
Mahonia aquifolium Extracts Promote Doxorubicin Effects against Lung Adenocarcinoma Cells In Vitro

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Cytotoxic Activity In Vitro
2.1.1. Cytotoxic Activity of Extracts and DOX
2.1.2. Cytotoxic Activity of DOX in Combination with Extracts
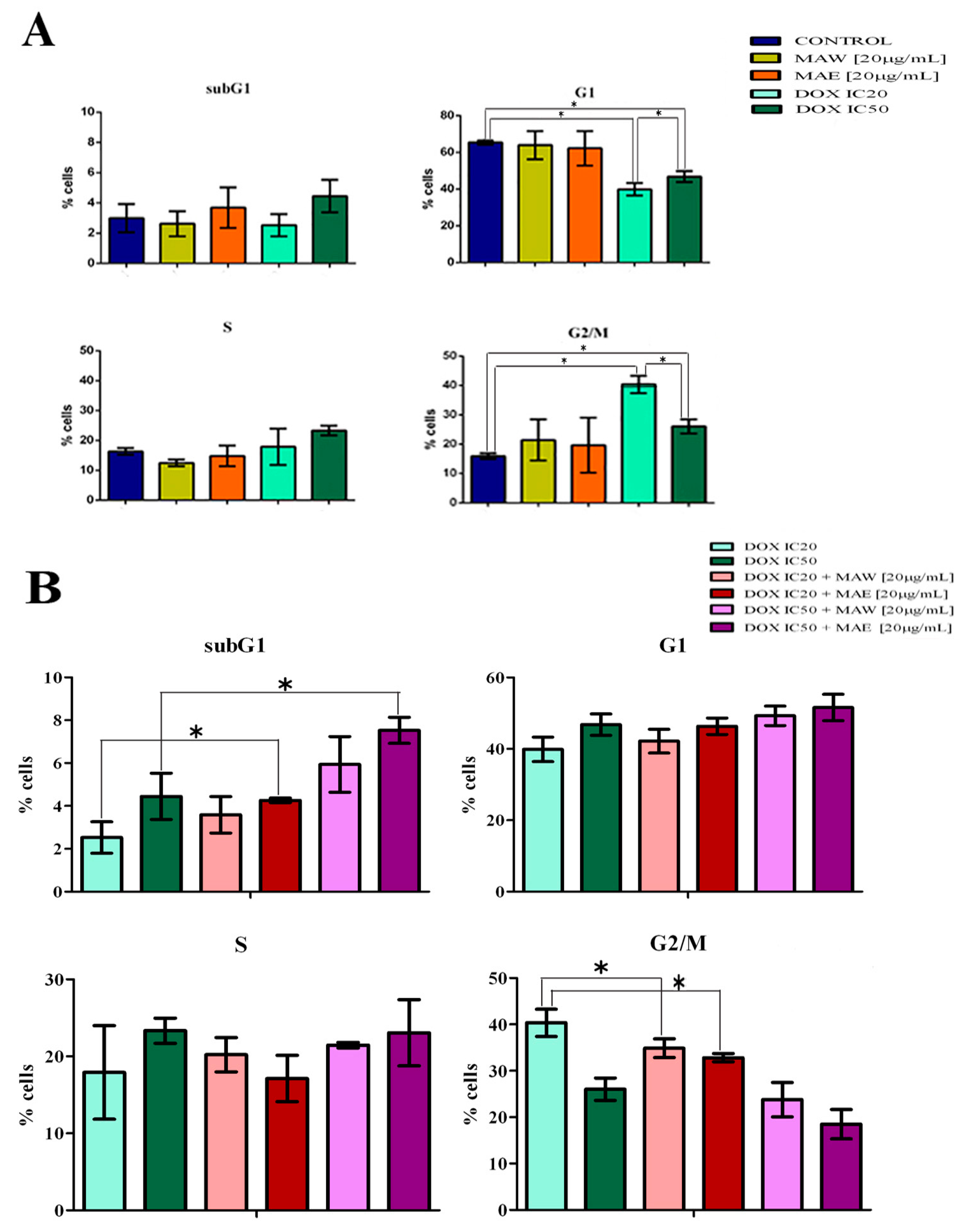
2.2. Cell Cycle Analysis
2.3. Cellular Uptake and DOX Retention
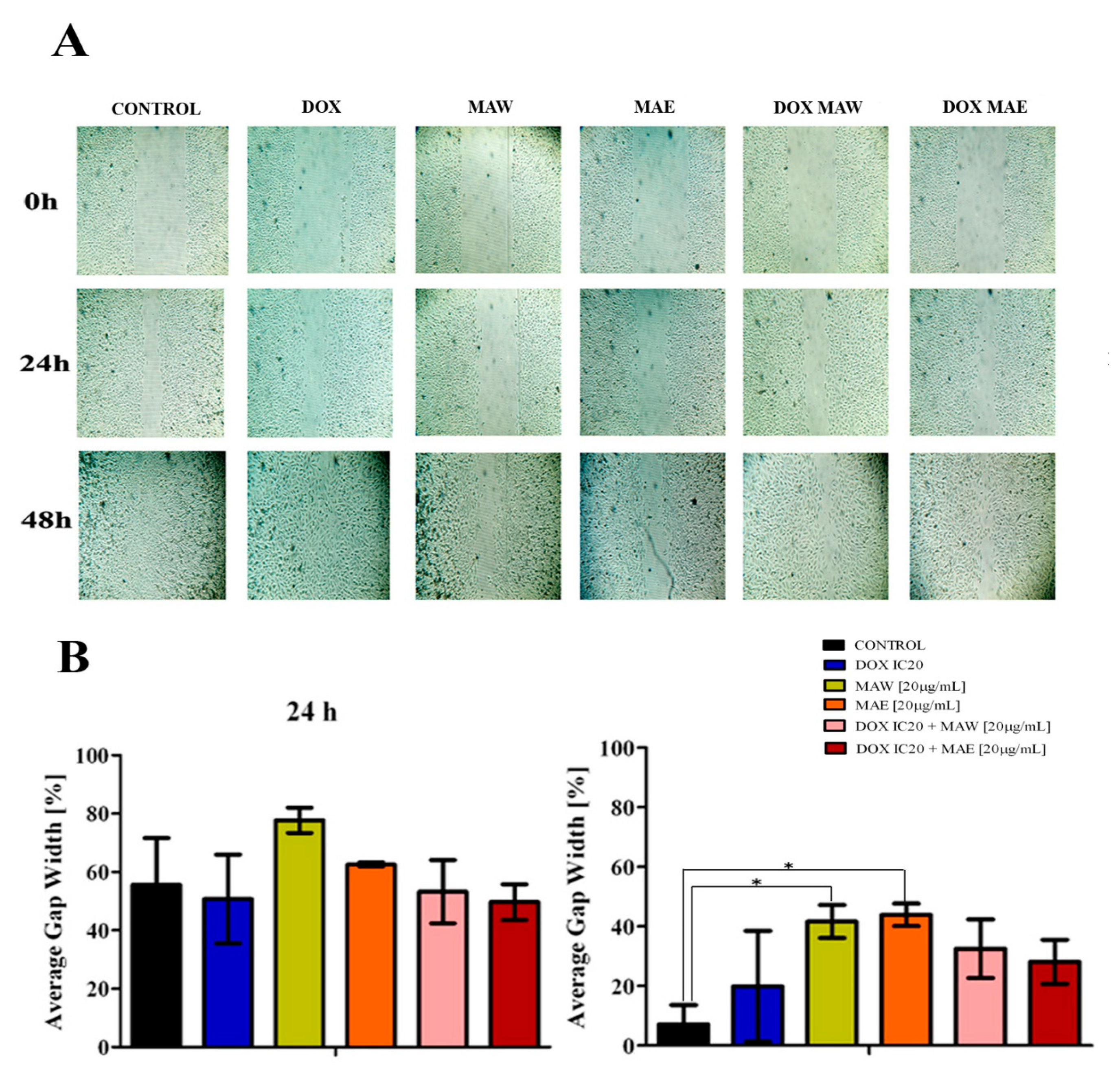
2.4. Cell Migration
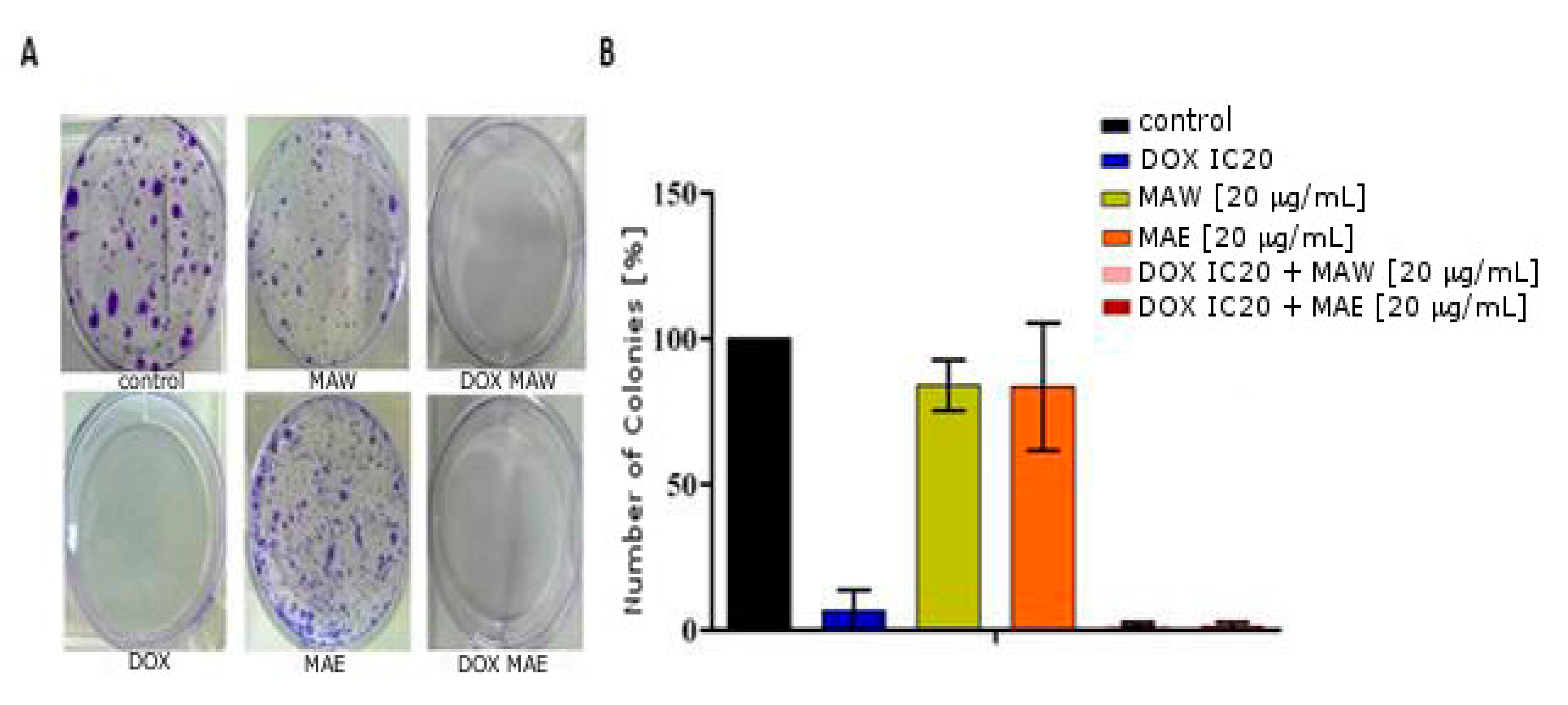
2.5. Colony Formation
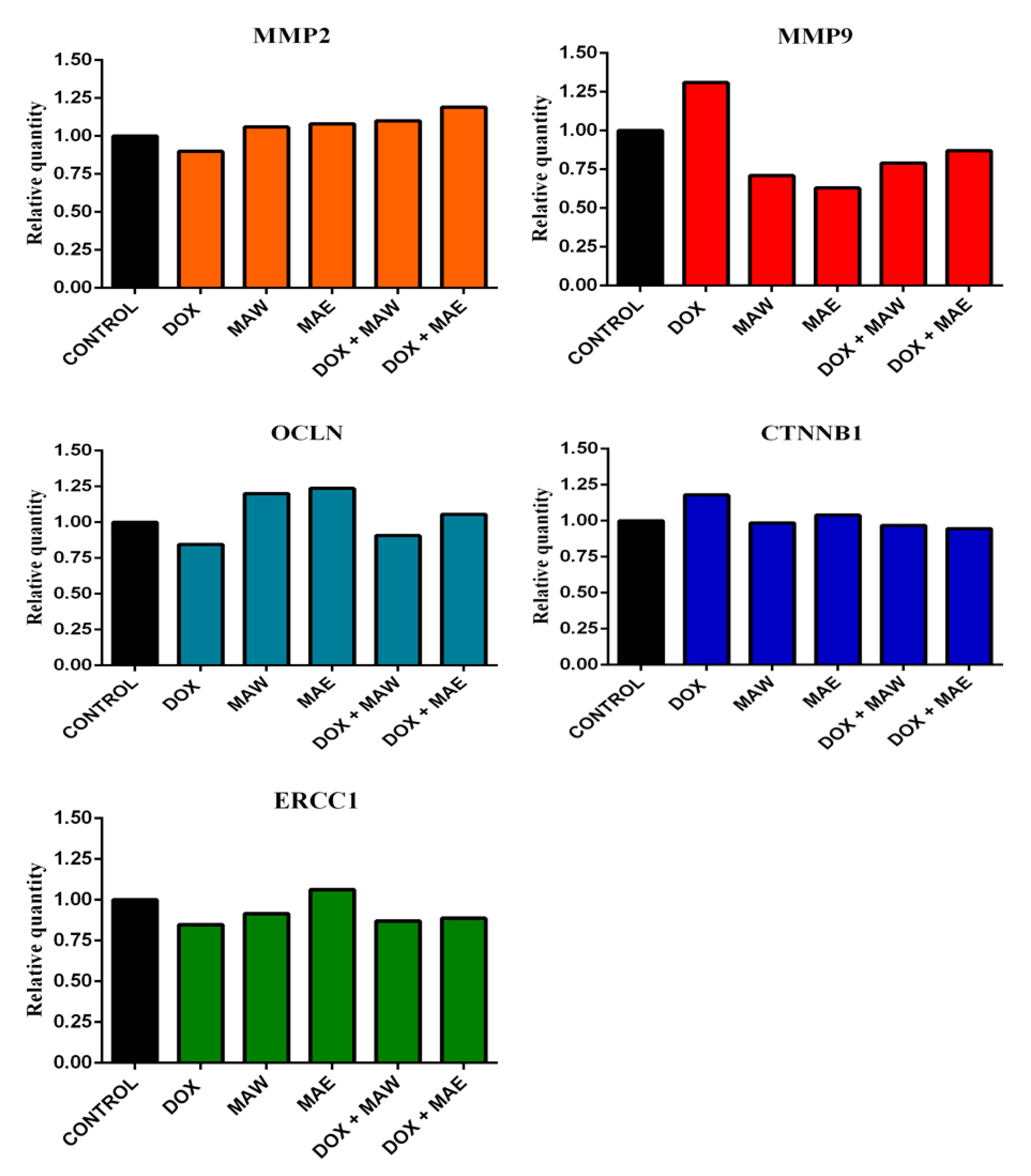
2.6. Gene Expression Analyses
3. Discussion
4. Materials and Methods
4.1. Plant Extracts/LC-MC Analyses
4.2. Reagents
4.3. Cell Lines
4.4. MTT Assay
4.5. Cell Cycle Analysis
4.6. Cellular Uptake and Retention of Doxorubicin
4.7. Scratch Assay
4.8. Colony Formation Assay
4.9. Gene Expression Analyses
4.10. Real-Time PCR Amplification
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Carvalho, C.; Santos, R.X.; Cardoso, S.; Correia, S.; Oliveira, P.J.; Santos, M.S.; Moreira, P.I. Doxorubicin: The Good, the Bad and the Ugly Effect. Curr. Med. Chem. 2009, 16, 3267–3285. [Google Scholar] [CrossRef] [PubMed]
- Thorn, C.F.; Oshiro, C.; Marsh, S.; Hernandez-Boussard, T.; McLeod, H.; Klein, T.E.; Altman, R.B. Doxorubicin pathways: Pharmacodynamics and adverse effects. Pharm. Genom. 2011, 21, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Bamodu, O.A.; Huang, W.C.; Tzeng, D.T.; Wu, A.; Wang, L.S.; Yeh, C.T.; Chao., T.Y. Ovatodiolide sensitizes aggressive breast cancer cells to doxorubicin, eliminates their cancer stem cell-like phenotype, and reduces doxorubicin-associated toxicity. Cancer Lett. 2015, 364, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, J.; Tong, N.; Liao, X..; Wang, E.; Li, Z.; Luo, Y.; Zuo, H. Berberine