
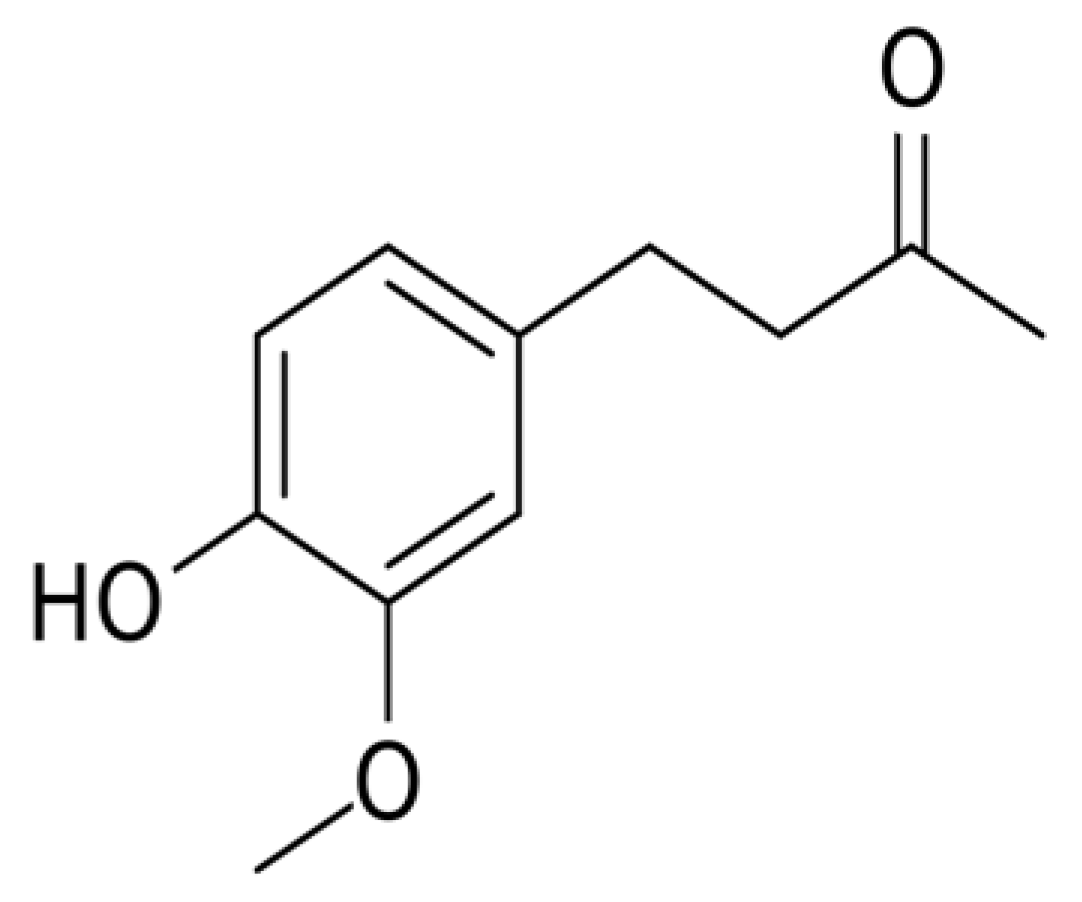
Zingerone [4-(3-Methoxy-4-hydroxyphenyl)-butan-2] Attenuates Lipopolysaccharide-Induced Inflammation and Protects Rats from Sepsis Associated Multi Organ Damage

 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Results
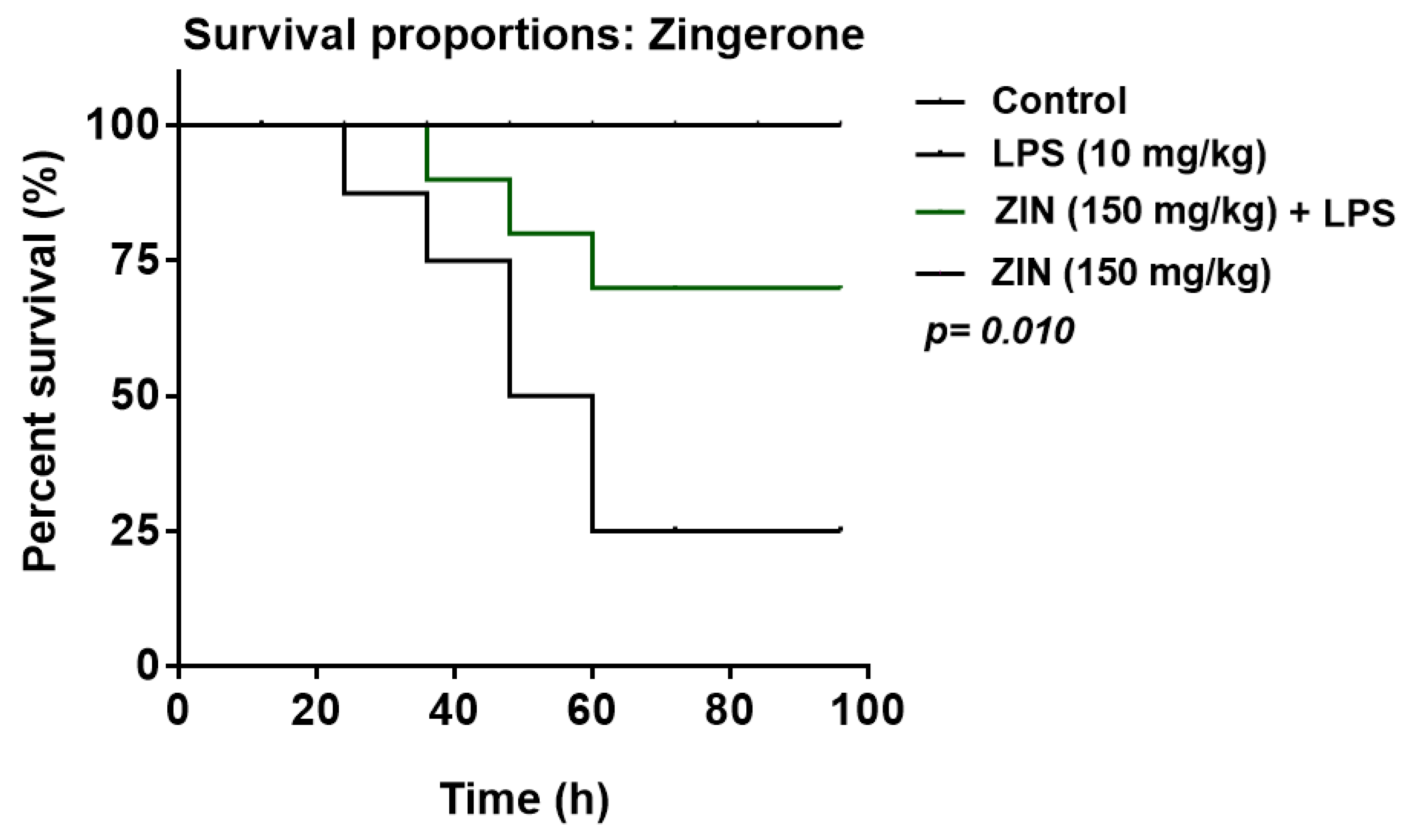
2.1. Effect of ZIN on Survival Rate of Animals
2.2. Effect of ZIN on Biochemical Markers
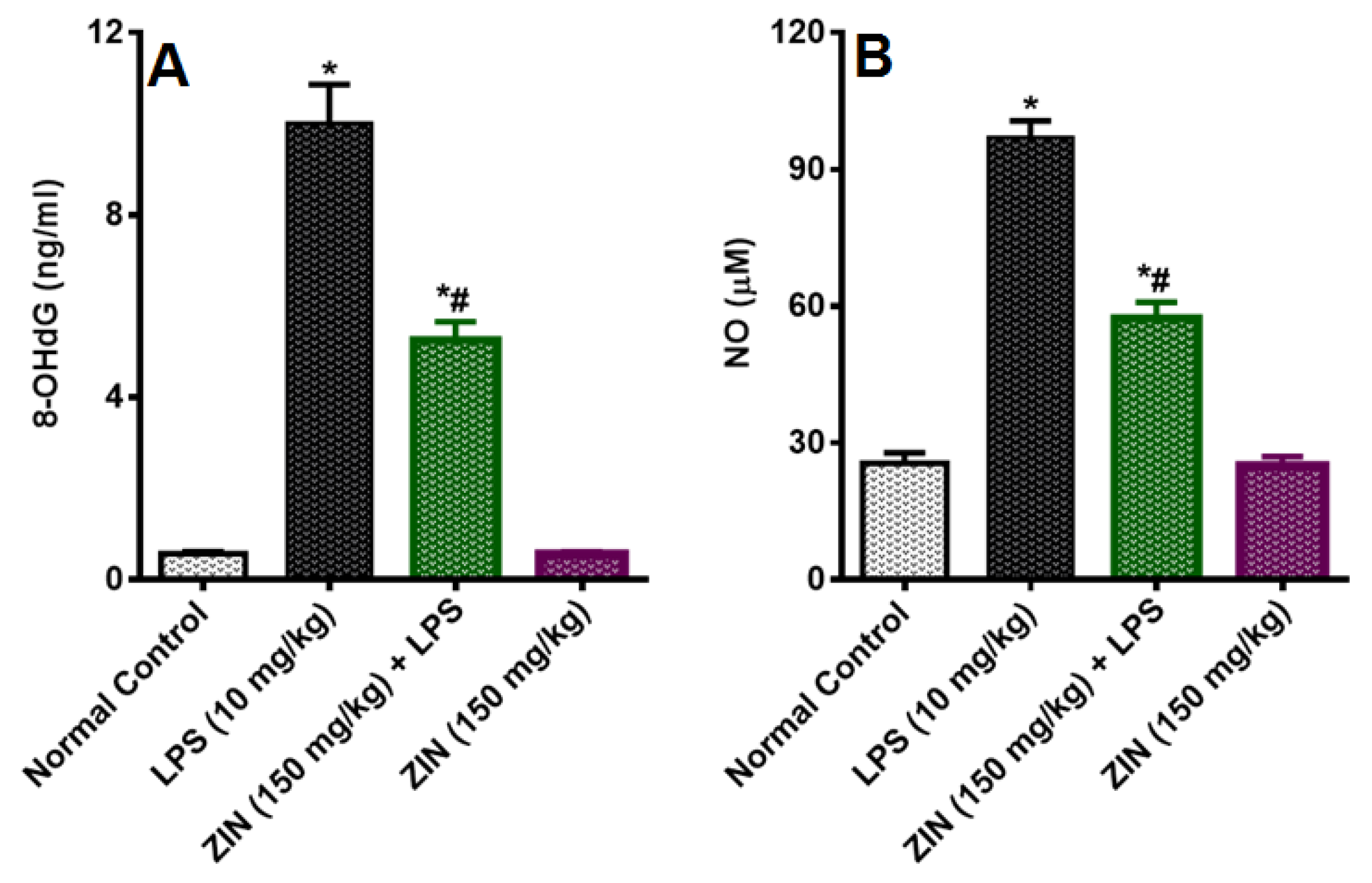
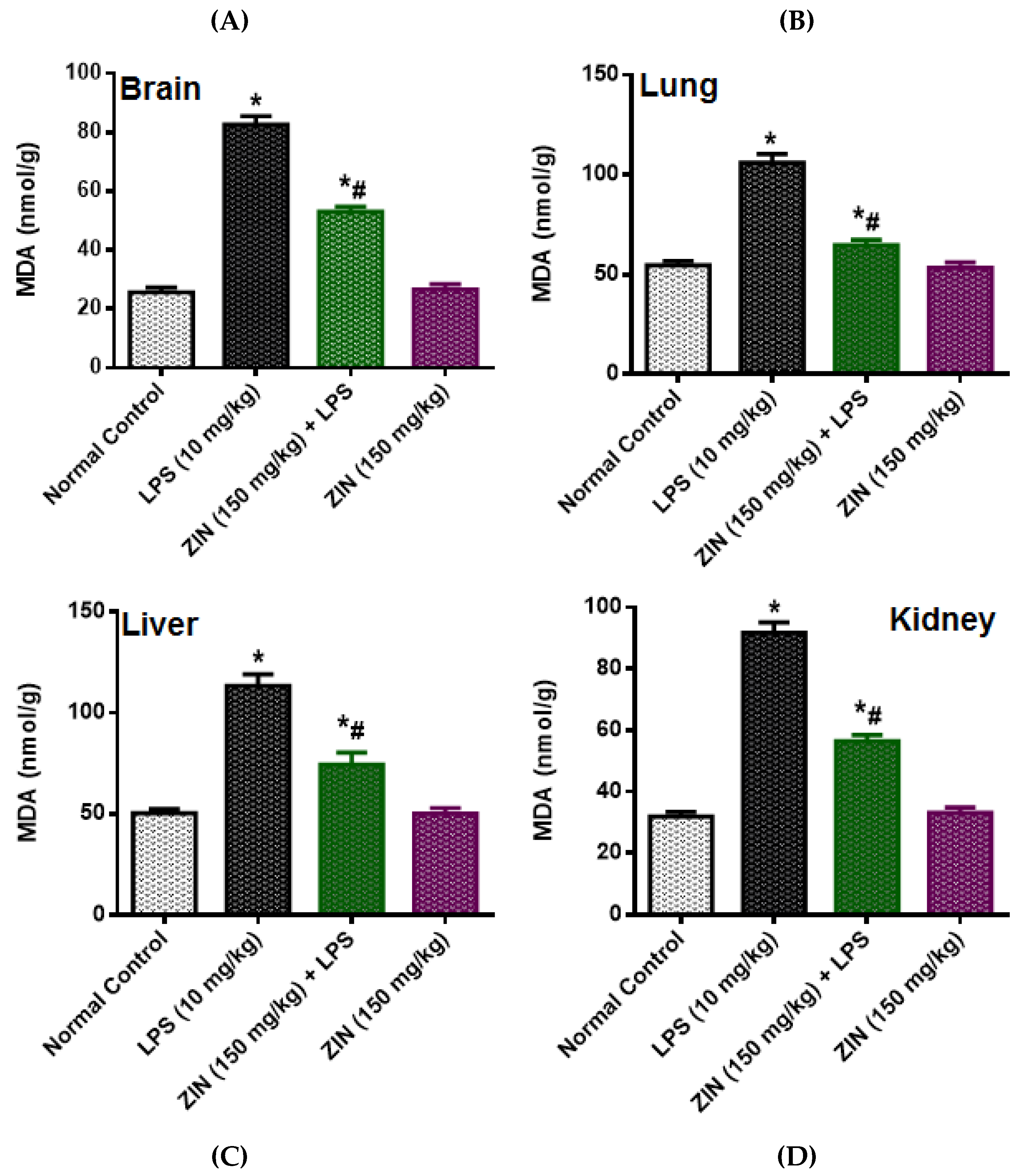
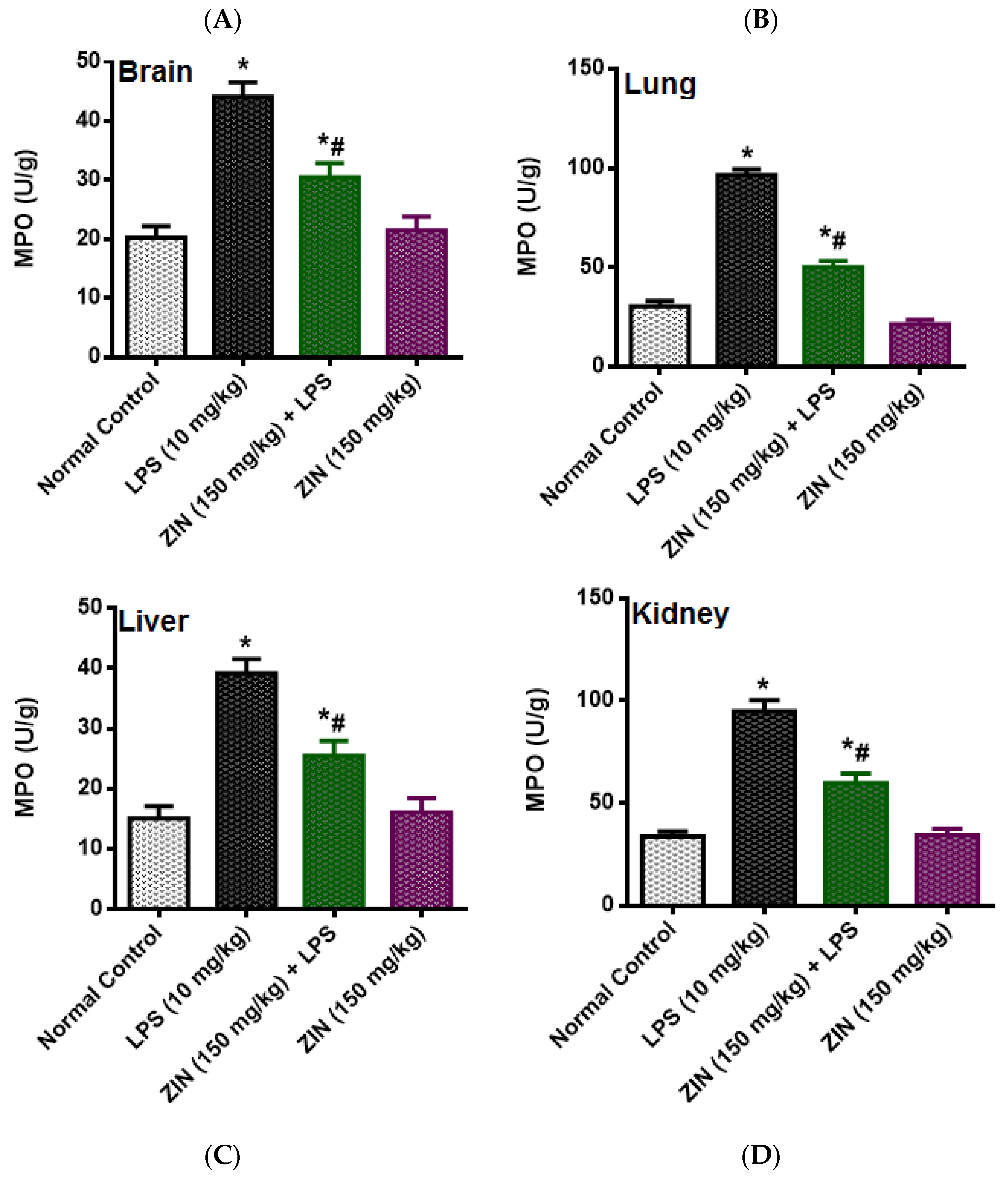
2.3. Effect of ZIN on Oxidative Stress and Antioxidant Enzyme Markers
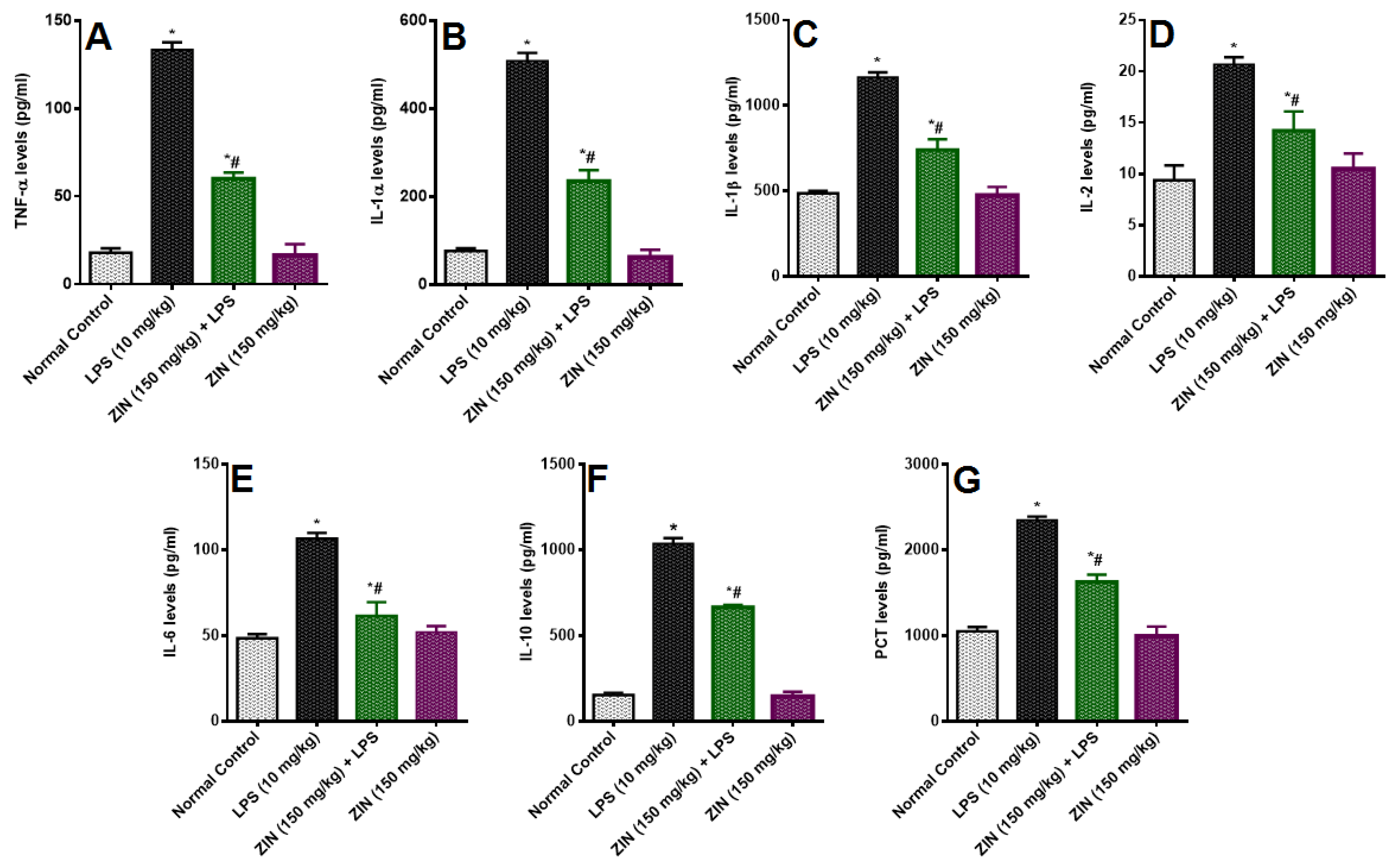
2.4. Effect of ZIN on Cytokines and Inflammatory Markers
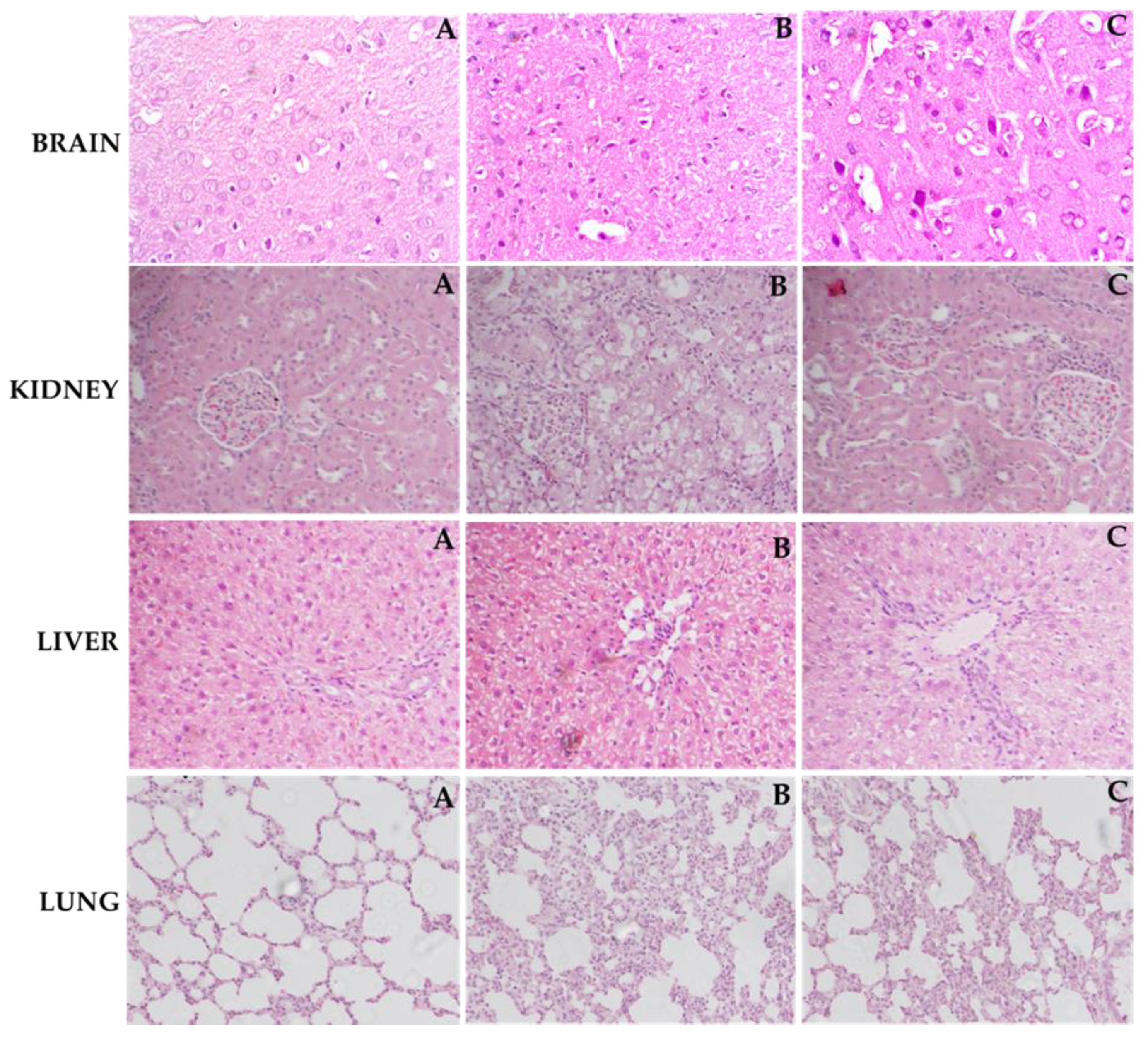
2.5. Effect of ZIN on Histoarchitecture of Different Organs
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Animals
4.3. LPS-Induced Endotoxemia and Survival Study
4.4. Experimental Model
4.5. Determination of Biochemical, Oxidative Stress and Inflammatory Markers
4.6. Histological Evaluation
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- El-Seedi, H.R.; Khalifa, S.A.; Yosri, N.; Khatib, A.; Chen, L.; Saeed, A.; Efferth, T.; Verpoorte, R. Plants mentioned in the Islamic Scriptures (Holy Qur’ân and Ahadith): Traditional uses and medicinal importance in contemporary times. J. Ethnopharmacol. 2019, 243, 112007. [Google Scholar] [CrossRef]
- Ahmad, B.; Rehman, M.U.; Amin, I.; Arif, A.; Rasool, S.; Bhat, S.A.; Afzal, I.; Hussain, I.; Bilal, S.; Mir, M.U.R. A Review on Pharmacological Properties of Zingerone (4-(4-Hydroxy-3-methoxyphenyl)-2-butanone). Sci. World J. 2015, 2015, 816364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takizawa, M.; Sato, M.; Kusuoku, H.; Sakasai, M. Lipolysis Stimulator. U.S. Patent US008197859B2, 12 June 2012. [Google Scholar]
