
Verbascoside-Rich Abeliophyllum distichum Nakai Leaf Extracts Prevent LPS-Induced Preterm Birth Through Inhibiting the Expression of Proinflammatory Cytokines from Macrophages and the Cell Death of Trophoblasts Induced by TNF-α

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Results
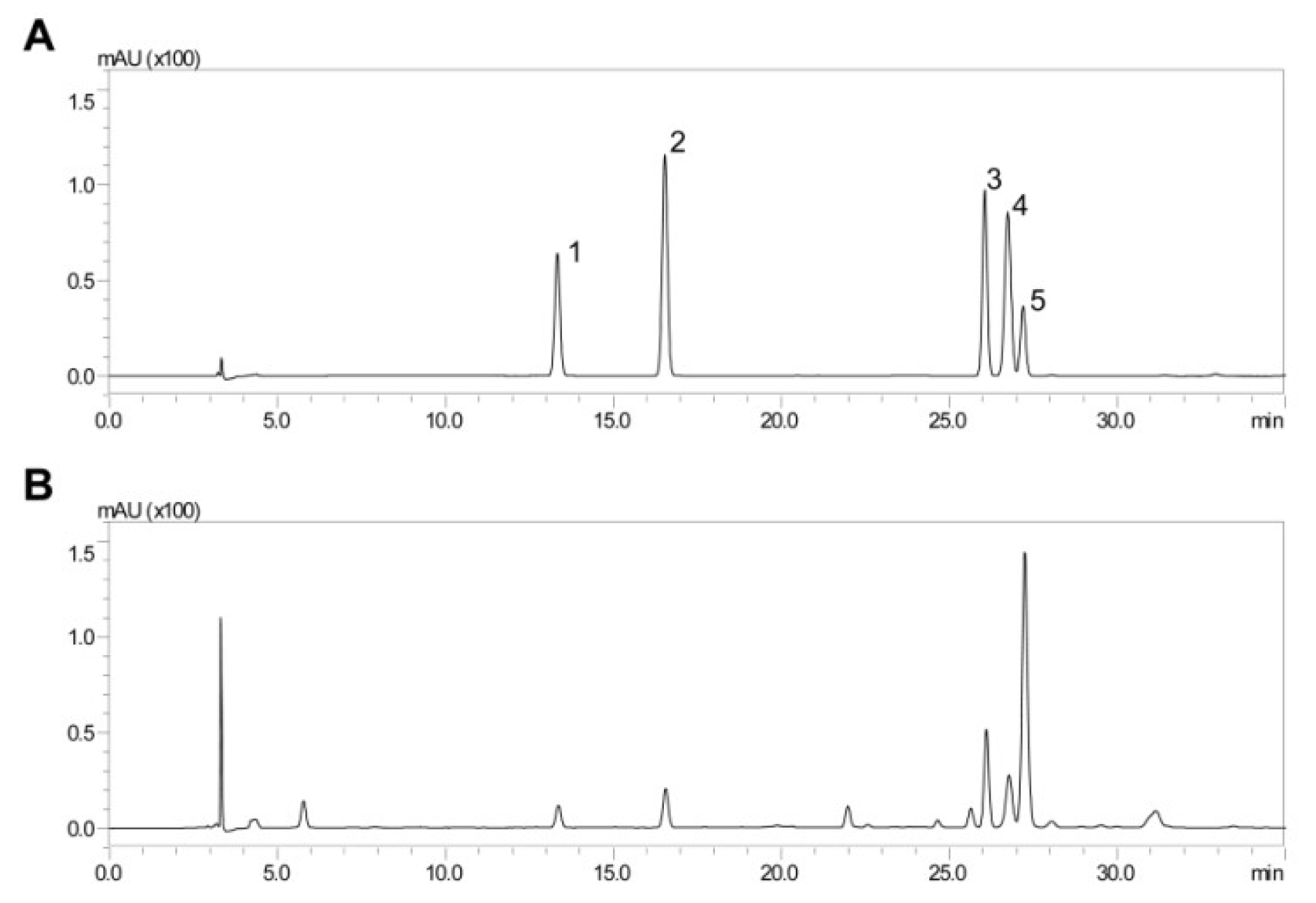
2.1. HPLC Analysis of ADL Extract
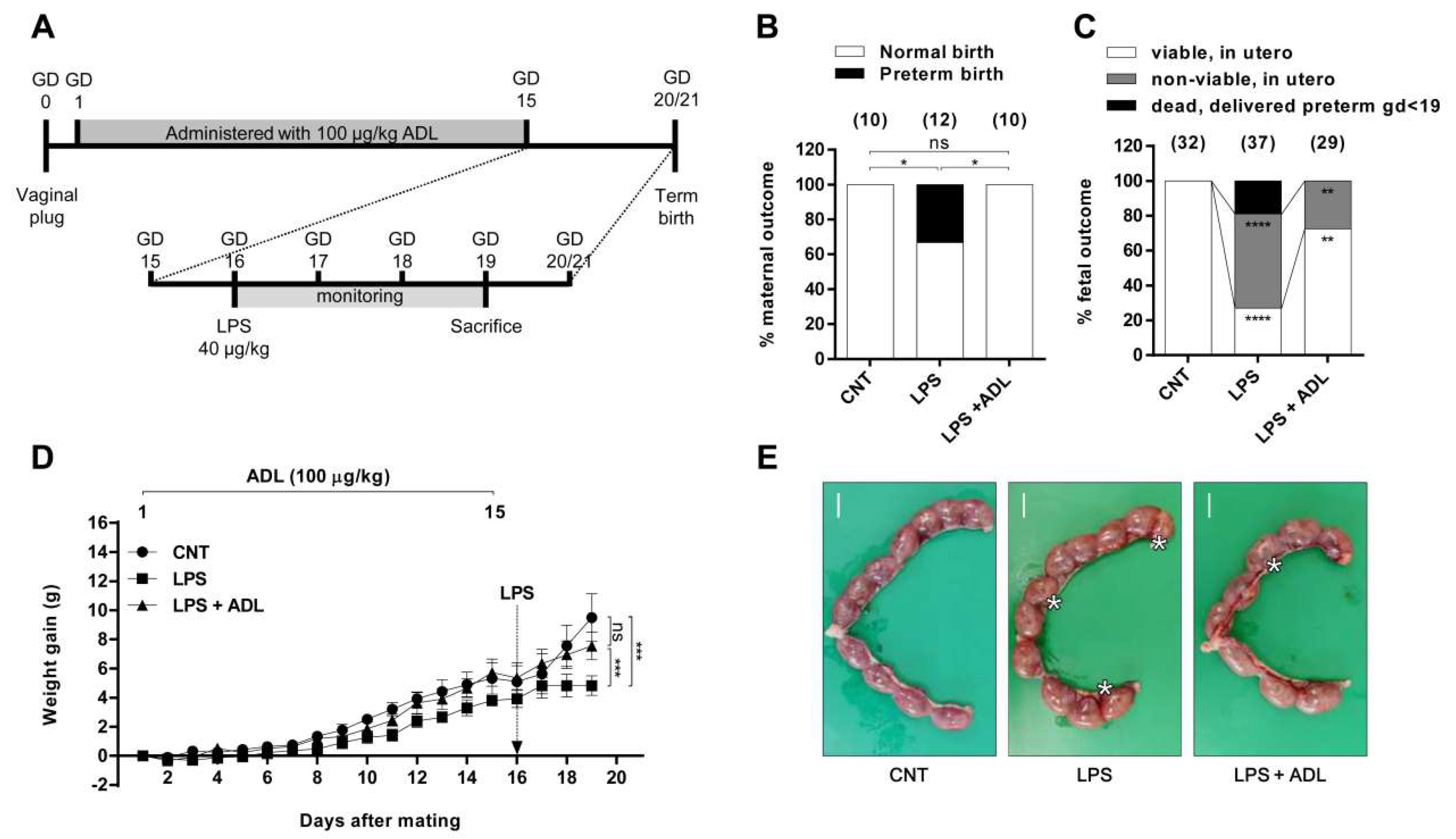
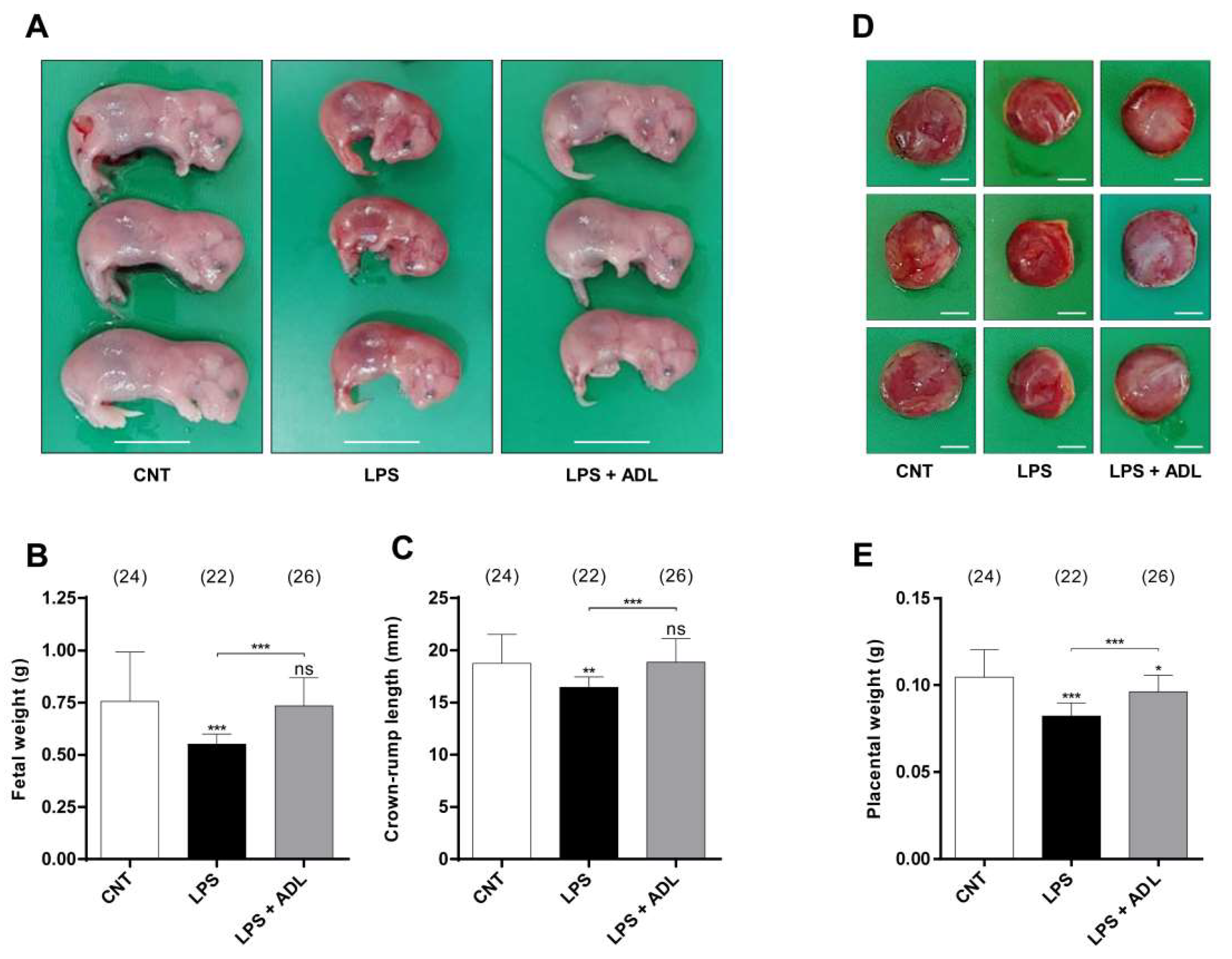
2.2. ADL Extract Prevents LPS-Induced Preterm Birth and Fetal Growth Restriction
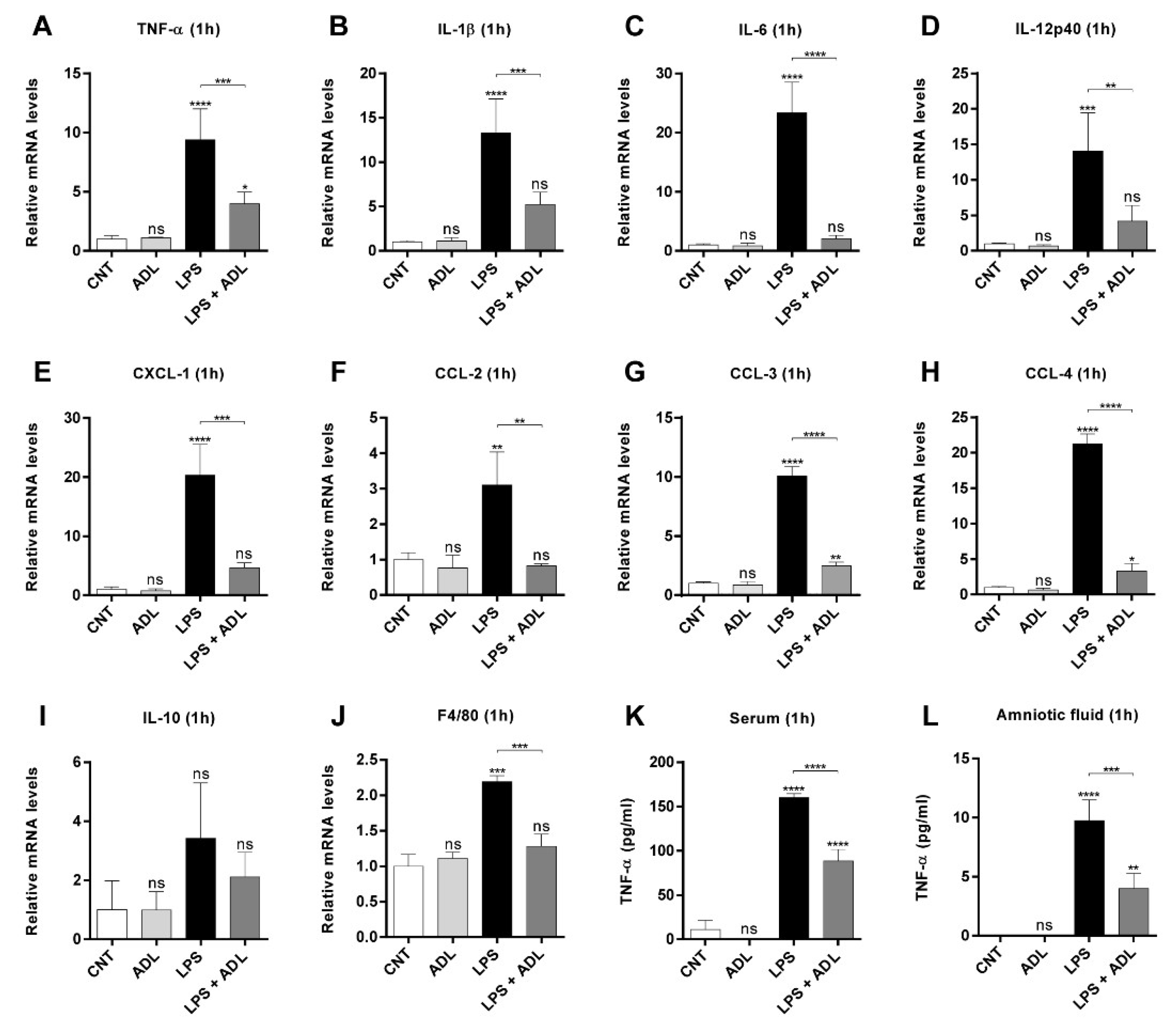
2.3. ADL Extract Pretreatment Downregulates LPS-Induced Expression of Proinflammatory Cytokines and Chemokines in Placental Tissue
2.4. ADL Extract Inhibits the Production of TNF-α in Macrophages by Regulating Mitogen-Activated Protein Kinases (MAPKs) and NF-κB
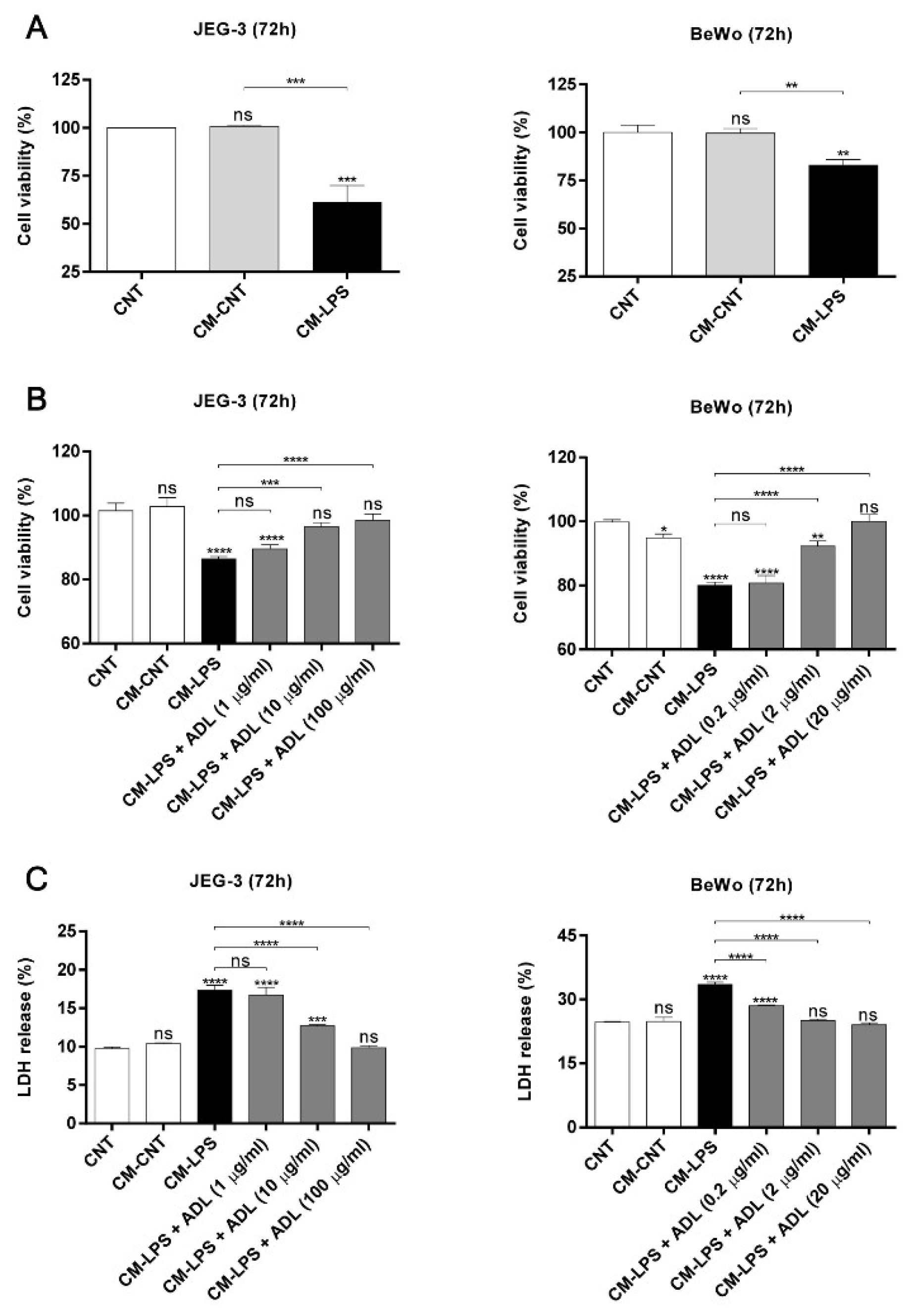
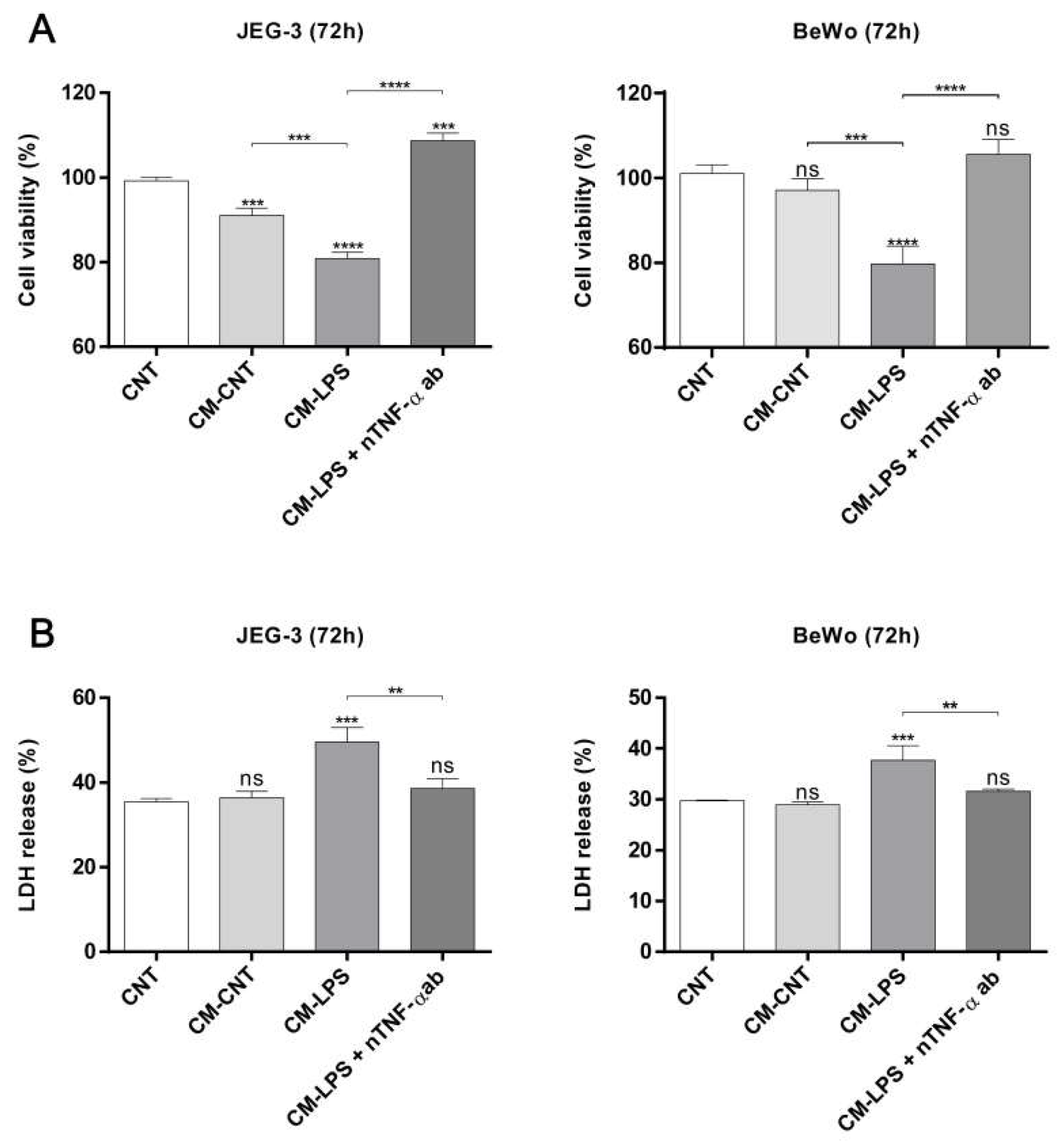
2.5. ADL Extract Inhibits TNF-α-Induced Trophoblast Cell Death Resulting from Exposure to LPS-Treated THP-1 Cell-Conditioned and Medium
2.6. Effect of ADL Extract on TNF-α-Induced Cell Death in Trophoblasts
3. Discussion
4. Materials and Methods
4.1. Plant Materials and HLPC Analysis
4.2. Mice
4.3. Animal Experiments
4.4. Cell Culture
4.5. Generation of Mouse Bone Marrow-Derived Macrophages
4.6. Conditioned Medium Preparation
4.7. Cell Viability and Cytotoxicity Assay
4.8. Quantification of mRNA by Real-Time RT-PCR
4.9. ELISA
4.10. Western Blot
4.11. Immunofluorescence
4.12. Assessment of Dead Cell Count
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Born Too Soon: The Global Action Report on Preterm Birth; WTO: Geneve, Switzerland, 2012.
- Romero, R.; Roslansky, P.; Oyarzun, E.; Wan, M.; Emamian, M.; Novitsky, T.J.; Gould, M.J.; Hobbins, J.C. Labor and infection: II. Bacterial endotoxin in amniotic fluid and its relationship to the onset of preterm labor. Am. J. Obstet. Gynecol. 1988, 158, 1044–1049. [Google Scholar] [CrossRef]