attenuates doxorubicin-induced cardiotoxicity in mice. J. Int. Med. Res. 2011, 39, 1720–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osman, A.M.; Bayoumi, H.M.; Al-Harthi, S.E.; Damanhouri, Z.A.; Elshal, M.F. Modulation of doxorubicin cytotoxicity by resveratrol in a human breast cancer cell line. Cancer Cell Int. 2012, 12, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.M.; Mu, Q. The medicinal uses of the genus Mahonia in traditional Chinese medicine: An ethnopharmacological, phytochemical and pharmacological review. J. Ethnopharmacol. 2015, 175, 668–683. [Google Scholar] [CrossRef] [PubMed]
- Wong, B.S.; Hsiao, Y.C.; Lin, T.W.; Chen, K.S.; Chen, P.N.; Kuo, W.H.; Hsieh, Y.S. The in vitro and in vivo apoptotic effects of Mahonia oiwakensis on human lung cancer cells. Chem. Biol. Interact. 2009, 180, 165–174. [Google Scholar] [CrossRef]
- Hu, W.; Yu, L.; Wang, M.H. Antioxidant and antiproliferative properties of water extract from Mahonia bealei (Fort.) Carr. leaves. Food Chem. Toxicol. 2011, 49. [Google Scholar] [CrossRef]
- Kumar, A.; Ekavali, K.; Chopra, M.; Mukherjee, R.; Pottabathini, M.; Dhull, D.K. Current knowledge and pharmacological profile of berberine: An update. Eur. J. Pharmacol. 2015, 761, 288–297. [Google Scholar] [CrossRef]
- Zhao, T.F.; Wang, X.K.; Rimando, A.M.; Che, C.T. Folkloric medicinal plants: Tinospora sagittata var. cravaniana and Mahonia bealei. Planta Med. 1991, 57, 505–510. [Google Scholar] [CrossRef]
- Cernáková, M.; Kost’álová, D.; Kettmann, V.; Plodová, M.; Tóth, J.; Drímal, J. Potential antimutagenic activity of berberine, a constituent of Mahonia aquifolium. BMC Complementary Altern. Med. 2002, 2, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Racková, L.; Májeková, M.; Kost’álová, D.; Stefek, M. Antiradical and antioxidant activities of alkaloids isolated from Mahonia aquifolium. Structural aspects. Bioorganic Med Chem. 2004, 12, 4709–4715. [Google Scholar] [CrossRef] [PubMed]