- Moran, A.P.; Prendergast, M.M.; Appelmelk, B.J. Molecular mimicry of host structures by bacterial lipopolysaccharides and its contribution to disease. FEMS Immunol. Med. Mic. 1996, 16, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Frantz, S.; Kobzik, L.; Kim, Y.-D.; Fukazawa, R.; Medzhitov, R.; Lee, R.T.; Kelly, R.A. Toll4 (TLR4) expression in cardiac myocytes in normal and failing myocardium. J. Clin. Investig. 1999, 104, 271–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavener, S.A.; Long, E.M.; Robbins, S.M.; McRae, K.M.; Van Remmen, H.; Kubes, P. Immune Cell Toll-Like Receptor 4 Is Required for Cardiac Myocyte Impairment during Endotoxemia. Circ. Res. 2004, 95, 700–707. [Google Scholar] [CrossRef]
- Sweet, M.J.; Hume, D.A. Endotoxin signal transduction in macrophages. J. Leukoc. Biol. 1996, 60, 8–26. [Google Scholar] [CrossRef]
- Di Penta, A.; Moreno, B.; Reix, S.; Fernandez-Diez, B.; Villanueva, M.; Errea, O.; Escala, N.; Vandenbroeck, K.; Comella, J.X.; Villoslada, P. Oxidative Stress and Proinflammatory Cytokines Contribute to Demyelination and Axonal Damage in a Cerebellar Culture Model of Neuroinflammation. PLoS ONE 2013, 8, e54722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Victor, V.M.; Rocha, M.; De La Fuente, M. Immune cells: Free radicals and antioxidants in sepsis. Int. Immunopharmacol. 2004, 4, 327–347. [Google Scholar] [CrossRef]
- Kallapura, G.; Pumford, N.R.; Hernandez-Velasco, X.; Hargis, B.; Tellez, G. Mechanisms Involved in Lipopolysaccharide Derived ROS and RNS Oxidative Stress and Septic Shock. J. Microbiol. Res. Rev. 2014, 2, 6–11. [Google Scholar]
