
Effect of Interaction between 17β-Estradiol, 2-Methoxyestradiol and 16α-Hydroxyestrone with Chromium (VI) on Ovary Cancer Line SKOV-3: Preliminary Study



Abstract
:1. Introduction
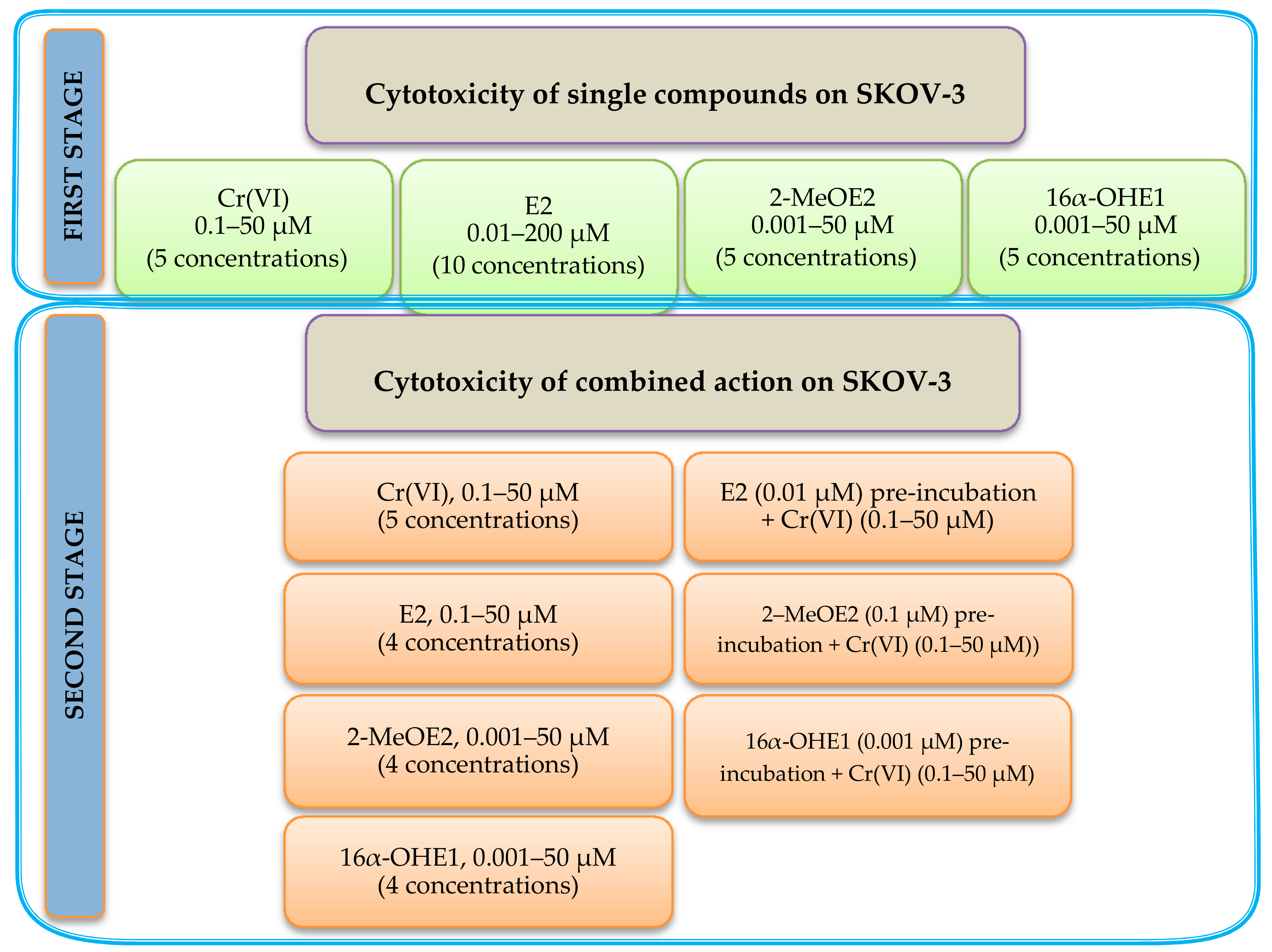
2. Results
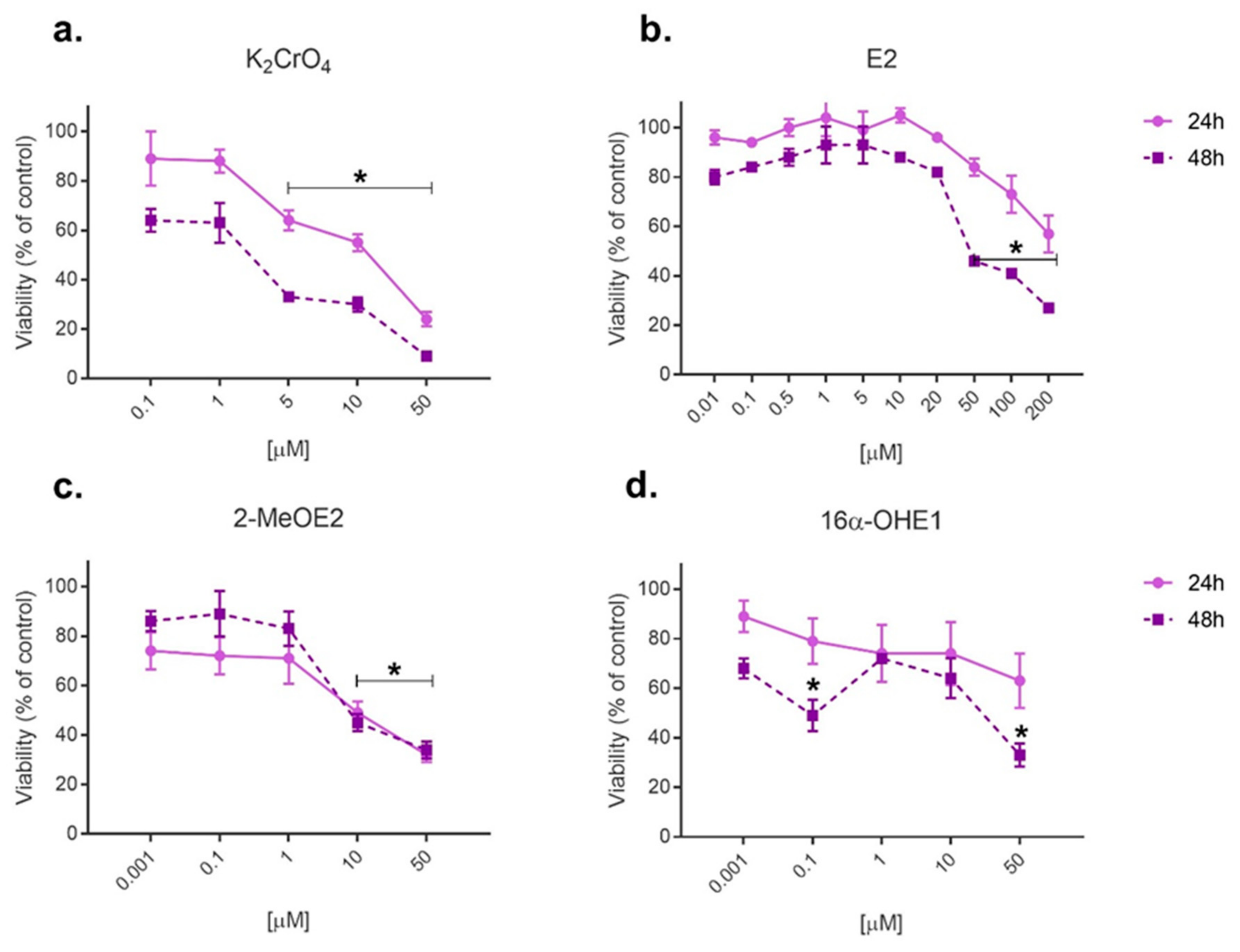
2.1. Cytotoxicity Evaluation by MTT Assay
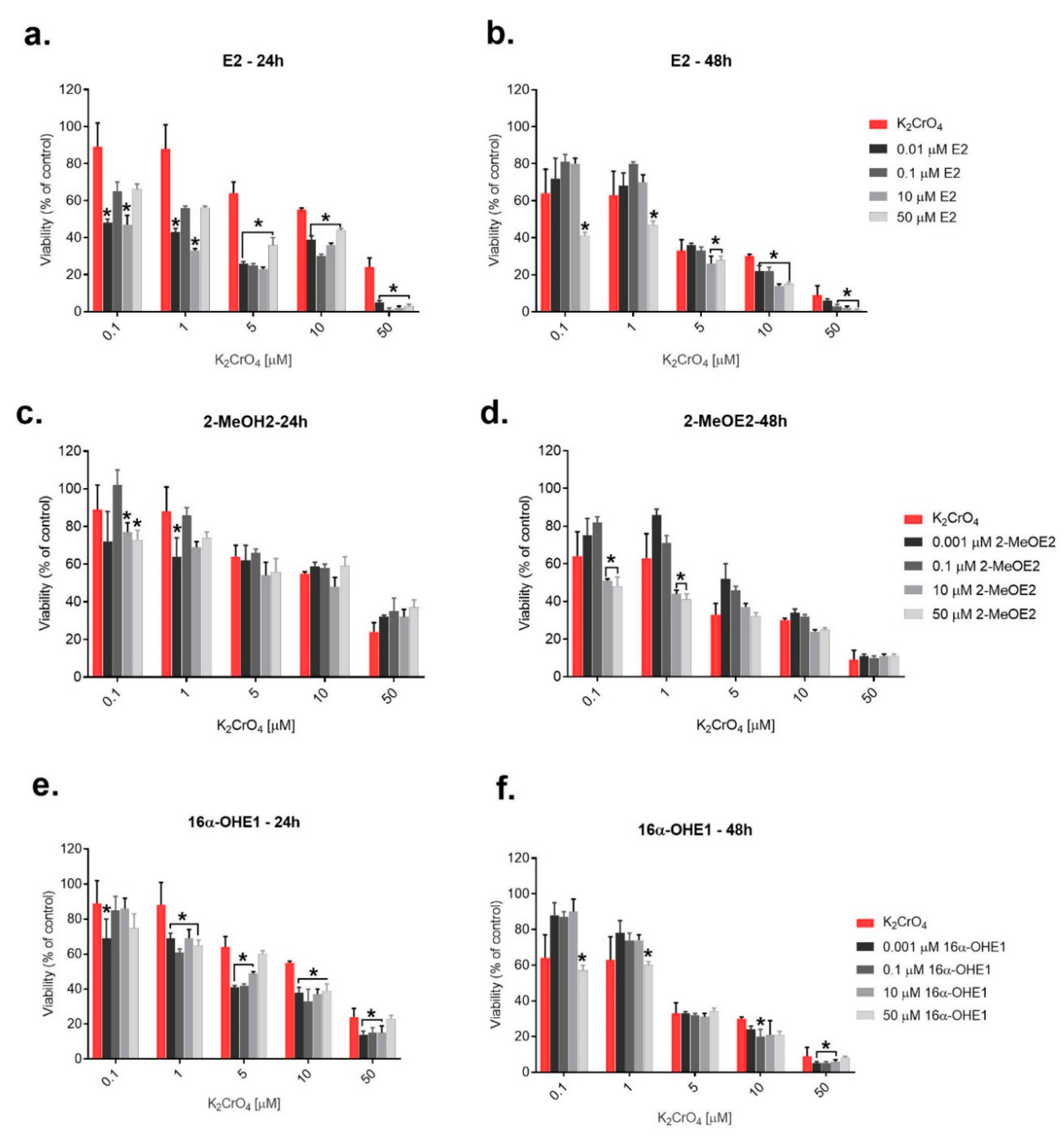
2.2. The Evaluation of Simultaneous Effect of 17β-Estradiol and Its Metabolites as Well as K2CrO4—Interaction Examination
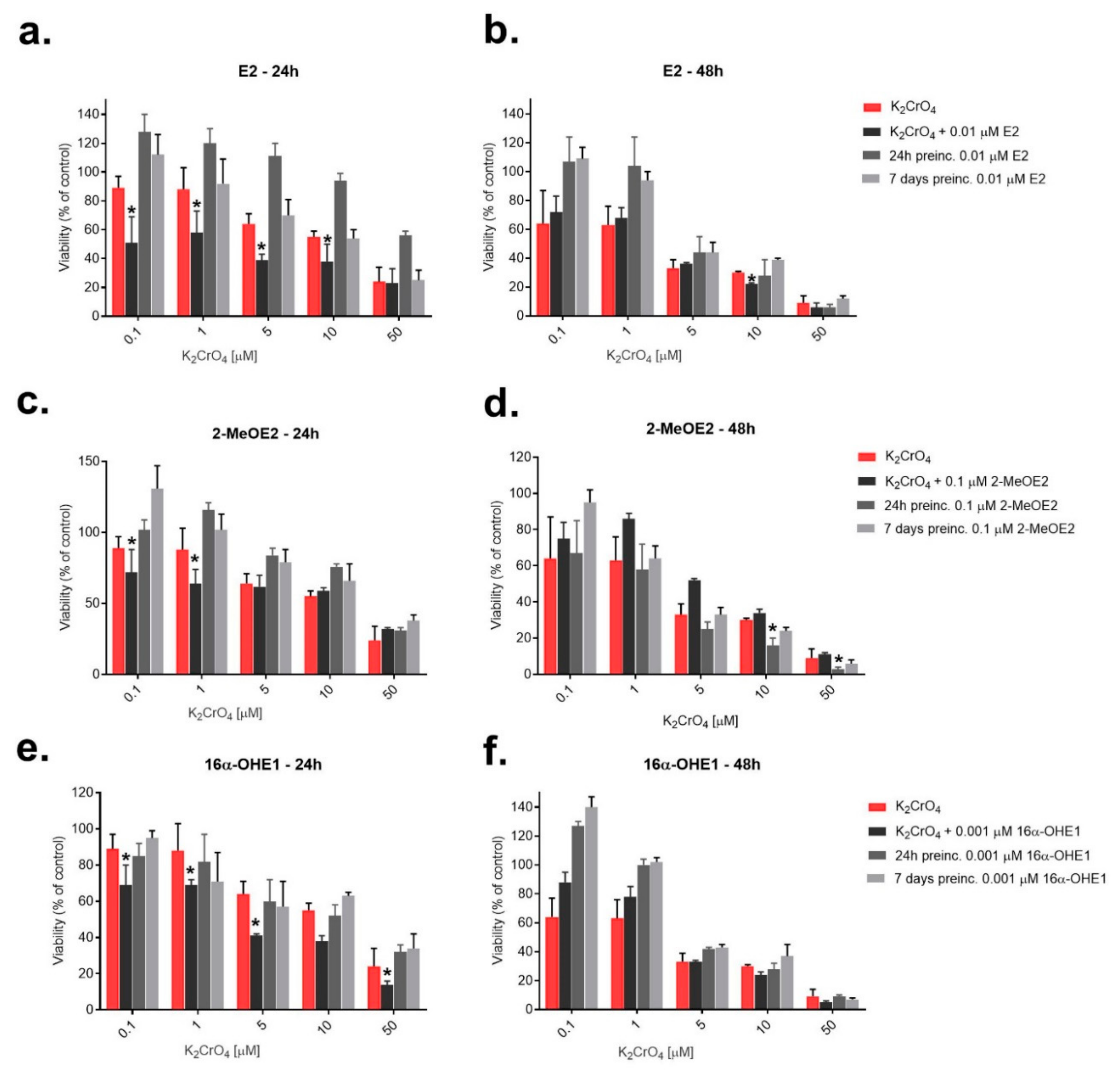
2.3. The Evaluation of Effect of Potassium Chromate (VI) on SKOV-3 Cells after Pre-Incubation (24 h and 7 Days) with 17β-Estradiol and Its Metabolites
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Compounds
4.3. Cytotoxicity Assay of Single Action of Estrogens and Cr(VI)
4.4. Combination Effect of Estrogen–Chromium (VI) Estimation
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Reid, M.B.; Permuth, J.B.; Sellers, T.A. Epidemiology of ovarian cancer: A review. Cancer Biol. Med. 2017, 4, 9–32. [Google Scholar]
- Momenimovahed, Z.; Tiznobaik, A.; Taheri, S.; Salehiniya, H. Ovarian cancer in the world: Epidemiology and risk factors. Int. J. Womens Health 2019, 11, 287–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukanova, A.; Kaaks, R. Endogenous hormones and ovarian cancer: Epidemiology and current hypotheses. Cancer Epidemiol. Biomarkers Prev. 2005, 14, 98–107. [Google Scholar] [PubMed]
- Dallal, C.M.; Lacey, J.V., Jr.; Pfeiffer, R.M.; Bauer, D.C.; Falk, R.T.; Buist, D.S.M.; Cauley, J.A.; Hue, T.F.; LaCroix, A.Z.; Tice, J.A.; et al. Estrogen Metabolism and Risk of Postmenopausal Endometrial and Ovarian Cancer: The B∼FIT Cohort. Horm. Cancer 2016, 7, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Falk, R.T.; Manson, J.E.; Barnabei, V.M.; Anderson, G.L.; Brinton, L.A.; Rohan, T.E.; Cauley, J.A.; Chen, C.; Coburn, S.B.; Pfeiffer, R.M.; et al. Estrogen metabolism in menopausal hormone users in the Women’s Health Initiative Observational Study: Does it differ between estrogen plus progestin and estrogen alone. Int. J. Cancer 2019, 144, 730–740. [Google Scholar] [CrossRef] [PubMed]
- Modugno, F.; Laskey, R.; Smith, A.L.; Andersen, C.L.; Haluska, P.; Oesterreich, S. Hormone response in ovarian cancer: Time to reconsider as a clinica ltarget? Endocr. Relat. Cancer 2012, 19, R255–R279. [Google Scholar] [CrossRef] [Green Version]
- Schuler, S.; Ponnath, M.; Engel, J.; Ortmann, O. Ovarian epithelial tumors and reproductive factors: Asystematic review. Arch. Gynecol. Obstet. 2013, 287, 1187–1204. [Google Scholar] [CrossRef]
- Zhou, B.; Sun, Q.; Cong, R.; Gu, H.; Tang, N.; Yang, L.; Wang, B. Hormone replacement therapy and ovarian cancer risk: A meta-analysis. Gynecol. Oncol. 2008, 108, 641–651. [Google Scholar] [CrossRef]
- Mungenast, F.; Thalhammer, T. Estrogen biosynthesis and action in ovarian cancer. Front. Endocrinol. 2014, 5, 192. [Google Scholar] [CrossRef] [Green Version]
- Park, S.H.; Cheung, L.W.; Wong, A.S.; Leung, P.C. Estrogen regulates snail and slug in the down-regulation of E-cadherin and induces metastatic potential of ovarian cancer cells through estrogen receptor alpha. Mol. Endocrinol. 2008, 22, 2085–2098. [Google Scholar] [CrossRef]
- Wellejus, A.; Olsen, A.; Tjonneland, A.; Thomsen, B.L.; Overvad, K.; Loft, S. Urinary hydroxyestrogens and breast cancer risk among postmenopausal women: A prospective study. Cancer Epidemiol. Biomarkers Prev. 2005, 14, 2137–2142. [Google Scholar] [CrossRef] [Green Version]
