Capsaicin Causes Vasorelaxation of Rat Aorta through Blocking of L-type Ca2+ Channels and Activation of CB1 Receptors


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
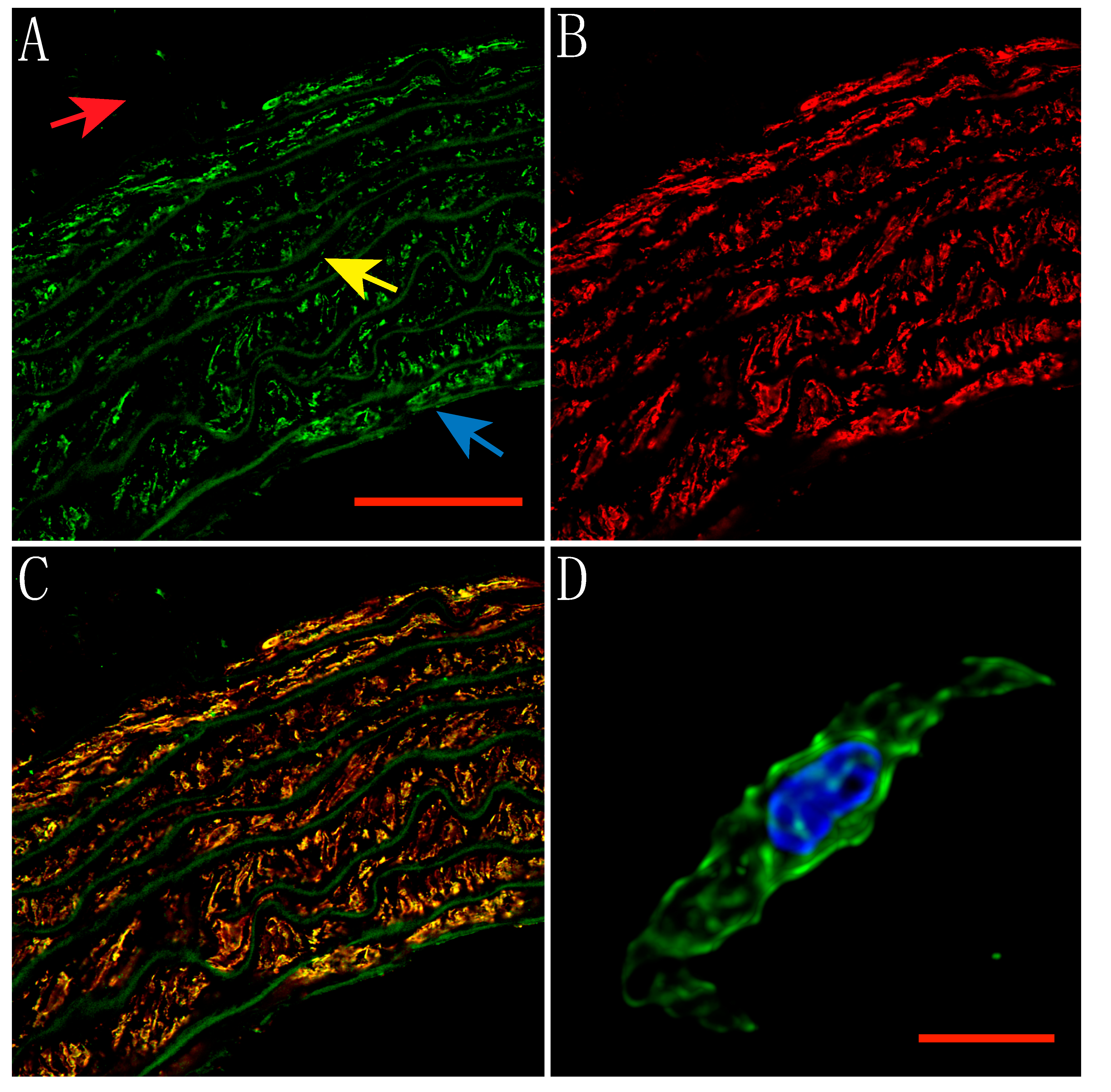
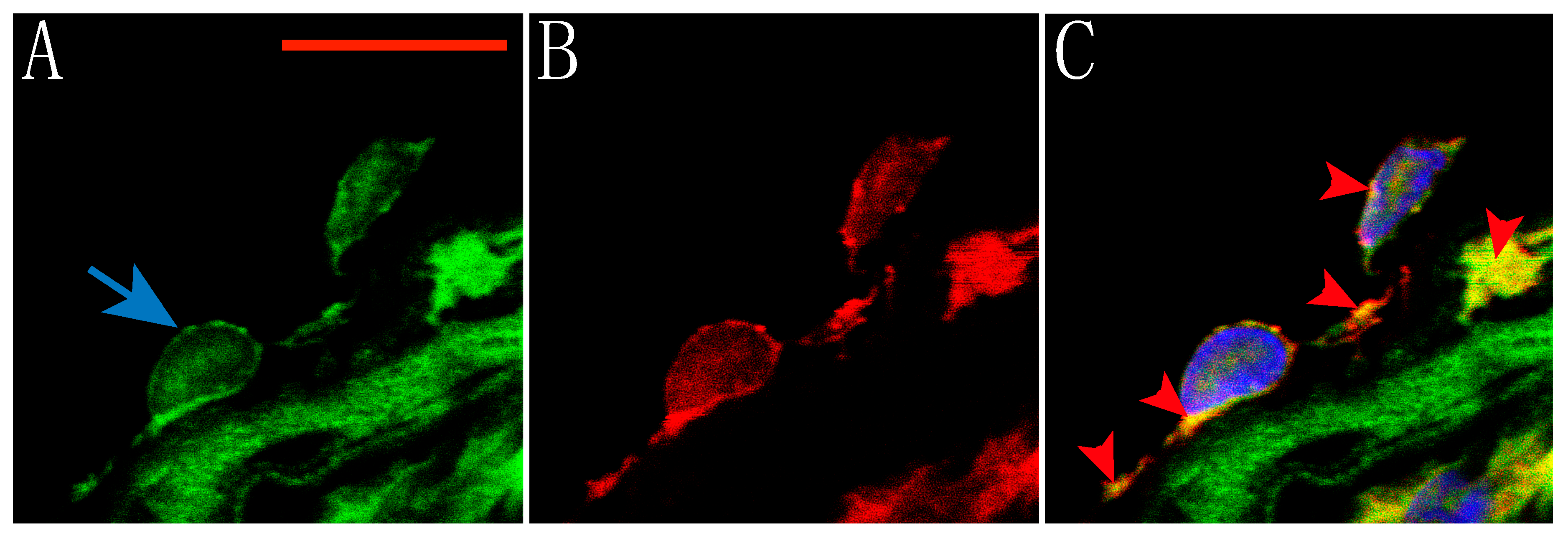
2.1. Expression of TRPV1 Receptors in Smooth Muscle and Endothelial Cells of Rat Aorta
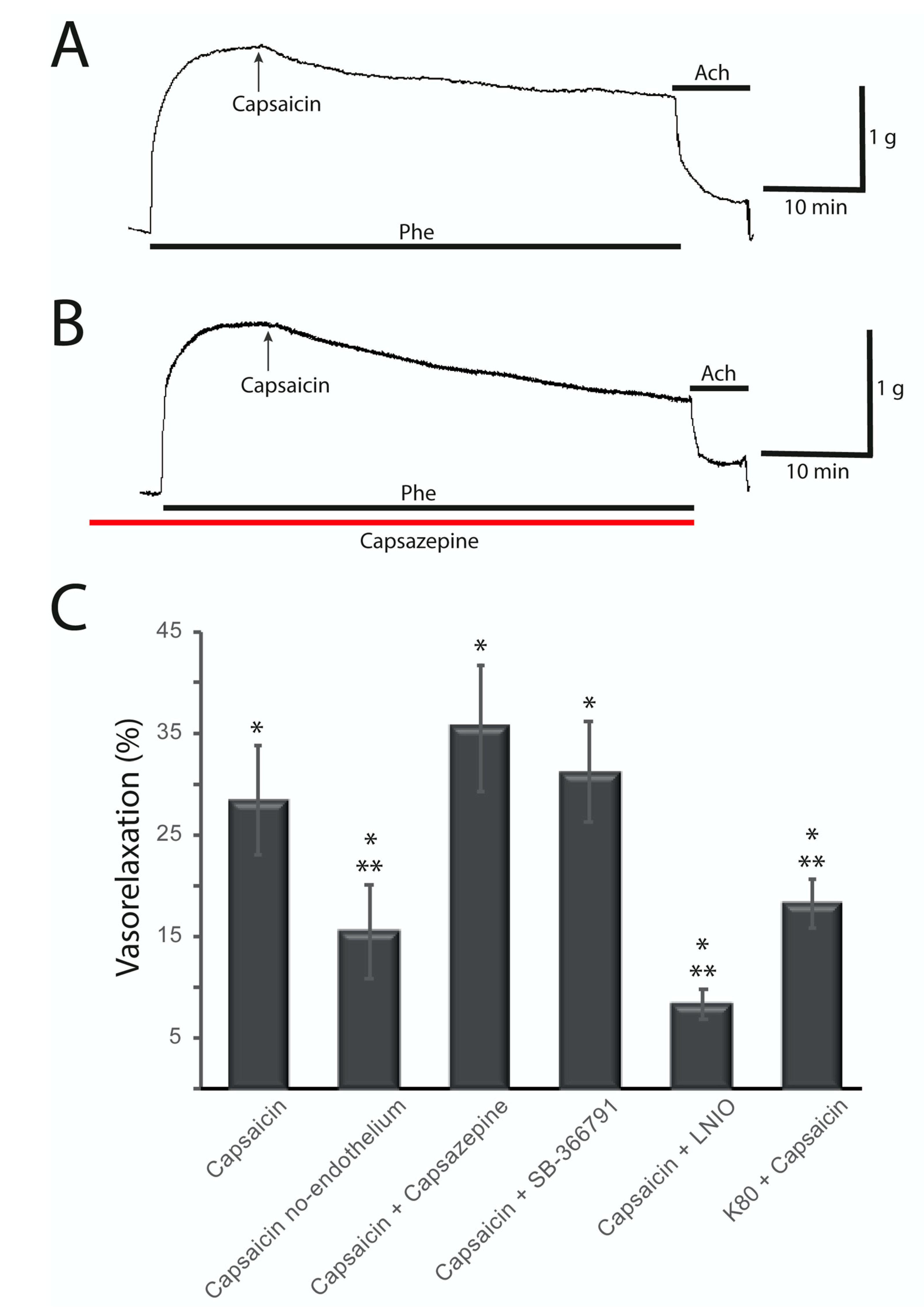
2.2. Capsaicin-Induced Vasorelaxation of the Rat Aorta Does not Involve TRPV1 Receptors and Is Partially Endothelium Dependent
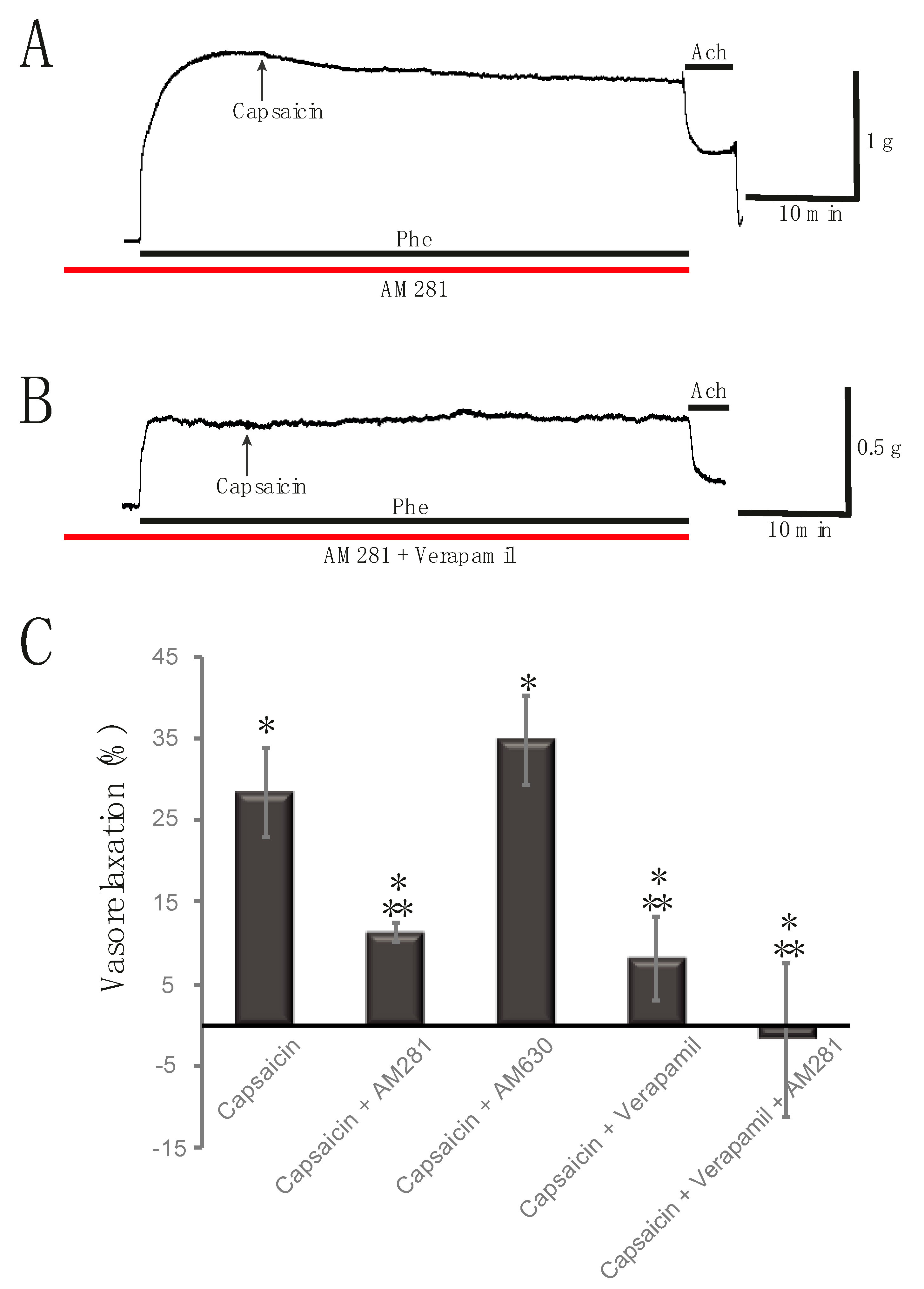
2.3. Capsaicin-Induced Rat Aortic Ring Vasorelaxation Involves CB1 Receptors and Voltage-Gated Calcium Channels
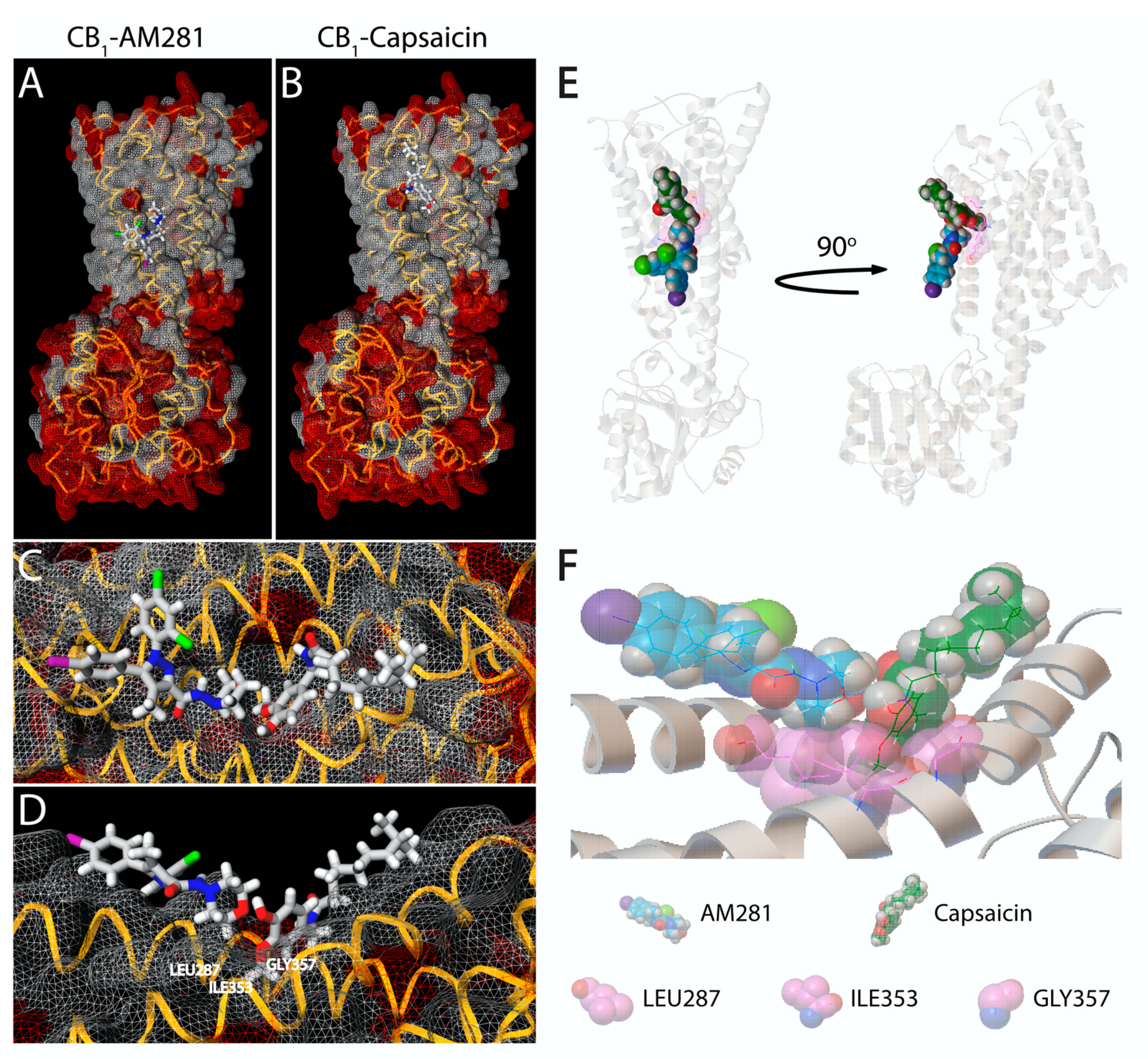
2.4. Capsaicin and AM281 Binding Site Regions in the Human CB1 Receptor Interpose at the Same Binding Cavity
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Dissection
4.3. Experimental Procedure
4.4. Immunolabeling and Confocal Microscopy
4.5. Solutions
4.6. Structure-Based Simulation of CB1 Receptor Ligands
4.7. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eichhorn, B.; Dobrev, D. Vascular large conductance calcium-activated potassium channels: Functional role and therapeutic potential. Naunyn. Schmiedebergs. Arch. Pharmacol. 2007, 376, 145–155. [Google Scholar] [CrossRef]
- Veronesi, B.; Oortgiesen, M. The TRPV1 receptor: Target of toxicants and therapeutics. Toxicol. Sci. 2006, 89, 1–3. [Google Scholar] [CrossRef]