- Sena, L.A.; Chandel, N.S. Physiological Roles of Mitochondrial Reactive Oxygen Species. Mol. Cell 2012, 48, 158–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forni, C.; Facchiano, F.; Bartoli, M.; Pieretti, S.; Facchiano, A.; D’Arcangelo, D.; Norelli, S.; Valle, G.; Nisini, R.; Beninati, S.; et al. Beneficial Role of Phytochemicals on Oxidative Stress and Age-Related Diseases. BioMed Res. Int. 2019, 2019, 8748253. [Google Scholar] [CrossRef] [Green Version]
- Rajan, I.; Narayanan, N.; Rabindran, R.; Jayasree, P.R.; Kumar, P.R.M. Zingerone Protects Against Stannous Chloride-Induced and Hydrogen Peroxide-Induced Oxidative DNA Damage in Vitro. Biol. Trace Element Res. 2013, 155, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Mu, E.; Ding, R.; An, X.; Li, X.; Chen, S.; Ma, X. Heparin attenuates lipopolysaccharide-induced acute lung injury by inhibiting nitric oxide synthase and TGF-β/Smad signaling pathway. Thromb. Res. 2012, 129, 479–485. [Google Scholar] [CrossRef]
- Amin, I.; Hussain, I.; Rehman, M.U.; Mir, B.A.; Ganaie, S.A.; Ahmad, S.B.; Mir, M.U.R.; Shanaz, S.; Muzamil, S.; Arafah, A.; et al. Zingerone prevents lead-induced toxicity in liver and kidney tissues by regulating the oxidative damage in Wistar rats. J. Food Biochem. 2020, e13241. [Google Scholar] [CrossRef]
- Kaur, G.; Tirkey, N.; Chopra, K. Beneficial effect of hesperidin on lipopolysaccharide-induced hepatotoxicity. Toxicology 2006, 226, 152–160. [Google Scholar] [CrossRef]
- Kamisaki, Y.; Wada, K.; Ataka, M.; Yamada, Y.; Nakamoto, K.; Ashida, K.; Kishimoto, Y. Lipopolysaccharide-induced increase in plasma nitrotyrosine concentrations in rats. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 1997, 1362, 24–28. [Google Scholar] [CrossRef] [Green Version]
