
Pogostemon cablin Triggered ROS-Induced DNA Damage to Arrest Cell Cycle Progression and Induce Apoptosis on Human Hepatocellular Carcinoma In Vitro and In Vivo


 , ,
, ,
Abstract
:1. Introduction
2. Results
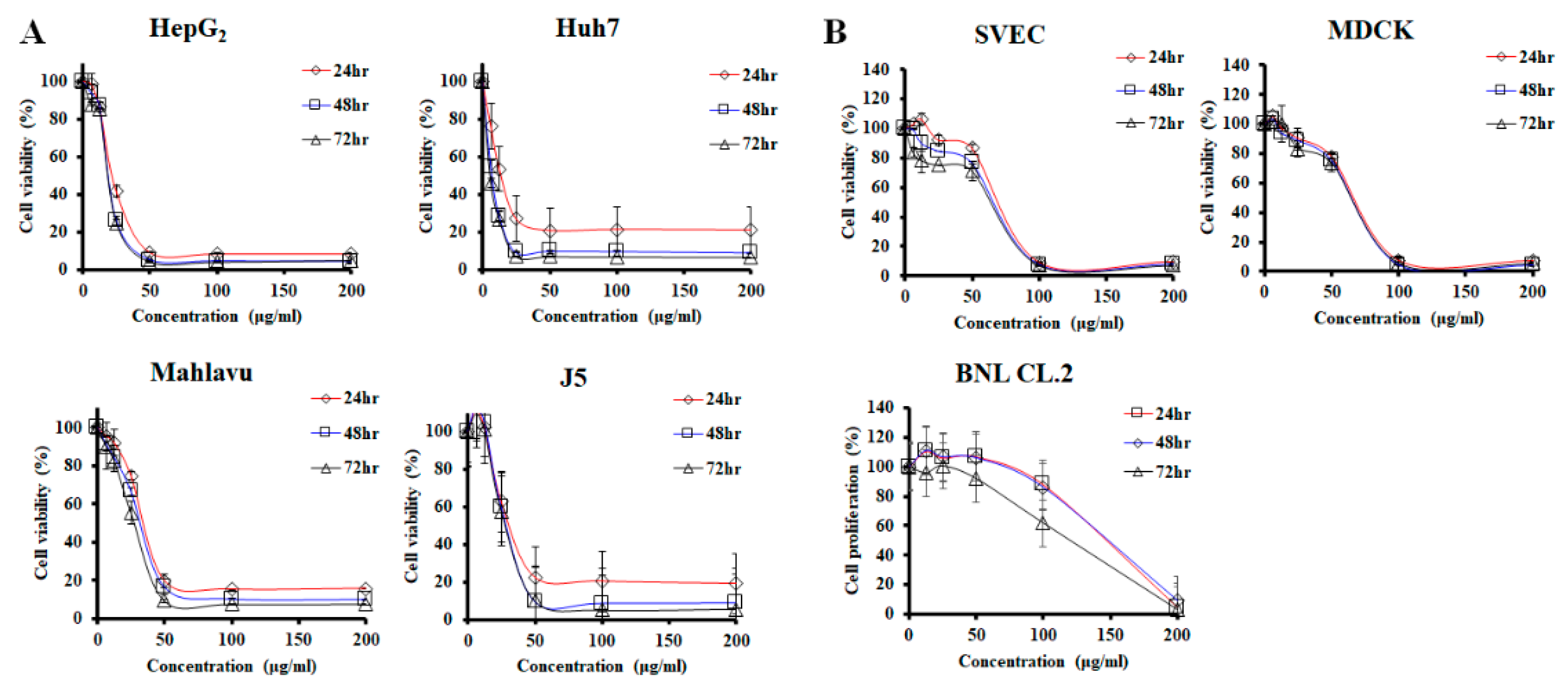
2.1. PPa Extract Inhibited HCC Cell Growth
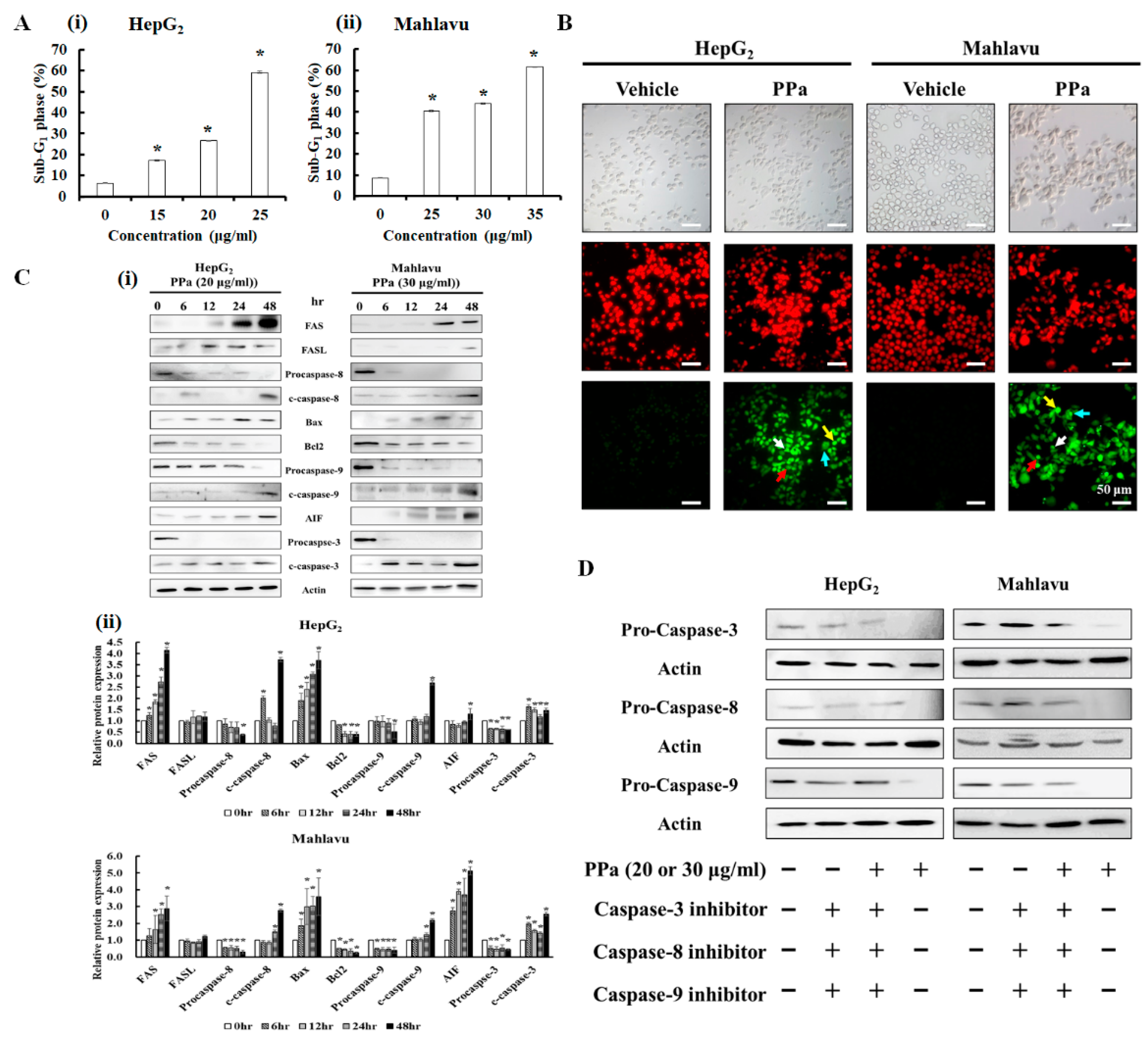
2.2. PPa extract Altered the Cell Cycle Distribution in HCC cells
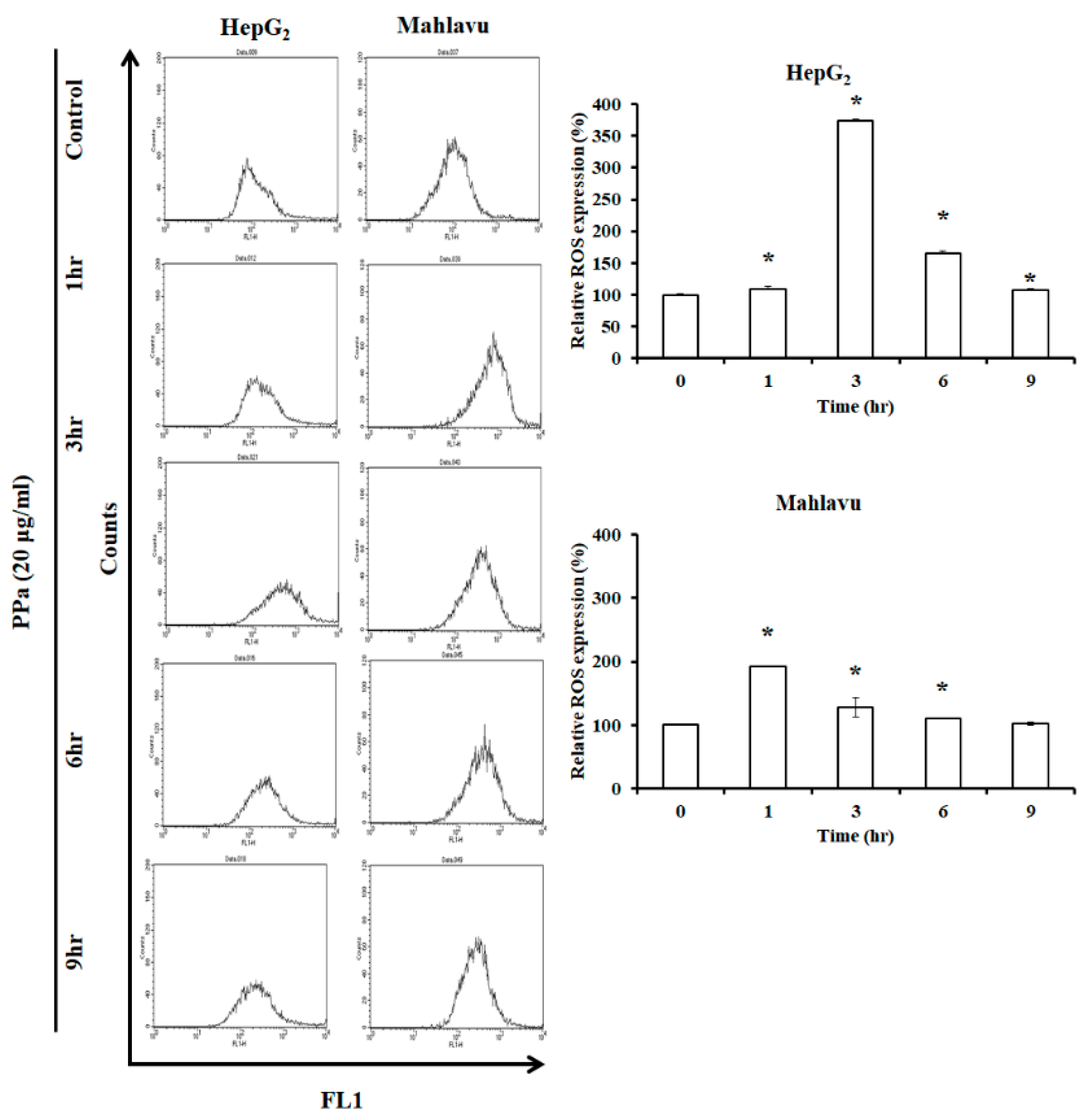
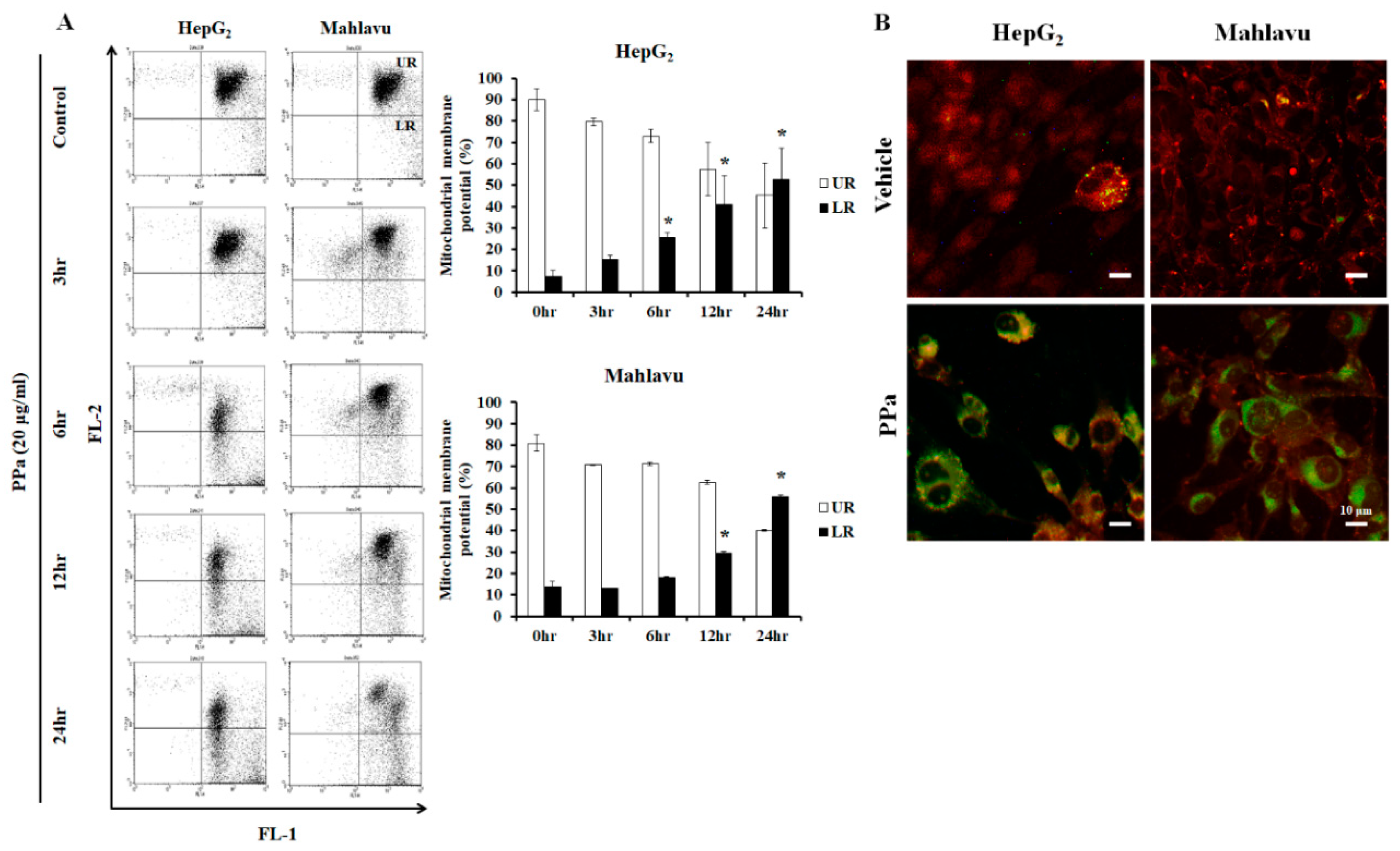
2.3. Ppa Extract Stimulated ROS Production and Imbalanced Mitochondrial Membrane Potential in HCC Cells
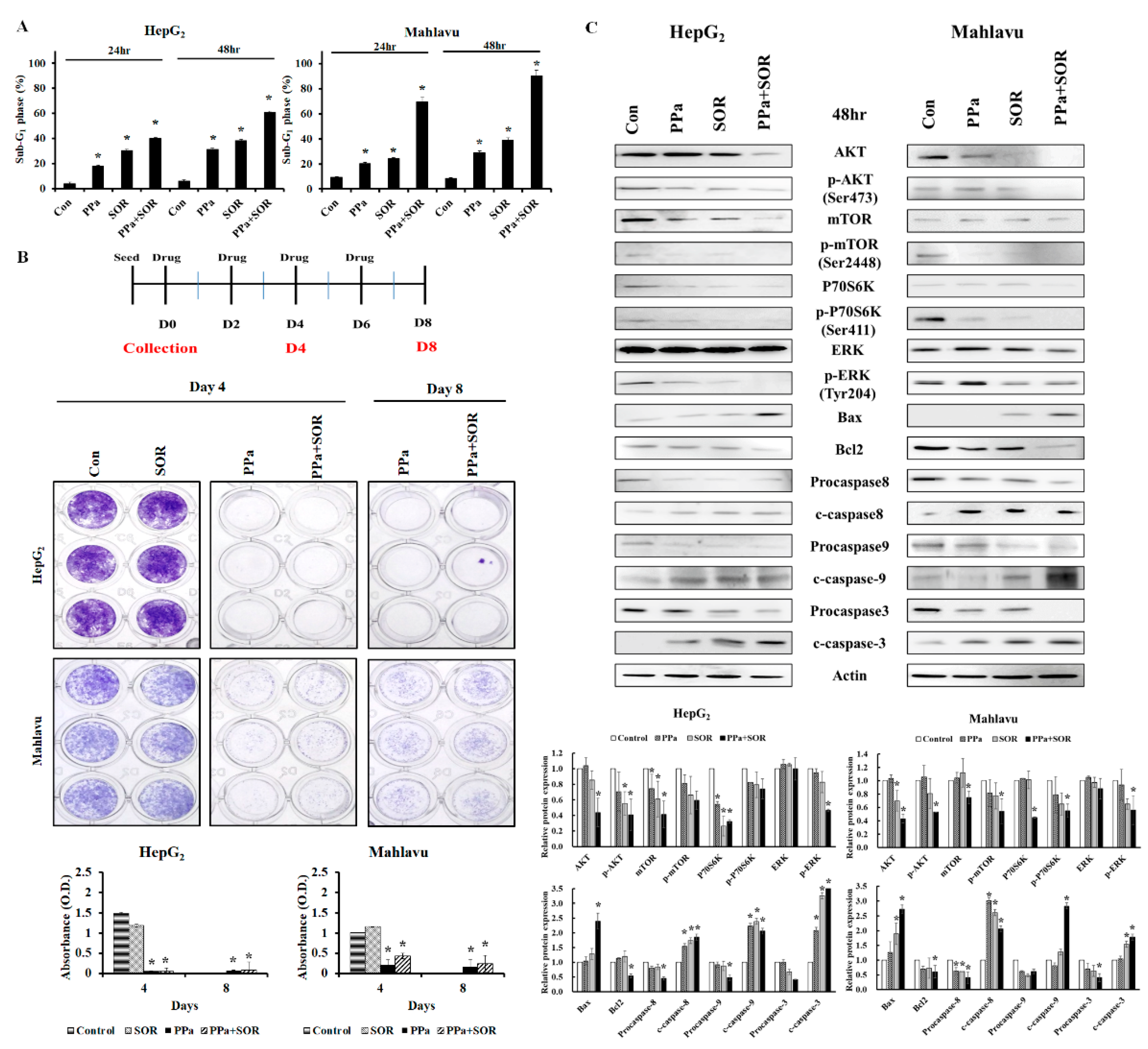
2.4. PPa Extract Induced Extrinsic and Intrinsic Apoptosis in HCC Cells
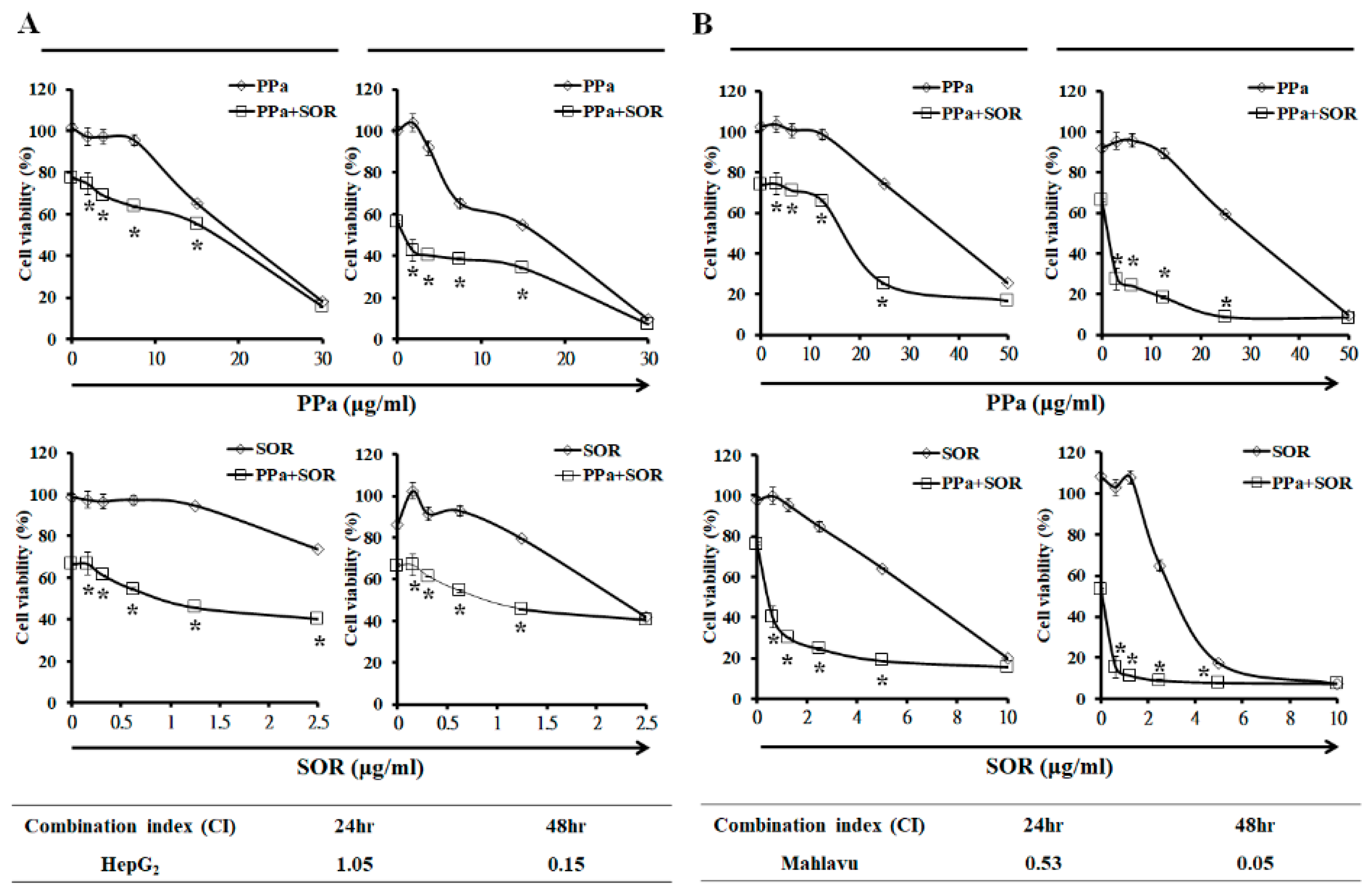
2.5. Synergistic Inhibitory Effects Induced by PPa Extract Plus Sorafenib in Hepatoma Cells
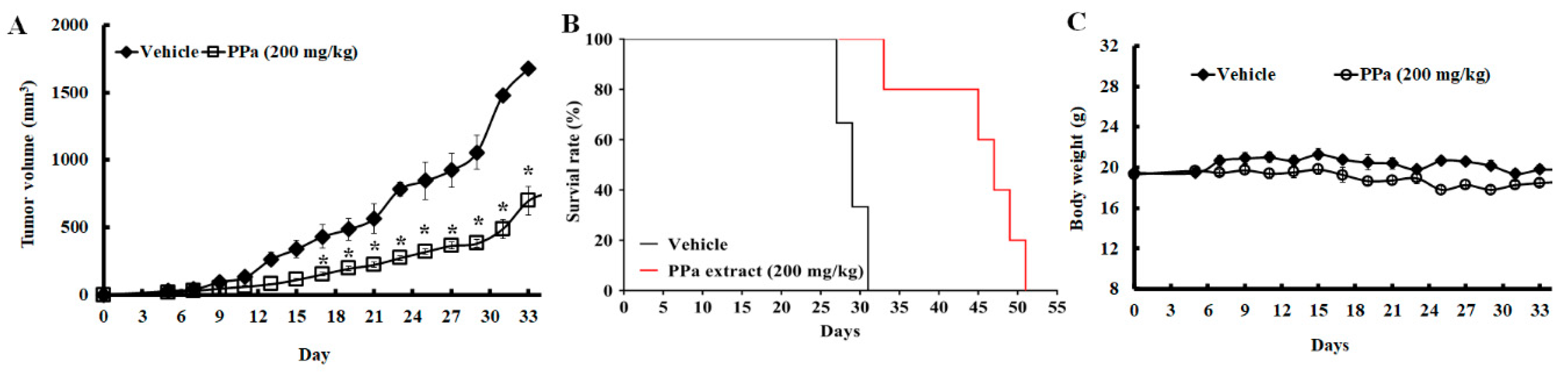
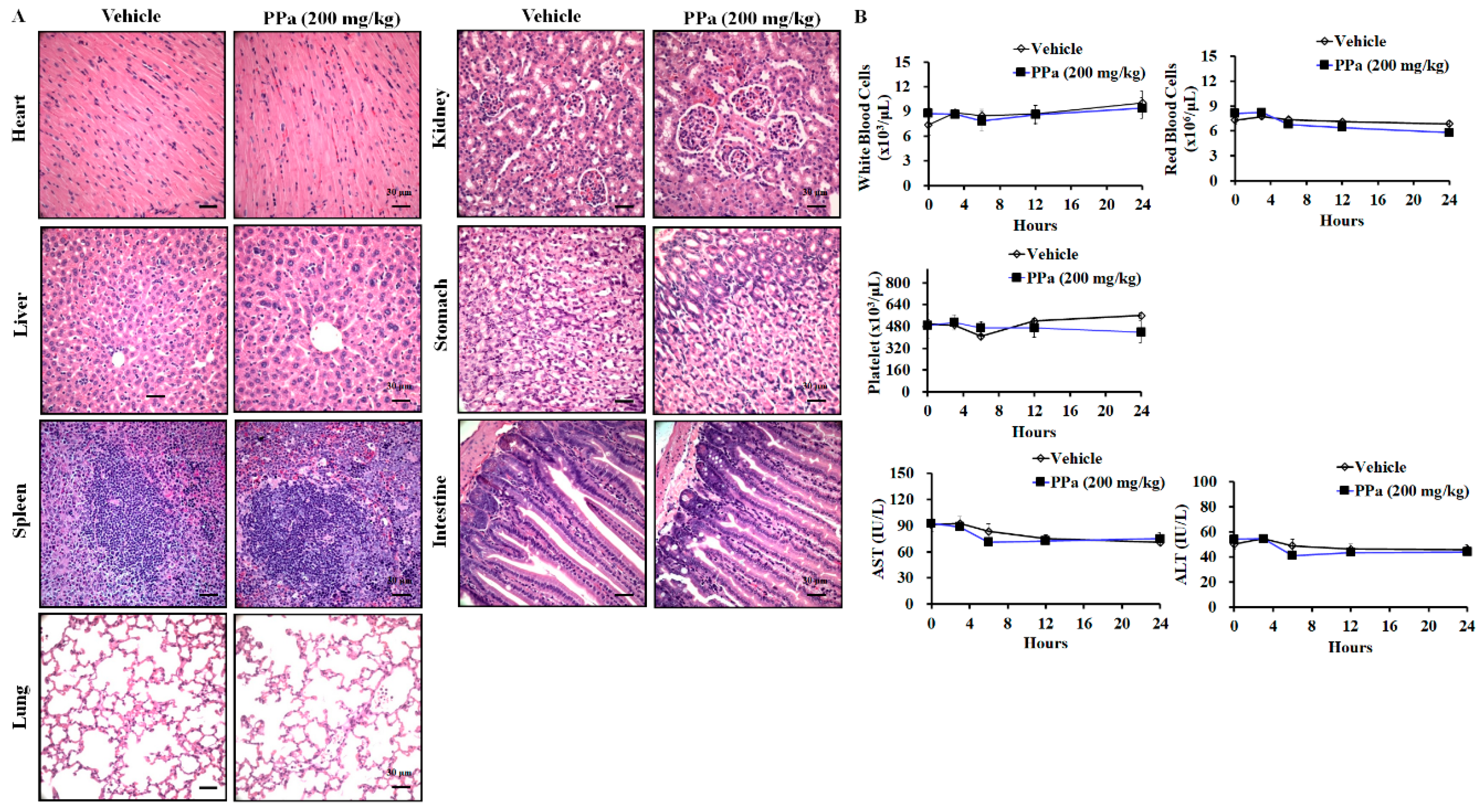
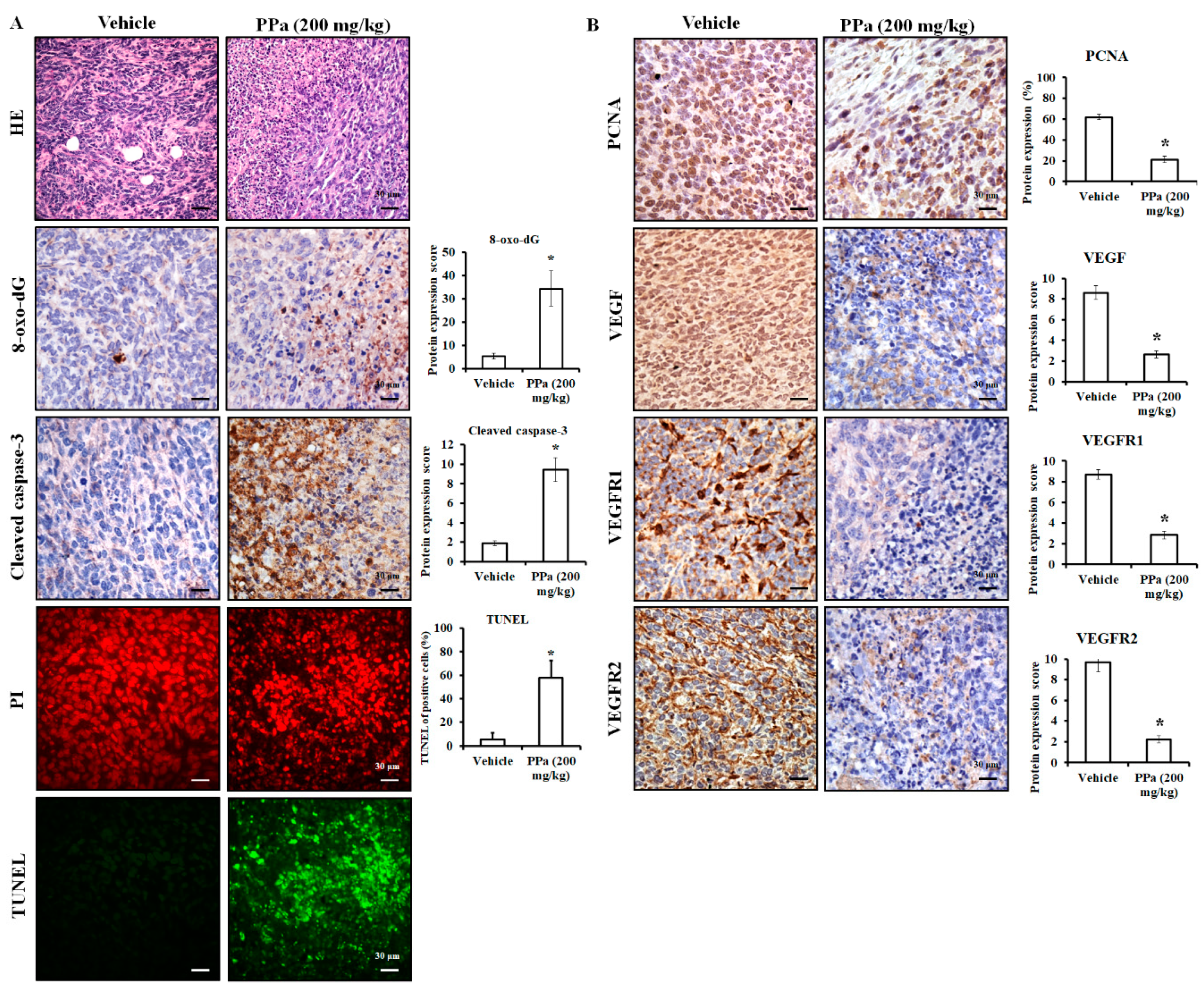
2.6. PPa Extract Suppressed Hepg2 Tumor Growth and Exhibited Less Toxicity in HCC Xenograft Model
2.7. PPa Extract Induced Apoptosis and Reduced Autocrine Proliferation in Xenograft Model
3. Discussion
4. Materials and Methods
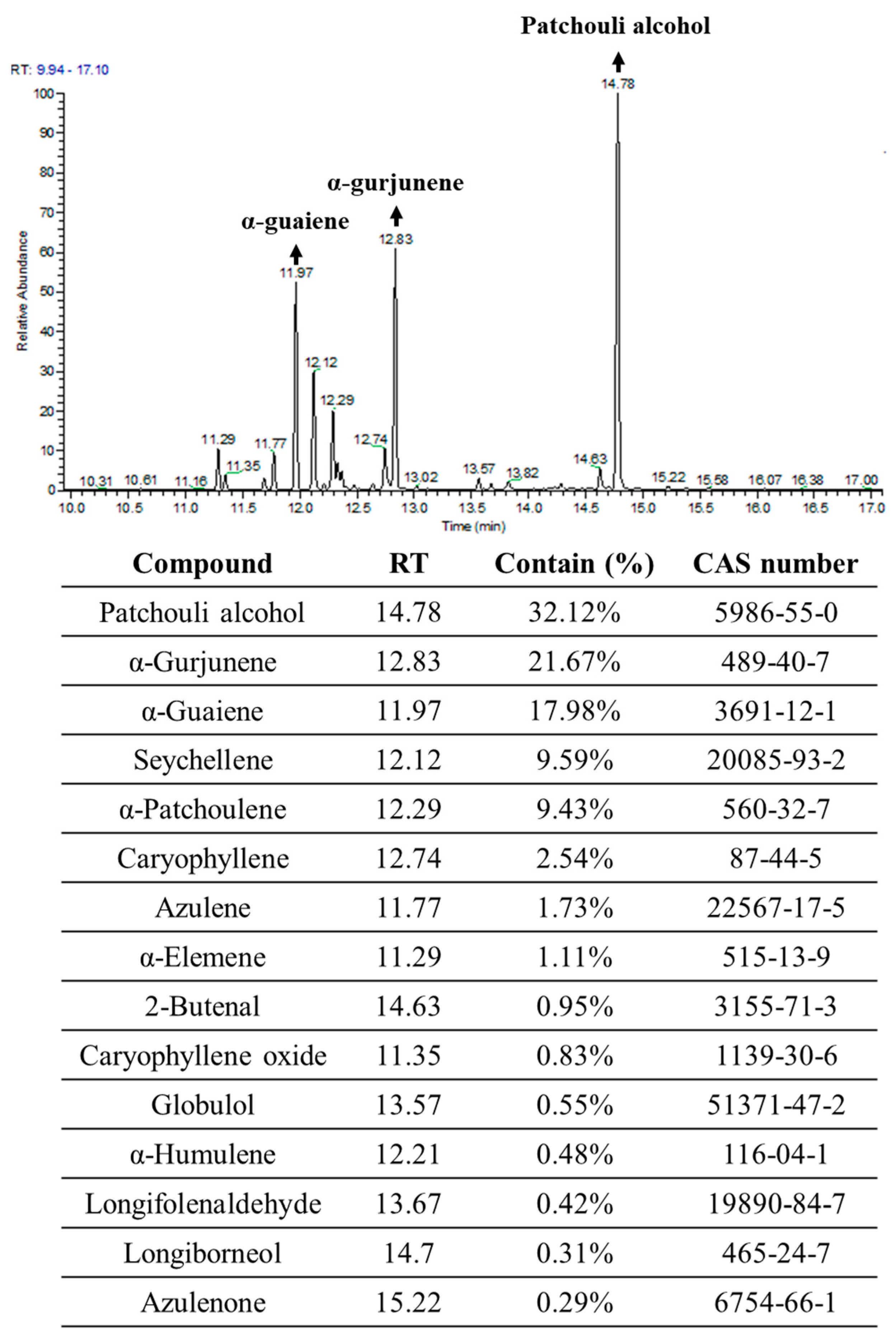
4.1. Extration Essential Oils of Pogostemon Cablin (PPa Extract)
4.2. Cell Culture
4.3. Cell Viability
4.4. Cell Cycle Analysis
4.5. TUNEL Assay
4.6. Western Blot Analysis
4.7. Detection of Reactive Oxygen Species (ROS)
4.8. Mitochondrial Membrane Potential (MMPs, ΔΨm) Assay
4.9. Synergistic Effect of Ppa Extract Plus Sorafenib on Cell Proliferation and Regrowth
4.10. Xenograft Animal Study
4.11. H&E and Immunohistochemistry Staining
4.12. Gas Chromatography-Mass Spectrometry Analysis
4.13. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yu, W.-B.; Rao, A.; Vu, V.; Xu, L.; Rao, J.-Y.; Wu, J.-X. Management of centrally located hepatocellular carcinoma: Update 2016. World J. Hepatol. 2017, 9, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Stagos, D.; Amoutzias, G.D.; Matakos, A.; Spyrou, A.; Tsatsakis, A.M.; Kouretas, D. Chemoprevention of liver cancer by plant polyphenols. Food Chem. Toxicol. 2012, 50, 2155–2170. [Google Scholar] [CrossRef] [PubMed]
