
Biochemical and Computational Approach of Selected Phytocompounds from Tinospora crispa in the Management of COVID-19

 ,
,  ,
,  ,
,  ,
, 
 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
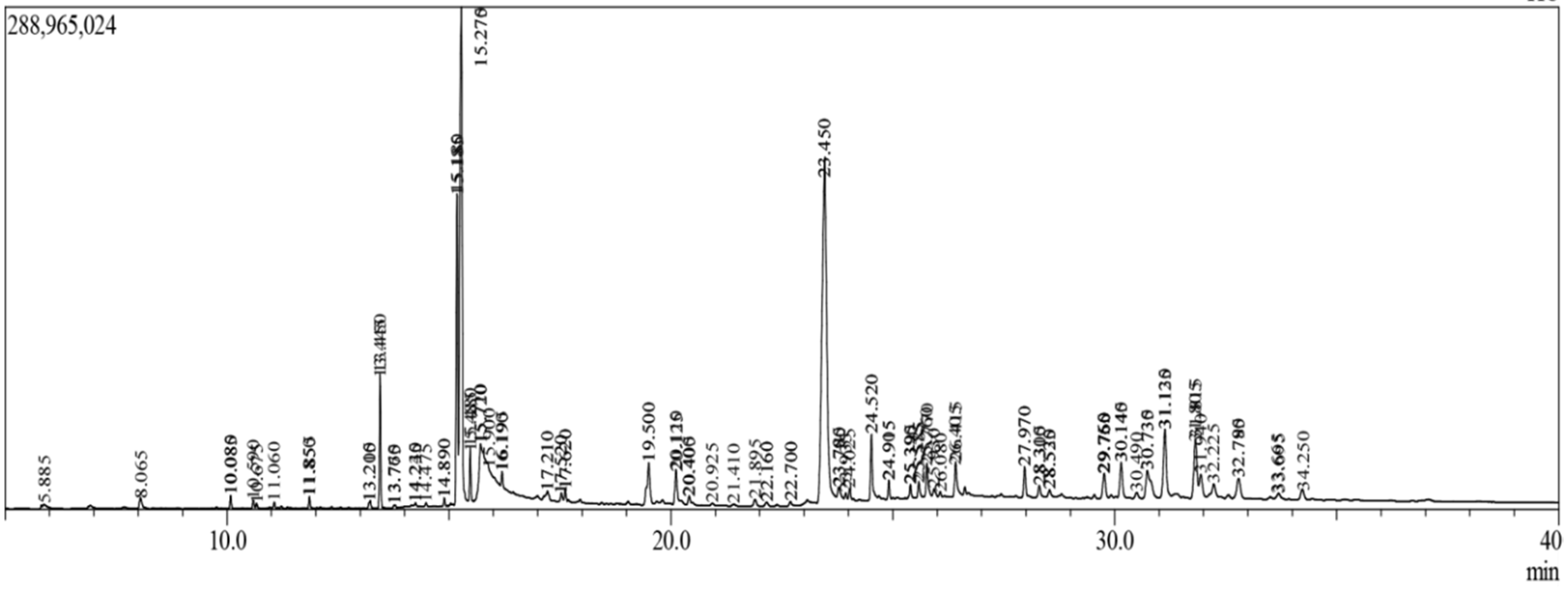
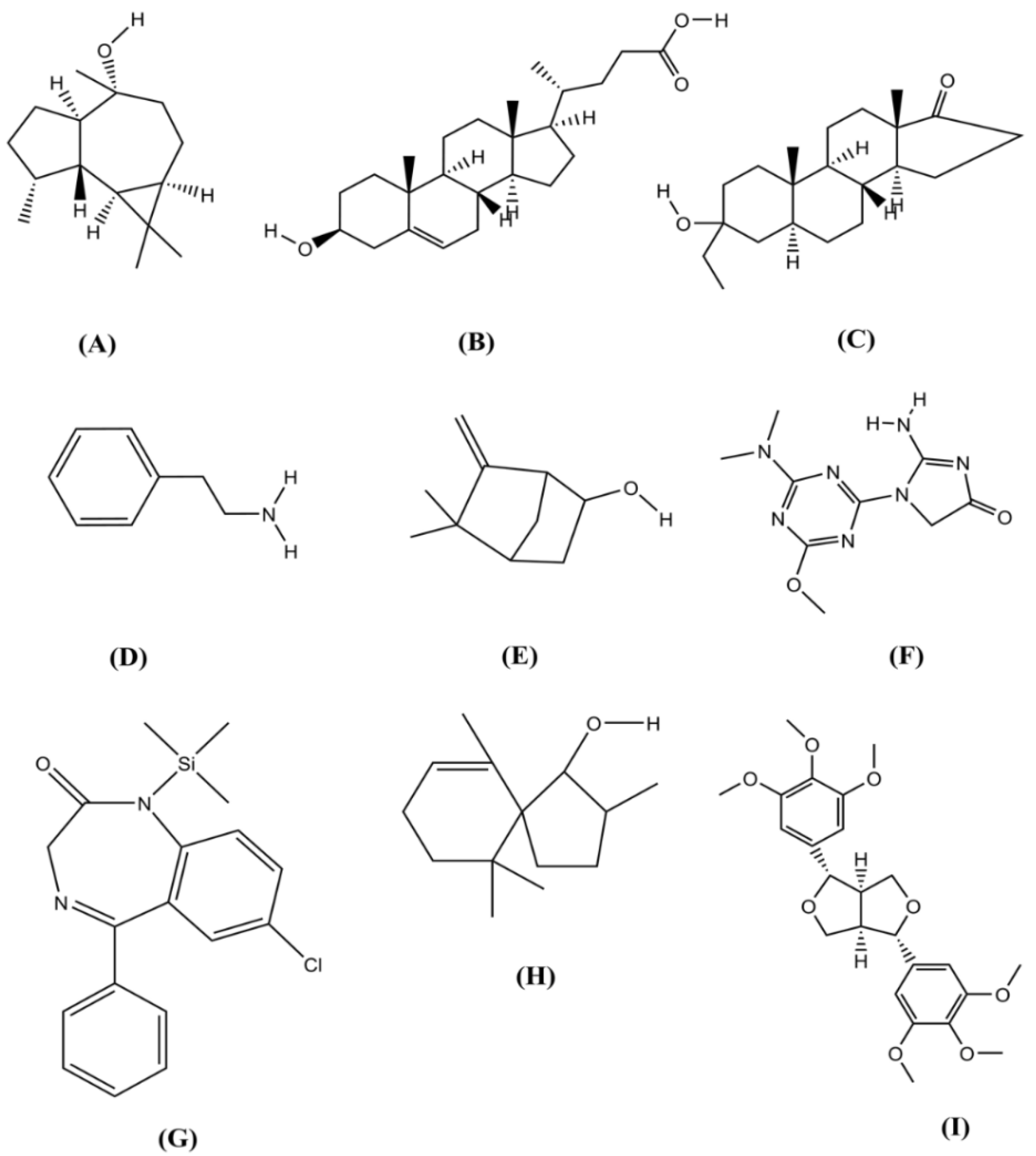
2.1. GC-MS Analysis
2.2. Prediction of Active Site
2.3. Ligand-Based ADME/T Prediction
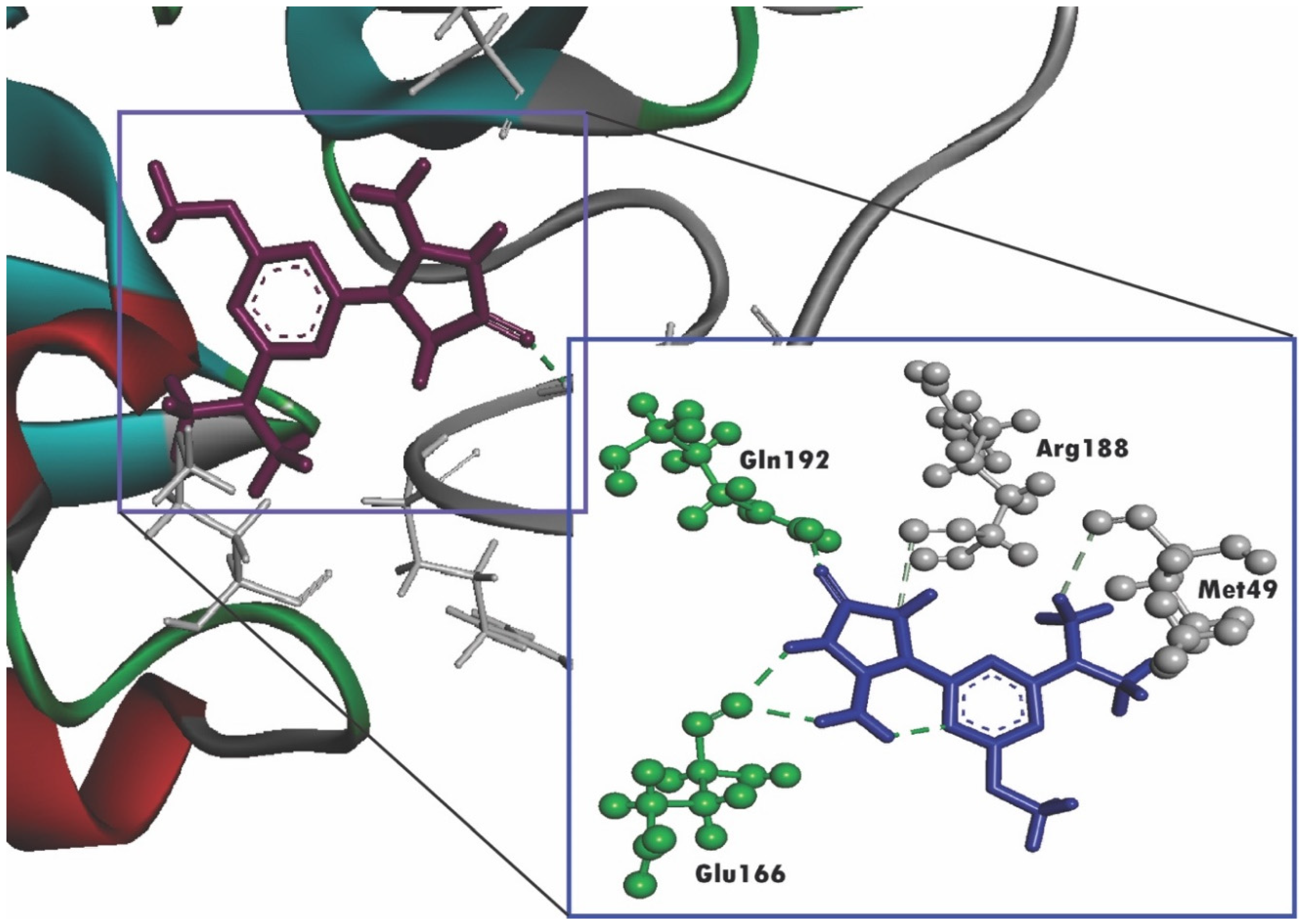
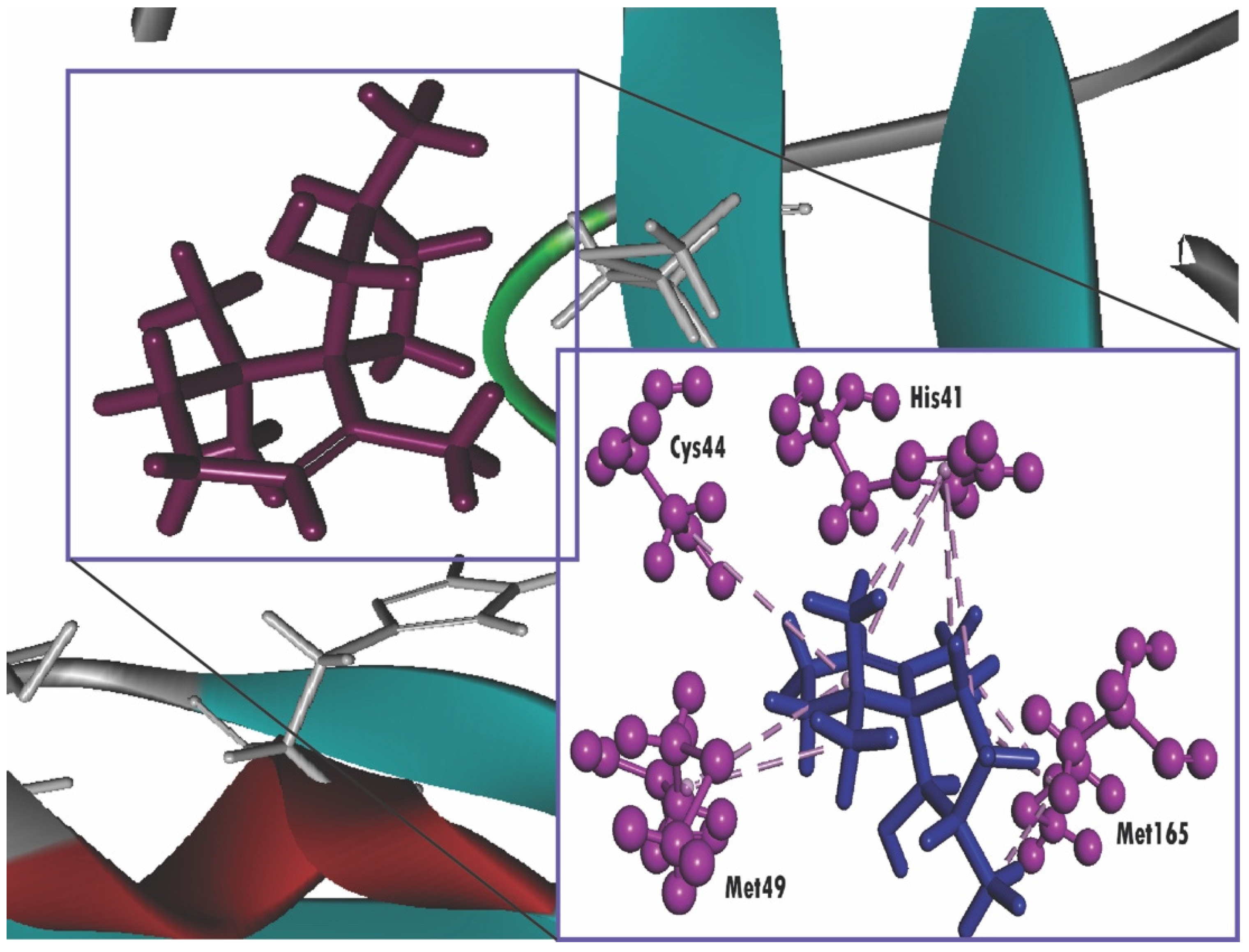
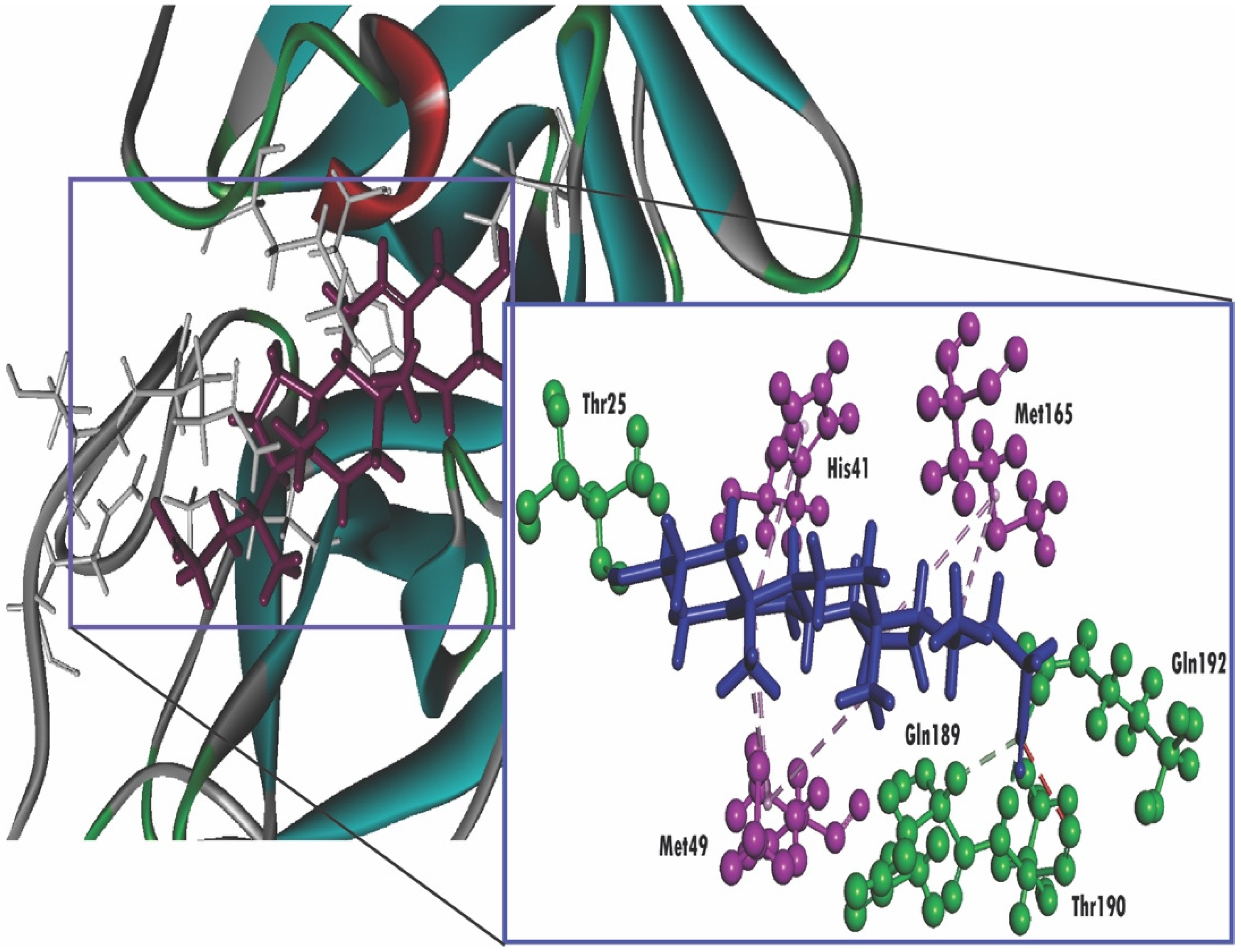
2.4. Molecular Docking
2.5. Prediction of Biological Activity
3. Discussion
4. Materials and Methods
4.1. Plant Collection
4.2. Preparation of Extracts
4.3. Gas Chromatography–Mass Spectroscopy (GC-MS) Analysis
4.4. Active Site Prediction
4.5. In Silico ADME Analysis
4.6. Computational Molecular Docking Analysis
4.6.1. Ligand Preparation
4.6.2. Protein Preparation
4.6.3. Receptor Grid Generation and Molecular Docking
4.7. Biological Activity Prediction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yang, Y.; Islam, M.S.; Wang, J.; Li, Y.; Chen, X. Traditional Chinese Medicine in the Treatment of Patients Infected with 2019-New Coronavirus (SARS-CoV-2): A Review and Perspective. Int. J. Biol. Sci. 2020, 16, 1708. [Google Scholar] [CrossRef]
- Chhikara, B.S.; Rathi, B.; Singh, J.; Poonam, F.N.U. Corona virus SARS-CoV-2 disease COVID-19: Infection, prevention and clinical advances of the prospective chemical drug therapeutics. Chem. Biol. Lett. 2020, 7, 63–72. [Google Scholar]
- Agbowuro, A.A.; Huston, W.M.; Gamble, A.B.; Tyndall, J.D.A. Proteases and protease inhibitors in infectious diseases. Med. Res. Rev. 2018, 38, 1295–1331. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, B.; Jin, Z.; Yang, H.; Rao, Z. The Crytal Structure of 2019-NCoV Main Protease in Complex with an Inhibitor N3. RCSB Protein Data Bank 2020. [Google Scholar]
- Ullrich, S.; Nitsche, C. The SARS-CoV-2 main protease as drug target. Bioorg. Med. Chem. Lett. 2020, 127377. [Google Scholar] [CrossRef]
- Ziebuhr, J.; Snijder, E.J.; Gorbalenya, A.E. Virus-encoded proteinases and proteolytic processing in the Nidovirales. J. Gen. Virol. 2000, 81, 853–879. [Google Scholar] [CrossRef]
- Hegyi, A.; Ziebuhr, J. Conservation of substrate specificities among coronavirus main proteases. J. Gen. Virol. 2002, 83, 595–599. [Google Scholar] [CrossRef]
