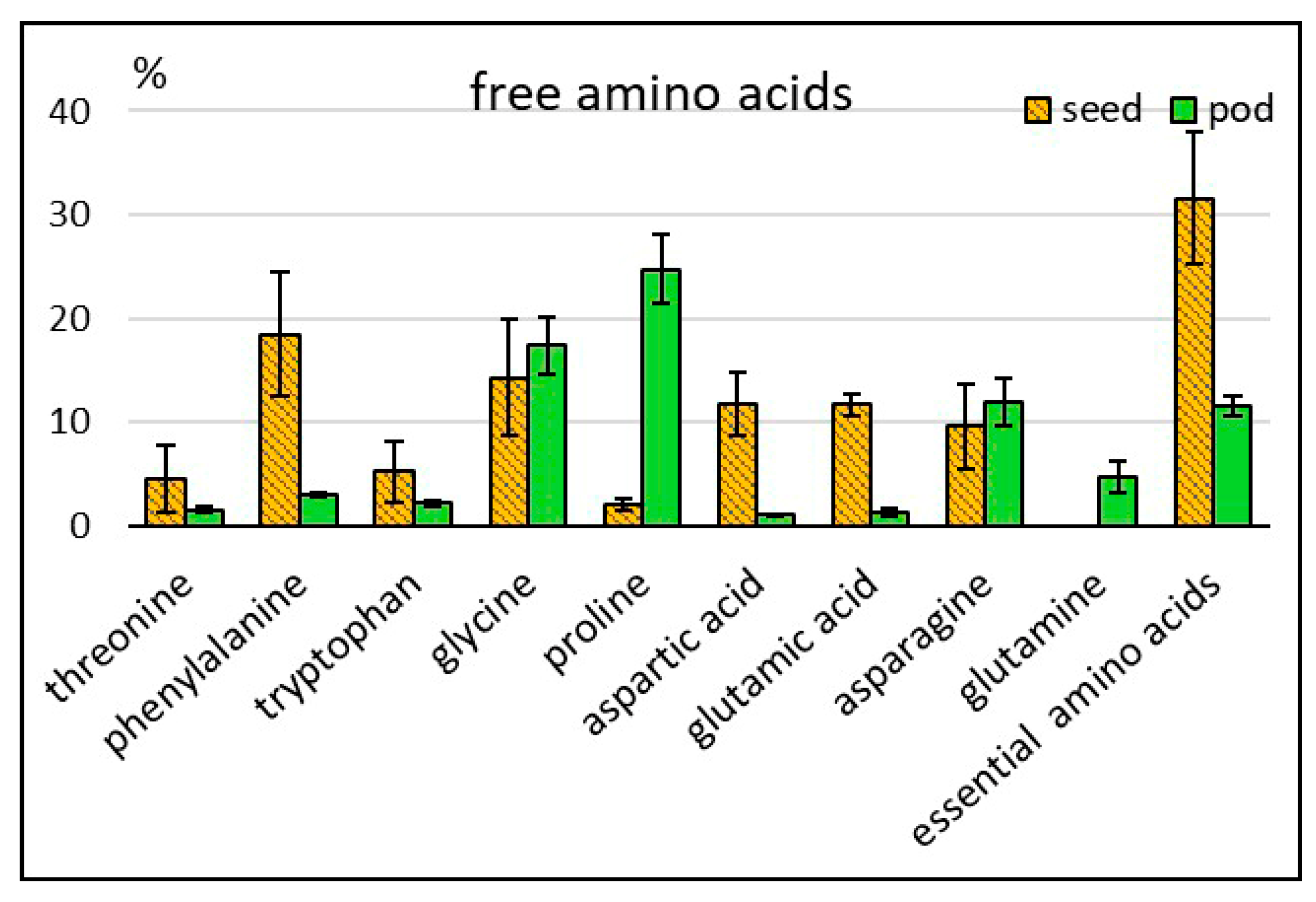
2.2.1. Free Amino Acids
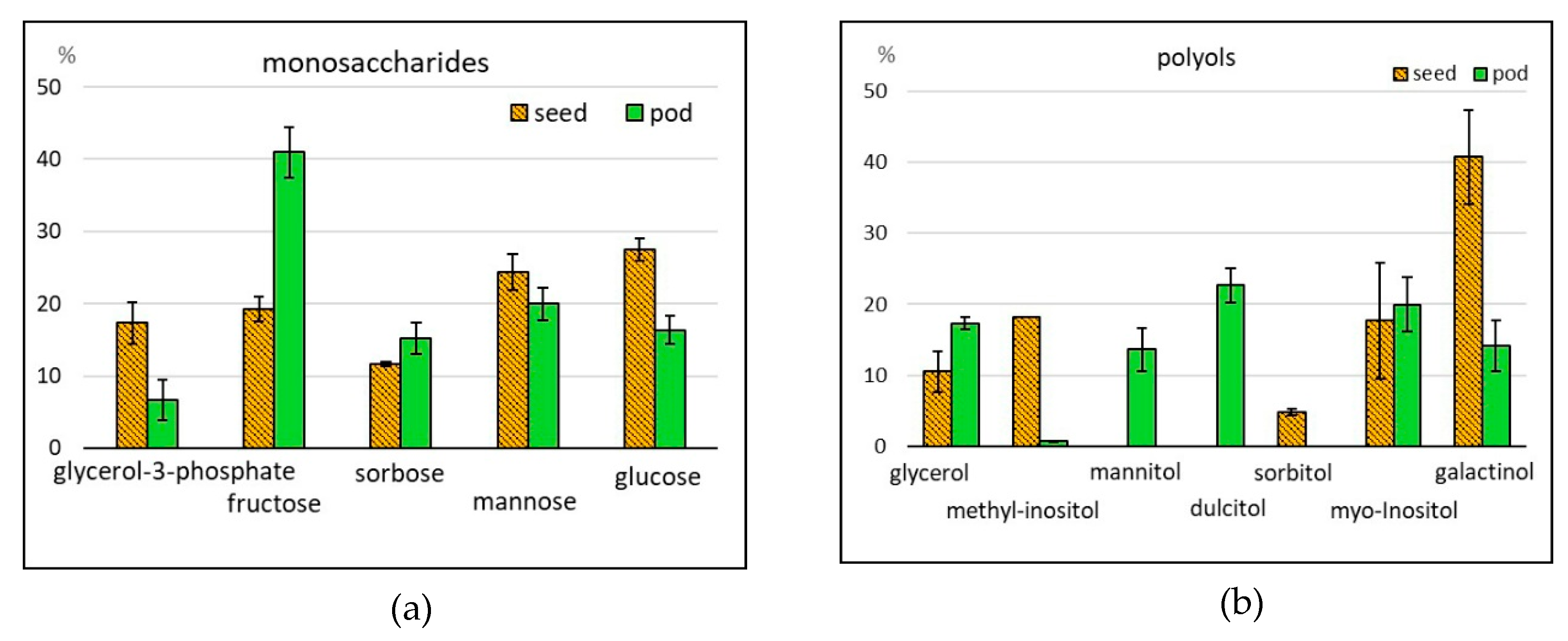
2.2.2. Saccharides
2.2.3. Alcohols
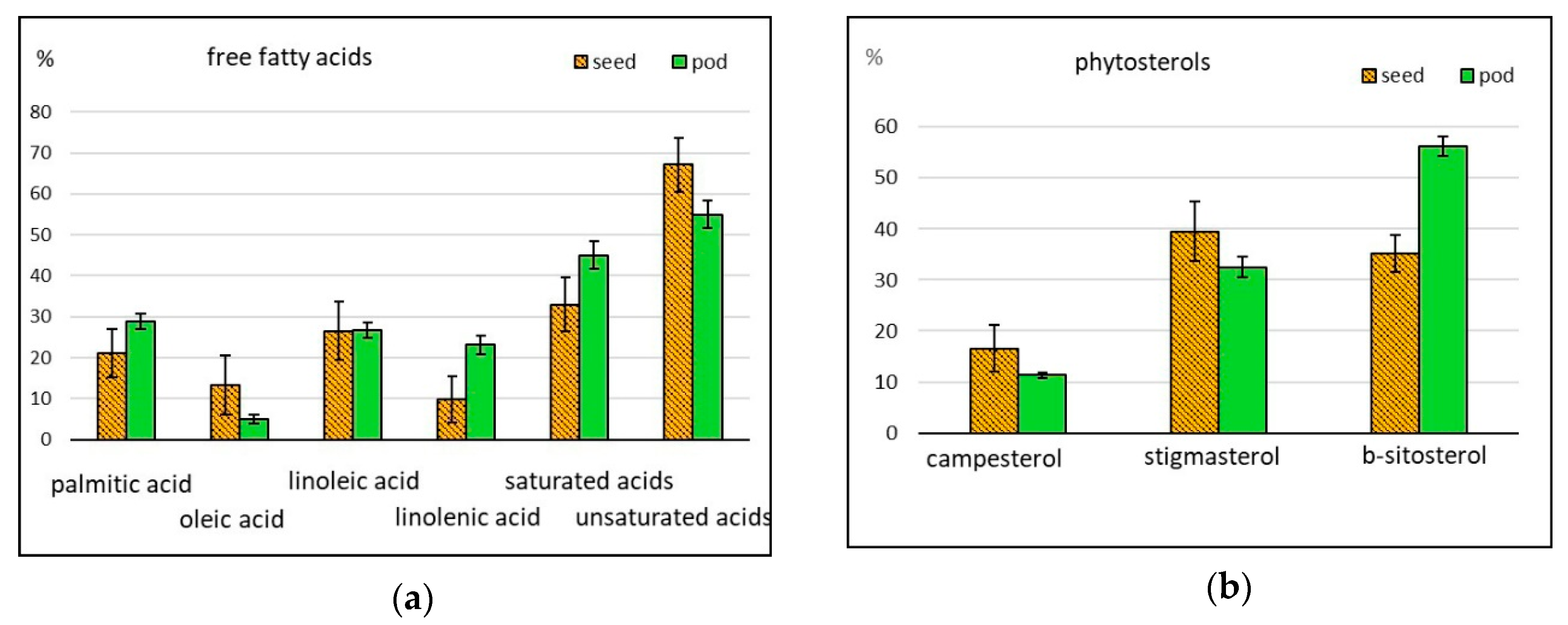
2.2.4. Free Fatty Acids
2.2.5. Phytosterols