- Kudo, M. Systemic Therapy for Hepatocellular Carcinoma: 2017 Update. Oncology 2017, 93 (Suppl. 1), 135–146. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Ricci, S.; Mazzaferro, V.; Hilgard, P.; Gane, E.; Blanc, J.F.; de Oliveira, A.C.; Santoro, A.; Raoul, J.L.; Forner, A.; et al. Sorafenib in advanced hepatocellular carcinoma. N. Engl. J. Med. 2008, 359, 378–390. [Google Scholar] [CrossRef]
- Cheng, A.L.; Kang, Y.K.; Chen, Z.; Tsao, C.J.; Qin, S.; Kim, J.S.; Luo, R.; Feng, J.; Ye, S.; Yang, T.S.; et al. Efficacy and safety of sorafenib in patients in the Asia-Pacific region with advanced hepatocellular carcinoma: A phase III randomised, double-blind, placebo-controlled trial. Lancet Oncol. 2009, 10, 25–34. [Google Scholar] [CrossRef]
- Tsai, N.M.; Lin, S.Z.; Lee, C.C.; Chen, S.P.; Su, H.C.; Chang, W.L.; Harn, H.J. The antitumor effects of Angelica sinensis on malignant brain tumors in vitro and in vivo. Clin. Cancer. Res. 2005, 11, 3475–3484. [Google Scholar] [CrossRef] [Green Version]
- Tsai, N.M.; Chen, Y.L.; Lee, C.C.; Lin, P.C.; Cheng, Y.L.; Chang, W.L.; Lin, S.Z.; Harn, H.J. The natural compound n-butylidenephthalide derived from Angelica sinensis inhibits malignant brain tumor growth in vitro and in vivo. J. Neurochem. 2006, 99, 1251–1262. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Hamid, N.M.; Abass, S.A.; Mohamed, A.A.; Muneam Hamid, D. Herbal management of hepatocellular carcinoma through cutting the pathways of the common risk factors. Biomed. Pharmacother. 2018, 107, 1246–1258. [Google Scholar] [CrossRef]
- Feng, X.X.; Yu, X.T.; Li, W.J.; Kong, S.Z.; Liu, Y.H.; Zhang, X.; Xian, Y.F.; Zhang, X.J.; Su, Z.R.; Lin, Z.X. Effects of topical application of patchouli alcohol on the UV-induced skin photoaging in mice. Eur. J. Pharm. Sci. 2014, 63, 113–123. [Google Scholar] [CrossRef]
- Sangeet Pilkhwal Sah, C.S.M. Kanwaljit Chopra, Antidepressant effect of Valeriana wallichii patchouli alcohol chemotype in mice: Behavioural and biochemical evidence. J. Ethnopharmacol. 2011, 135, 197–200. [Google Scholar] [CrossRef]
- Sah, S.P.; Mathela, C.S.; Chopra, K. Involvement of nitric oxide (NO) signalling pathway in the antidepressant activity of essential oil of Valeriana wallichii Patchouli alcohol chemotype. Phytomedicine 2011, 18, 1269–1275. [Google Scholar] [CrossRef] [PubMed]