- Ursin, G.; London, S.; Stanczyk, F.Z.; Gentzschein, E.; Paganini-Hill, A.; Ross, R.K.; Pike, M.C. A pilot study of urinary estrogen metabolites (16alpha-OHE1 and 2-OHE1) in postmenopausal women with and without breast cancer. Environ. Health Perspect. 1997, 105, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Parada-Bustamante, A.; Valencia, C.; Reuquen, P.; Diaz, P.; Rincion-Rodriguez, R.; Orihuela, P.A. Role of 2-methoxyestradiol, an Endogenous Estrogen Metabolite, in Health and Disease. Mini Rev. Med. Chem. 2015, 15, 427–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saczko, J.; Choromańska, A.; Rembiałkowska, N.; Dubińska-Magiera, M.; Bednarz-Misa, I.; Bar, J.; Marcinkowska, A.; Kulbacka, J. Oxidative modification induced by photodynamic therapy with Photofrin®II and 2-methoxyestradiol in human ovarian clear carcinoma (OvBH-1) and human breast adenocarcinoma (MCF-7) cells. Biomed. Pharmacother. 2015, 71, 30–36. [Google Scholar] [CrossRef]
- Pribluda, V.S.; Gubish, E.R., Jr.; Lavallee, T.M.; Treston, A.; Swartz, G.M.; Green, S.J. 2-Methoxyestradiol: An endogenous antiangiogenic and antiproliferative drug candidate. Cancer Metastasis Rev. 2000, 19, 173–179. [Google Scholar] [CrossRef]
- Mueck, A.O.; Seeger, H.; Wallwiener, D.; Huober, J. Is the combination with 2- methoxyestradiol able to reduce the dosages of chemotherapeutics in the treatment of human ovarian cancer? Preliminary in vitro investigations. Eur. J. Gynaecol. Oncol. 2004, 25, 699–701. [Google Scholar] [PubMed]
- Dikshit, A.; Hales, K.; Hales, D.B. Whole flaxseed diet alters estrogen metabolism to promote 2-methoxtestradiol-induced apoptosis in hen ovarian cancer. J. Nutr. Biochem. 