- Du, Q.-S.; Wang, S.-Q.; Zhu, Y.; Wei, D.-Q.; Guo, H.; Sirois, S.; Chou, K.-C. Polyprotein cleavage mechanism of SARS-CoV Mpro and chemical modification of the octapeptide. Peptides 2004, 25, 1857–1864. [Google Scholar] [CrossRef]
- Thiel, V.; Ivanov, K.A.; Putics, A.; Hertzig, T.; Schelle, B.; Bayer, S.; Weißbrich, B.; Snijder, E.J.; Rabenau, H.; Doerr, H.W.; et al. Mechanisms and enzymes involved in SARS coronavirus genome expression. J. Gen. Virol. 2003, 84, 2305–2315. [Google Scholar] [CrossRef]
- Lu, H. Drug treatment options for the 2019-new coronavirus (2019-nCoV). Biosci. Trends 2020, 14, 69–71. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Peng, C.; Shi, Y.; Zhu, Z.; Mu, K.; Wang, X.; Zhu, W. Nelfinavir was predicted to be a potential inhibitor of 2019-nCov main protease by an integrative approach combining homology modelling, molecular docking and binding free energy calculation. BioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Khaerunnisa, S.; Kurniawan, H.; Awaluddin, R.; Suhartati, S.; Soetjipto, S. Potential Inhibitor of COVID-19 Main Protease (Mpro) from Several Medicinal Plant Compounds by Molecular Docking Study. Preprints 2020, 20944, 1–14. [Google Scholar]
- Khanam, B.H.; Rakib, A.; Faiz, F.B.; Uddin, M.G.; Hossain, M.K.; Sultan, R.A. Highly Anticancer and Moderate Thrombolytic Property of Accacia rugata of Mimosaceae Family. J. Plant. Sci. 2020, 8, 12–16. [Google Scholar]
- Dutta, T.; Paul, A.; Majumder, M.; Sultan, R.A.; Emran, T.B. Pharmacological evidence for the use of Cissus assamica as a medicinal plant in the management of pain and pyrexia. Biochem. Biophys. Rep. 2020, 21, 100715. [Google Scholar]
- Tareq, A.M.; Farhad, S.; Neshar Uddin, A.B.M.; Hoque, M.; Nasrin, M.S.; Uddin, M.M.R.; Hasan, M.; Sultana, A.; Munira, M.S.; Lyzu, C.; et al. Chemical profiles, pharmacological properties, and in silico studies provide new insights on Cycas pectinata. Heliyon 2020, 6, e04061. [Google Scholar] [CrossRef]
- Shifah, F.; Tareq, A.M.; Sayeed, M.A.; Islam, M.N.; Emran, T.B.; Ullah, M.A.; Mukit, M.A.; Ullah, A. Antidiarrheal, cytotoxic and thrombolytic activities of methanolic extract of Hedychium coccineum leaves. J. Adv. Biotechnol. Experiment. Therap. 2020, 11, 12. [Google Scholar] [CrossRef]