- Kocevski, D.; Du, M.; Kan, J.; Jing, C.; Lačanin, I.; Pavlović, H. Antifungal effect of Allium tuberosum, Cinnamomum cassia, and Pogostemon cablin essential oils and their components against population of Aspergillus species. J. Food Sci. 2013, 78, M731–M737. [Google Scholar] [CrossRef] [PubMed]
- Farisa Banu, S.; Rubini, D.; Shanmugavelan, P.; Murugan, R.; Gowrishankar, S.; Karutha Pandian, S.; Nithyanand, P. Effects of patchouli and cinnamon essential oils on biofilm and hyphae formation by Candida species. J. Mycol. Med. 2018, 28, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.L.; Ju, D.H.; Chen, J.; Yu, B.; Liu, K.L.; He, J.X.; Dai, C.Q.; Wu, S.; Chang, Z.; Wang, Y.P.; et al. Immunologic Mechanism of Patchouli Alcohol Anti-H1N1 Influenza Virus May through Regulation of the RLH Signal Pathway In Vitro. Curr. Microbiol. 2013, 67, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Lin, Z.; Xian, Y.; Kong, S.; Lai, Z.; Ip, S.; Chen, H.; Guo, H.; Su, Z.; Yang, X.; et al. (-)-Patchouli alcohol protects against Helicobacter pylori urease-induced apoptosis, oxidative stress and inflammatory response in human gastric epithelial cells. Int. Immunopharmacol. 2016, 35, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.; Dou, Y.; Wu, X.; Li, H.; Wu, J.; Huang, Q.; Luo, D.; Yi, T.; Liu, Y.; Su, Z.; et al. Prophylactic efficacy of patchoulene epoxide against ethanol-induced gastric ulcer in rats: Influence on oxidative stress, inflammation and apoptosis. Chem. Biol. Interact. 2018, 283, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Z.; Liu, Y.H.; Liang, J.L.; Huang, Q.H.; Dou, Y.X.; Nie, J.; Zhuo, J.Y.; Wu, X.; Chen, J.N.; Su, Z.R.; et al. Protective role of beta-patchoulene from Pogostemon cablin against indomethacin-induced gastric ulcer in rats: Involvement of anti-inflammation and angiogenesis. Phytomedicine 2018, 39, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Beaumont, C.; Stevens, N. Chemical composition analysis and in vitro biological activities of ten essential oils in human skin cells. Biochimie Open 2017, 5, 1–7. [Google Scholar] [CrossRef]