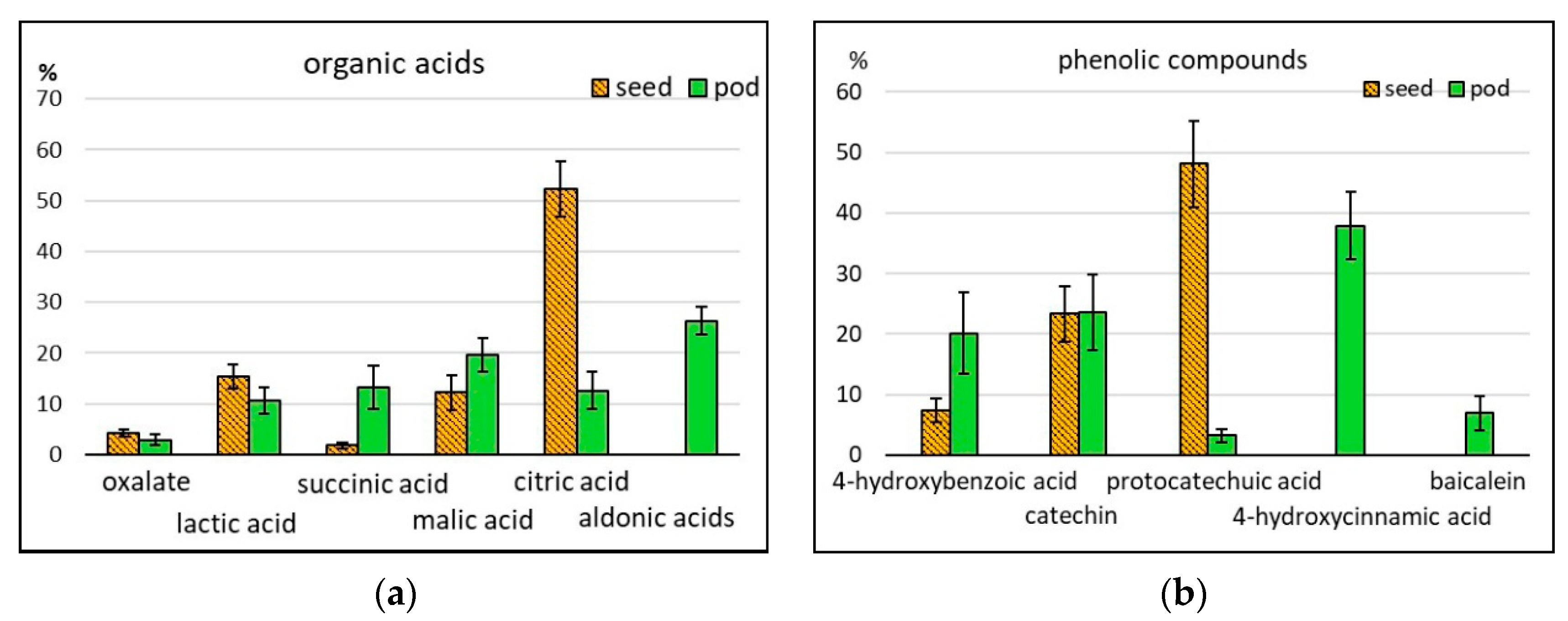
2.2.6. Organic Acids
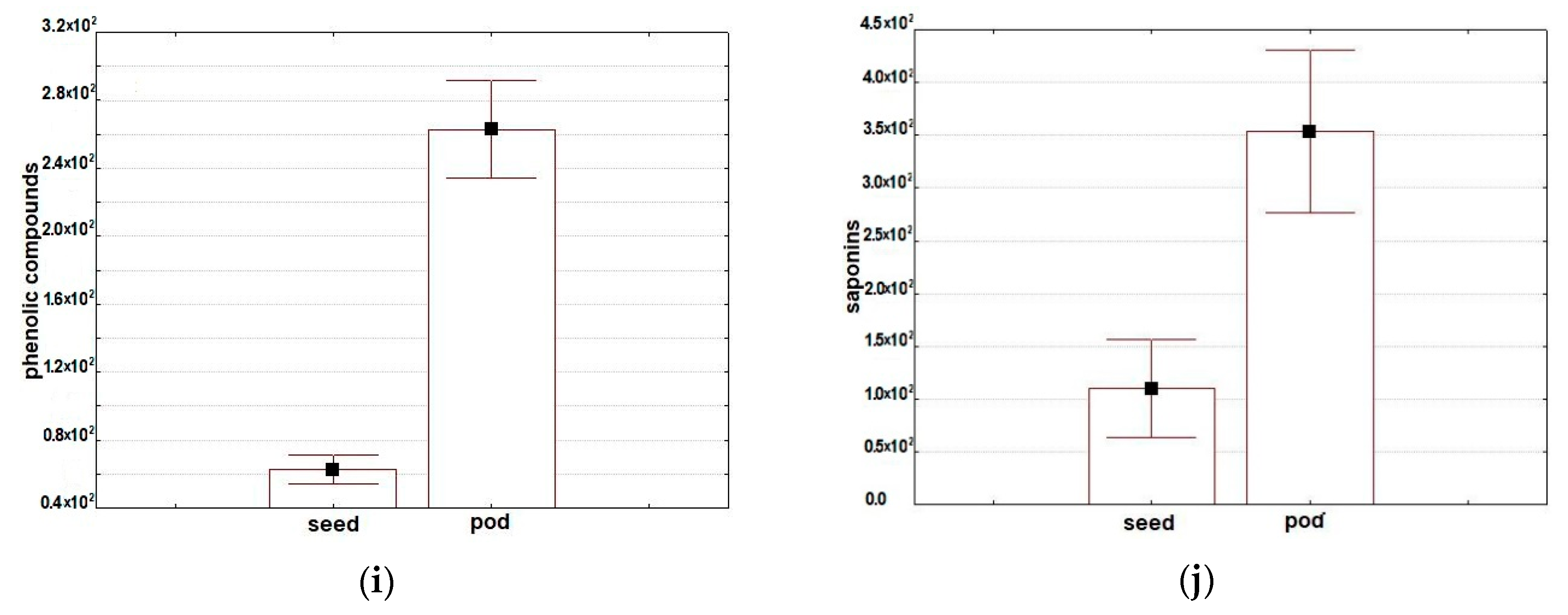
2.2.7. Phenolic Compounds
2.2.8. Saponins
2.2.9. Other Compounds
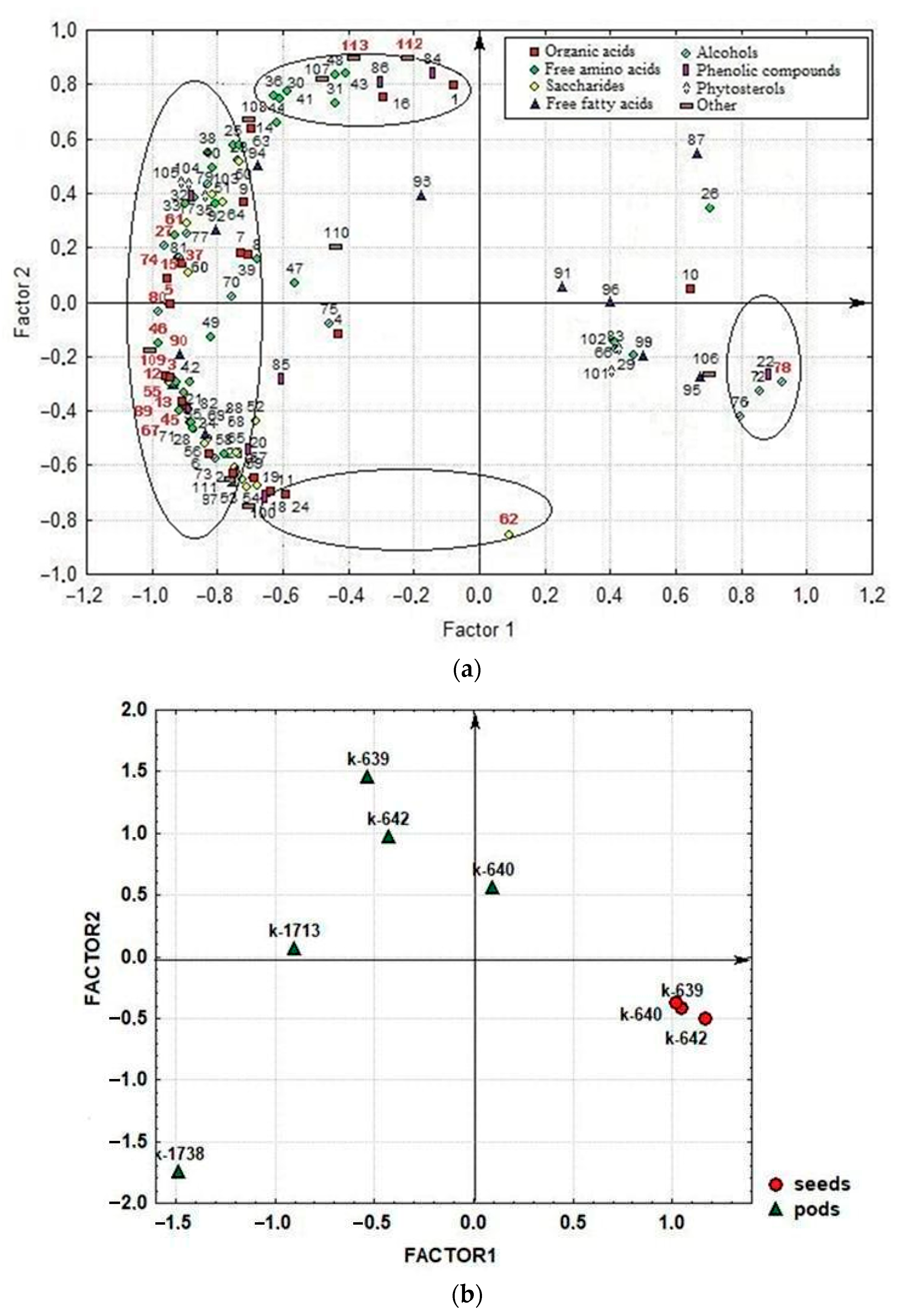
2.3. Results of the Factor Analysis
3. Materials and Methods
3.1. Materials
3.2. Chemicals and Reagents
3.3. Protein Determination
3.4. GC-MS Analysis
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Vavilov, N.I. Botanical and Geographical Fundamentals of Plant Breeding; USSR Academy of Sciences: Moscow/Leningrad, Russia, 1935; p. 60. (In Russian) [Google Scholar]
- FAO. FAO Departments and Offices. 2019. Available online: http://www.fao.org/faostat/ru/ (accessed on 21 May 2020).