- Kost’álová, D.; Kardosová, A.; Hajnická, V. Effect of Mahonia aquifolium stem bark crude extract and one of its polysaccharide components on production of IL-8. Fitoterapia 2001, 72, 802–806. [Google Scholar] [CrossRef]
- Damjanović, A.; Zdunić, G.; Šavikin, K.; Mandić, B.; Jadranin, M.; Matić, I.Z.; Stanojković, T.P. Evaluation of the anti-cancer potential of Mahonia aquifolium extracts via apoptosis and anti-angiogenesis. Bangladesh J. Pharmacol. 2016, 11, 741–774. [Google Scholar] [CrossRef] [Green Version]
- Gođevac, D.; Damjanović, A.; Stanojković, T.P.; Anđelković, B.; Zdunić, G. Identification of cytotoxic metabolites from Mahonia aquifolium using1H NMR-based metabolomics approach. J. Pharm. Biomed. Anal. 2017, 150, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Ihde, D.C.; Minna, J.D. Non-small cell lung cancer. Part II: Treatment. Curr. Probl. Cancer 1991, 15, 105–154. [Google Scholar] [CrossRef]
- Borst, P.; Evers, R.; Kool, M.; Wijnholds, J. A family of drug transporters: The multidrug resistance-associated proteins. J. Natl. Cancer Inst. 2000, 92, 1295–1302. [Google Scholar] [CrossRef]
- Salomon, J.J.; Ehrhardt, C. Nanoparticles attenuate P-glycoprotein/MDR1function in A549 human alveolar epithelial cells. Eur. J. Pharm. Biopharm. 2011, 77, 392–397. [Google Scholar] [CrossRef]
- Lee, E.; Lim, S.J. The association of increased lung resistance protein expression with acquired etoposide resistance in human H460 lung cancer cell lines. Arch. Pharmacal Res. 2006, 29, 1018–1023. [Google Scholar] [CrossRef]
