CB1 Receptor-Dependent and Independent Induction of Lipolysis in Primary Rat Adipocytes by the Inverse Agonist Rimonabant (SR141716A)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
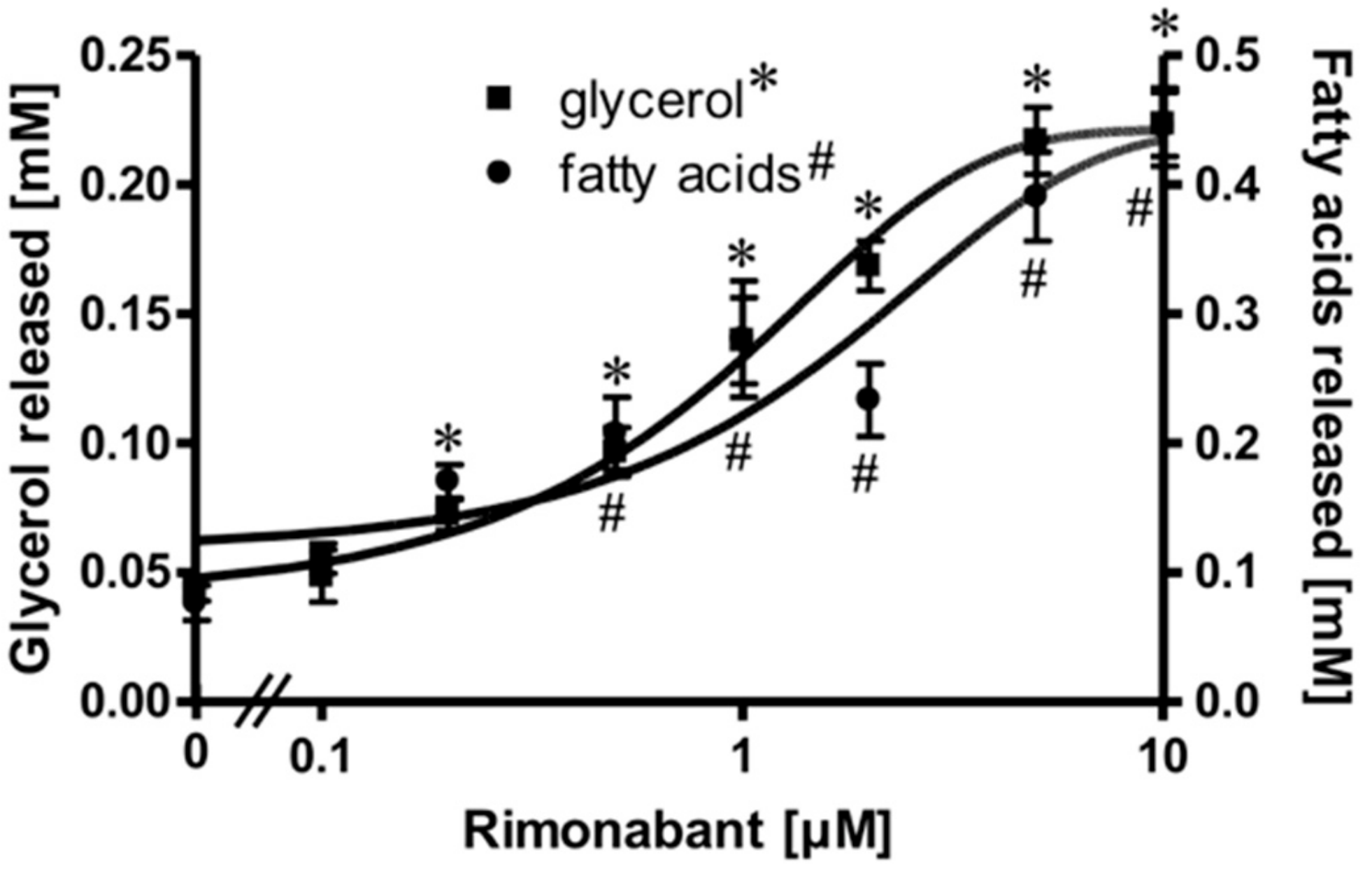
2.1. Rimonabant Stimulates Lipolysis in Primary Rat Adipocytes
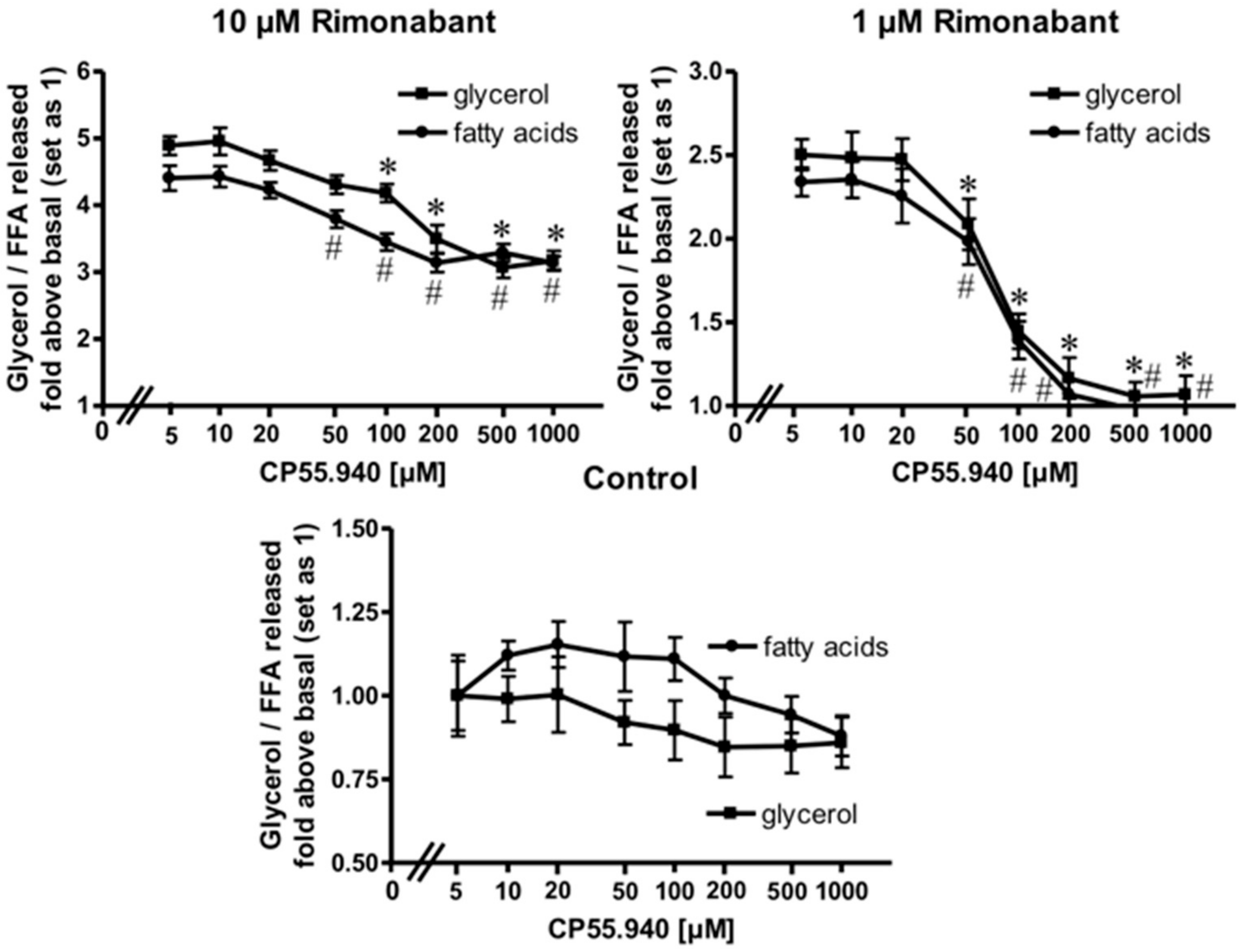
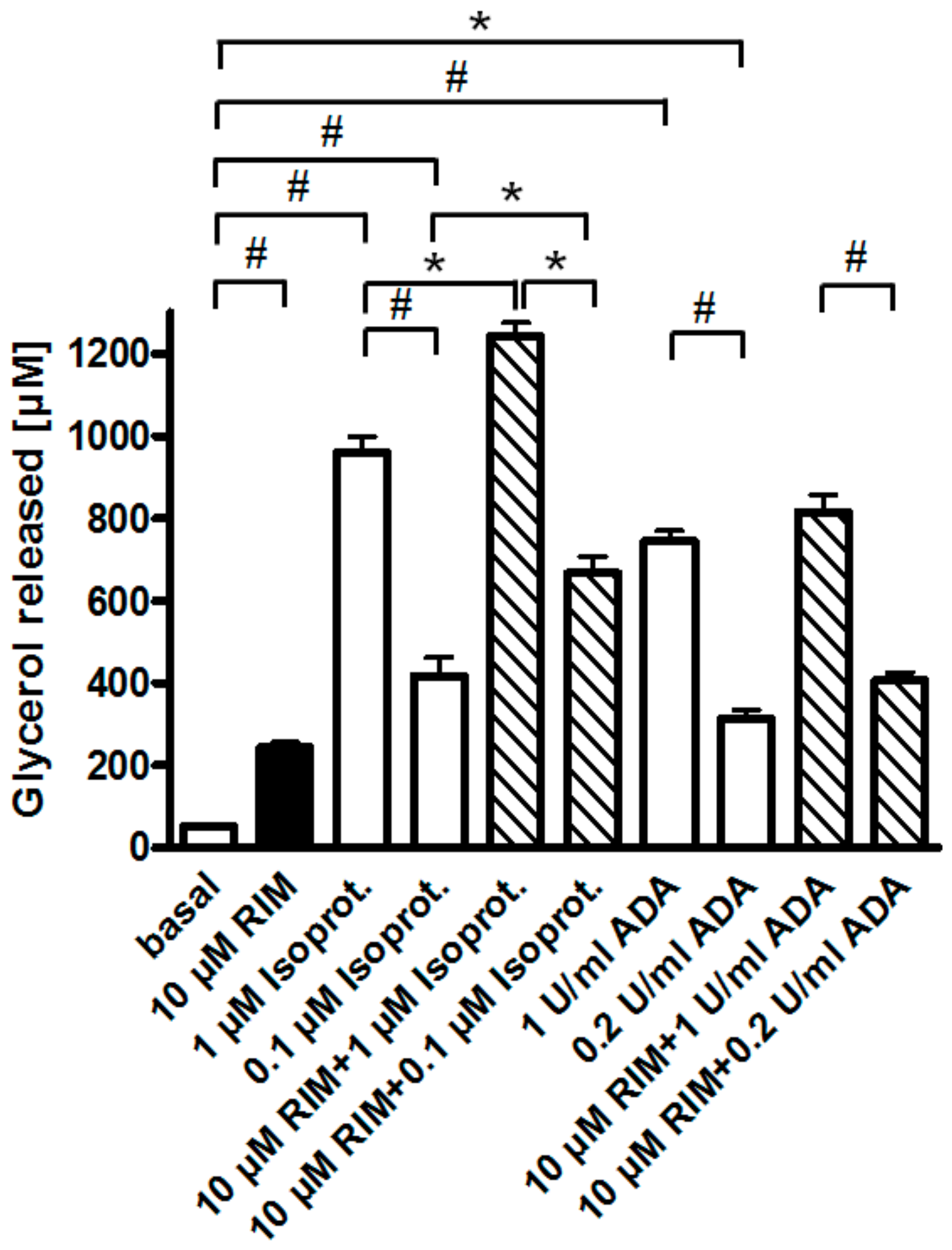
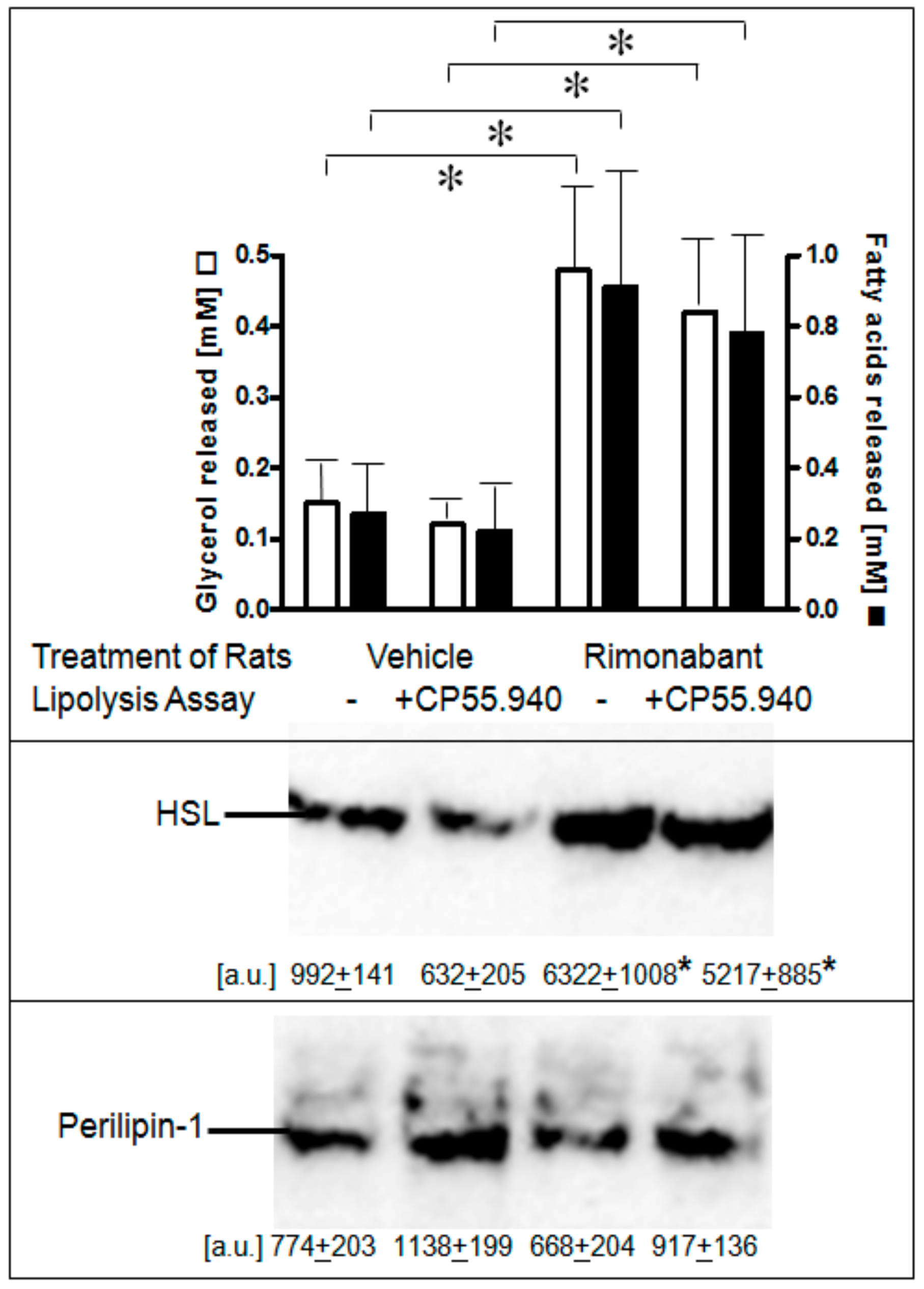
2.2. Rimonabant Stimulates Lipolysis by CB1R-Dependent and Independent Mechanisms
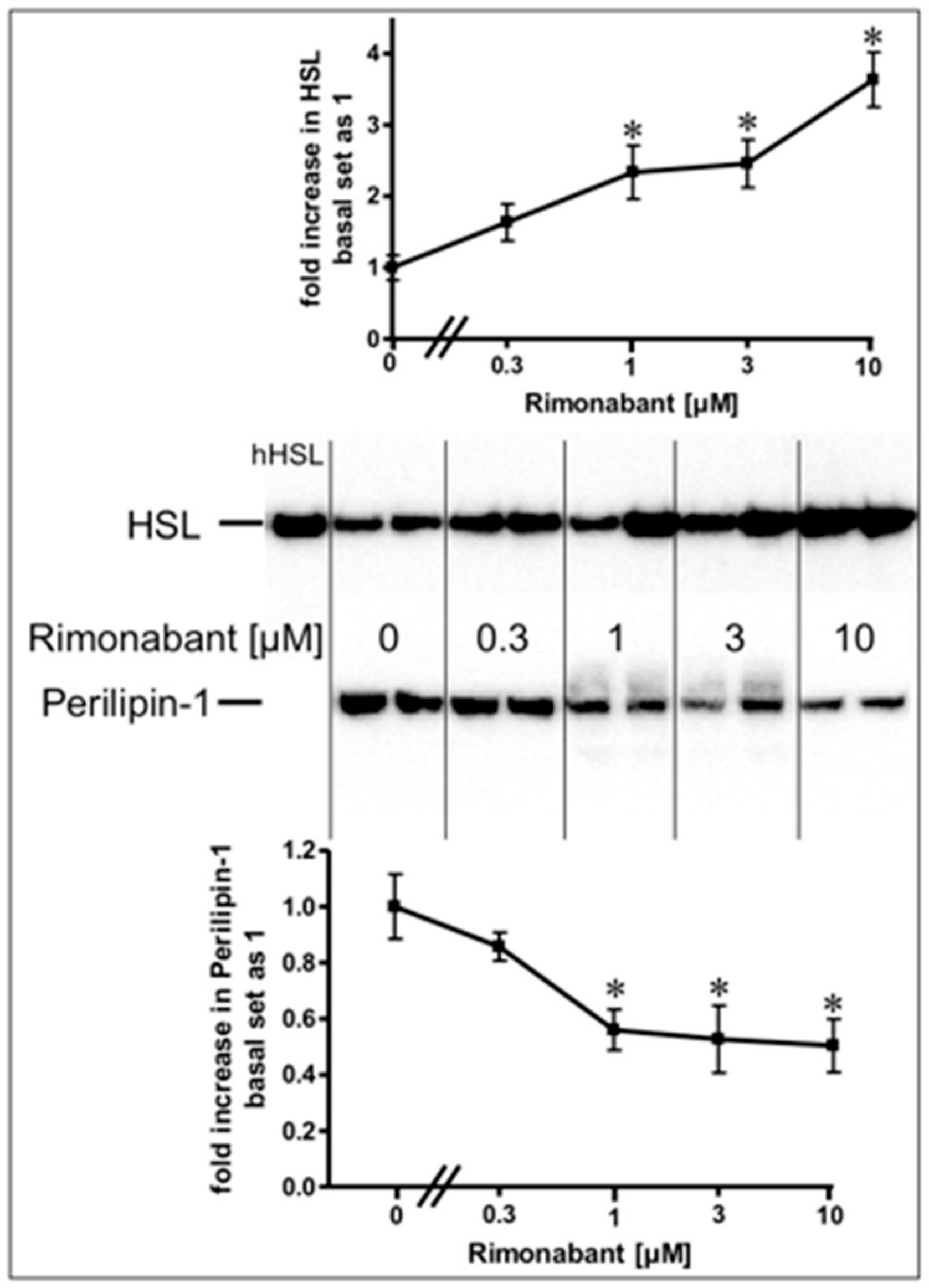
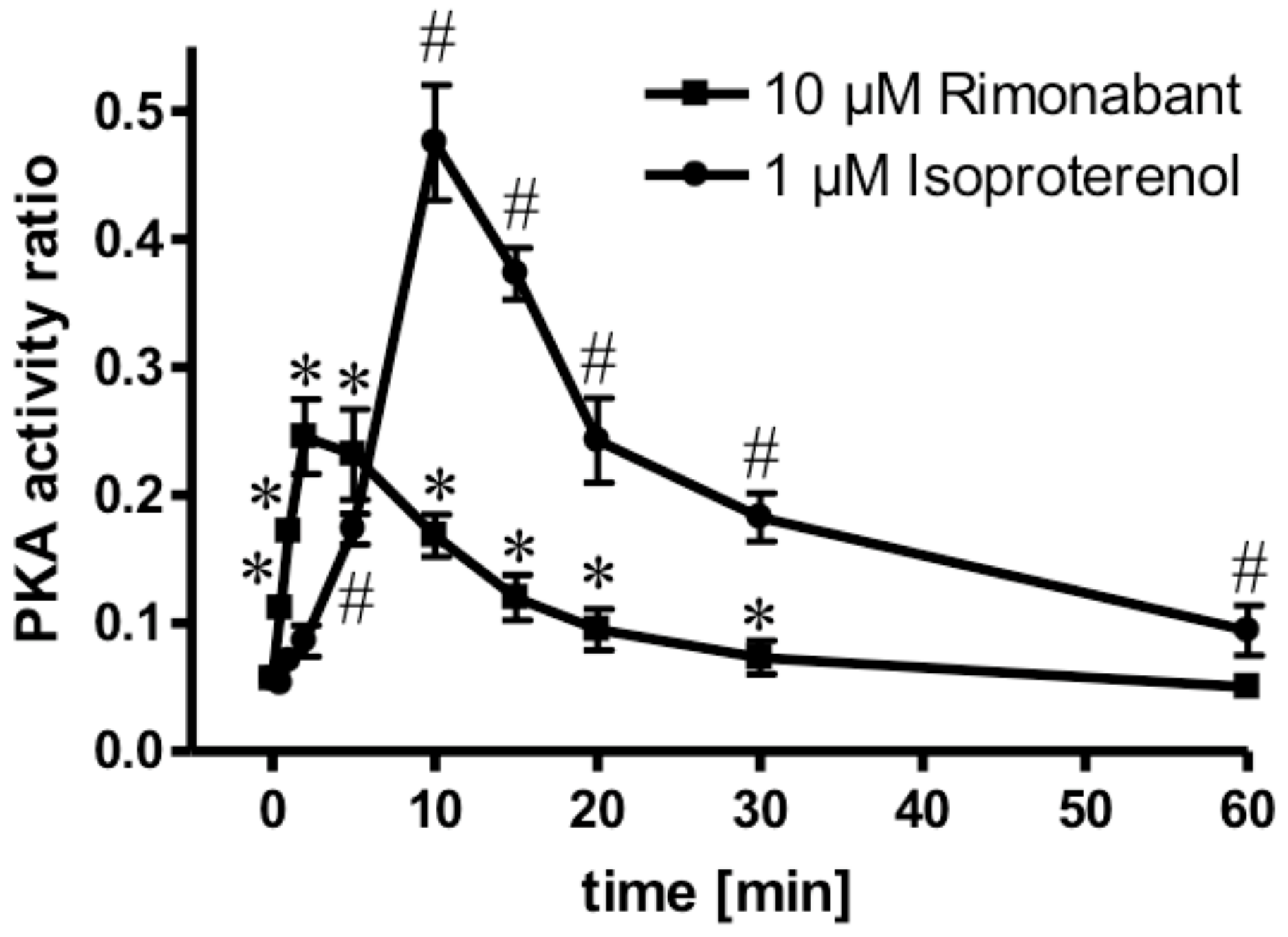
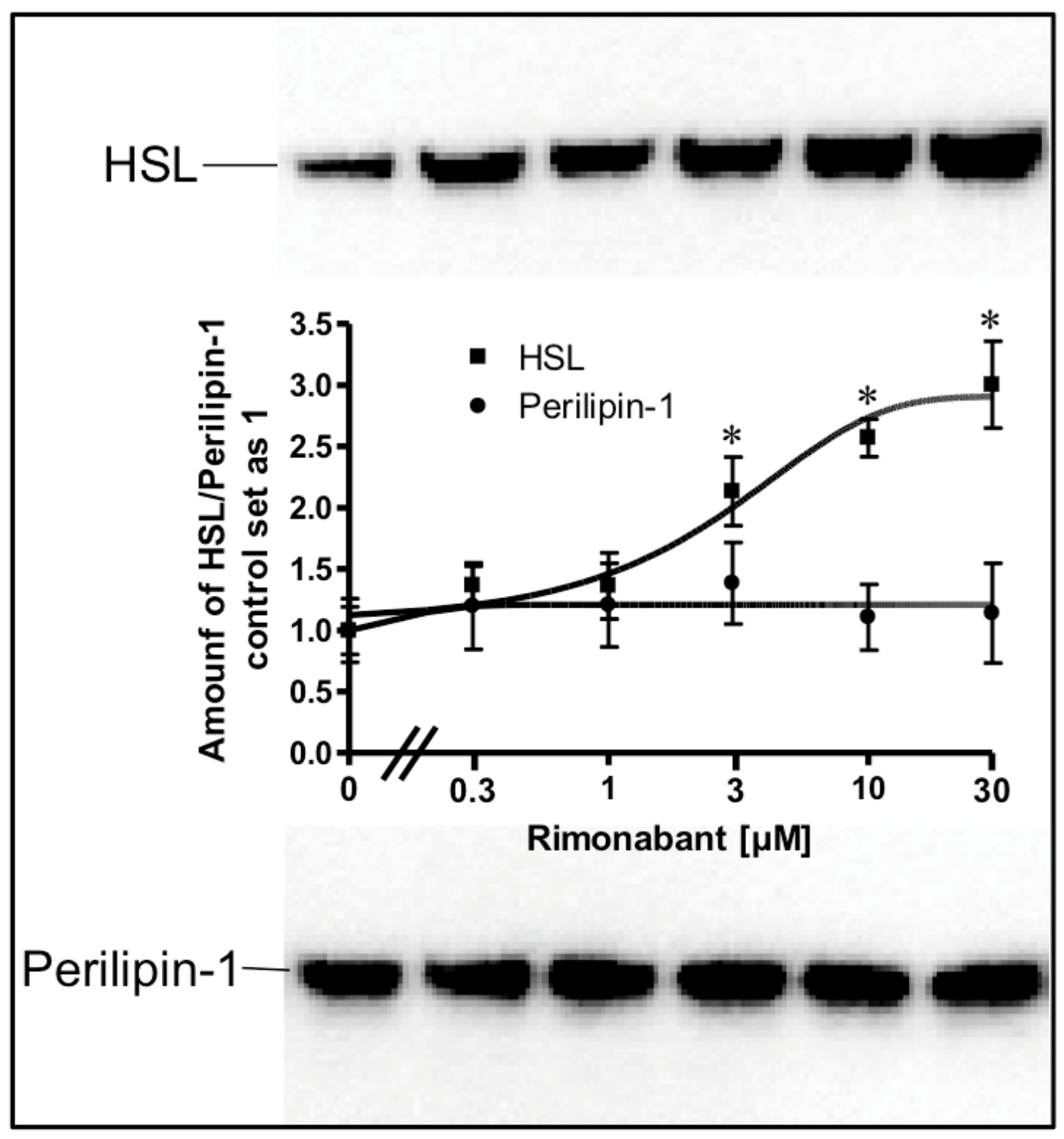
2.3. Rimonabant Stimulates PKA (Protein Kinase A) in Adipocytes
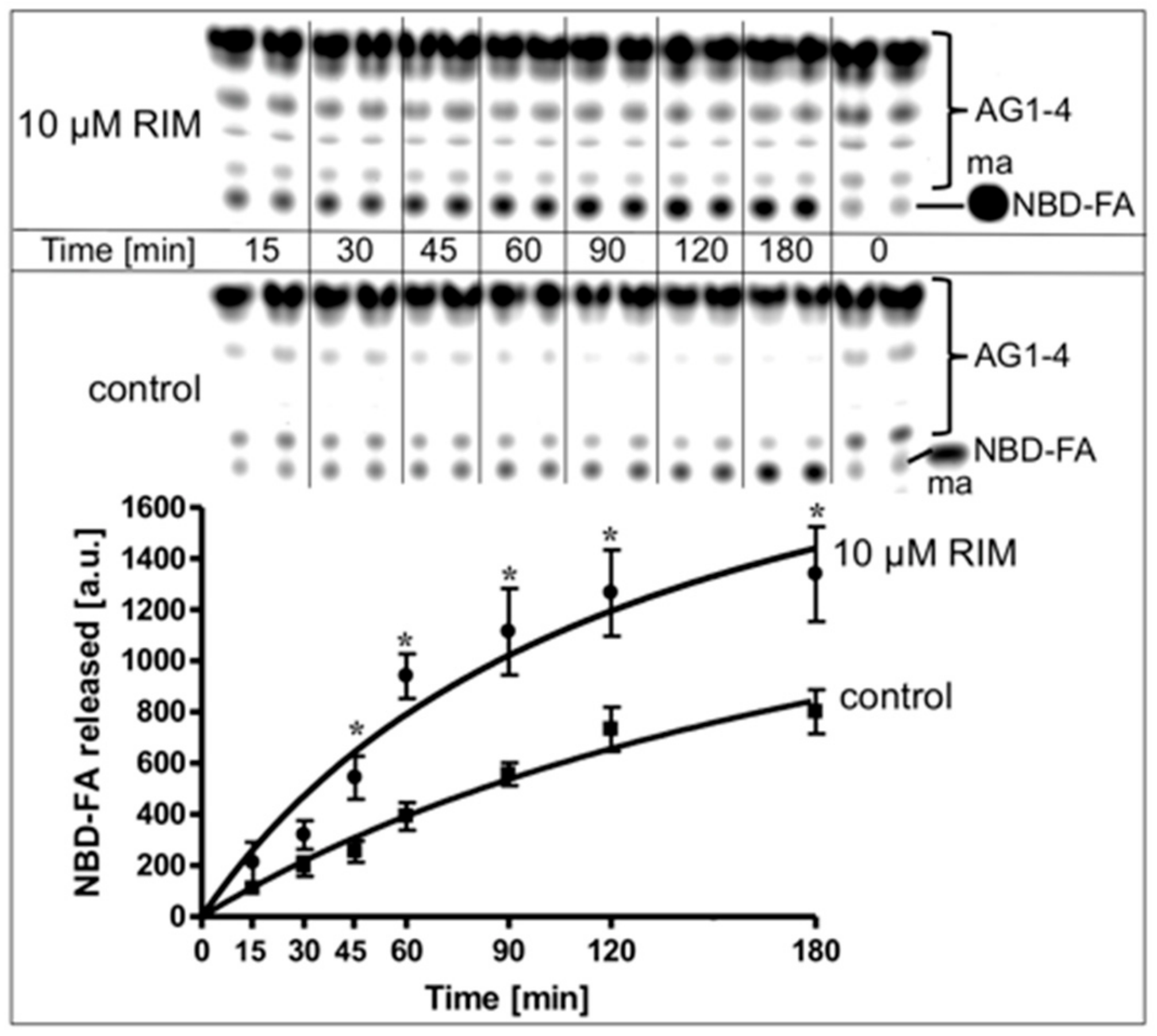
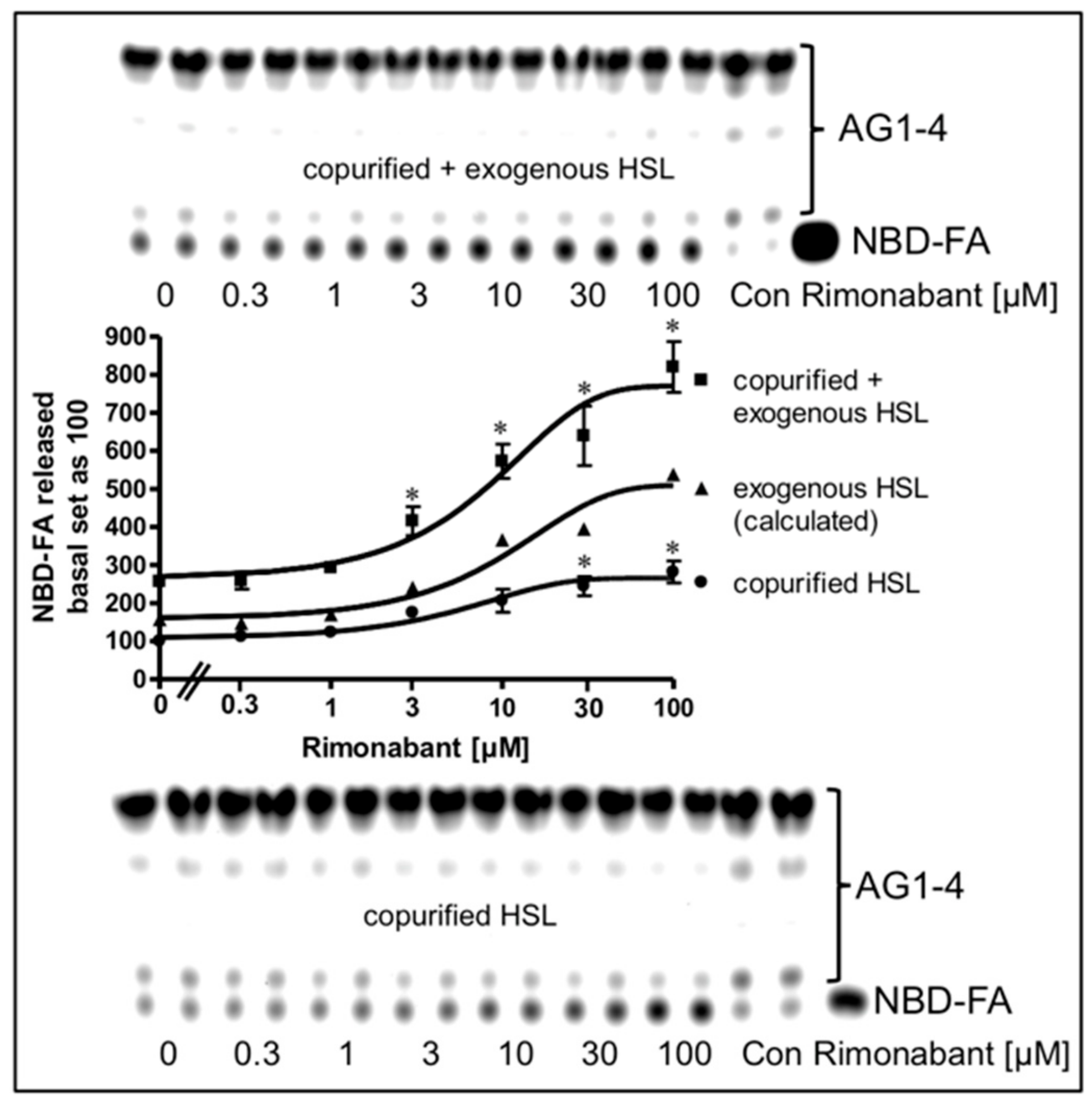
2.4. Rimonabant Increases Accessibility of LD for Lipolytic Cleavage in a Cell-Free System
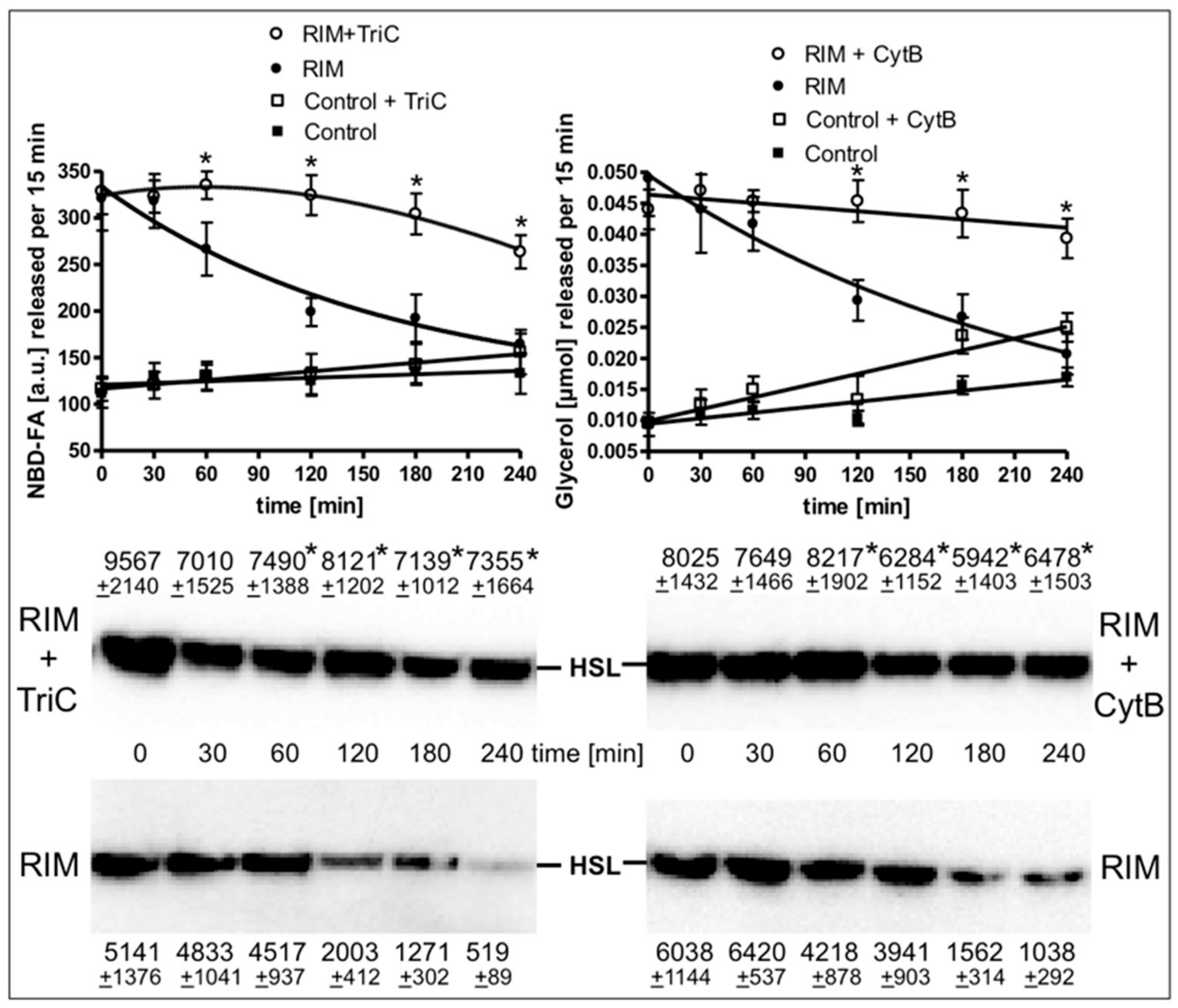
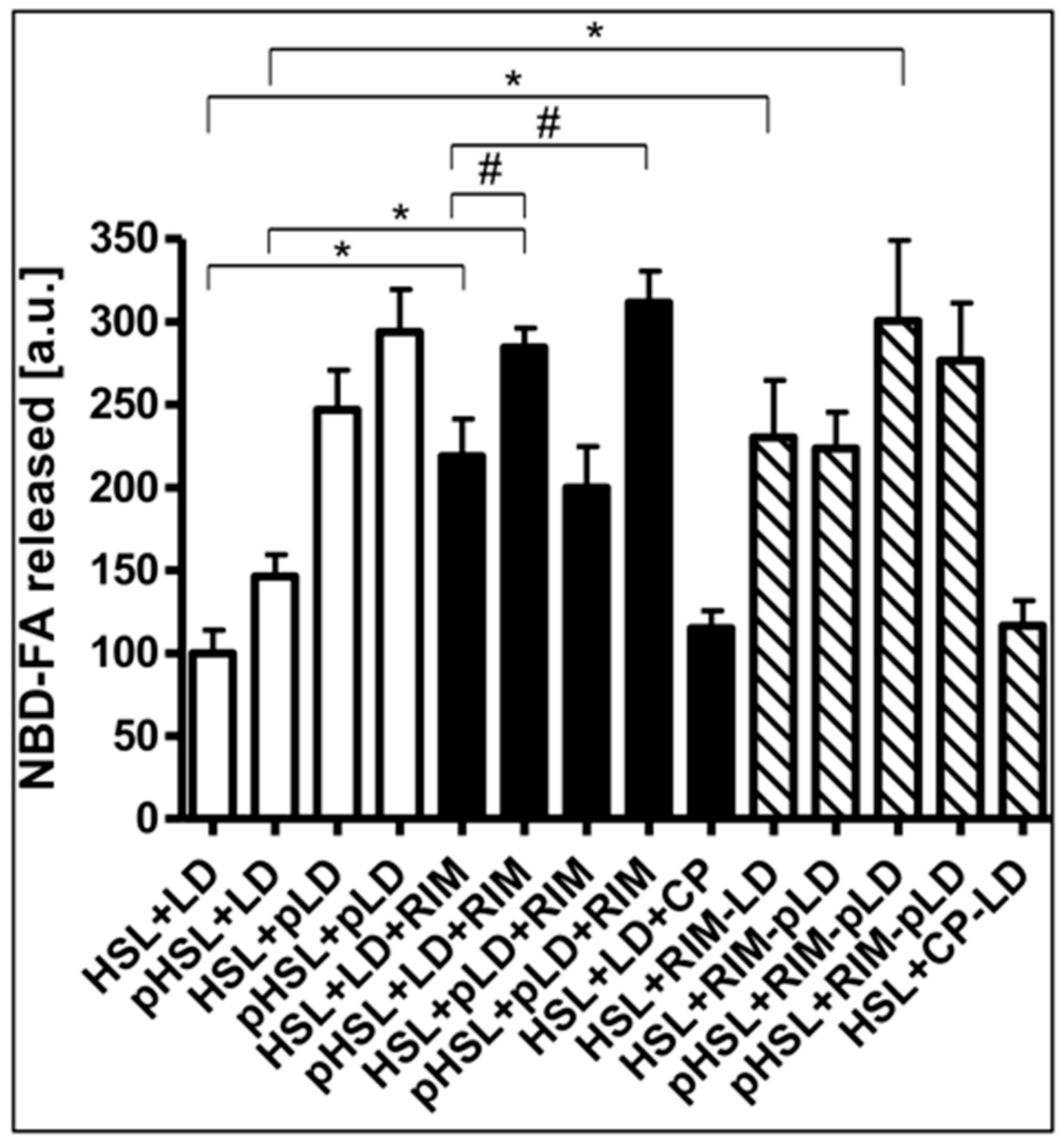
2.5. Rimonabant Improves Accessibility of LD for Lipolytic Cleavage in Intact Adipocytes
3. Discussion
3.1. The Direct CB1R-Dependent Lipolytic Activity of Rimonabant
3.2. The Direct CB1R-Independent Lipolytic Activity of Rimonabant
3.3. Physiological Relevance of the Direct Lipolytic Activity of Rimonabant
4. Materials and Methods
4.1. Animal Handling and Rimonabant Treatment
4.2. Preparation and Incubation of Primary Rat Adipocytes
4.3. Protein and Lipid Composition of LD from Adipocytes
4.4. Cell-Free Lipolysis System
4.5. Phosphorylation of LD and HSL In Vitro
4.6. Lipid Analysis
4.7. PKA Activity Ratio
4.8. Statistics
4.9. Miscellaneous
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Horn, H.; Böhme, B.; Dietrich, L.; Koch, M. Endocannabinoids in body weight control. Pharmaceuticals 2018, 11, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devane, W.A.; Dysarz, F.A.; Johnson, M.R.; Melvin, L.S.; Howlett, A.C. Determination and characterization of a cannabinoid receptor in rat brain. Mol. Pharmacol. 1988, 34, 605–613. [Google Scholar] [PubMed]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Galiegue, S.; Mary, S.; Marchand, J.; Dussossoy, D.; Carriere, D.; Carayon, P.; Bouaboula, M.; Shire, D.; Le Fur, G.; Casellas, P. Expression of central and peripheral cannabinoid receptors in human tissues and leukocyte subpopulations. Eur. J. Biochem. 1995, 232, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Simon, V.; Cota, D. Mechanisms in Endocrinology: Endocannabinoids and metabolism: Past, present and future. Eur. J. Endocrinol. 2017, 176, R309–R324. [Google Scholar] [CrossRef] [Green Version]