- Sethi, S.; Dikshit, M. Modulation of Polymorphonuclear Leukocytes Function by Nitric Oxide. Thromb. Res. 2000, 100, 223–247. [Google Scholar] [CrossRef]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric Oxide and Peroxynitrite in Health and Disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [Green Version]
- Alkharfy, K.M.; Ahmad, A.; Raish, M.; Vanhoutte, P.M. Thymoquinone modulates nitric oxide production and improves organ dysfunction of sepsis. Life Sci. 2015, 143, 131–138. [Google Scholar] [CrossRef]
- Halliwell, B. Oxidative stress and neurodegeneration: Where are we now? J. Neurochem. 2006, 97, 1634–1658. [Google Scholar] [CrossRef]
- van Klaveren, R.J.; Demedts, M.; Nemery, B. Cellular glutathione turnover in vitro, with emphasis on type II pneumocytes. Eur. Respir. J. 1997, 10, 1392–1400. [Google Scholar] [CrossRef] [Green Version]
- Luppi, F.; Aarbiou, J.; Van Wetering, S.; Rahman, I.; I De Boer, W.; Rabe, K.F.; Hiemstra, P. Effects of cigarette smoke condensate on proliferation and wound closure of bronchial epithelial cells in vitro: Role of glutathione. Respir. Res. 2005, 6, 140. [Google Scholar] [CrossRef] [Green Version]
- Vargas, M.R.; Johnson, D.A.; Johnson, J.A. Decreased glutathione accelerates neurological deficit and mitochondrial pathology in familial ALS-linked hSOD1G93A mice model. Neurobiol. Dis. 2011, 43, 543–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.; Ji, W.; Fu, Q.; Ma, S.-P. Formononetin Inhibited the Inflammation of LPS-Induced Acute Lung Injury in Mice Associated with Induction of PPAR Gamma Expression. Inflammation 2013, 36, 1560–1566. [Google Scholar] [CrossRef]