- Bratz, I.N.; Dick, G.M.; Tune, J.D.; Edwards, J.M.; Neeb, Z.P.; Dincer, U.D.; Sturek, M. Impaired capsaicin-induced relaxation of coronary arteries in a porcine model of the metabolic syndrome. Am. J. Physiol.-Hear. Circ. Physiol. 2008, 294, H2489–H2496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Sullivan, S.E.; Kendall, D.A.; Randall, M.D. Vascular effects of Δ9-tetrahydrocannabinol (THC), anandamide and N-arachidonoyldopamine (NADA) in the rat isolated aorta. Eur. J. Pharmacol. 2005, 507, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Hopps, J.J.; Dunn, W.R.; Randall, M.D. Vasorelaxation to capsaicin and its effects on calcium influx in arteries. Eur. J. Pharmacol. 2012, 681, 88–93. [Google Scholar] [CrossRef]
- Bondarenko, A.I.; Montecucco, F.; Panasiuk, O.; Sagach, V.; Sidoryak, N.; Brandt, K.J.; Mach, F. GPR55 agonist lysophosphatidylinositol and lysophosphatidylcholine inhibit endothelial cell hyperpolarization via GPR-independent suppression of Na+-Ca2 + exchanger and endoplasmic reticulum Ca2 + refilling. Vascul. Pharmacol. 2017, 89, 39–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeon, D.; Kwon, S.; Lee, Y.; Leem, J.; Nam, T.; Ahn, D. Capsaicin-induced Relaxation in Rabbit Coronary Artery. J. Vet. Med. Sci. 2001, 63, 499–503. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Lozano-Cuenca, J.; Villalón, C.M.; De Vries, R.; Garrelds, I.M.; Avezaat, C.J.J.; Van Kats, J.P.; Saxena, P.R.; MaassenVanDenBrink, A. Pharmacological characterisation of capsaicin-induced relaxations in human and porcine isolated arteries. Naunyn. Schmiedebergs. Arch. Pharmacol. 