- Timko, M.P.; Ehlers, J.D.; Roberts, P.A. Cowpea. In Genome Mapping and Molecular Breeding in Plants. Pulses. Sugar and Tuber Crops; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; Volume 3, pp. 49–67. [Google Scholar]
- Awika, J.M.; Duodu, K.G. Bioactive polyphenols and peptides in cowpea (Vigna unguiculata) and their health promoting properties: A review. J. Funct. Foods 2017, 38, 686–697. [Google Scholar] [CrossRef]
- Marques, M.R.; Freitas, R.A.M.S.; Carlos, A.C.C.; Siguemoto, E.S.; Fontanari, G.G.; Arкas, J.A.G. Peptides from cowpea present antioxidant activity. Inhibit cholesterol synthesis and its solubilisation into micelles. Food Chem. 2015, 168, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.; Goufo, P.; Domínguez-Perles, R.; Trindade, H.; Rosa, E.A.; Ferreira, L.; Rodrigues, M. Cowpea (Vigna unguiculata L. Walp) a renewed multipurpose crop for a more sustainable agri-food system: Nutritional advantages and constraints. J. Sci. Food Agric. 2016, 96, 2941–2951. [Google Scholar] [CrossRef]
- Belane, A.K.; Dakora, F.D. Elevated concentrations of dietarily-important trace elements and macronutrients in edible leaves and grain of 27 cowpea (Vigna unguiculata L. Walp) genotypes: Implications for human nutrition and health. Food Nutr. Sci. 2012, 3, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, A.; Khalil, I.A.; Ateeq, N.; Khan, M.S. Nutritional quality of important food legumes. Food Chem. 2006, 97, 331–335. [Google Scholar] [CrossRef]
- Bessada, S.M.F.; Barreira, J.C.M.; Oliveira, M.B.P.P. Pulses and food security: Dietary protein, digestibility, bioactive and functional properties. Trends Food Sci. Tech. 2019, 93, 53–68. [Google Scholar] [CrossRef]
- Shevkani, K.; Kaur, A.; Kumar, S.; Singh, N. Cowpea protein isolates: Functional properties and application in gluten-free rice muffins. LWT-Food Sci. Technol. 2015, 63, 927–933. [Google Scholar] [CrossRef]
- Phillips, R.D.; McWatters, K.H.; Chinnan, M.S.; Hung, Y.-C.; Beuchat, L.R.; Sefa-Dedeh, S.; Sakyi-Dawson, E.; Ngoddy, P.; Nnanyelugo, D.; Enwere, J.; et al. Utilization of cowpeas for human food. Field Crop. Res. 2003, 82, 193–213. [Google Scholar] [CrossRef]
- Van Breemen, R.B. Role of mass spectrometry in establishing safety and efficacy of botanical dietary supplements. Clin. Mass Spect. 2020, 15, 25–28. [Google Scholar] [CrossRef]
- Onuha, J.O.; Alukob, R.E. Metabolomics as a tool to study the mechanism of action of bioactive protein hydrolysates and peptides: A review of current literature. Trends Food Sci. Tech. 2019, 91, 625–633. [Google Scholar] [CrossRef]
- Morris, J.B.; Tonnis, B.D.; Wang, M.L. Protein content and seed trait analysis in a subset of the USDA, ARS, PGRCU cowpea [Vigna unguiculata (L.) Walp] core collection. Leg. Res.-Int. J. 2020, LR-529. [Google Scholar] [CrossRef]
- Zhuzhukin, V.I.; Gorbunov, V.S.; Bagdalova, A.Z. Study of biochemical composition of seeds and green beans of vegetable cowpea (Vigna Ung. ssp. Sesquipedalis). Rus. Agric. Sci. 2017, 43, 390–394. [Google Scholar] [CrossRef]
- Gupta, P.; Singh, R.; Malhotra, S.; Boora, K.S.; Singal, H.R. Characterization of seed storage proteins in high protein genotypes of cowpea [Vigna unguiculata (L.) Walp]. Physiol. Mol. Biol. Plants. 2010, 16, 53–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, E.S.; Capraro, J.; Sessa, F.; Magni, C.; Demonte, A.; Consonni, A.; Neves, V.A.; Cilli, E.M.; Marcello Duranti, M.; Scarafoni, A. New molecular features of cowpea bean (Vigna unguiculata. l. Walp) β-vignin. Biosci. Biotech. Biochem. 2018, 82, 285–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasconcelos, I.M.; Maia, F.M.M.; Farias, D.F.; Campello, C.C.; Carvalho, A.F.U.; Moreira, R.A.; de Oliveira, J.T.A. Protein fractions. Aamino acid composition and antinutritional constituents of high-yielding cowpea cultivars. J. Food Comp. Anal. 2010, 23, 54–60. [Google Scholar] [CrossRef]