- Mills-Davies, N.; Butler, D.; Norton, E.; Thompson, D.; Sarwar, M.I.; Guo, J.; Gill, R.; Azim, N.; Coker, A.R.; Wood, S.P.; et al. Structural studies of substrate and product complexes of 5-aminolaevulinic acid dehydratase from humans, Escherichia coli and the hyperthermophile Pyrobaculum calidifontis. Acta Crystallogr. Sect. D Struct. Biol. 2017, 73, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Sarker, S.; Vashistha, D.; Sarker, M.S.; Sarkar, A. DNA damage in marine rock oyster (Saccostrea Cucullata) exposed to environmentally available PAHs and heavy metals along the Arabian Sea coast. Ecotoxicol. Environ. Saf. 2018, 151, 132–143. [Google Scholar] [CrossRef]
- Torun, A.N.; Kulaksizoglu, S.; Kulaksizoglu, M.; Pamuk, B.O.; Isbilen, E.; Tutuncu, N.B. Serum total antioxidant status and lipid peroxidation marker malondialdehyde levels in overt and subclinical hypothyroidism. Clin. Endocrinol. 2009, 70, 469–474. [Google Scholar] [CrossRef]
- Vaibhav, K.; Shrivastava, P.; Tabassum, R.; Khan, A.; Javed, H.; Ahmed, E.; Islam, F.; Safhi, M.M.; Islam, F. Delayed administration of zingerone mitigates the behavioral and histological alteration via repression of oxidative stress and intrinsic programmed cell death in focal transient ischemic rats. Pharmacol. Biochem. Behav. 2013, 113, 53–62. [Google Scholar] [CrossRef]
- Shin, S.-G.; Kim, J.Y.; Chung, H.Y.; Jeong, J.-C. Zingerone as an Antioxidant against Peroxynitrite. J. Agric. Food Chem. 2005, 53, 7617–7622. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2′ -deoxyguanosine (8-OHdG): A Critical Biomarker of Oxidative Stress and Carcinogenesis. J. Environ. Sci. Health Part C 2009, 27, 120–139. [Google Scholar] [CrossRef] [Green Version]