- Buhimschi, I.A.; Buhimschi, C.S.; Weiner, C.P. Protective effect of N-acetylcysteine against fetal death and preterm labor induced by maternal inflammation. Am. J. Obstet. Gynecol. 2003, 188, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Xu, D.X.; Wang, J.P.; Wang, H.; Wei, L.Z.; Sun, M.F.; Wei, W. Melatonin protects against lipopolysaccharide-induced intra-uterine fetal death and growth retardation in mice. J. Pineal Res. 2006, 40, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Rivera, D.; Olister, S.; Liu, X.; Thompson, J.; Zhang, X.J.; Pennline, K.; Azuero, R.; Clark, D.; Miller, M. Interleukin-10 attenuates experimental fetal growth restriction and demise. FASEB J. 1998, 12, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.-X.; Chen, Y.-H.; Wang, H.; Zhao, L.; Wang, J.-P.; Wei, W. Effect of N-acetylcysteine on lipopolysaccharide-induced intra-uterine fetal death and intra-uterine growth retardation in mice. Toxicol. Sci. 2005, 88, 525–533. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.-X.; Wang, H.; Zhao, L.; Ning, H.; Chen, Y.-H.; Zhang, C. Effects of low-dose lipopolysaccharide (LPS) pretreatment on LPS-induced intra-uterine fetal death and preterm labor. Toxicology 2007, 234, 167–175. [Google Scholar] [CrossRef]
- Xu, D.X.; Wang, H.; Ning, H.; Zhao, L.; Chen, Y.H. Maternally administered melatonin differentially regulates lipopolysaccharide-induced proinflammatory and anti-inflammatory cytokines in maternal serum, amniotic fluid, fetal liver, and fetal brain. J. Pineal Res. 2007, 43, 74–79. [Google Scholar] [CrossRef]
- Williams, P.; Searle, R.; Robson, S.; Innes, B.; Bulmer, J. Decidual leucocyte populations in early to late gestation normal human pregnancy. J. Reprod. Immunol. 2009, 82, 24–31. [Google Scholar] [CrossRef]
- Care, A.S.; Diener, K.R.; Jasper, M.J.; Brown, H.M.; Ingman, W.V.; Robertson, S.A. Macrophages regulate corpus luteum development during embryo implantation in mice. J. Clin. Investig. 2013, 123, 3472–3487. [Google Scholar] [CrossRef]
- Co, E.C.; Gormley, M.; Kapidzic, M.; Rosen, D.B.; Scott, M.A.; Stolp, H.A.; McMaster, M.; Lanier, L.L.; Bárcena, A.; Fisher, S.J. Maternal decidual macrophages inhibit NK cell killing of invasive cytotrophoblasts during human pregnancy. Biol. Reprod. 2013, 88, 151–159. [Google Scholar] [CrossRef]