- Cohen, K.; Weizman, A.; Weinstein, A. Positive and negative effects of cannabis and cannabinoids on health. Clin. Pharmacol. Ther. 2019. [Google Scholar] [CrossRef]
- Cota, D.; Marsicano, G.; Tschöp, M.; Grubler, Y.; Flachskamm, C.; Schubert, M.; Auer, D.; Yassouridis, A.; Thone-Reineke, C.; Ortmann, S.; et al. The endogenous cannabinoid system affects energy balance via central orexigenic drive and peripheral lipogenesis. J. Clin. Invest. 2003, 112, 423–431. [Google Scholar] [CrossRef]
- Dinh, T.P.; Carpenter, D.; Leslie, F.M.; Freund, T.F.; Katona, I.; Sensi, S.L.; Kathuria, S.; Piomelli, D. Brain monoglyceride lipase participating in endocannabinoid inactivation. Proc. Natl. Acad. Sci. USA 2002, 99, 10819–10824. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, M.; Contreras, J.A.; Hellman, U.; Tornqvist, H.; Holm, C. cDNA cloning, tissue distribution, and identification of the catalytic triad of monoglyceride lipase, Evolutionary relationship to esterases, lysophospholipases, and haloperoxidases. J. Biol. Chem. 1997, 272, 27218–27223. [Google Scholar] [CrossRef] [Green Version]
- Gruden, G.; Barutta, F.; Kunos, G.; Pacher, P. Role of the endocannabinoid system in diabetic and diabetic complications. Br. J. Pharmacol. 2016, 173, 1116–1127. [Google Scholar] [CrossRef]
- Krott, L.M.; Piscitelli, F.; Heine, M.; Borrino, S.; Scheja, L.; Silvestri, C.; Heeren, J.; Di Marzo, V. Endocannabinoid regulation in white and brown adipose tissue following thermogenic activation. J. Lipid Res. 2016, 57, 464–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engeli, S.; Bohnke, J.; Feldpausch, M.; Gorzelniak, K.; Janke, J.; Batkai, S.; Pacher, P.; Harvey-White, J.; Luft, F.C.; Sharma, A.M.; et al. Activation of the peripheral endocannabinoid system in human obesity. Diabetes 2005, 54, 2838–2843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunos, G.; Osei-Hyiaman, D.; Liu, J.; Godlewski, G.; Batkai, S. Endocannabinoids and the control of energy homeostasis. J. Biol. Chem. 2008, 283, 33021–33025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, A.P.; Vemuri, V.K.; Peng, Y.; Wood, J.T.; Olszewska, T.; Pittman, Q.J.; Makriyannis, A.; Sharkey, K.A. A neutral CB1 receptor antagonist reduces weight gain in rat. Am. J. Physiol. Regul. Integr. Omp. Physiol. 2007, 293, R2185–R2193. [Google Scholar] [CrossRef]