- Tsai, C.C.; Chang, Y.H.; Chang, C.C.; Cheng, Y.M.; Ou, Y.C.; Chien, C.C.; Hsu, Y.C. Induction of Apoptosis in Endometrial Cancer (Ishikawa) Cells by Pogostemon cablin Aqueous Extract (PCAE). Int. J. Mol. Sci. 2015, 16, 12424–12435. [Google Scholar] [CrossRef] [Green Version]
- Chien, J.-H.; Lee, S.-C.; Chang, K.-F.; Huang, X.-F.; Chen, Y.-T.; Tsai, N.-M. Extract of Pogostemon cablin Possesses Potent Anticancer Activity against Colorectal Cancer Cells In Vitro and In Vivo. Evid. Based Complement. Alternat. Med. 2020, 2020, 9758156. [Google Scholar] [CrossRef]
- Singh, K.; Gangrade, A.; Jana, A.; Mandal, B.B.; Das, N. Design, Synthesis, Characterization, and Antiproliferative Activity of Organoplatinum Compounds Bearing a 1,2,3-Triazole Ring. ACS Omega 2019, 4, 835–841. [Google Scholar] [CrossRef]
- Keller, D.M.; Zeng, X.; Wang, Y.; Zhang, Q.H.; Kapoor, M.; Shu, H.; Goodman, R.; Lozano, G.; Zhao, Y.; Lu, H. A DNA damage-induced p53 serine 392 kinase complex contains CK2, hSpt16, and SSRP1. Mol. Cell 2001, 7, 283–292. [Google Scholar] [CrossRef]
- Sova, H.; Jukkola-Vuorinen, A.; Puistola, U.; Kauppila, S.; Karihtala, P. 8-Hydroxydeoxyguanosine: A new potential independent prognostic factor in breast cancer. Br. J. Cancer 2010, 102, 1018–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.F.; Pervaiz, S. Assessment of oxidative stress-induced DNA damage by immunoflourescent analysis of 8-oxodG. Methods Cell Biol. 2011, 103, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Costantini, S.; Di Bernardo, G.; Cammarota, M.; Castello, G.; Colonna, G. Gene expression signature of human HepG2 cell line. Gene 2013, 518, 335–345. [Google Scholar] [CrossRef]
- Lin, Y.; Ong, L.K.; Chan, S.H. Differential in situ hybridization for determination of mutational specific expression of the p53 gene in human hepatoma cell lines. Pathology 1995, 27, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.P.; Schwank, J.; Staib, F.; Wang, X.W.; Harris, C.C. TP53 mutations and hepatocellular carcinoma: Insights into the etiology and pathogenesis of liver cancer. Oncogene 2007, 26, 2166–2176. [Google Scholar] [CrossRef] [Green Version]