- Erlebacher, A. Immunology of the maternal-fetal interface. Annu. Rev. Immunol. 2013, 31, 387–411. [Google Scholar] [CrossRef] [PubMed]
- Nagamatsu, T.; Schust, D.J. The Immunomodulatory Roles of Macrophages at the Maternal—Fetal Interface. Reprod. Sci. 2010, 17, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, J.M.; Franzke, C.-W.; Yang, F.; Romero, R.; Girardi, G. Complement activation triggers metalloproteinases release inducing cervical remodeling and preterm birth in mice. Am J. Pathol. 2011, 179, 838–849. [Google Scholar] [CrossRef] [PubMed]
- Guenther, S.; Vrekoussis, T.; Heublein, S.; Bayer, B.; Anz, D.; Knabl, J.; Navrozoglou, I.; Dian, D.; Friese, K.; Makrigiannakis, A. Decidual macrophages are significantly increased in spontaneous miscarriages and over-express FasL: A potential role for macrophages in trophoblast apoptosis. Int. J. Mol. Sci. 2012, 13, 9069–9080. [Google Scholar] [CrossRef]
- Schonkeren, D.; van der Hoorn, M.-L.; Khedoe, P.; Swings, G.; van Beelen, E.; Claas, F.; van Kooten, C.; de Heer, E.; Scherjon, S. Differential distribution and phenotype of decidual macrophages in preeclamptic versus control pregnancies. Am. J. Pathol. 2011, 178, 709–717. [Google Scholar] [CrossRef] [Green Version]
- Oh, H.; Kang, D.G.; Kwon, T.O.; Jang, K.K.; Chai, K.Y.; Yun, Y.G.; Chung, H.T.; Lee, H.S. Four glycosides from the leaves of Abeliophyllum distichum with inhibitory effects on angiotensin converting enzyme. Phytother. Res. 2003, 17, 811–813. [Google Scholar] [CrossRef]
- Park, G.H.; Park, J.H.; Eo, H.J.; Song, H.M.; Woo, S.H.; Kim, M.K.; Lee, J.W.; Lee, M.H.; Lee, J.R.; Koo, J.S. The induction of activating transcription factor 3 (ATF3) contributes to anti-cancer activity of Abeliophyllum distichum Nakai in human colorectal cancer cells. BMC Complement. Altern. Med. 2014, 14, 487. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [Green Version]