- Haldar, S.; Dru, C.; Choudhury, D.; Mishra, R.; Fernández, A.; Biondi, S.; Liu, Z.; Shimada, K.; Arditi, M.; Bhowmick, N.A. Inflammation and Pyroptosis Mediate Muscle Expansion in an Interleukin-1β (IL-1β)-dependent Manner. J. Biol. Chem. 2015, 290, 6574–6583. [Google Scholar] [CrossRef] [Green Version]
- Ghezzi, P.; Cerami, A.; Corti, A. Tumor Necrosis Factor as a Pharmacological Target. Tumor Necrosis Factor 2004, 98, 1–8. [Google Scholar] [CrossRef]
- Faix, J.D. Biomarkers of sepsis. Crit. Rev. Clin. Lab. Sci. 2013, 50, 23–36. [Google Scholar] [CrossRef]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef] [Green Version]
- Parameswaran, N.; Patial, S. Tumor Necrosis Factor-α Signaling in Macrophages. Crit. Rev. Eukaryot. Gene Expr. 2010, 20, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Cannon, J.G.; Tompkins, R.G.; Gelfand, J.A.; Michie, H.R.; Stanford, G.G.; Van Der Meer, J.W.M.; Endres, S.; Lonnemann, G.; Corsetti, J.; Chernow, B.; et al. Circulating Interleukin-1 and Tumor Necrosis Factor in Septic Shock and Experimental Endotoxin Fever. J. Infect. Dis. 1990, 161, 79–84. [Google Scholar] [CrossRef]
- Michie, H.R.; Manogue, K.R.; Spriggs, D.R.; Revhaug, A.; O’Dwyer, S.; Dinarello, C.A.; Cerami, A.; Wolff, S.M.; Wilmore, D.W. Detection of Circulating Tumor Necrosis Factor after Endotoxin Administration. N. Engl. J. Med. 1988, 318, 1481–1486. [Google Scholar] [CrossRef]