- Uddin, M.Z.; Rana, M.S.; Hossain, S.; Ferdous, S.; Dutta, E.; Dutta, M.; Emran, T. Bin In vivo neuroprotective, antinociceptive, anti-inflammatory potential in Swiss albino mice and in vitro antioxidant and clot lysis activities of fractionated Holigarna longifolia Roxb. bark extract. J. Complement. Integr. Med. 2019, 17. [Google Scholar] [CrossRef]
- Uddin, M.M.N.; Ahmed, S.; Kabir, M.S.H.; Rahman, M.S.; Sultan, R.A.; Emran, T. Bin In vivo analgesic, anti-inflammatory potential in Swiss albino mice and in vitro thrombolytic activity of hydroalcoholic fruits extract from Daemonorops robusta Warb. J. Appl. Pharm. Sci. 2017, 7, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Jahan, I.; Tona, M.R.; Sharmin, S.; Sayeed, M.A.; Tania, F.Z.; Paul, A.; Chy, M.N.U.; Rakib, A.; Emran, T.B.; Simal-Gandara, J. GC-MS Phytochemical Profiling, Pharmacological Properties, and In Silico Studies of Chukrasia velutina Leaves: A Novel Source for Bioactive Agents. Molecules 2020, 25, 3536. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Khatun, A.; Sharmin, T.; Mobin, F.; Tanu, A.R.; Morshed, T.; Fakir, T.A.; Begum, R.A.; Nabi, A.N. MPDB 1.0: A medicinal plant database of Bangladesh. Bioinformation 2014, 10, 384–386. [Google Scholar]
- Rakib, A.; Ahmed, S.; Islam, M.A.; Haye, A.; Uddin, S.M.N.; Uddin, M.M.N.; Hossain, M.K.; Paul, A.; Emran, T. Bin Antipyretic and hepatoprotective potential of Tinospora crispa and investigation of possible lead compounds through in silico approaches. Food Sci. Nutr. 2020, 8, 547–556. [Google Scholar] [CrossRef] [Green Version]
- Rakib, A.; Ahmed, S.; Islam, M.A.; Uddin, M.M.N.; Paul, A.; Chy, M.N.U.; Emran, T.B.; Seidel, V. Pharmacological studies on the antinociceptive, anxiolytic and antidepressant activity of Tinospora crispa. Phyther. Res. 2020, 1–7. [Google Scholar] [CrossRef]
- Niljan, J.; Jaihan, U.; Srichairatanakool, S.; Uthaipibull, C.; Somsak, V. Antimalarial activity of stem extract of Tinospora crispa against Plasmodium berghei infection in mice. J. Heal. Res. 2014, 28, 199–204. [Google Scholar]
- Gautret, P.; Lagier, J.-C.; Parola, P.; Meddeb, L.; Mailhe, M.; Doudier, B.; Courjon, J.; Giordanengo, V.; Vieira, V.E.; Dupont, H.T.; et al. Hydroxychloroquine and azithromycin as a treatment of COVID-19: Results of an open-label non-randomized clinical trial. Int. J. Antimicrob. Agents 2020, 56, 105949. [Google Scholar] [CrossRef]
- Colson, P.; Rolain, J.-M.; Lagier, J.-C.; Brouqui, P.; Raoult, D. Chloroquine and hydroxychloroquine as available weapons to fight COVID-19. Int. J. Antimicrob. Agents 2020, 10, 105932. [Google Scholar] [CrossRef]