- Neta, G.I.; Von Ehrenstein, O.S.; Goldman, L.R.; Lum, K.; Sundaram, R.; Andrews, W.; Zhang, J. Umbilical cord serum cytokine levels and risks of small-for-gestational-age and preterm birth. Am. J. Epidemiol. 2010, 171, 859–867. [Google Scholar] [CrossRef] [Green Version]
- Rode, L.; Klein, K.; Larsen, H.; Holmskov, A.; Andreasen, K.R.; Uldbjerg, N.; Ramb, J.; Bødker, B.; Skibsted, L.; Sperling, L. Cytokines and the risk of preterm delivery in twin pregnancies. Obstet. Gynecol. 2012, 120, 60–68. [Google Scholar] [CrossRef]
- Duriez, M.; Quillay, H.; Madec, Y.; El Costa, H.; Cannou, C.; Marlin, R.; De Truchis, C.; Rahmati, M.; Barré-Sinoussi, F.; Nugeyre, M.-T. Human decidual macrophages and NK cells differentially express Toll-like receptors and display distinct cytokine profiles upon TLR stimulation. Front. Microbial. 2014, 5, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reister, F.; Frank, H.-G.; Kingdom, J.C.; Heyl, W.; Kaufmann, P.; Rath, W.; Huppertz, B. Macrophage-induced apoptosis limits endovascular trophoblast invasion in the uterine wall of preeclamptic women. Lab. Investig. 2001, 81, 1143–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilczyński, J.R.; Tchórzewski, H.; Głowacka, E.; Banasik, M.; Lewkowicz, P.; Szpakowski, M.; Zeman, K.; Wilczyński, J. Cytokine secretion by decidual lymphocytes in transient hypertension of pregnancy and pre-eclampsia. Med. Inflamm. 2002, 11, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Minas, V.; Jeschke, U.; Kalantaridou, S.; Richter, D.; Reimer, T.; Mylonas, I.; Friese, K.; Makrigiannakis, A. Abortion is associated with increased expression of FasL in decidual leukocytes and apoptosis of extravillous trophoblasts: A role for CRH and urocortin. Mol. Hum. Reprod. 2007, 13, 663–673. [Google Scholar] [CrossRef]
- Cotechini, T.; Komisarenko, M.; Sperou, A.; Macdonald-Goodfellow, S.; Adams, M.A.; Graham, C.H. Inflammation in rat pregnancy inhibits spiral artery remodeling leading to fetal growth restriction and features of preeclampsia. J. Exp. Med. 2014, 211, 165–179. [Google Scholar] [CrossRef]