2007, 375, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Tóth, A.; Czikora, Á.; Pásztor, E.T.; Dienes, B.; Bai, P.; Csernoch, L.; Rutkai, I.; Csató, V.; Mányiné, I.S.; Pórszász, R.; et al. Vanilloid Receptor-1 (TRPV1) Expression and Function in the Vasculature of the Rat. J. Histochem. Cytochem. 2014, 62, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Torres-Narváez, J.C.; Del Valle Mondragón, L.; López, E.V.; Pérez-Torres, I.; Díaz Juárez, J.A.; Suarez, J.; Hernández, G.P. Role of the transient receptor potential vanilloid type 1 receptor and stretch-activated ion channels in nitric oxide release from endothelial cells of the aorta and heart in rats. Exp. Clin. Cardiol. 2012, 17, 89–94. [Google Scholar]
- Gifford, A.N.; Tang, Y.; John Gatley, S.; Volkow, N.D.; Lan, R.; Makriyannis, A. Effect of the cannabinoid receptor SPECT agent, AM 281, on hippocampal acetylcholine release from rat brain slices. Neurosci. Lett. 1997, 238, 84–86. [Google Scholar] [CrossRef]
- Bisogno, T.; Hanuš, L.; De Petrocellis, L.; Tchilibon, S.; Ponde, D.E.; Brandi, I.; Moriello, A.S.; Davis, J.B.; Mechoulam, R.; Di Marzo, V. Molecular targets for cannabidiol and its synthetic analogues: Effect on vanilloid VR1 receptors and on the cellular uptake and enzymatic hydrolysis of anandamide. Br. J. Pharmacol. 2001, 134, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Zygmunt, P.M.; Petersson, J.; Andersson, D.A.; Chuang, H.H.; Sørgård, M.; Di Marzo, V.; Julius, D.; Högestätt, E.D. Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature 1999, 400, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Muller, C.; Morales, P.; Reggio, P.H. Cannabinoid ligands targeting TRP channels. Front. Mol. Neurosci. 2019, 11, 487. [Google Scholar] [CrossRef]