2017, 42, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.U.; Rhee, M.; Min, Y.K.; Kim, S.H. Involvement of peroxiredoxin IV in the 16ahydroxyestrone-induced proliferation of human MCF-7 breast cancer cells. Cell Biol. Int. 2008, 32, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kim, K.Y.; An, B.S.; Choi, J.H.; Jeung, E.B.; Leung, P.C.; Choi, K.C. Cell growth of ovarian cancer cells is stimulated by xenoestrogens through an estrogen-dependent pathway, but their stimulation of cell growth appears not to be involved in the activation of the mitogen-activated protein kinases ERK-1 and p38. J. Reprod. Dev. 2009, 55, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Martin, B.M.; Reiter, R.; Pham, T.; Avellanet, J.C.; Lahm, M.; Pentecost, E.; Pratap, K.; Gilmore, B.A.; Divekar, S.; Dagata, R.S.; et al. Estrogen-like activity of metals in MCF-7 breast cancer cells. Endocrinology 2003, 144, 2425–2436. [Google Scholar] [CrossRef] [Green Version]
- Byrne, C.; Divekar, S.D.; Storchan, G.B.; Parody, D.; Martin, M.B. Metals and breast cancer. J. Mammary Gland Biol. Neoplasia 2013, 18, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Jomova, K.; Valko, M. Advances in metal-induced oxidative stress and human disease. Toxicology 2011, 283, 65–87. [Google Scholar] [CrossRef]
- Fu, S.C.; Liu, J.M.; Lee, K.I.; Tang, F.C.; Fang, K.M.; Yang, C.Y.; Su, C.C.; Chen, H.H.; Hsu, R.J.; Chen, Y.W. Cr(VI) induces ROS-mediated mitochondrial-dependent apoptosis in neuronal cells via the activation of Akt/ERK/AMPK signaling pathway. Toxicol. In Vitro 2020, 65, 104795. [Google Scholar] [CrossRef]
- Mishra, A.K.; Mohanty, B. Effect of sublethal hexavalent chromium exposure on the pituitary-ovarian axis of a teleost, channapunctatus (bloch). Environ. Toxicol. 2010, 27, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Rembacz, K.; Sawicka, E.; Długosz, A. Role of estradiol in chromium–induced oxidative stress. Acta Pol. Pharm. 2012, 69, 1372–1379. [Google Scholar]