- Gayle, D.A.; Beloosesky, R.; Desai, M.; Amidi, F.; Nunez, S.E.; Ross, M.G. Maternal LPS induces cytokines in the amniotic fluid and corticotropin releasing hormone in the fetal rat brain. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286, R1024–R1029. [Google Scholar] [CrossRef]
- Leazer, T.M.; Barbee, B.; Ebron-McCoy, M.; Henry-Sam, G.A.; Rogers, J.M. Role of the maternal acute phase response and tumor necrosis factor alpha in the developmental toxicity of lipopolysaccharide in the CD-1 mouse. Reprod. Toxicol. 2002, 16, 173–179. [Google Scholar] [CrossRef]
- Xu, D.-X.; Chen, Y.-H.; Wang, H.; Zhao, L.; Wang, J.-P.; Wei, W. Tumor necrosis factor alpha partially contributes to lipopolysaccharide-induced intra-uterine fetal growth restriction and skeletal development retardation in mice. Toxicol. Lett. 2006, 163, 20–29. [Google Scholar] [CrossRef]
- Al-Dhabi, N.A.; Valan Arasu, M.; Park, C.H.; Park, S.U. An up-to-date review of rutin and its biological and pharmacological activities. EXCLI J. 2015, 14, 59. [Google Scholar]
- Bazzaz, B.S.F.; Khameneh, B.; Ostad, M.R.Z.; Hosseinzadeh, H. In vitro evaluation of antibacterial activity of verbascoside, lemon verbena extract and caffeine in combination with gentamicin against drug-resistant Staphylococcus aureus and Escherichia coli clinical isolates. Avicenna J. Phytomed. 2018, 8, 246. [Google Scholar]
- Liu, M.; Song, S.; Li, H.; Jiang, X.; Yin, P.; Wan, C.; Liu, X.; Liu, F.; Xu, J. The protective effect of caffeic acid against inflammation injury of primary bovine mammary epithelial cells induced by lipopolysaccharide. J. Dairy Sci. 2014, 97, 2856–2865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, M.; Sudheer, A.R.; Menon, V.P. Ferulic acid: Therapeutic potential through its antioxidant property. J. Clin. Biochem. Nutr. 2007, 40, 92–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, R.; Wei, P. Studies on the antioxidant effect of rutin in the microenvironment of cationic micelles. Microchim. Acta 2008, 161, 233–239. [Google Scholar] [CrossRef]