- Ligresti, A.; de Petrocellis, L.; de la Ossa, D.H.P.; Aberturas, R.; Cristino, L.; Moriello, A.S.; Finizio, A.; Gil, E.E.; Torres, A.I.; Molpeceres, J.; et al. Exploiting Nanotechnologies and TRPV1 Channels to Investigate the Putative Anandamide Membrane Transporter. PLoS ONE 2010, 5, e10239. [Google Scholar] [CrossRef] [Green Version]
- Raisinghani, M.; Pabbidi, R.M.; Premkumar, L.S. Activation of transient receptor potential vanilloid 1 (TRPV1) by resiniferatoxin. J. Physiol. 2005, 567, 771–786. [Google Scholar] [CrossRef]
- Dunn, W.R.; Hardy, T.A.; Brock, J.A. Electrophysiological effects of activating the peptidergic primary afferent innervation of rat mesenteric arteries. Br. J. Pharmacol. 2003, 140, 231–238. [Google Scholar] [CrossRef] [Green Version]
- Franco-Cereceda, A.; Henke, H.; Lundberg, J.M.; Petermann, J.B.; Hökfelt, T.; Fischer, J.A. Calcitonin gene-related peptide (CGRP) in capsaicin-sensitive substance P-immunoreactive sensory neurons in animals and man: Distribution and release by capsaicin. Peptides 1987, 8, 399–410. [Google Scholar] [CrossRef]
- Franco-Cereceda, A.; Rudehill, A. Capsaicin-induced vasodilatation of human coronary arteries in vitro is mediated by calcitonin gene-related peptide rather than substance P or neurokinin A. Acta Physiol. Scand. 1989, 136, 575–580. [Google Scholar] [CrossRef]
- Jansen, I.; Alafaci, C.; Uddman, R.; Edvinsson, L. Evidence that calcitonin gene-related peptide contributes to the capsaicin-induced relaxation of guinea pig cerebral arteries. Regul. Pept. 1990, 31, 167–178. [Google Scholar] [CrossRef]