- Fernandez, J.R.; Allison, D.B. Rimonabant sanofi-synthelabo. Curr. Opin. Investig. Drugs 2004, 5, 430–435. [Google Scholar]
- Sam, A.H.; Salem, V.; Ghatei, M.A. Rimonabant: From rion to ban. J. Obes. 2011, 2011, 432607. [Google Scholar] [CrossRef]
- Ward, S.J.; Raffa, R.B. Rimonabant redux and strategies to improve the future outlook of CB1 receptor neutral-antagonist/inverse-agonist therapies. Obesity 2011, 19, 1325–1334. [Google Scholar] [CrossRef]
- Jbilo, O.; Ravinet-Trillou, C.; Arnone, M.; Buisson, I.; Bribes, E.; Peleraux, A.; Penarier, G.; Soubrie, P.; Le Fur, G.; Galiegue, S.; et al. The CB1 receptor antagonist rimonabant reverses the diet-induced obesity phenotype through the regulation of lipolysis and energy balance. FASEB J. 2005, 19, 1567–1569. [Google Scholar] [CrossRef]
- Boon, M.R.; Kooijman, S.; van Dam, A.D.; Pelgrom, L.R.; Berbee, J.F.; Visseren, C.A.; van Aggele, R.C.; van den Hoek, A.M.; Sips, H.C.M.; Lombes, M.; et al. Peripheral cannabinoid 1 receptor blockade activates brown adipose tissue and diminishes dyslipidemia and obesity. FASEB J. 2014, 28, 5361–5375. [Google Scholar] [CrossRef] [Green Version]
- Herling, A.W.; Kilp, S.; Juretschke, H.P.; Neumann-Haefelin, C.; Gerl, M.; Kramer, W. Reversal of visceral adiposity in candy-diet fed female Wistar rats by the CB1 receptor antagonist rimonabant. Int. J. Obes. 2008, 32, 1363–1372. [Google Scholar] [CrossRef] [Green Version]
- Ben Ali, Y.; Chahinian, H.; Petry, S.; Müller, G.; Carriere, F.; Verger, R.; Abousalham, A. Might the kinetic behavior of hormone-sensitive lipase reflect the absence of the lid domain? Biochemistry 2004, 43, 9298–9306. [Google Scholar] [CrossRef]
- Londos, C.; Honnor, R.C.; Dhillon, G.S. cAMP-dependent protein kinase and lipolysis in rat adipocytes. III. Multiple modes of insulin regulation of lipolysis and regulation of insulin responses by adenylate cyclase regulators. J. Biol. Chem. 1985, 260, 15139–15145. [Google Scholar] [PubMed]
- Müller, G.; Schulz, A.; Wied, S.; Frick, W. Regulation of lipid raft proteins by glimepiride- and insulin-induced glycosylphosphatidylinositol-specific phospholipase C in rat adipocytes. Biochem. Pharmacol. 2005, 69, 761–780. [Google Scholar] [CrossRef] [PubMed]