- Korkmaz, A.; Kolankaya, D. Protective effect of rutin on the ischemia/reperfusion induced damage in rat kidney. J. Surg. Res. 2010, 164, 309–315. [Google Scholar] [CrossRef]
- Lee, S.-R.; Suh, S.-I.; Kim, S.-P. Protective effects of the green tea polyphenol (−)-epigallocatechin gallate against hippocampal neuronal damage after transient global ischemia in gerbils. Neurosci. Lett. 2000, 287, 191–194. [Google Scholar] [CrossRef]
- Novakovic, A.; Gojkovic-Bukarica, L.; Peric, M.; Nezic, D.; Djukanovic, B.; Markovic-Lipkovski, J.; Heinle, H. The mechanism of endothelium-independent relaxation induced by the wine polyphenol resveratrol in human internal mammary artery. J. Pharmacol. Sci. 2006, 101, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Pesce, M.; Franceschelli, S.; Ferrone, A.; De Lutiis, M.A.; Patruno, A.; Grilli, A.; Felaco, M.; Speranza, L. Verbascoside down-regulates some pro-inflammatory signal transduction pathways by increasing the activity of tyrosine phosphatase SHP-1 in the U937 cell line. J. Cell. Mol. Med. 2015, 19, 1548–1556. [Google Scholar] [CrossRef]
- Trinh, P.T.N.; Giang, B.L.; Tuan, N.T.; Hang, H.T.T.; Thuy, N.T.L.; Tuan, N.N.; Dung, L.T. Alfa glucosidase inhibitory, anti inflammatory activities and a new furanocoumarin derivative of Ruellia tuberosa. Natl. Prod. Res. 2019, 1–8. [Google Scholar] [CrossRef]
- Wu, Y.-S.; Shi, L.; Liu, X.-G.; Li, W.; Wang, R.; Huang, S.; Li, Y.; Yan, D.-L.; Wang, H.-Y.; Tian, Y. Chemical profiling of Callicarpa nudiflora and its effective compounds identification by compound-target network analysis. J. Pharm. Biomed. Anal. 2020, 182, 113110. [Google Scholar] [CrossRef]
- Khong, T. Immunohistologic study of the leukocytic infiltrate in maternal uterine tissues in normal and preeclamptic pregnancies at term. Am. J. Reprod. Immunol. Microbial. 1987, 15, 1–8. [Google Scholar] [CrossRef]