- Poblete, I.M.; Orliac, M.L.; Briones, R.; Adler-Graschinsky, E.; Huidobro-Toro, J.P. Anandamide elicits an acute release of nitric oxide through endothelial TRPV1 receptor activation in the rat arterial mesenteric bed. J. Physiol. 2005, 568, 539–551. [Google Scholar] [CrossRef] [PubMed]
- Ralevic, V.; Kendall, D.A.; Randall, M.D.; Zygmunt, P.M.; Movahed, P.; Högestätt, E.D. Vanilloid receptors on capsaicin-sensitive sensory nerves mediate relaxation to methanandamide in the rat isolated mesenteric arterial bed and small mesenteric arteries. Br. J. Pharmacol. 2000, 130, 1483–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanessa Ho, W.-S.; Robin Hiley, C. Endothelium-independent relaxation to cannabinoids in rat-isolated mesenteric artery and role of Ca 2+ influx. Br. J. Pharmacol. 2003, 139, 585–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, R.; Vanessa Ho, W.S.V.; Bottrill, F.E.; Ford, W.R.; Hiley, C.R. Mechanisms of anandamide-induced vasorelaxation in rat isolated coronary arteries. Br. J. Pharmacol. 2001, 134, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Yi-Ching Lo; Sheng-Nan Wu; Jiunn-Ren Wu; Ing-Jun Chen Effect of capsaicin on membrane currents in cultured vascular smooth muscle cells of rat aorta. Eur. J. Pharmacol. Environ. Toxicol. 1995, 292, 321–328. [CrossRef]
- Docherty, R.J.; Yeats, J.C.; Piper, A.S. Capsazepine block of voltage-activated calcium channels in adult rat dorsal root ganglion neurones in culture. Br. J. Pharmacol. 1997, 121, 1461–1467. [Google Scholar] [CrossRef]