- Egan, J.J.; Greenberg, A.S.; Chang, M.K.; Wek, S.A.; Moos, M.C.; Londos, C. Mechanism of hormone-stimulated lipolysis in adipocytes: Translocation of hormone-sensitive lipase to the lipid storage droplet. Proc. Natl. Acad. Sci. USA 1992, 89, 8537–8541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Londos, C.; Brasaemle, D.L.; Schultz, C.J.; Adler-Wailes, D.C.; Levin, D.M.; Kimmel, A.R.; Rondinone, C.M. On the control of lipolysis in adipocytes. Ann. N. Y. Acad. Sci. 1999, 892, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Müller, G.; Petry, S. Physiological and pharmacological regulation of triacylglycerol storage and mobilization. In Lipases and Phospholipases in Drug Development; Wiley-VCH: Weinheim, Germany, 2004; pp. 231–330. [Google Scholar]
- Clifford, G.M.; Londos, C.; Kraemer, F.B.; Vernon, R.G.; Yeaman, S.J. Translocation of hormone-sensitive lipase and perilipin upon lipolytic stimulation of rat adipocytes. J. Biol. Chem. 2000, 275, 5011–5015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bensaid, M.; Gary-Bobo, M.; Esclangon, A.; Maffrand, J.P.; Le Fur, G.; Oury-Donat, F.; Soubrie, P. The cannabinoid CB1 receptor antagonist SR141716 increases Acrp30 mRNA expression in adipose tissue of obese fa/fa rats and in cultured adipocyte cells. Mol. Pharmacol. 2003, 63, 908–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, J.; Hinden, L.; Drori, A.; Udi, S.; Azar, S.; Baraghithy, S. The therapeutic potential of targeting the peripheral endocannabinoid/CB1 receptor system. Eur. J. Intern. Med. 2018, 49, 23–29. [Google Scholar] [CrossRef]
- Howlett, A.C. Cannabinoid inhibition of adenylate cyclase. Biochemistry of the response in neuroblastoma cell membranes. Mol. Pharmacol. 1985, 27, 429–436. [Google Scholar]
- Belfrage, P.; Fredrikson, G.; Olsson, H.; Stralfors, P. Hormonal regulation of adipose tissue lipolysis by reversible phosphorylation of hormone-sensitive lipase. Prog. Clin. Biol. Res. 1982, 102, 213–223. [Google Scholar]
- Egan, J.J.; Greenberg, A.S.; Chang, M.K.; Londos, C. Control of endogenous phosphorylation of the major cAMP-dependent protein kinase substrate in adipocytes by insulin and beta-adrenergic stimulation. J. Biol. Chem. 1990, 265, 18769–18775. [Google Scholar]