- Lessin, D.L.; Hunt, J.S.; King, C.R.; Wood, G.W. Antigen expression by cells near the maternal-fetal interface. Am. J. Reprod. Immunol. Microbiol. 1988, 16, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Vento-Tormo, R.; Efremova, M.; Botting, R.A.; Turco, M.Y.; Vento-Tormo, M.; Meyer, K.B.; Park, J.-E.; Stephenson, E.; Polański, K.; Goncalves, A. Single-cell reconstruction of the early maternal–fetal interface in humans. Nature 2018, 563, 347–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allaire, A.D.; Ballenger, K.A.; Wells, S.R.; McMahon, M.J.; Lessey, B.A. Placental apoptosis in preeclampsia. Obstet. Gynecol. 2000, 96, 271–276. [Google Scholar] [PubMed]
- Crocker, I.P.; Cooper, S.; Ong, S.C.; Baker, P.N. Differences in apoptotic susceptibility of cytotrophoblasts and syncytiotrophoblasts in normal pregnancy to those complicated with preeclampsia and intrauterine growth restriction. Am. J. Pathol. 2003, 162, 637–643. [Google Scholar] [CrossRef] [Green Version]
- DiFederico, E.; Genbacev, O.; Fisher, S.J. Preeclampsia is associated with widespread apoptosis of placental cytotrophoblasts within the uterine wall. Am. J. Pathol. 1999, 155, 293–301. [Google Scholar] [CrossRef] [Green Version]
- Genbacev, O.; DiFederico, E.; McMaster, M.; Fisher, S.J. Invasive cytotrophoblast apoptosis in pre-eclampsia. Hum. Reprod. 1999, 14, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Ishihara, N.; Matsuo, H.; Murakoshi, H.; Laoag-Fernandez, J.B.; Samoto, T.; Maruo, T. Increased apoptosis in the syncytiotrophoblast in human term placentas complicated by either preeclampsia or intrauterine growth retardation. Am. J. Obstet. Gynecol. 2002, 186, 158–166. [Google Scholar] [CrossRef]
- Smith, S.C.; Baker, P.N.; Symonds, E.M. Increased placental apoptosis in intrauterine growth restriction. Am. J. Obstet. Gynecol. 1997, 177, 1395–1401. [Google Scholar] [CrossRef]
- Pijnenborg, R.; McLaughlin, P.; Vercruysse, L.; Hanssens, M.; Johnson, P.; Keith Jr, J.; Van Assche, F.A. Immunolocalization of tumour necrosis factor-α (TNF-α) in the placental bed of normotensive and hypertensive human pregnancies. Placenta 1998, 19, 231–239. [Google Scholar] [CrossRef]