- Meschini, S.; Marra, M.; Calcabrini, A.; Monti, E.; Gariboldi, M.; Dolfini, E.; Arancia, G. Role of the lung resistance-related protein (LRP) in the drug sensitivity of cultured tumor cells. Toxicol. Vitr. 2002, 16, 389–398. [Google Scholar] [CrossRef]
- Chatterjee, K.; Zhang, J.; Honbo, N.; Karliner, J.S. Doxorubicin cardiomyopathy. Cardiology 2009, 115, 155–162. [Google Scholar] [CrossRef]
- De Angelis, A.; Urbanek, K.; Cappetta, D.; Piegari, E.; Ciuffreda, L.P.; Rivellino, A.; Esposito, G.; Rossi, F.; Berrino, L. Doxorubicin cardiotoxicity and target cells: A broader perspective. Cardio-Oncol. 2016, 2, 1. [Google Scholar] [CrossRef] [Green Version]
- Lipshultz, S.E.; Lipsitz, S.R.; Sallan, S.E.; Dalton, V.M.; Mone, S.M.; Gelber, R.D.; Colan, S.D. Chronic progressive cardiac dysfunction years after doxorubicin therapy for childhood acute lymphoblastic leukemia. J. Clin. Oncol. 2005, 23, 2629–2636. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Long, Y.; Ni, L. Anticancer effect of berberine based on experimental animal models of various cancers: A systematic review and meta-analysis. BMC Cancer 2019, 19, 589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Z.B.; Lu, K.J.; Wu, X.L.; Chen, C.; Huang, X.E.; Yin, H.T. In vitro and in vivo antitumor evaluation of berbamine for lung cancer treatment. Asian Pac. J. Cancer Prev. 2014, 15, 1767–1769. [Google Scholar] [CrossRef] [PubMed]
- Miwa, S.; Yano, S.; Kimura, H.; Yamamoto, M.; Toneri, M.; Matsumoto, Y.; Uehara, F.; Hiroshima, Y.; Murakami, T.; Hayashi, K.; et al. Cell-cycle fate-monitoring distinguishes individual chemosensitive and chemoresistant cancer cells in drug-treated heterogeneous populations demonstrated by real-time FUCCI imaging. Cell Cycle (Georget. Tex.) 2014, 14, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Wlodkowic, D.; Telford, W.; Skommer, J.; Darzynkiewicz, Z. Apoptosis and beyond: Cytometry in studies of programmed cell death. Methods Cell Biol. 2011, 103, 55–98. [Google Scholar]