- Frota, K.M.G.; Mendonca, S.; Saldiva, P.H.N.; Cruz, R.J.; Areas, J.A.G. Cholesterol-lowering properties of whole cowpea seed and its protein isolate in hamsters. J. Food Sci. 2008, 73, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Marques, M.R.; Fontanari, G.G.; Pimenta, D.C.; Freitas, R.A.M.S.; Areas, J.A.G. Proteolytic hydrolysis of cowpea proteins is able to release peptides with hypocholesterolemic activity. Food Res. Inter. 2015, 77, 43–48. [Google Scholar] [CrossRef]
- Castaneda-Perez, E.; Jimenez-Morales, K.; Quintal-Novelo, C.; Moo-Puc, R.; Chel-Guerrero, L.; Betancur-Ancona, D. Enzymatic protein hydrolysates and ultrafiltered peptide fractions from cowpea Vigna unguiculata L. bean with in vitro antidiabetic potential. J. Iran. Chem. Soc. 2019, 16, 1773–1781. [Google Scholar] [CrossRef]
- De Souza Rocha, T.S.; Hernandez, L.M.R.; Chang, Y.K.; González de Mejía, C. Impact of germination and enzymatic hydrolysis of cowpea bean (Vigna unguiculata) on the generation of peptides capable of inhibiting dipeptidyl peptidase IV. Food Res. Inter. 2014, 64, 799–809. [Google Scholar] [CrossRef]
- Abdel-Shafi, S.; Al-Mohammadi, A.-R.; Osman, A.; Gamal Enan, G.; Abdel-Hameid, S.; Sitohy, M. Characterization and antibacterial activity of 7S and 11S globulins isolated from cowpea seed protein. Molecules 2019, 24, 1082. [Google Scholar] [CrossRef] [Green Version]
- Elhardallou, S.B.; Khalid, I.I.; Gobouri, A.A.; Abdel-Hafez, S.H. Amino acid composition of cowpea (Vigna ungiculata L. Walp) flour and its protein isolates. Food Nutr. Sci. 2015, 6, 790–797. [Google Scholar] [CrossRef] [Green Version]
- Ibragimova, S.S.; Gorelova, V.V.; Kochetov, A.V.; Shumny, V.K. Role of plant metabolites in mechanisms of stress tolerance. Bull. Novosib. State Univeristy Ser. Biol. Clin. Med. 2010, 8, 98–103. (In Russian) [Google Scholar]
- Liu, W.R.; Lim, G.-H.; de Lorenzo, L.; Yu, K.; Zhang, K.; Hunt, A.G.; Kachroo, A.; Kachroo, P. Pipecolic acid confers systemic immunity by regulating free radicals. Sci. Adv. 2018, 4, eaar4509. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Jander, G.; de Vos, M. Non-protein amino acids in plant defense against insect herbivores: Representative cases and opportunities for further functional analysis. Phytochemistry 2011, 72, 1531–1537. [Google Scholar] [CrossRef]
- Deol, J.K.; Bains, K. Effect of household cooking methods on nutritional and antinutritional factors in green cowpea (Vigna unguiculata) pods. J. Food Sci. Tech. 2010, 47, 579–581. [Google Scholar] [CrossRef] [Green Version]
- Amadioha, A.C.; Nwazuo, E.D. Biochemical composition of seed and husk of cowpea (Vigna unguiculata (L.) Walp) infected by Colletotrichum destructivum O’Gara in storage. Ann. Res. Rev. Biol. 2019, 31, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Onyenekwe, P.C.; Njoku, G.C.; Ameh, D.A. Effect of cowpea (Vigna unguiculata) processing methods on flatus causing oligosaccharides. Nutr. Res. 2000, 20, 349–358. [Google Scholar] [CrossRef]
- Egounlety, M.; Aworh, O.C. Effect of soaking, dehulling, cooking and fermentation with Rhizopus oligosporus on the oligosaccharides, trypsin inhibitor, phytic acid and tannins of soybean (Glycine max Merr.), cowpea (Vigna unguiculata L. Walp) and groundbean (Macrotyloma geocarpa Harms). J. Food Eng. 2003, 56, 249–254. [Google Scholar]
- Dumschott, K.; Richter, A.; Loescher, W.; Merchant, A. Post photosynthetic carbon partitioning to sugar alcohols and consequences for plant growth. Phytochemistry 2017, 144, 243–252. [Google Scholar] [CrossRef]
- Gangola, M.P.; Jaiswal, S.; Kannan, U.; Gaur, P.M.; Båga, M.; Chibbar, R.N. Galactinol synthase enzyme activity influences raffinose family oligosaccharides (RFO) accumulation in developing chickpea (Cicer arietinum L.) seeds. Phytochemistry 2016, 125, 88–98. [Google Scholar] [CrossRef]
- Limanova, O.A.; Gromova, O.A.; Torshin, I.Y.; Gromov, A.N.; Grishina, N.R. Systematic analysis of molecular mechanisms and physiological effects of myo-inositol: Findings of molecular biology. Experimental and clinical medicine. Eff. Pharm. Obs. Gynecol. 2013, 3, 32–41. [Google Scholar]
- Kalpanadevi, V.; Mohan, V.R. Effect of processing on antinutrients and in vitro protein digestibility of the underutilized legume. Vigna unguiculata (L.) Walp subsp. unguiculata. Lwt Food Sci. Technol. 2013, 51, 455–461. [Google Scholar] [CrossRef]