- Edey, L.F.; O’Dea, K.P.; Herbert, B.R.; Hua, R.; Waddington, S.N.; MacIntyre, D.A.; Bennett, P.R.; Takata, M.; Johnson, M.R. The local and systemic immune response to intrauterine LPS in the prepartum mouse. Biol. Reprod. 2016, 95, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Toyama, R.P.; Xikota, J.C.; Schwarzbold, M.L.; Frode, T.S.; Buss, Z.d.S.; Nunes, J.C.; Funchal, G.D.G.; Nunes, F.C.; Walz, R.; Pires, M.M.d.S. Dose-dependent sickness behavior, abortion and inflammation induced by systemic LPS injection in pregnant mice. J. Matern. Fetal Neonatal Med. 2015, 28, 426–430. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Compounds | Contents (μg/mg) |
|---|---|
| Chlorogenic acid | 11.60 ± 0.02 |
| Caffeic acid | 11.26 ± 0.02 |
| Rutin | 32.72 ± 0.01 |
| Ferulic acid | 20.70 ± 0.04 |
| Verbascoside | 260.43 ± 0.10 |
| Mean ± SD (n = 3) | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.W.; Yu, A.-R.; Kang, M.; Sung, N.-Y.; Lee, B.S.; Park, S.-Y.; Han, I.-J.; Kim, D.-S.; Oh, S.-M.; Lee, Y.I.; et al. Verbascoside-Rich Abeliophyllum distichum Nakai Leaf Extracts Prevent LPS-Induced Preterm Birth Through Inhibiting the Expression of Proinflammatory Cytokines from Macrophages and the Cell Death of Trophoblasts Induced by TNF-α. Molecules 2020, 25, 4579. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25194579
Kim HW, Yu A-R, Kang M, Sung N-Y, Lee BS, Park S-Y, Han I-J, Kim D-S, Oh S-M, Lee YI, et al. Verbascoside-Rich Abeliophyllum distichum Nakai Leaf Extracts Prevent LPS-Induced Preterm Birth Through Inhibiting the Expression of Proinflammatory Cytokines from Macrophages and the Cell Death of Trophoblasts Induced by TNF-α. Molecules. 2020; 25(19):4579. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25194579
Chicago/Turabian StyleKim, Ho Won, A-Reum Yu, Minji Kang, Nak-Yun Sung, Byung Soo Lee, Sang-Yun Park, In-Jun Han, Dong-Sub Kim, Sang-Muk Oh, Young Ik Lee, and et al. 2020. "Verbascoside-Rich Abeliophyllum distichum Nakai Leaf Extracts Prevent LPS-Induced Preterm Birth Through Inhibiting the Expression of Proinflammatory Cytokines from Macrophages and the Cell Death of Trophoblasts Induced by TNF-α" Molecules 25, no. 19: 4579. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25194579