- Yeaman, S.J. Hormone-sensitive lipase – new roles for an old enzyme. Biochem. J. 2004, 379, 11–22. [Google Scholar] [CrossRef]
- Okuda, H.; Morimoto, C.; Tsujita, T. Role of endogenous LD in lipolysis in rat adipocytes. J. Lipid Res. 1994, 35, 36–44. [Google Scholar]
- Morimoto, C.; Tsujita, T.; Sumida, M.; Okuda, H. Substrate-dependent lipolysis induced by isoproterenol. Biochem. Biophys. Res. Commun. 2000, 274, 631–634. [Google Scholar] [CrossRef]
- Londos, C.; Brasaemle, D.L.; Schultz, C.J.; Segrest, J.; Kimmel, A.R. Perilipins, ADRP, and other proteins that associate with intracellular neutral lipid droplets in animal cells. Sem. Cell. Dev. Biol. 1999, 10, 51–58. [Google Scholar] [CrossRef]
- Londos, C.; Sztalryd, C.; Tansey, J.T.; Kimmel, A.R. Role of PAT proteins in lipid metabolism. Biochimie 2005, 87, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Tauchi-Sato, K.; Ozeki, S.; Houjou, T.; Taguchi, R.; Fujimoto, T. The surface of lipid droplets is a phospholipid monolayer with a unique fatty acid composition. J. Biol. Chem. 2002, 277, 44507–44512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarda, L.; Desnuelle, P. Action de la lipase pancreatique sur les esters en emulsion. Biochim. Biophys. Acta 1958, 30, 513–521. [Google Scholar] [CrossRef]
- Verger, R. ‘Interfacial activation’ of lipases: Facts and artifacts. Trends Biotechnol. 1997, 15, 32–38. [Google Scholar] [CrossRef]
- Pertwee, R.G. Inverse agonism and neutral antagonism at cannabinoid CB(1) receptors. Life Sci. 2005, 76, 1307–1324. [Google Scholar] [CrossRef]
- Jarai, Z.; Wagner, J.A.; Varga, K.; Compton, D.R.; Martin, B.R.; Zimmer, A.M.; Bonner, T.I.; Buckley, N.E.; Mezey, E.; Razdan, R.K.; et al. Cannabinoid-induced mesenteric vasodilation through an endothelial site distinct from CB1 or CB2 receptors. Proc. Natl. Acad. Sci. USA 1999, 96, 14136–14141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smart, D.; Gunthorpe, M.J.; Jerman, J.C.; Nasir, S.; Gray, J.; Muir, A.I.; Chambers, J.K.; Randall, A.D.; Davis, J.B. The endogenous lipid anandamide is a full agonist at the human vanilloid receptor (hVR1). Br. J. Pharmacol. 