- Sawicka, E.; Piwowar, A.; Musiała, T.; Długosz, A. The estrogens/chromium interaction in the nitric oxide generation. Acta Pol. Pharm. 2017, 74, 785–791. [Google Scholar]
- Russo, J.; Lareef, M.H.; Balogh, G.; Guo, S.; Russo, I.H. Estrogen and its metabolites are carcinogenic agents in human breast epithelial cells. J. Steroid. Biochem. Mol. Biol. 2003, 87, 1–25. [Google Scholar] [CrossRef]
- Kajiyama, H.; Suzuki, S.; Utsumi, F.; Nishino, K.; Niimi, K.; Mizuno, M.; Yoshikawa, N.; Kawai, M.; Oguchi, H.; Mizuno, K.; et al. Epidemiological overview of metastatic ovarian carcinoma: Long-term experience of TOTSG database. Nagoya J. Med. Sci. 2019, 81, 193–198. [Google Scholar]
- Vuong, N.H.; Salah, O.S.; Vanderhyden, B.C. 17β-Estradiol sensitizes ovarian surface epithelium to transformation by suppressing Disabled-2 expression. Sci. Rep. 2017, 716702. [Google Scholar] [CrossRef] [Green Version]
- Sassarini, J.; Lumsden, M.A. Estrogen replacement in postmenopausal women. Age Ageing. 2015, 44, 551–558. [Google Scholar] [CrossRef] [Green Version]
- Marino, M.; Pellegrini, M.; La Rosa, P.; Acconcia, F. Susceptibility of estrogen receptor rapid responses to xenoestrogens: Physiological outcomes. Steroids 2012, 77, 910–917. [Google Scholar] [CrossRef]
- Deng, Y.; Wang, M.; Tian, T.; Lin, S.; Xu, P.; Zhou, L.; Dai, C.; Hao, Q.; Wu, Y.; Zhai, Z.; et al. The Effect of Hexavalent Chromium on the Incidence and Mortality of Human Cancers: A Meta-Analysis Based on Published Epidemiological Cohort Studies. Front. Oncol. 2019, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.L.; Siu, M.K.Y.; Jiang, Y.; Wang, J.; Wang, Y.; Leung, T.Y.; Liu, S.S.; Cheung, A.N.Y.; Ngan, H.Y.S. Differential expression of estrogen receptor subtypes and variants in ovarian cancer: Effects on cell invasion, proliferation and prognosis. BMC Cancer 2017, 17, 606. [Google Scholar] [CrossRef]