- Conte, D.; Holcik, M.; Lefebvre, C.A.; Lacasse, E.; Picketts, D.J.; Wright, K.E.; Korneluk, R.G. Inhibitor of Apoptosis Protein cIAP2 Is Essential for Lipopolysaccharide-Induced Macrophage Survival. Mol. Cell. Biol. 2006, 26, 699–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, J. The immunopathogenesis of sepsis. Nat. Cell Biol. 2002, 420, 885–891. [Google Scholar] [CrossRef]
- Pestka, S.; Krause, C.D.; Sarkar, D.; Walter, M.R.; Shi, Y.; Fisher, P.B. Interleukin-10 and Related Cytokines and Receptors. Annu. Rev. Immunol. 2004, 22, 929–979. [Google Scholar] [CrossRef]
- Zhou, X.; Schmidtke, P.; Zepp, F.; Meyer, C.U. Boosting Interleukin-10 Production: Therapeutic Effects and Mechanisms. Curr. Drug Targets-Immune Endocr. Metab. Disord. 2005, 5, 465–475. [Google Scholar] [CrossRef]
- Vijayan, A.L.; Ravindran, S.; Saikant, R.; Lakshmi, S.; Kartik, R. Procalcitonin: A promising diagnostic marker for sepsis and antibiotic therapy. J. Intensiv. Care 2017, 5, 51. [Google Scholar] [CrossRef]
- Badawy, G.M.; Atallah, M.N.; Sakr, S.A. Effect of gabapentin on fetal rat brain and its amelioration by ginger. Heliyon 2019, 5, e02387. [Google Scholar] [CrossRef]
- Kaid, F.; AlAbsi, A.M.; Al-Afifi, N.; Ali-Saeed, R.; Al-Koshab, M.A.; Ramanathan, A.; Ali, A.M. Histological, Biochemical, and Hematological Effects of Goniothalamin on Selective Internal Organs of Male Sprague-Dawley Rats. J. Toxicol. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No. | Parameters | Control | LPS (10 mg/kg) | ZIN (150 mg/kg) + LPS | ZIN Only (150 mg/kg) |
|---|---|---|---|---|---|
| 1 | Creatinine Kinase (U/L) | 68.32 ± 3.92 | 855.47 ± 18.84 * | 553.83 ± 19.61 *# | 66.67 ± 3.22 |
| 2 | Serum Creatinine (mg/dL) | 0.30 ± 0.03 | 0.59 ± 0.09 * | 0.38 ± 0.05 *# | 0.27 ± 0.01 |
| 3 | BUN (mg/dL) | 42.93 ± 1.92 | 153.18 ± 7.34 * | 67.71 ± 2.27 *# | 41.05 ± 0.59 |