- Devi1, C.B.; Kushwaha, A.; Kumar, A. Sprouting characteristics and associated changes in nutritional composition of cowpea (Vigna unguiculata). J. Food Sci. Tech. 2015, 52, 6821–6827. [Google Scholar] [CrossRef]
- Kumar, V.; Sinha, A.K.; Makkar, H.P.S.; Becker, K. Dietary roles of phytate and phytase in human nutrition: A review. Food Chem. 2010, 120, 945–959. [Google Scholar] [CrossRef]
- Antova, G.A.; Stoilova, T.D.; Ivanova, M.M. Proximate and lipid composition of cowpea (Vigna unguiculata L.) cultivated in Bulgaria. J. Food Comp. Anal. 2014, 33, 146–152. [Google Scholar] [CrossRef]
- Liu, X.; Ma, D.; Zhang, Z.; Wang, S.; Du, S.; Deng, X.; Yin, L. Plant lipid remodeling in response to abiotic stresses. Env. Exp. Bot. 2019, 165, 174–184. [Google Scholar] [CrossRef]
- Sharova, E.I. Plant Antioxidants; St. Petersburg State University: St. Petersburg, Russia, 2016; p. 140. (In Russian) [Google Scholar]
- Karateev, A.E.; Aleinikova, T.L. Eicosanoids and inflammation. Mod. Rheumatol. J. 2016, 10, 73–86. [Google Scholar] [CrossRef] [Green Version]
- Plat, J.; Baumgartner, S.; Vanmierlo, T.; Lutjohann, D.; Calkins, K.L.; Burrin, D.G.; Guthrie, G.; Thijs, C.; Te Velde, A.A.; Vreugdenhil, A.C.E.; et al. Plant-based sterols and stanols in health & disease: “Consequences of human development in a plant-based environment?”. Prog. Lipid Res. 2019, 74, 87–102. [Google Scholar] [CrossRef]
- Shahzad, N.; Khan, W.; Shadab, M.D.; Ali, A.; Saluja, S.S.; Sharma, S.; Al-Allaf, F.A.; Abduljaleel, Z.; Ibrahim, I.A.A.; Abdel-Wahab, A.F.; et al. Phytosterols as a natural anticancer agent: Current status and future perspective. Biomed. Pharm. 2017, 88, 786–794. [Google Scholar] [CrossRef]
- Borisova, G.G. (Ed.) Fundamentals of Secondary Metabolism Biochemistry in Plants; Ural University: Yekaterinburg, Russia, 2014; p. 128. (In Russian) [Google Scholar]
- Corso, M.; Perreau, F.; Mouille, G.; Lepiniec, L. Specialized phenolic compounds in seeds: Structures, functions, and regulations. Plant Sci. 2020, 296, 110471. [Google Scholar] [CrossRef]
- Azmir, J.; Zaidul, I.S.M.; Rahman, M.M.; Sharif, K.M.; Mohamed, A.; Sahena, F.; Jahurul, M.H.A.; Ghafoor, K.; Norulaini, N.A.N.; Omar, A.K.M. Techniques for extraction of bioactive compounds from plant materials: A review. J. Food Eng. 2013, 117, 426–436. [Google Scholar] [CrossRef]
- Zia-Ul-Haq, M.; Ahmad, S.; Amarowicz, R.; De Feo, V. Antioxidant activity of the extracts of some cowpea (Vigna unguiculata (L) Walp) cultivars commonly consumed in Pakistan. Molecules 2013, 18, 2005–2017. [Google Scholar] [CrossRef] [PubMed]
- Khang, D.T.; Dung, T.N.; Elzaawely, A.A.; Xuan, T.D. Phenolic profiles and antioxidant activity of germinated legumes. Foods 2016, 5, 27. [Google Scholar] [CrossRef]
- Gutie’rrez-Uribe, J.A.; Romo-Lopez, I.; Serna-Saldıvar, S.O. Phenolic composition and mammary cancer cell inhibition of extracts of whole cowpeas (Vigna unguiculata) and its anatomical parts. J. Funct. Foods 2011, 3, 290–297. [Google Scholar] [CrossRef]
- Teixeira-Guedes, C.I.; Oppolzer, D.; Barros, A.I.; Pereira-Wilson, C. Phenolic rich extracts from cowpea sprouts decrease cell proliferation and enhance 5-fluorouracil effect in human colorectal cancer cell line. J. Funct. Foods 2019, 60, 103452. [Google Scholar] [CrossRef]
- Nderitu, A.M.; Dykes, L.; Awika, J.M.; Minnaar, A.; Duodu, K.G. Phenolic composition and inhibitory effect against oxidative DNA damage of cooked cowpeas as affected by simulated in vitro gastrointestinal digestion. Food Chem. 2013, 141, 1763–1771. [Google Scholar] [CrossRef]
- Fan, F.-Y.; Sang, L.-X.; Jiang, M. Catechins and their therapeutic benefits to inflammatory bowel disease. Molecules 2017, 22, 484. [Google Scholar] [CrossRef] [Green Version]
- Procházková, D.; Boušová, I.; Wilhelmová, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar] [CrossRef]
- Ojwang, L.O.; Yang, L.; Dykes, L.; Awika, J. Proanthocyanidin profile of cowpea (Vigna unguiculata) reveals catechin-O-glucoside as the dominant compound. Food Chem. 2013, 139, 35–43. [Google Scholar] [CrossRef]