- Laing, N.M.; Belinsky, M.G.; Kruh, G.D.; Bell, D.W.; Boyd, J.T.; Barone, L.; Tew, K.D. Amplification of the ATP-Binding Cassette 2 Transporter Gene Is Functionally Linked with Enhanced Efflux of Estramustine in Ovarian Carcinoma Cells. Cancer Res. 1998, 58, 1332–1337. [Google Scholar]
- Liu, X.; Li, Q.; Zhou, J.S.; Zhang, S. ATP binding cassette transporter A7 accelerates epithelial to mesenchymal transition in ovarian cancer cells by upregulating the transforming growth factor β signaling pathway. Oncol. Lett. 2018, 16, 5868–5874. [Google Scholar] [CrossRef]
- Mueck, A.O.; Seeger, H.; Wallwiener, D.; Kraemer, E. Estradiol metabolites are potent mitogenic substances for human ovarian cancer cells. Eur. J. Gynaecol. Oncol. 2005, 26, 383–385. [Google Scholar]
- Lindgren, P.R.; Cajander, S.; Bäckström, T.; Gustafsson, J.A.; Mäkelä, S.; Olofsson, J.I. Estrogen and progesterone receptors in ovarian epithelial tumors. Mol. Cell. Endocrinol. 2004, 221, 97–104. [Google Scholar] [CrossRef] [PubMed]
- De Stefano, I.; Zannoni, G.F.; Prisco, M.G.; Fagotti, A.; Tortorella, L.; Vizzielli, G.; Mencaglia, L.; Scambia, G.; Gallo, D. Cytoplasmic expression of estrogen receptor beta (ERβ) predicts poor clinical outcome in advanced serous ovarian cancer. Gynecol. Oncol. 2011, 122, 573–579. [Google Scholar] [CrossRef]
- Kato, S.; Sadarangani, A.; Lange, S.; Delpiano, A.M.; Vargas, M.; Brañes, J.; Carvajal, J.; Lipkowitz, S.; Owen, G.I.; Cuello, M.A. 2-methoxyestradiol mediates apoptosis through caspase-dependent and independent mechanisms in ovarian cancer cells but not in normal counterparts. Reprod. Sci. 2008, 15, 878–894. [Google Scholar] [CrossRef]