- Thakore, P.; Earley, S. Transient receptor potential channels and endothelial cell calcium signaling. Compr. Physiol. 2019, 9, 1249–1277. [Google Scholar] [CrossRef]
- Barthó, L.; Benkó, R.; Patacchini, R.; Pethö, G.; Holzer-Petsche, U.; Holzer, P.; Lázár, Z.; Undi, S.; Illényi, L.; Antal, A.; et al. Effects of capsaicin on visceral smooth muscle: A valuable tool for sensory neurotransmitter identification. Eur. J. Pharmacol. 2004, 500, 143–157. [Google Scholar] [CrossRef]
- Mitchell, J.A.; Williams, F.M.; Williams, T.J.; Larkin, S.W. Role of nitric oxide in the dilator actions of capsaicin-sensitive nerves in the rabbit coronary circulation. Neuropeptides 1997, 31, 333–338. [Google Scholar] [CrossRef]
- Diniz, C.R.A.F.; Biojone, C.; Joca, S.R.L.; Rantamäki, T.; Castrén, E.; Guimarães, F.S.; Casarotto, P.C. Dual mechanism of TRKB activation by anandamide through CB1 and TRPV1 receptors. PeerJ 2019, 7, e6493. [Google Scholar] [CrossRef]
- Moreira, F.A.; Wotjak, C.T. Cannabinoids and anxiety. Curr. Top. Behav. Neurosci. 2010, 2, 429–450. [Google Scholar] [CrossRef]
- Perchuk, A.; Bierbower, S.M.; Canseco-Alba, A.; Mora, Z.; Tyrell, L.; Joshi, N.; Schanz, N.; Gould, G.G.; Onaivi, E.S. Developmental and behavioral effects in neonatal and adult mice following prenatal activation of endocannabinoid receptors by capsaicin. Acta Pharmacol. Sin. 2019, 40, 418–424. [Google Scholar] [CrossRef] [Green Version]