- Direkbusarakom, S.; Herunsalee, A.; Yoshimizu, M.; Ezura, Y. Antiviral activity of several Thai traditional herb extracts against fish pathogenic viruses. Fish. Pathol. 1996, 31, 209–213. [Google Scholar] [CrossRef]
- Saha, S.; Ghosh, S. Tinospora cordifolia: One plant, many roles. Anc. Sci. Life 2012, 31, 151. [Google Scholar]
- Diao, K.; Han, P.; Pang, T.; Li, Y.; Yang, Z. HRCT Imaging Features in Representative Imported Cases of 2019 Novel Coronavirus Pneumonia. Precis. Clin. Med. 2020, 3, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Chang, D.; Lin, M.; Wei, L.; Xie, L.; Zhu, G.; Dela Cruz, C.S.; Sharma, L. Epidemiologic and Clinical Characteristics of Novel Coronavirus Infections Involving 13 Patients Outside Wuhan, China. JAMA-J. Am. Med. Assoc. 2020, 323, 1092–1093. [Google Scholar] [CrossRef]
- Holshue, M.L.; DeBolt, C.; Lindquist, S.; Lofy, K.H.; Wiesman, J.; Bruce, H.; Spitters, C.; Ericson, K.; Wilkerson, S.; Tural, A.; et al. First case of 2019 novel coronavirus in the United States. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Liu, Y.; Gayle, A.A.; Wilder-Smith, A.; Rocklöv, J. The reproductive number of COVID-19 is higher compared to SARS coronavirus. J. Travel Med. 2020. [Google Scholar] [CrossRef] [Green Version]
- Mahase, E. Coronavirus: Covid-19 has killed more people than SARS and MERS combined, despite lower case fatality rate. BMJ 2020, 368, m641. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Tang, Y.; Bao, B.; Zhang, P. Exploring the Active Compounds of Traditional Mongolian Medicine Agsirga in Intervention of Novel Coronavirus (2019-nCoV) Based on HPLC-Q-Exactive-MS/MS and Molecular Docking Method. 2020. Available online: https://chemrxiv.org/articles/Exploring_the_Active_Compounds_of_Traditional_Mongolian_Medicine_Agsirga_in_Intervention_of_Novel_Coronavirus_2019-nCoV_Based_on_HPLC-Q-Exactive-MS_MS_and_Molecular_Docking_Method/11955273 (accessed on 10 August 2020).
- Rakib, A.; Sami, S.A.; Islam, M.A.; Ahmed, S.; Faiz, F.B.; Khanam, B.H.; Marma, K.K.S.; Rahman, M.; Uddin, M.M.N.; Emran, T.B.; et al. Epitope-based Immunoinformatics Approach on Nucleocapsid Protein of Severe Acute Respiratory Syndrome-Coronavirus-2. 2020. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pmc/articles/PMC7423576/ (accessed on 10 August 2020).