- Ding, L.; Wang, X.O.; Zhang, J.; Mu, Z.L.; Zhou, X.X.; Liu, P.S. Underlying mechanism of 2-methoxyestradiolinduced apoptosis and growth arrest in SKOV-3 human ovarian cancer cells. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 2084–2090. [Google Scholar]
- Seeger, H.; Wallwiener, D.; Kraemer, E.; Mueck, A.O. Comparison of possible carcinogenic estradiol metabolites: Effects on proliferation, apoptosis and metastasis of human breast cancer cells. Maturitas 2006, 54, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Mueck, A.O.; Seeger, H.; Lippert, T.H. Estradiol metabolism and malignant disease. Maturitas 2002, 43, 1–10. [Google Scholar] [CrossRef]
- Zhao, L.; Au, J.L.; Wientjes, M.G. Comparison of methods for evaluating drug-drug interaction. Front. Biosci. 2010, 2, 241–249. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Incubation (h) | 17β-Estradiol (µM) | |||||||
|---|---|---|---|---|---|---|---|---|
| Chromium (VI) (µM) | 24 h | 48 h | ||||||
| 50 | 10 | 0.1 | 0.01 | 50 | 10 | 0.1 | 0.01 | |
| 50 | S | S | S | S | S | S | S | S |
| 10 | S | S | S | S | S | S | S | S |
| 5 | S | S | S | S | S | S | A | A |
| 1 | S | S | S | S | S | A | A | A |
| 0.1 | S | S | S | S | S | A | A | S |
| Incubation (h) | 2-Methoxyestradiol (µM) | |||||||
|---|---|---|---|---|---|---|---|---|
| Chromium (VI) (µM) | 24 h | 48 h | ||||||
| 50 | 10 | 0.1 | 0.001 | 50 | 10 | 0.1 | 0.001 | |
| 50 | A | A | A | A | S | S | S | S |
| 10 | A | A | A | A | A | S | A | A |
| 5 | A | A | A | A | A | A | A | A |
| 1 | A | A | A | S | A | A | A | A |
| 0.1 | A | A | A | S | A | A | A | A |
| Incubation (h) | 16α-Hydroxyestrone (µM) | |||||||
|---|---|---|---|---|---|---|---|---|
| Chromium (VI) (µM) | 24 h | 48 h | ||||||