- Oda, T.; Tsuda, H.; Scarpa, A.; Sakamoto, M.; Hirohashi, S. p53 gene mutation spectrum in hepatocellular carcinoma. Cancer Res. 1992, 52, 6358–6364. [Google Scholar]
- Honda, K.; Sbisa, E.; Tullo, A.; Papeo, P.A.; Saccone, C.; Poole, S.; Pignatelli, M.; Mitry, R.R.; Ding, S.; Isla, A.; et al. p53 mutation is a poor prognostic indicator for survival in patients with hepatocellular carcinoma undergoing surgical tumour ablation. Br. J. Cancer 1998, 77, 776–782. [Google Scholar] [CrossRef] [Green Version]
- Tornesello, M.L.; Buonaguro, L.; Tatangelo, F.; Botti, G.; Izzo, F.; Buonaguro, F.M. Mutations in TP53, CTNNB1 and PIK3CA genes in hepatocellular carcinoma associated with hepatitis B and hepatitis C virus infections. Genomics 2013, 102, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Karimian, A.; Ahmadi, Y.; Yousefi, B. Multiple functions of p21 in cell cycle, apoptosis and transcriptional regulation after DNA damage. DNA Repair 2016, 42, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Sherr, C.J.; McCormick, F. The RB and p53 pathways in cancer. Cancer Cell 2002, 2, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Abbas, T.; Dutta, A. p21 in cancer: Intricate networks and multiple activities. Nat. Rev. Cancer 2009, 9, 400–414. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Chen, Y.; St Clair, D.K. ROS and p53: A versatile partnership. Free Radic. Biol. Med. 2008, 44, 1529–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef] [PubMed]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Shin, M.S.; Lee, H.S.; Bae, J.H.; Lee, H.K.; Kim, H.S.; Kim, S.Y.; Jang, J.J.; Joo, M.; Kang, Y.K.; et al. Expression of Fas and Fas-related molecules in human hepatocellular carcinoma. Hum. Pathol. 2001, 32, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Jodo, S.; Kobayashi, S.; Nakajima, Y.; Matsunaga, T.; Nakayama, N.; Ogura, N.; Kayagaki, N.; Okumura, K.; Koike, T. Elevated serum levels of soluble Fas/APO-1 (CD95) in patients with hepatocellular carcinoma. Clin. Exp. Immunol. 1998, 112, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Cande, C.; Cohen, I.; Daugas, E.; Ravagnan, L.; Larochette, N.; Zamzami, N.; Kroemer, G. Apoptosis-inducing factor (AIF): A novel caspase-independent death effector released from mitochondria. Biochimie 2002, 84, 215–222. [Google Scholar] [CrossRef]