- Muzquiz, M.; Varela, A.; Burbano, C.; Cuadrado, C.; Guillamón, E.; Pedrosa, M.M. Bioactive compounds in legumes: Pronutritive and antinutritive actions. Implications for nutrition and health. Phytochem. Rev. 2012, 11, 227–244. [Google Scholar] [CrossRef]
- Kretovich, V.L. Biochemistry of Plants; Vysshaya shkola: Moscow, Russia, 1986; p. 503. (in Russian) [Google Scholar]
- Ermakov, A.I. (Ed.) Methods of Biochemical Research of Plants, 3rd ed.; Agropromizdat: Leningrad, Russia, 1987; p. 430. (In Russian) [Google Scholar]
- Loskutov, I.G.; Shelenga, T.V.; Konarev, A.V.; Vargach, Y.I.; Porokhovinova, E.A.; Blinova, E.V.; Gnutikov, A.A.; Rodionov, A.V. Modern approach of structuring the varietal diversity of the naked and covered forms of cultural oats (Avena sativa L.). Ecol. Genet. 2020, 18, 27–41. (In Russian) [Google Scholar] [CrossRef]
- Shtark, O.Y.; Puzanskiy, R.K.; Avdeeva, G.S.; Yurkov, A.P.; Smolikova, G.N.; Yemelyanov, V.V.; Kliukova, M.S.; Shavarda, A.L.; Kirpichnikova, A.A.; Zhernakov, A.I.; et al. Metabolic alterations in pea leaves during arbuscular mycorrhiza development. Peer J. 2019, 7, 1–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puzanskiy, R.; Tarakhovskaya, E.; Shavarda, A.; Shishova, M. Metabolomic and physiological changes of Chlamydomonas reinhardtii (Chlorophyceae, Chlorophyta) during batch culture development. J. Appl. Phycol. 2018, 30, 803–818. [Google Scholar] [CrossRef]
- StatSoft Inc. Electronic Statistics Textbook; StatSoft: Tulsa, OK, USA, 2013; Available online: http://www.statsoft.com/textbook/ (accessed on 4 April 2020).
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | Seeds | Pods | ||
|---|---|---|---|---|
| Mean ± SE | Min–Max | Mean ± SE | Min–Max | |
| Protein* | 27.10 ± 0.22 | 26.75–27.51 | 29.98 ± 1.10 | 26.90–33.50 |
| Free amino acids** | 433 ± 69 | 340–567 | 20,433 ± 2310 | 14,016–25,727 |
| Total saccharides** | 15,596 ± 1574 | 12,720–18,142 | 37,072 ± 4032 | 27,331–50,046 |
| Oligosaccharides** | 15,522 ± 1568 | 12,669–18,075 | 32,602 ± 2066 | 26,500–38,462 |
| Monosaccharides** | 74 ± 16 | 51–105 | 4409 ± 2625 | 807–14,764 |
| Polyols** | 1127 ± 125 | 897–1327 | 5678 ± 1557 | 2822–11,110 |
| Free fatty acids** | 1827 ± 213 | 1472–2209 | 3588 ± 897 | 1992–6807 |
| Phytosterols** | 584 ± 113 | 400–790 | 2972 ± 307 | 1954–3785 |
| Organic acids** | 808 ± 143 | 634–1093 | 2474 ± 447 | 1495–3867 |
| Phenolic compounds** | 63 ± 9 | 48–79 | 263 ± 30 | 193–364 |
| Saponins** | 110 ± 48 | 43–204 | 354 ± 81 | 212–643 |
| Phosphoric acids** | 2407 ± 240 | 2127–2885 | 1667 ± 236 | 1037–2274 |
| Urea** | 289 ± 15 | 271–319 | 446 ± 102 | 181–780 |
| No. | VIR Catalogue Number | Accession Name | Origin | Year of Acquisition | Comments |
|---|---|---|---|---|---|
| 1 | k-639 | Landrace | China | 1929 | N.I. Vavilov’s expedition to China |
| 2 | k-640 | Landrace | China | 1929 | N.I. Vavilov’s expedition to China |
| 3 | k-642 | Landrace | China | 1929 | N.I. Vavilov’s expedition to China |
| 4 | k-1713 | Cv. ‘Yunanskaya’ | Novosibirsk Province, Russia | 2006 | |
| 5 | k-1738 | Cv. ‘Sibirskiy razmer’ | Novosibirsk Province, Russia | 2006 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perchuk, I.; Shelenga, T.; Gurkina, M.; Miroshnichenko, E.; Burlyaeva, M. Composition of Primary and Secondary Metabolite Compounds in Seeds and Pods of Asparagus Bean (Vigna unguiculata (L.) Walp.) from China. Molecules 2020, 25, 3778. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25173778
Perchuk I, Shelenga T, Gurkina M, Miroshnichenko E, Burlyaeva M. Composition of Primary and Secondary Metabolite Compounds in Seeds and Pods of Asparagus Bean (Vigna unguiculata (L.) Walp.) from China. Molecules. 2020; 25(17):3778. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25173778
Chicago/Turabian StylePerchuk, Irina, Tatyana Shelenga, Maria Gurkina, Elena Miroshnichenko, and Marina Burlyaeva. 2020. "Composition of Primary and Secondary Metabolite Compounds in Seeds and Pods of Asparagus Bean (Vigna unguiculata (L.) Walp.) from China" Molecules 25, no. 17: 3778. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25173778