| 50 | 10 | 0.1 | 0.001 | 50 | 10 | 0.1 | 0.001 | |
| 50 | S | S | S | S | S | S | S | S |
| 10 | S | S | S | S | S | S | S | S |
| 5 | A | S | S | S | A | S | A | A |
| 1 | A | A | S | S | A | A | A | A |
| 0.1 | A | A | A | S | A | A | A | A |
| The Name of the Compound | The Structure and the Manufacturer’s Name | Stock Solution | The Molecular Mass of the Compound |
|---|---|---|---|
| 17β-estradiol |  Sigma—Aldrich | 50 mM ethanol solution | 272.38 g/mol |
| 2-MeOE2 |  Sigma—Aldrich | 10 mM ethanol solution | 302.41 g/mol |
| 16α-OHE1 |  Steraloids | 10 mM ethanol solution | 286.40 g/mol |
| Potassium chromate (VI) | K2CrO4 Sigma-Aldrich | 50 mM water solution | 194.19 g/mol |
Sample Availability: Samples of the compounds are not available from the authors. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sawicka, E.; Saczko, J.; Roik, J.; Kulbacka, J.; Piwowar, A. Effect of Interaction between 17β-Estradiol, 2-Methoxyestradiol and 16α-Hydroxyestrone with Chromium (VI) on Ovary Cancer Line SKOV-3: Preliminary Study. Molecules 2020, 25, 5214. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25215214
Sawicka E, Saczko J, Roik J, Kulbacka J, Piwowar A. Effect of Interaction between 17β-Estradiol, 2-Methoxyestradiol and 16α-Hydroxyestrone with Chromium (VI) on Ovary Cancer Line SKOV-3: Preliminary Study. Molecules. 2020; 25(21):5214. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25215214
Chicago/Turabian StyleSawicka, Ewa, Jolanta Saczko, Joanna Roik, Julita Kulbacka, and Agnieszka Piwowar. 2020. "Effect of Interaction between 17β-Estradiol, 2-Methoxyestradiol and 16α-Hydroxyestrone with Chromium (VI) on Ovary Cancer Line SKOV-3: Preliminary Study" Molecules 25, no. 21: 5214. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25215214