| 4 | LDH (U/L) | 307.61 ± 12.27 | 972.75 ± 20.74 * | 656.59 ± 7.46 *# | 279.28 ± 7.50 |
| 5 | ALT (U/L) | 27.69 ± 3.20 | 145.02 ± 3.84 | 74.99 ± 6.69 *# | 30.23 ± 1.43 |
| 6 | ALP (U/L) | 64.24 ± 4.32 | 186.54 ± 8.76 * | 91.29 ± 5.34 *# | 61.91 ± 2.65 |
| 7 | AST (U/L) | 57.65 ± 3.76 | 206.49 ± 11.45 * | 87.35 ± 8.29 *# | 52.24 ± 3.56 |
| 8 | BIL (µmol/L) | 1.69 ± 0.52 | 8.55 ± 1.07 * | 4.56 ± 0.76*# | 1.56 ± 0.75 |
| 9 | GGT (U/L) | 1.34 ± 0.24 | 3.89 ± 0.65 * | 2.16 ± 0.66 *# | 1.22 ± 0.34 |
| 10 | Albumin (g/dL) | 3.33 ± 0.12 | 2.36 ± 0.12 * | 3.18 ± 0.013 *# | 3.37 ± 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wali, A.F.; Rehman, M.U.; Raish, M.; Kazi, M.; Rao, P.G.M.; Alnemer, O.; Ahmad, P.; Ahmad, A. Zingerone [4-(3-Methoxy-4-hydroxyphenyl)-butan-2] Attenuates Lipopolysaccharide-Induced Inflammation and Protects Rats from Sepsis Associated Multi Organ Damage. Molecules 2020, 25, 5127. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25215127
Wali AF, Rehman MU, Raish M, Kazi M, Rao PGM, Alnemer O, Ahmad P, Ahmad A. Zingerone [4-(3-Methoxy-4-hydroxyphenyl)-butan-2] Attenuates Lipopolysaccharide-Induced Inflammation and Protects Rats from Sepsis Associated Multi Organ Damage. Molecules. 2020; 25(21):5127. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25215127
Chicago/Turabian StyleWali, Adil Farooq, Muneeb U Rehman, Mohammad Raish, Mohsin Kazi, Padma G. M. Rao, Osamah Alnemer, Parvaiz Ahmad, and Ajaz Ahmad. 2020. "Zingerone [4-(3-Methoxy-4-hydroxyphenyl)-butan-2] Attenuates Lipopolysaccharide-Induced Inflammation and Protects Rats from Sepsis Associated Multi Organ Damage" Molecules 25, no. 21: 5127. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25215127