- Vermes, I.; Haanen, C. Reutelingsperger. Flow cytometry of apoptotic cell death. J. Immunol. Methods 2000, 243, 167–190. [Google Scholar] [CrossRef]
- DiPaola, R.S. To arrest or not to G(2)-M Cell-cycle arrest: Commentary re: A. K. Tyagi et al., Silibinin strongly synergizes human prostate carcinoma DU145 cells to doxorubicin-induced growth inhibition, G(2)-M arrest, and apoptosis. Clin. Cancer Res. 2002, 8, 3311–3314. [Google Scholar]
- Tipton, J.M. Side effects of cancer chemotherapy. In Handbook of Cancer Chemotherapy; Skeel, R.T., Ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2003; pp. 561–580. [Google Scholar]
- Wong, H.L.; Bendayan, R.; Rauth, A.M.; Xue, H.Y.; Babakhanian, K.; Wu, X.Y. A mechanistic study of enhanced doxorubicin uptake and retention in multidrug resistant breast cancer cells using a polymer-lipid hybrid nanoparticle system. J. Pharmacol. Exp. Ther. 2006, 317, 1372–1381. [Google Scholar] [CrossRef] [Green Version]
- Millenbaugh, N.J.; Wientjes, M.G.; Au, J.L. A pharmacodynamic analysis method to determine the relative importance of drug concentration and treatment time on effect. Cancer Chemother. Pharmacol. 2000, 45, 265–272. [Google Scholar] [PubMed]
- Gonçalves, C.; Martins-Neves, S.R.; Paiva-Oliveira, D.; Oliveira, V.E.; Fontes-Ribeiro, C.; Gomes, C.M. Sensitizing osteosarcoma stem cells to doxorubicin-induced apoptosis through retention of doxorubicin and modulation of apoptotic-related proteins. Life Sci. 2015, 130, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Su, Y.; Fingleton, B.; Acuff, H.; Matrisian, L.M.; Zent, R.; Pozzi, A. Increased plasma MMP9 in integrin alpha1-null mice enhances lung metastasis of colon carcinoma cells. Int. J. Cancer 2005, 116, 52–61. [Google Scholar] [CrossRef]