- Oyebode, O.; Kandala, N.-B.; Chilton, P.J.; Lilford, R.J. Use of traditional medicine in middle-income countries: A WHO-SAGE study. Health Policy Plan. 2016, 31, 984–991. [Google Scholar] [CrossRef] [Green Version]
- Gwynn, J.; Hylands, P.J. Plants as a source of new medicines. Drug Discov. 2000, 55. Available online: https://www.researchgate.net/profile/Jennie_Gwynn2/publication/325417739_Plants_as_a_Source_of_New_Medicines_’Reproduced_with_kind_permission_of_Drug_Discovery_World_Summer_2000_Vol_1_No_1’/links/5b7bf331a6fdcc5f8b586555/Plants-as-a-Source-of-New-Medicines-Reproduced-with-kind-permission-of-Drug-Discovery-World-Summer-2000-Vol-1-No-1.pdf (accessed on 10 August 2020).
- Al Mahmud, Z.; Bachar, S.C.; Hasan, C.M.; Emran, T.B.; Qais, N.; Uddin, M.M.N. Phytochemical investigations and antioxidant potential of roots of Leea macrophylla (Roxb.). BMC Res. Notes 2017, 10, 245. [Google Scholar] [CrossRef]
- Al Mahmud, Z.; Emran, T.B.; Qais, N.; Bachar, S.C.; Sarker, M.; Uddin, M.M.N. Evaluation of analgesic, anti-inflammatory, thrombolytic and hepatoprotective activities of roots of Premna esculenta (Roxb). J. Basic Clin. Physiol. Pharmacol. 2016, 27, 63–70. [Google Scholar] [CrossRef]
- Emran, T.; E.-Mowla, T.; Ahmed, S.; Zahan, S.; Rakib, A.; Hasan, M.; Amin, M.; Mow, T.; Uddin, M. Sedative, Anxiolytic, Antinociceptive, Anti-inflammatory and Antipyretic Effects of a Chloroform Extract from the Leaves of Urena sinuata in Rodents. J. Appl. Life Sci. Int. 2018, 16, 1–19. [Google Scholar] [CrossRef]
- Wagner, H.K.; Wolff, P.M. New Natural Products and Plant. In Drugs with Pharmacological, Biological or Therapeutical Activity: Proceedings of the First International Congress on Medicinal Plant; Farnsworth, N.R., Bingel, A.S., Eds.; Springer Science & Business Media: New York, NY, USA, 2012; pp. 6–10. [Google Scholar]
- Emran, T.B.; Rahman, M.A.; Uddin, M.M.N.; Rahman, M.M.; Uddin, M.Z.; Dash, R.; Layzu, C. Effects of organic extracts and their different fractions of five Bangladeshi plants on in vitro thrombolysis. BMC Complement. Altern. Med. 2015, 15, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Vasanthi, H.; ShriShriMal, N.; Das, D.K. Phytochemicals from plants to combat cardiovascular disease. Curr. Med. Chem. 2012, 19, 2242–2251. [Google Scholar] [CrossRef]
- Lee, K.W.; Bode, A.M.; Dong, Z. Molecular targets of phytochemicals for cancer prevention. Nat. Rev. Cancer 2011, 11, 211–218. [Google Scholar] [CrossRef]
- Singh, R.; Arif, T.; Khan, I.; Sharma, P. Phytochemicals in antidiabetic drug discovery. J. Biomed. Ther. Sci. 2014, 1, 1–33. [Google Scholar]
- Korkina, L.; Kostyuk, V.; Potapovich, A.; Mayer, W.; Talib, N.; De Luca, C. Secondary Plant Metabolites for Sun Protective Cosmetics: From Pre-Selection to Product Formulation. Cosmetics 2018, 5, 32. [Google Scholar] [CrossRef] [Green Version]
- Sermakkani, M.; Thangapandian, V. GC-MS analysis of Cassia italica leaf methanol extract. Asian, J. Pharm. Clin. Res. 2012, 5, 90–94. [Google Scholar]
- Barua, N.; Aziz, M.A.I.; Tareq, A.M.; Sayeed, M.A.; Alam, N.; ul Alam, N.; Uddin, M.A.; Lyzu, C.; Emran, T.B. In vivo and in vitro evaluation of pharmacological activities of Adenia trilobata (Roxb.). Biochem. Biophys. Rep. 2020, 23, 100772. [Google Scholar] [CrossRef]
- ul Qamar, M.T.; Alqahtani, S.M.; Alamri, M.A.; Chen, L.-L. Structural basis of SARS-CoV-2 3CLpro and anti-COVID-19 drug discovery from medicinal plants. J. Pharm. Anal. 2020. [Google Scholar] [CrossRef]
- Adem, S.; Eyupoglu, V.; Sarfraz, I.; Rasul, A.; Ali, M. Identification of potent COVID-19 main protease (Mpro) inhibitors from natural polyphenols: An in silico strategy unveils a hope against CORONA. Preprints 2020, 2020030333. [Google Scholar] [CrossRef] [Green Version]
- Phoebe Chen, Y.-P.; Chen, F. Identifying targets for drug discovery using bioinformatics. Expert Opin. Ther. Targets 2008, 12, 383–389. [Google Scholar] [CrossRef]
- Ahmed, S.; Rakib, A.; Islam, M.A.; Khanam, B.H.; Faiz, F.B.; Paul, A.; Chy, M.N.U.; Bhuiya, N.M.M.A.; Uddin, M.M.N.; Ullah, S.M.A.; et al. In vivo and in vitro pharmacological activities of Tacca integrifolia rhizome and investigation of possible lead compounds against breast cancer through in silico approaches. Clin. Phytosci. 2019, 5, 36. [Google Scholar] [CrossRef]
- Brooijmans, N.; Kuntz, I.D. Molecular recognition and docking algorithms. Annu. Rev. Biophys. Biomol. Struct. 2003, 32, 335–373. [Google Scholar] [CrossRef]
- Rakib, A.; Sami, S.A.; Mimi, N.J.; Chowdhury, M.M.; Eva, T.A.; Nainu, F.; Paul, A.; Shahriar, A.; Tareq, A.M.; Emon, N.U.; et al. Immunoinformatics-guided design of an epitope-based vaccine against severe acute respiratory syndrome coronavirus 2 spike glycoprotein. Comput. Biol. Med. 2020, 124, 103967. [Google Scholar] [CrossRef]
- Yamamoto, N.; Matsuyama, S.; Hoshino, T.; Yamamoto, N. Nelfinavir inhibits replication of severe acute respiratory syndrome coronavirus 2 in vitro. BioRxiv 2020. [Google Scholar] [CrossRef]