- Man, Y.N.; Liu, X.H.; Wu, X.Z. Chinese medicine herbal treatment based on syndrome differentiation improves the overall survival of patients with unresectable hepatocellular carcinoma. Chin. J. Integr. Med. 2015, 21, 49–57. [Google Scholar] [CrossRef]
- Shu, X.; McCulloch, M.; Xiao, H.; Broffman, M.; Gao, J. Chinese herbal medicine and chemotherapy in the treatment of hepatocellular carcinoma: A meta-analysis of randomized controlled trials. Integr. Cancer Ther. 2005, 4, 219–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriwaki, H. Prevention of liver cancer: Basic and clinical aspects. Exp. Mol. Med. 2002, 34, 319–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coriat, R.; Nicco, C.; Chéreau, C.; Mir, O.; Alexandre, J.; Ropert, S.; Weill, B.; Chaussade, S.; Goldwasser, F.; Batteux, F. Sorafenib-induced hepatocellular carcinoma cell death depends on reactive oxygen species production in vitro and in vivo. Mol. Cancer Ther. 2012, 11, 2284–2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxid. Med. Cell. Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Lui, V.W.; Yeo, W. Targeting the PI3K/Akt/mTOR pathway in hepatocellular carcinoma. Future Oncol. 2011, 7, 1149–1167. [Google Scholar] [CrossRef]
- Zhai, B.; Hu, F.; Jiang, X.; Xu, J.; Zhao, D.; Liu, B.; Pan, S.; Dong, X.; Tan, G.; Wei, Z.; et al. Inhibition of Akt reverses the acquired resistance to sorafenib by switching protective autophagy to autophagic cell death in hepatocellular carcinoma. Mol. Cancer Ther. 2014, 13, 1589–1598. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhou, X.; Shen, H.; Wang, D.; Wang, Y. Phosphorylated ERK is a potential predictor of sensitivity to sorafenib when treating hepatocellular carcinoma: Evidence from an in vitro study. BMC Med. 2009, 7, 41. [Google Scholar] [CrossRef] [Green Version]