2000, 129, 227–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Marzo, V.; De Petrocellis, L.; Fezza, F.; Ligresti, A.; Bisogno, T. Anandamide receptors. Prostaglandins Leukot. Essent. Fatty Acids 2002, 66, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Wijkander, J.; Landström, T.R.; Manganiello, V.; Belfrage, P.; Degerman, E. Insulin-induced phosphorylation and activation of phosphodiesterase 3B in rat adipocytes: Possible role of protein kinase B but not mitogen-activated protein kinase or p70 S6 kinase. Endocrinology 1998, 139, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Kola, B.; Hubina, E.; Tucci, S.A.; Kirkham, T.C.; Garcia, E.A.; Mitchell, S.E.; Williams, L.M.; Hawley, S.A.; Hardie, D.G.; Grossman, A.B.; et al. Cannabinoids and ghrelin have both central and peripheral metabolic and cardiac effects via AMP-activated protein kinase. J. Biol. Chem. 2005, 280, 25196–25201. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, J.E.; Brocklehurst, K.J.; Marley, A.E.; Carey, F.; Carling, D.; Beri, R.K. Inhibition of lipolysis and lipogenesis in isolated rat adipocytes with AICAR, a cell-permeable activator of AMP-activated protein kinase. FEBS Lett. 1996, 382, 43–47. [Google Scholar] [CrossRef] [Green Version]
- Daval, M.; Diot-Duputy, F.; Bazin, R.; Hainault, I.; Viollet, B.; Vaulont, S.; Hajduch, E.; Ferre, P.; Foufelle, F. Anti-lipolytic action of cAMP-activated protein kinase in rodent adipocytes. J. Biol. Chem. 2005, 280, 25250–25257. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.A. Lipid droplets: Proteins floating on a pool of fat. Curr. Biol. 2001, 11, R446–R449. [Google Scholar] [CrossRef] [Green Version]
- Blanchette-Mackie, E.J.; Dwyer, N.K.; Barber, T.; Coxey, R.A.; Takeda, T.; Rondinone, C.M.; Theodorakis, J.L.; Greenberg, A.S.; Londos, C. Perilipin is located on the surface layer of intracellular lipid droplets in adipocytes. J. Lipid Res. 1995, 36, 1211–1226. [Google Scholar]
- Winkler, F.K.; D’Arcy, A.; Hunziker, W. Structure of human pancreatic lipase. Nature 1995, 343, 771–774. [Google Scholar] [CrossRef]
- Brady, L.; Brzozowski, A.M.; Derewenda, Z.S.; Dodson, E.; Dodson, G.; Tolley, S.; Turkenburg, J.P.; Christiansen, L.; Huge-Jensen, B.; Norskov, L. A serine protease triad forms the catalytic centre of a triacylglycerol lipase. Nature 1990, 343, 767–770. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.T.; Soulages, J.L.; Hariharasundaram, B.; Arrese, E.L. Activation of the lipid droplet controls the rate of lipolysis of triglycerides in the insect fat body. J. Biol. Chem. 2005, 280, 22624–22631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravinet, T.C.; Delgorge, C.; Menet, C.; Arnone, M.; Soubrie, P. CB1 cannabinoid receptor knockout in mice leads to leanness, resistance to diet-induced obesity and enhanced leptin sensitivity. Int. J. Obes. Relat. Metab. Disord. 2004, 28, 640–648. [Google Scholar] [CrossRef] [Green Version]
- Han, J.H.; Shin, H.; Park, J.Y.; Rho, J.G.; Son, D.H.; Kim, K.W.; Seong, J.K.; Yoon, S.H.; Kim, W. A novel peripheral cannabinoid 1 receptor antagonist, AJ5012, improves metabolic outcomes and suppresses adipose tissue inflammation in obese mice. FASEB J. 2019, 33, 4314–4326. [Google Scholar] [CrossRef]
- Mollica, A.; Pelliccia, S.; Famiglini, V.; Stefanucci, A.; Macedonio, G.; Chiavaroli, A.; Orlando, G.; Brunetti, L.; Ferrante, C.; Pieretti, S.; et al. Exploring the first rimonabant analog-opoid peptide hybrid compound, as bivalent ligand for CB1 and opoid receptors. J. Enzyme Inhib. Med. Chem. 2017, 32, 444–451. [Google Scholar] [CrossRef] [Green Version]
- Stanucci, A.; Macedonio, G.; Dvoracsko, S.; Tömböly, C.; Mollica, A. Novel fubinaca/rimonabant hybrids as endocannabinoid system modulators. Amino Acids 2018, 50, 1595–1605. [Google Scholar] [CrossRef] [PubMed]
- Dimmito, M.P.; Stefanucci, A.; Pieretti, S.; Minosi, P.; Dvoracsko, S.; Tömböly, C.; Zengin, G.; Mollica, A. Discovery of orexant and anorexant agents with indazole scaffold endowed with peripheral antiedema activity. Biomolecules 2019, 9, 492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, G.; Wied, S. The sulfonylurea drug, glimepiride, stimulates glucose transport, glucose transporter translocation, and dephosphorylation in insulin-resistant rat adipocytes in vitro. Diabetes 1993, 42, 1852–1867. [Google Scholar] [CrossRef] [Green Version]
- Müller, G.; Jordan, H.; Jung, C.; Kleine, H.; Petry, S. Analysis of lipolysis in adipocytes using a fluorescent fatty acid derivative. Biochimie 2003, 85, 1245–1256. [Google Scholar] [CrossRef]
- Müller, G.; Jordan, H.; Petry, S.; Wetekam, E.-M.; Schindler, P. Analysis of lipid metabolism in adipocytes using fluorescent fatty acids I. Insulin stimulation of lipogenesis. Biochim. Biophys. Acta 1997, 1347, 23–39. [Google Scholar] [CrossRef]