- Yamamoto, N.; Yang, R.; Yoshinaka, Y.; Amari, S.; Nakano, T.; Cinatl, J.; Rabenau, H.; Doerr, H.W.; Hunsmann, G.; Otaka, A.; et al. HIV protease inhibitor nelfinavir inhibits replication of SARS-associated coronavirus. Biochem. Biophys. Res. Commun. 2004, 318, 719–725. [Google Scholar] [CrossRef]
- Kang, C.K.; Seong, M.-W.; Choi, S.-J.; Kim, T.S.; Choe, P.G.; Song, S.H.; Kim, N.-J.; Park, W.B.; Oh, M. In vitro activity of lopinavir/ritonavir and hydroxychloroquine against severe acute respiratory syndrome coronavirus 2 at concentrations achievable by usual doses. Korean J. Intern. Med. 2020, 35, 728–787. [Google Scholar] [CrossRef]
- Goyal, B.; Goyal, D. Targeting the dimerization of main protease of coronaviruses: A potential broad-spectrum therapeutic strategy. ACS Comb. Sci. 2020, 22, 297–305. [Google Scholar] [CrossRef]
- Muramatsu, T.; Takemoto, C.; Kim, Y.-T.; Wang, H.; Nishii, W.; Terada, T.; Shirouzu, M.; Yokoyama, S. SARS-CoV 3CL protease cleaves its C-terminal autoprocessing site by novel subsite cooperativity. Proc. Natl. Acad. Sci. USA 2016, 113, 12997–13002. [Google Scholar] [CrossRef] [Green Version]
- Hsu, M.-F.; Kuo, C.-J.; Chang, K.-T.; Chang, H.-C.; Chou, C.-C.; Ko, T.-P.; Shr, H.-L.; Chang, G.-G.; Wang, A.H.-J.; Liang, P.-H. Mechanism of the maturation process of SARS-CoV 3CL protease. J. Biol. Chem. 2005, 280, 31257–31266. [Google Scholar] [CrossRef] [Green Version]
- Volkamer, A.; Kuhn, D.; Rippmann, F.; Rarey, M. DoGSiteScorer: A web server for automatic binding site prediction, analysis and druggability assessment. Bioinformatics 2012, 28, 2074–2075. [Google Scholar] [CrossRef] [Green Version]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 1997, 23, 3–25. [Google Scholar] [CrossRef]
- Lubomirov, R.; di Iulio, J.; Fayet, A.; Colombo, S.; Martinez, R.; Marzolini, C.; Furrer, H.; Vernazza, P.; Calmy, A.; Cavassini, M.; et al. ADME pharmacogenetics: Investigation of the pharmacokinetics of the antiretroviral agent lopinavir coformulated with ritonavir. Pharmacogenet. Genomics 2010, 20, 217–230. [Google Scholar] [CrossRef]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra precision glide: Docking and scoring incorporating a model of hydrophobic enclosure for protein-ligand complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [Green Version]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A New Approach for Rapid, Accurate Docking and Scoring. 1. Method and Assessment of Docking Accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | Retention Time | m/z | Area | PA (%) |
|---|---|---|---|---|
| Benzeneethanamine | 10.083 | 73.00 | 3196250 | 0.106518 |
| Camphenol | 13.761 | 107.00 | 431753 | 0.014389 |
| Strophanthidin | 22.159 | 79.00 | 334706 | 0.011154 |
| Retinal | 22.159 | 79.00 | 334706 | 0.011154 |
| Trans-Geranylgeraniol | 24.911 | 69.00 | 4724235 | 0.15744 |
| 3,4-Dihydroxymandelic acid | 17.261 | 73.00 | 2174852 | 0.072479 |
| Imidazolidin-4-one, 2-imino-1-(4-methoxy-6-dimethylamino-1,3,5-triazin-2-yl) | 25.586 | 251.00 | 4374165 | 0.145774 |
| Cholest-22-ene-21-ol, 3,5-dehydro-6-methoxy | 25.771 | 95.00 | 5058177 | 0.168569 |
| d-Mannitol, 1-O-(16-hydroxyhexadecyl)- | 26.431 | 207.00 | 153759 | 0.005124 |
| Heneicosanoic acid, methyl ester | 26.431 | 207.00 | 153759 | 0.005124 |
| Gorgost-5-en-3-ol, (3.beta)-, TMS derivative | 26.431 | 207.00 | 153759 | 0.005124 |
| Retinol | 26.414 | 91.00 | 1957701 | 0.065242 |
| Octacosanol | 27.969 | 97.00 | 3033614 | 0.101098 |
| Alpha-Santalol | 28.532 | 94.00 | 3133161 | 0.104416 |
| Santalol, E-cis,epi-beta- | 28.532 | 94.00 | 3133161 | 0.104416 |
| Spiro[4,5]dec-6-en-1-ol, 2,6,10,10-tetramethyl | 28.532 | 94.00 | 3133161 | 0.104416 |
| Campesterol | 29.551 | 207.00 | 243053 | 0.0081 |
| Cholesterol | 29.551 | 207.00 | 243053 | 0.0081 |
| 9,19-Cyclocholestan-3-ol, 14-methyl-, (3.beta) | 29.551 | 207.00 | 243053 | 0.0081 |
| Cholest-5-en-3-ol, 6-methyl-, (3.beta)- | 29.551 | 207.00 | 243053 | 0.0081 |
| 26-Hydroxycholesterol | 29.551 | 207.00 | 243053 | 0.0081 |
| Beta-Sitosterol | 29.551 | 207.00 | 243053 | 0.0081 |
| Lathosterol | 29.551 | 207.00 | 243053 | 0.0081 |
| Ergost-7-en-3-ol | 29.551 | 207.00 | 243053 | 0.0081 |
| Cholest-5-en-3-ol (3.beta)-, carbonochloridate | 29.551 | 207.00 | 243053 | 0.0081 |
| Stigmasterol | 29.551 | 207.00 | 243053 | 0.0081 |
| Cholesta-5,22-dien-3-ol, (3.beta)- | 29.551 | 207.00 | 243053 | 0.0081 |
| Ergosta-5,24(28)-dien-3-ol, (3.beta)- | 29.551 | 207.00 | 243053 | 0.0081 |
| Lathosterol | 29.551 | 207.00 | 243053 | 0.0081 |
| Cholestane-3,5-diol, 5-acetate, (3.beta,5.alpha | 29.551 | 207.00 | 243053 | 0.0081 |
| 26,27-Dinorergosta-5,23-dien-3-ol, (3.beta)- | 30.145 | 55.00 | 2543634 | 0.084769 |
| Desmosterol | 30.145 | 55.00 | 2543634 | 0.084769 |
| 5,6-Dihydroergosterol | 30.145 | 55.00 | 2543634 | 0.084769 |
| 9,19-Cyclolanost-23-ene-3,25-diol, 3-acetate | 30.787 | 207.00 | 142424 | 0.004746 |
| Lupeol | 30.787 | 207.00 | 142424 | 0.004746 |
| 3.beta-Hydroxy-5-cholen-24-oic acid | 31.132 | 43.00 | 4086523 | 0.136188 |
| 26,27-Dinorergost-5-ene-3,24-diol, (3.beta)- | 31.132 | 43.00 | 4086523 | 0.136188 |
| 9,19-Cyclolanostan-3-ol, 24-methylene-, (3.beta) | 29.551 | 95.00 | 3611829 | 0.120368 |