- Fatahi, Z.; Reisi, Z.; Rainer, G.; Haghparast, A.; Khani, A. Cannabinoids induce apathetic and impulsive patterns of choice through CB1 receptors and TRPV1 channels. Neuropharmacology 2018, 133, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Ho, W.S.V.; Zheng, X.; Zhang, D.X. Role of endothelial TRPV4 channels in vascular actions of the endocannabinoid, 2-arachidonoylglycerol. Br. J. Pharmacol. 2015, 172, 5251–5264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhopadhyay, S.; Chapnick, B.M.; Howlett, A.C. Anandamide-induced vasorelaxation in rabbit aortic rings has two components: G protein dependent and independent. Am. J. Physiol. Hear. Circ. Physiol. 2002, 282, H2046–H2054. [Google Scholar] [CrossRef]
- Bhatta, P.; Dhukhwa, A.; Sheehan, K.; Al Aameri, R.F.H.; Borse, V.; Ghosh, S.; Sheth, S.; Mamillapalli, C.; Rybak, L.; Ramkumar, V.; et al. Capsaicin Protects Against Cisplatin Ototoxicity by Changing the STAT3/STAT1 Ratio and Activating Cannabinoid (CB2) Receptors in the Cochlea. Sci. Rep. 2019, 9, 4131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Dyck, E.; Andrade-Urzúa, F.; Elizalde, A.; Ferrer-Villada, T.; Dagnino-Acosta, A.; Huerta, M.; Osuna-Calleros, Z.; Rangel-Sandoval, C.; Sánchez-Pastor, E. ACPA and JWH-133 modulate the vascular tone of superior mesenteric arteries through cannabinoid receptors, BKCa channels, and nitric oxide dependent mechanisms. Pharmacol. Rep. 2017, 69, 1131–1139. [Google Scholar] [CrossRef]
- Sánchez-Pastor, E.; Andrade, F.; Sánchez-Pastor, J.M.; Elizalde, A.; Huerta, M.; Virgen-Ortiz, A.; Trujillo, X.; Rodríguez-Hernández, A. Cannabinoid receptor type 1 activation by arachidonylcyclopropylamide in rat aortic rings causes vasorelaxation involving calcium-activated potassium channel subunit alpha-1 and calcium channel, voltage-dependent, L type, alpha 1C subunit. Eur. J. Pharmacol. 