- Hiraoka, N.; Allen, E.; Apel, I.J.; Gyetko, M.R.; Weiss, S.J. Matrix metalloproteinases regulate neovascularization by acting as pericellular Fibrinolysins. Cell 1998, 95, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Cornelius, L.A.; Nehring, L.C.; Harding, E.; Bolanowski, M.; Welgus, H.G.; Kobayashi, D.K.; Shapiro, S.D. Matrix metalloproteinases generate angiostatin: Effects on neovascularization. J. Immunol. 1998, 161, 6845–6852. [Google Scholar]
- Sang, Q.X. Complex role of matrix metalloproteinases in angiogenesis. Cell Res. 1998, 8, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Hamano, Y.M.; Zeisberg, Y.H.; Sugimoto, H.J.C.; Lively, J.C.Y.; Maeshima, Y.C.; Yang, C.; Kalluri, R. Physiological levels of tumstatin, a fragment of collagen IV alpha3 chain, are generated by MMP-9 proteolysis and suppress angiogenesis via alphaV beta3 integrin. Cancer Cell 2003, 3, 589–601. [Google Scholar] [CrossRef] [Green Version]
- Webb, A.H.; Gao, B.T.; Goldsmith, Z.K.; Morales-Tirado, V.M. Inhibition of MMP-2 and MMP-9 decreases cellular migration, and angiogenesis in in vitro models of retinoblastoma. BMC Cancer 2017, 17, 434. [Google Scholar] [CrossRef]
- Park, K.S.; Kim, J.B.; Bae, J.; Park, S.Y.; Jee, H.G.; Lee, K.E.; Youn, Y.K. Berberine inhibited the growth of thyroid cancer cell lines 8505C and TPC1. Yonsei Med. J. 2012, 53, 346–351. [Google Scholar] [CrossRef]
- Albring, K.F.; Weidemüller, J.; Mittag, S.; Weiske, J.F.; Geroni, M.C.; Lombardi, P.; Huber, O. Berberine acts as a natural inhibitor of Wnt/β-catenin signaling--identification of more active 13-arylalkyl derivatives. Biofactors 2013, 39, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Kaboli, J.; Rahmat, P.; Ismail, A.; Ling, K.H. Targets and mechanisms of berberine, a natural drug with potential to treat cancer with special focus on breast cancer. Eur. J. Pharmacol. 