| Lupeol, trifluoroacetate | 31.820 | 95.00 | 3611829 | 0.120368 |
| Lup-20(29)-en-3-ol, acetate, (3beta)- | 32.780 | 207.00 | 1038162 | 0.034598 |
| Phosphonoacetic Acid, 3TMS derivative | 5.819 | 73.00 | 653645 | 0.021783 |
| Nordazepam, TMS derivative | 5.819 | 73.00 | 653645 | 0.021783 |
| 2,6-Dihydroxybenzoic acid, 3TMS derivative | 10.083 | 73.00 | 3196250 | 0.106518 |
| aR-Turmerone | 10.592 | 83.00 | 2528243 | 0.084256 |
| (Z)-.gamma.-Atlantone | 10.592 | 83.00 | 2528243 | 0.084256 |
| Verbenylangelate, cis- | 10.592 | 83.00 | 2528243 | 0.084256 |
| Tumerone | 10.592 | 83.00 | 2548801 | 0.084941 |
| Dibutyl phthalate | 13.793 | 149.00 | 1573158 | 0.052427 |
| (−)-Globulol | 22.159 | 79.00 | 334706 | 0.011154 |
| Androstan-17-one, 3-ethyl-3-hydroxy-, (5.alpha) | 22.159 | 79.00 | 334706 | 0.011154 |
| Eudesma-4(15),7-dien-1.beta –ol | 22.159 | 79.00 | 334706 | 0.011154 |
| 5.alpha-Cholest-8-en-3-one, 14-methyl- | 25.771 | 95.00 | 5058177 | 0.168569 |
| 25-Hydroxycholesterol, 3-methyl ether | 26.622 | 207.00 | 362457 | 0.012079 |
| 26-Homo-25-hydroxycholesterol | 29.551 | 207.00 | 243053 | 0.0081 |
| Betulin | 32.780 | 207.00 | 1038162 | 0.034598 |
| Yangambin | 34.225 | 207.00 | 1108879 | 0.036955 |
| Compound Name | Molecular Weight | Num. H-Bond Acceptors | Num. H-Bond Donors | Log P | Molar Refractivity | No. of Violation |
|---|---|---|---|---|---|---|
| Benzeneethanamine | 121.18 | 1 | 1 | 1.7 | 38.92 | 0 |
| Camphenol | 152.23 | 1 | 1 | 2.30 | 46.38 | 0 |
| Strophanthidin | 404.5 | 6 | 3 | 1.82 | 106.16 | 0 |
| Retinal | 284.44 | 1 | 0 | 4.39 | 93.71 | 0 |
| Trans-Geranylgeraniol | 290.48 | 1 | 1 | 4.95 | 97.52 | 0 |
| 3,4-Dihydroxymandelic acid | 184.15 | 5 | 4 | −0.36 | 43.19 | 0 |
| Imidazolidin-4-one, 2-imino-1-(4-methoxy-6-dimethylamino-1,3,5-triazin-2-yl) | 251.25 | 6 | 1 | −0.59 | 70.28 | 0 |
| Cholest-22-ene-21-ol, 3,5-dehydro-6-methoxy | 498.78 | 3 | 0 | 6.39 | 151.03 | 1 |
| d-Mannitol, 1-O-(16-hydroxyhexadecyl)- | 422.60 | 7 | 6 | 0.74 | 115.92 | 1 |
| Heneicosanoic acid, methyl ester | 340.58 | 2 | 0 | 5.58 | 109.15 | 1 |
| Gorgost-5-en-3-ol, (3.beta)-, TMS derivative | 498.90 | 1 | 0 | 7.49 | 157.87 | 2 |
| Retinol | 286.45 | 1 | 1 | 4.48 | 94.67 | 0 |
| Octacosanol | 410.76 | 1 | 1 | 7.07 | 137.87 | 2 |
| Alpha-Santalol | 220.35 | 1 | 1 | 3.67 | 68.04 | 0 |
| Santalol, E-cis,epi-.beta- | 220.35 | 1 | 1 | 3.56 | 69.94 | 0 |
| Spiro[4,5]dec-6-en-1-ol, 2,6,10,10-tetramethyl | 208.34 | 1 | 1 | 3.41 | 65.35 | 0 |
| Campesterol | 400.68 | 1 | 1 | 6.54 | 128.42 | 1 |
| Cholesterol | 386.65 | 1 | 1 | 6.34 | 123.61 | 1 |
| 9,19-Cyclocholestan-3-ol, 14-methyl-, (3.beta) | 400.68 | 1 | 1 | 6.68 | 126.26 | 1 |
| Cholest-5-en-3-ol, 6-methyl-, (3.beta)- | 400.68 | 1 | 1 | 6.54 | 128.42 | 1 |
| 26-Hydroxycholesterol | 402.65 | 2 | 2 | 5.41 | 124.78 | 1 |
| Beta-Sitosterol | 414.71 | 1 | 1 | 6.73 | 133.23 | 2 |
| Lathosterol | 386.65 | 1 | 1 | 6.34 | 123.61 | 1 |
| Ergost-7-en-3-ol | 400.68 | 1 | 1 | 6.54 | 128.42 | 1 |
| Cholest-5-en-3-ol (3.beta)-, carbonochloridate | 449.11 | 2 | 0 | 6.51 | 133.73 | 2 |
| Stigmasterol | 412.69 | 1 | 1 | 6.62 | 132.75 | 2 |
| Cholesta-5,22-dien-3-ol, (3.beta)- | 384.64 | 1 | 1 | 6.23 | 123.14 | 1 |
| Ergosta-5,24(28)-dien-3-ol, (3.beta)- | 398.66 | 1 | 1 | 6.43 | 127.95 | 1 |
| Lathosterol | 386.65 | 1 | 1 | 6.34 | 123.61 | 1 |
| Cholestane-3,5-diol, 5-acetate, (3.beta,5.alpha | 446.71 | 3 | 1 | 5.74 | 135.03 | 2 |
| 26,27-Dinorergosta-5,23-dien-3-ol, (3.beta)- | 370.61 | 1 | 1 | 6.03 | 118.33 | 1 |
| Desmosterol | 384.64 | 1 | 1 | 6.23 | 123.14 | 1 |
| 5,6-Dihydroergosterol | 398.66 | 1 | 1 | 6.43 | 127.95 | 1 |
| 9,19-Cyclolanost-23-ene-3,25-diol, 3-acetate | 484.75 | 3 | 1 | 6.20 | 146.08 | 2 |
| Lupeol | 426.72 | 1 | 1 | 6.92 | 135.14 | 2 |
| 3.beta-Hydroxy-5-cholen-24-oic acid | 374.56 | 3 | 2 | 4.62 | 110.97 | 0 |
| 26,27-Dinorergost-5-ene-3,24-diol, (3.beta)- | 388.63 | 2 | 2 | 5.21 | 120.01 | 1 |
| 9,19-Cyclolanostan-3-ol, 24-methylene-, (3.beta) | 440.74 | 1 | 1 | 7.12 | 139.95 | 2 |
| Lupeol, trifluoroacetate | 522.73 | 5 | 0 | 7.36 | 145.07 | 3 |
| Lup-20(29)-en-3-ol, acetate, (3beta)- | 468.75 | 2 | 0 | 7.08 | 144,.88 | 2 |
| Phosphonoacetic Acid, 3TMS derivative | 356.58 | 5 | 0 | 1.42 | 90.71 | 0 |
| Nordazepam, TMS derivative | 342.89 | 2 | 0 | 3.14 | 105.43 | 0 |
| 2,6-Dihydroxybenzoic acid, 3TMS derivative | 370.66 | 4 | 0 | 2.97 | 103.15 | 0 |
| aR-Turmerone | 216.32 | 1 | 0 | 3.68 | 69.75 | 0 |
| (Z)-gamma.-Atlantone | 218.33 | 1 | 0 | 3.37 | 70.88 | 0 |
| Verbenylangelate, cis- | 234.33 | 2 | 0 | 3.35 | 70.07 | 0 |
| Tumerone | 218.33 | 1 | 0 | 3.37 | 70.88 | 0 |
| Dibutyl phthalate | 278.34 | 4 | 0 | 3.43 | 77.84 | 0 |
| (−)-Globulol | 222.37 | 1 | 1 | 3.81 | 68.82 | 0 |
| Androstan-17-one, 3-ethyl-3-hydroxy-,(5.alpha) | 318.49 | 2 | 1 | 4.15 | 95.48 | 0 |
| Eudesma-4(15),7-dien-1.beta–ol | 220.35 | 1 | 1 | 3.56 | 69.94 | 0 |
| 5.alpha-Cholest-8-en-3-one, 14-methyl- | 398.66 | 1 | 0 | 6.43 | 127.20 | 1 |
| Compound Name | Docking Score | Residues Interacting with Ligand through H-Bonding (H-Bonds No.) | Hydrophobic Bonds (pi-Alkyl Stacked) | Hydrophobic Bonds (pi-pi Stacked) |