- Tseng, P.L.; Tai, M.H.; Huang, C.C.; Wang, C.C.; Lin, J.W.; Hung, C.H.; Chen, C.H.; Wang, J.H.; Lu, S.N.; Lee, C.M.; et al. Overexpression of VEGF is associated with positive p53 immunostaining in hepatocellular carcinoma (HCC) and adverse outcome of HCC patients. J. Surg. Oncol. 2008, 98, 349–357. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, J.N.; Tang, J.M.; Kong, X.; Yang, J.Y.; Zheng, F.; Guo, L.Y.; Huang, Y.Z.; Zhang, L.; Tian, L.; et al. VEGF is essential for the growth and migration of human hepatocellular carcinoma cells. Mol. Biol. Rep. 2012, 39, 5085–5093. [Google Scholar] [CrossRef] [Green Version]
- Mise, M.; Arii, S.; Higashituji, H.; Furutani, M.; Niwano, M.; Harada, T.; Ishigami, S.; Toda, Y.; Nakayama, H.; Fukumoto, M.; et al. Clinical significance of vascular endothelial growth factor and basic fibroblast growth factor gene expression in liver tumor. Hepatology 1996, 23, 455–464. [Google Scholar] [CrossRef]
- Zhan, P.; Qian, Q.; Yu, L.K. Prognostic significance of vascular endothelial growth factor expression in hepatocellular carcinoma tissue: A meta-analysis. Hepatobiliary Surg. Nutr. 2013, 2, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.B.; Choi, J.; Lou, Z.; Jiang, X.; Lee, S.H. Patchouli alcohol, an essential oil of Pogostemon cablin, exhibits anti-tumorigenic activity in human colorectal cancer cells. Int. Immunopharmacol. 2013, 16, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Yang, L.; Lu, C.; Xu, Z.; Qiu, H.; Wu, J.; Wang, J.; Tong, J.; Zhu, Y.; Shen, J. Molecular Role of EGFR-MAPK Pathway in Patchouli Alcohol-Induced Apoptosis and Cell Cycle Arrest on A549 Cells In Vitro and In Vivo. Biomed. Res. Int. 2016, 2016, 4567580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayaz, F.; Yuzer, A.; Ince, T.; Ince, M. Anti-Cancer and Anti-Inflammatory Activities of Bromo- and Cyano-Substituted Azulene Derivatives. Inflammation 2020, 43, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Colerangle, J.B. Chapter 25: Preclinical Development of Nononcogenic Drugs (Small and Large Molecules). In A Comprehensive Guide to Toxicology in Nonclinical Drug Development, 2nd ed.; Faqi, A.S., Ed.; Academic Press: Boston, MA, USA, 2017; pp. 659–683. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Tumor Type | PPa Extract | SOR | VP-16 | 5-FU |
|---|---|---|---|---|---|
| Hepatocellular Carcinoma Cells | |||||
| HepG2 | Human HCC cell | 20.09 ± 2.21 a,b | 3.52 ± 1.97 | 4.73 ± 3.57 | 2.09 ± 1.63 |