2014, 740, 584–595. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; He, C.; Zhou, K.; Wang, J.; Kang, J.X. Coptis extracts enhance the anticancer effect of estrogen receptor antagonists on human breast cancer cells. Biochem. Biophys. Res. Commun. 2009, 378, 174–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matić, I.Z.; Aljančić, I.; Žižak, Ž.; Vajs, V.; Jadranin, M.; Milosavljević, S.; Juranić, Z.D. In vitro antitumor actions of extracts from endemic plant Helichrysum zivojinii. Bmc Complementary Altern. Med. 2013, 13, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Ohno, M.; Abe, T. Rapid colorimetric assay for the quantification of leukemia inhibitory factor (LIF) and interleukin-6 (IL-6). J. Immunol. Methods 1991, 145, 199–203. [Google Scholar] [CrossRef]
- Huang, R.Y.; Pei, L.; Liu, Q. Isobologram Analysis: A Comprehensive Review of Methodology and Current Research. Front. Pharmacol. 2019, 10, 1222. [Google Scholar] [CrossRef]
- Forner, A.; Llovet, J.M.; Bruix, J. Hepatocellular carcinoma. Lancet 2012, 379, 1245–1255. [Google Scholar] [CrossRef]
- Khan, M.A.; Singh, M.; Khan, M.S.; Najmi, A.K.; Ahmad, S. Caspase mediated synergistic effect of Boswellia serrata extract in combination with doxorubicin against human hepatocellularcarcinoma. Biomed. Res. Int. 2014, 2014, 294143. [Google Scholar] [CrossRef] [Green Version]
- Zec, M.; Srdic-Rajic, T.; Krivokuca, A.; Jankovic, R.; Todorovic, T.; Andelkovic, K.; Radulovic, S. Novel selenosemicarbazone metal complexes exert anti-tumor effect via alternative, caspase- independent necroptotic cell death. Med. Chem. 2014, 10, 759–771. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 (µg/mL) | |
|---|---|
| A549 | |
| DOX | 0.4457 ± 0.0154 |