|---|---|---|---|---|
| Camphenol | −6.177 | His164 | His41, Met49 (3), Met165 | |
| Strophanthidin | −5.8 | Gln189 | Met165 | |
| Benzeneethanamine | −6.022 | His41 | His41 | |
| Retinal | −5.591 | His163 | His41, Cys145,Cys44, Met49 | |
| Trans-geranylgeraniol | −3.393 | His163, Ser144 | Met49 (2), Arg188 | |
| 3,4-Dihydroxymandelic acid | −5.51 | Gly143,Ser144 | ||
| Imidazolidin-4-one, 2-imino-1-(4-methoxy-6-dimethylamino-1,3,5-triazin-2-yl) | −7.013 | Glu166 (2), Gln192 | ||
| Retinol | −5.576 | Thr24 | Cys145, Met165, His41, Cys44, Met49 | |
| Alpha-Santalol | −5.595 | Gln192 | His41 (2), Met49 (3), Pro168, Met165 | |
| Santalol, E-cis, epi-beta- | −5.664 | Gln192 | His41, Met49 (2), Pro168, | |
| Spiro[4,5]dec-6-en-1-ol, 2,6,10,10-tetramethyl | −6.369 | Met165 (3), His41 (4), Met49 (2), Cys145 | ||
| 3.beta-Hydroxy-5-cholen-24-oic acid | −6.251 | Thr25, Thr190,Gln192 | Met49 (3), His41, Met165 (2) | |
| Phosphonoacetic Acid, 3TMS derivative | −4.273 | Glu166 | Cys145 (2), His41 (2), Met165, His163 | |
| Nordazepam, TMS derivative | −6.122 | Glu166 | Met165 | |
| 2,6-Dihydroxybenzoic acid, 3TMS derivative | −4.696 | Met165 (2), His41 (3), Cys145 | ||
| aR-Turmerone | −5.452 | Cys44 | Met165, Pro168, His41 (2), Met49 | |
| (Z)-gamma.-Atlantone | −5.708 | Gln189 | Met49, Met165, His41, Pro168 (2) | |
| Verbenyl angelate, cis- | −5.579 | Met165 (2), Pro52, Arg188, Cys44 (4), Met49 (4) | ||
| Tumerone | −5.131 | Met165 (2), His41 (2), Cys44 | ||
| Dibutyl phthalate | −2.279 | Gly143, Asn142 | His41, His163, His172 | |
| (−)-Globulol | −6.165 | Met49 (2), His41(4), Cys44 (2), Met165 (3) | ||
| Androstan-17-one, 3-ethyl-3-hydroxy-(5.alpha) | −6.218 | Thr26 | Cys145, Met49, His41 (3) | |
| Yangambin | −6.162 | Thr25 | Met49, Met165 | His41 |
| Nelfinavir | −7.596 | His41, Glu166 (2) | His41 | Met165 |
| Lopinavir | −8.251 | Gln189, Glu166, Cys141, Thr26 | Pro168, His41 (2) | Arg188, His41 |
| Compounds | GPCR Ligand | Ion Channel Inhibitor | Kinase Inhibitor | Nuclear Receptor Ligand | Protease Inhibitor | Enzyme Inhibitor |
|---|---|---|---|---|---|---|
| Benzeneethanamine | −1.71 | −1.16 | −1.95 | −2.61 | −1.85 | −1.43 |
| Camphenol | −0.66 | −0.43 | −1.53 | −0.62 | −1.06 | −0.37 |
| Strophanthidin | 0.08 | 0.07 | −0.46 | 0.52 | 0.01 | 0.79 |
| Retinal | −0.15 | 0.15 | −0.23 | 0.90 | 0.09 | 0.52 |
| Trans-Geranylgeraniol | 0.12 | 0.20 | −0.22 | 0.40 | −0.08 | 0.41 |
| 3,4-Dihydroxymandelic acid | −0.28 | −0.18 | −0.69 | −0.06 | −0.61 | −0.05 |
| Imidazolidin-4-one, 2-imino-1-(4-methoxy-6-dimethylamino-1,3,5-triazin-2-yl) | 0.15 | −0.15 | −0.20 | −0.63 | −0.55 | 0.08 |
| Retinol | −0.01 | 0.32 | −0.25 | 1.02 | −0.16 | 0.66 |
| Alpha-Santalol | −0.04 | 0.03 | −0.43 | 0.12 | −0.22 | 0.37 |
| Santalol, E-cis,epi-.beta- | −0.09 | −0.04 | −0.65 | 0.23 | −0.42 | 0.39 |
| Spiro[4,5]dec-6-en-1-ol, 2,6,10,10-tetramethyl | −0.23 | 0.00 | −0.79 | 0.40 | −0.27 | 0.43 |
| 3.beta-Hydroxy-5-cholen-24-oic acid | 0.20 | 0.03 | −0.57 | 0.87 | 0.07 | 0.64 |
| Phosphonoacetic Acid, 3TMS derivative | 0.43 | 0.59 | 0.12 | −0.05 | 0.71 | 1.08 |
| Nordazepam, TMS derivative | 0.48 | 0.58 | −0.22 | −0.07 | 0.33 | 0.35 |
| 2,6-Dihydroxybenzoic acid, 3TMS derivative | 0.34 | 0.12 | −0.03 | −0.06 | 0.42 | 0.72 |
| aR-Turmerone | −0.68 | −0.46 | −1.36 | −0.14 | −0.80 | −0.25 |
| (Z)-gamma.-Atlantone | −0.38 | 0.15 | −1.17 | 0.37 | −0.58 | 0.46 |
| Verbenyl angelate, cis- | −0.09 | −0.08 | −0.98 | 0.32 | −0.35 | 0.28 |
| Tumerone | −0.35 | −0.13 | −1.19 | 0.54 | −0.44 | 0.41 |
| Dibutyl phthalate | −0.16 | −0.09 | −0.27 | −0.12 | −0.25 | −0.07 |
| (−)-Globulol | −0.50 | −0.29 | −0.82 | −0.22 | −0.48 | −0.13 |
| Androstan-17-one, 3-ethyl-3-hydroxy-, (5.alpha) | 0.19 | 0.41 | −0.35 | 0.83 | 0.17 | 0.66 |
| Yangambin | −0.03 | −0.25 | −0.19 | −0.10 | −0.16 | 0.01 |
| Nelfinavir | 0.19 | −0.25 | −0.28 | −0.25 | 0.58 | −0.02 |
| Lopinavir | 0.04 | −0.78 | −0.55 | −0.66 | 0.42 | −0.37 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rakib, A.; Paul, A.; Chy, M.N.U.; Sami, S.A.; Baral, S.K.; Majumder, M.; Tareq, A.M.; Amin, M.N.; Shahriar, A.; Uddin, M.Z.; et al. Biochemical and Computational Approach of Selected Phytocompounds from Tinospora crispa in the Management of COVID-19. Molecules 2020, 25, 3936. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25173936
Rakib A, Paul A, Chy MNU, Sami SA, Baral SK, Majumder M, Tareq AM, Amin MN, Shahriar A, Uddin MZ, et al. Biochemical and Computational Approach of Selected Phytocompounds from Tinospora crispa in the Management of COVID-19. Molecules. 2020; 25(17):3936. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25173936
Chicago/Turabian StyleRakib, Ahmed, Arkajyoti Paul, Md. Nazim Uddin Chy, Saad Ahmed Sami, Sumit Kumar Baral, Mohuya Majumder, Abu Montakim Tareq, Mohammad Nurul Amin, Asif Shahriar, Md. Zia Uddin, and et al. 2020. "Biochemical and Computational Approach of Selected Phytocompounds from Tinospora crispa in the Management of COVID-19" Molecules 25, no. 17: 3936. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25173936