| Mahlavu | Human HCC cell | 33.29 ± 2.72 a,b | 6.07 ± 1.06 | 4.54 ± 2.17 | 14.97 ± 3.71 |
| J5 | Human HCC cell | 29.87 ± 3.62 a,b | 2.39 ± 1.91 | 3.01 ± 2.75 | 18.79 ± 0.91 |
| Huh7 | Human HCC cell | 7.34 ± 3.09 a,b | 1.77 ± 4.31 | 5.11 ± 2.08 | 4.88 ± 3.37 |
| Normal cells | |||||
| SVEC | Mouse vascular endothelial cell | 69.68 ± 4.63 c | 8.55 ± 2.73 | 1.85 ± 0.49 | 1.69 ± 2.8 |
| MDCK | Canine epithelial kidney cell | 73.61 ± 0.16 c | 6.95 ± 1.45 | 3.38 ± 0.43 | 8.74 ± 0.53 |
| BNL CL.2 | Mouse liver embryonic cell | 147.24 ± 7.71 c | 10.75 ± 5.17 | 6.43 ± 3.02 | >20 |
| Normal Cells | /Tumor Cells | PPa Extract | SOR | VP-16 |
|---|---|---|---|---|
| SVEC | /HepG | 3.5 | 2.4 | 0.4 |
| /Mahlavu | 2.1 | 1.4 | 0.4 | |
| /J5 | 2.3 | 3.6 | 0.6 | |
| /Huh7 | 9.5 | 4.8 | 0.4 | |
| MDCK | /HepG2 | 3.7 | 2.0 | 0.7 |
| /Mahlavu | 2.2 | 1.1 | 0.8 | |
| /J5 | 2.5 | 2.9 | 1.1 | |
| /Huh7 | 10 | 3.9 | 0.7 | |
| BNL CL.2 | /HepG2 | 7.4 | 3.1 | 1.4 |
| /Mahlavu | 4.4 | 1.8 | 1.4 | |
| /J5 | 4.9 | 4.5 | 2.1 | |
| /Huh7 | 20.1 | 6.1 | 1.3 |
Sample Availability: Samples of the essential oils of Pogostemon Cablin are available from the authors. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.-F.; Sheu, G.-T.; Chang, K.-F.; Huang, Y.-C.; Hung, P.-H.; Tsai, N.-M. Pogostemon cablin Triggered ROS-Induced DNA Damage to Arrest Cell Cycle Progression and Induce Apoptosis on Human Hepatocellular Carcinoma In Vitro and In Vivo. Molecules 2020, 25, 5639. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25235639
Huang X-F, Sheu G-T, Chang K-F, Huang Y-C, Hung P-H, Tsai N-M. Pogostemon cablin Triggered ROS-Induced DNA Damage to Arrest Cell Cycle Progression and Induce Apoptosis on Human Hepatocellular Carcinoma In Vitro and In Vivo. Molecules. 2020; 25(23):5639. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25235639
Chicago/Turabian StyleHuang, Xiao-Fan, Gwo-Tarng Sheu, Kai-Fu Chang, Ya-Chih Huang, Pei-Hsiu Hung, and Nu-Man Tsai. 2020. "Pogostemon cablin Triggered ROS-Induced DNA Damage to Arrest Cell Cycle Progression and Induce Apoptosis on Human Hepatocellular Carcinoma In Vitro and In Vivo" Molecules 25, no. 23: 5639. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25235639