| DOX + 5 µg/mL MAW | 0.0234 ± 0.0022 |
| DOX + 10 µg/mL MAW | 0.0141 ± 0.0046 |
| DOX + 20 µg/mL MAW | 0.0067 ± 0.0018 |
| DOX + 5 µg/mL MAE | 0.0113 ± 0.0006 |
| DOX + 10 µg/mL MAE | 0.0103 ± 0.0016 |
| DOX + 20 µg/mL MAE | 0.0036 ± 0.0012 |
| CI | |
|---|---|
| Treatment | A549 |
| DOX + 5 µg/mL MAW | 0.14 |
| DOX + 10 µg/mL MAW | 0.21 |
| DOX + 20 µg/mL MAW | 0.38 |
| DOX + 5 µg/mL MAE | 0.12 |
| DOX + 10 µg/mL MAE | 0.22 |
| DOX + 20 µg/mL MAE | 0.40 |
| Treatment | Uptake | Retention |
|---|---|---|
| DOX IC50 | 100 | 100 |
| DOX IC50 + MAW (40 μg/mL) | 92.34 ± 6.64 | 112.97 ± 0.25 |
| DOX IC50 + MAE (40 μg/mL) | 96.58 ± 2.77 | 122.23 ± 1.60 |
Sample Availability: Samples of the compounds are available from the authors. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damjanović, A.; Kolundžija, B.; Matić, I.Z.; Krivokuća, A.; Zdunić, G.; Šavikin, K.; Janković, R.; Stanković, J.A.; Stanojković, T.P. Mahonia aquifolium Extracts Promote Doxorubicin Effects against Lung Adenocarcinoma Cells In Vitro. Molecules 2020, 25, 5233. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25225233
Damjanović A, Kolundžija B, Matić IZ, Krivokuća A, Zdunić G, Šavikin K, Janković R, Stanković JA, Stanojković TP. Mahonia aquifolium Extracts Promote Doxorubicin Effects against Lung Adenocarcinoma Cells In Vitro. Molecules. 2020; 25(22):5233. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25225233
Chicago/Turabian StyleDamjanović, Ana, Branka Kolundžija, Ivana Z. Matić, Ana Krivokuća, Gordana Zdunić, Katarina Šavikin, Radmila Janković, Jelena Antić Stanković, and Tatjana P. Stanojković. 2020. "Mahonia aquifolium Extracts Promote Doxorubicin Effects against Lung Adenocarcinoma Cells In Vitro" Molecules 25, no. 22: 5233. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25225233