2014, 729, 100–106. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Sanner, M.F. Python: A programming language for software integration and development. J. Mol. Graph. Model. 1999, 17, 57–61. [Google Scholar]
Sample Availability: Samples of the compounds are not available from the authors. |





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrade, F.; Rangel-Sandoval, C.; Rodríguez-Hernández, A.; López-Dyck, E.; Elizalde, A.; Virgen-Ortiz, A.; Bonales-Alatorre, E.; Valencia-Cruz, G.; Sánchez-Pastor, E. Capsaicin Causes Vasorelaxation of Rat Aorta through Blocking of L-type Ca2+ Channels and Activation of CB1 Receptors. Molecules 2020, 25, 3957. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25173957
Andrade F, Rangel-Sandoval C, Rodríguez-Hernández A, López-Dyck E, Elizalde A, Virgen-Ortiz A, Bonales-Alatorre E, Valencia-Cruz G, Sánchez-Pastor E. Capsaicin Causes Vasorelaxation of Rat Aorta through Blocking of L-type Ca2+ Channels and Activation of CB1 Receptors. Molecules. 2020; 25(17):3957. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25173957
Chicago/Turabian StyleAndrade, Felipa, Cinthia Rangel-Sandoval, Alejandrina Rodríguez-Hernández, Evelyn López-Dyck, Alejandro Elizalde, Adolfo Virgen-Ortiz, Edgar Bonales-Alatorre, Georgina Valencia-Cruz, and Enrique Sánchez-Pastor. 2020. "Capsaicin Causes Vasorelaxation of Rat Aorta through Blocking of L-type Ca2+ Channels and Activation of CB1 Receptors" Molecules 25, no. 17: 3957. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25173957