- Greenberg, A.S.; Egan, J.J.; Wek, S.A.; Garty, N.B.; Blanchette-Mackie, E.J.; Londos, C. Perilipin, a major hormonally regulated adipocyte-specific phosphoprotein associated with the periphery of lipid storage droplets. J. Biol. Chem. 1991, 266, 11341–11346. [Google Scholar] [PubMed]
- Wolins, N.E.; Quaynor, B.K.; Skinner, J.R.; Schoenfish, M.J.; Tzekov, A.; Bickel, P.E. S3–12, Adipophilin, and TIP47 package lipid in adipocytes. J. Biol. Chem. 2005, 280, 19146–19155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holm, C.; Belfrage, P.; Fredrikson, G. Human adipose tissue hormone-sensitive lipase: Identification and comparison with other species. Biochim. Biophys. Acta 1989, 1006, 193–197. [Google Scholar] [CrossRef]
- Müller, G.; Jung, C.; Wied, S.; Welte, S.; Frick, W. Insulin-mimetic signaling by the sulfonylurea glimepiride and phosphoinositolglycans involves distinct mechanisms for redistribution of lipid raft components. Biochemistry 2001, 40, 14603–14620. [Google Scholar] [CrossRef] [PubMed]
- Petry, S.; Ali, Y.B.; Chahinian, H.; Jordan, H.; Kleine, H.; Müller, G.; Carriere, F.; Abousalham, A. Sensitive assay for hormone-sensitive lipase using NBD-labeled monoacylglycerol to detect low activities in rat adipocytes. J. Lipid Res. 2005, 46, 603–614. [Google Scholar] [CrossRef] [Green Version]
- Marzo, A.; Ghirardi, P.; Sardini, D.; Meroni, G. Simplified measurement of monoglycerides, diglycerides, triglycerides, and free fatty acids in biological samples. Clin. Chem. 1971, 17, 145–147. [Google Scholar] [CrossRef]
- Murphy, E.J.; Stiles, T.; Schroeder, F. Sterol carrier protein-2 expression alters phospholipid content and fatty acyl composition in L-cell fibroblasts. J. Lipid Res. 2000, 41, 788–796. [Google Scholar]
- Schölch, C.; Kuhlmann, J.; Gossel, M.; Müller, G.; Neumann-Häfelin, A.; Belz, U.; Kalisch, J.; Biemer-Daub, G.; Kramer, W.; Juretschke, H.-P.; et al. Characterization of adenosine-A1-receptor-mediated antilipolysis in rats by tissue-microdialysis, 1H-spectroscopy and glucose clamp studies. Diabetes 2004, 53, 1920–1926. [Google Scholar] [CrossRef]
- Rinaldi-Carmona, M.; Barth, F.; Heaulme, M.; Alonso, R.; Shire, D.; Congy, C.; Soubrie, P.; Breliere, J.C.; Le Fur, G. Biochemical and pharmacological characterization of SR141716, the first potent and selective brain cannabinoid receptor antagonist. Life Sci. 1995, 56, 1941–1947. [Google Scholar] [CrossRef]
- Müller, G.; Wied, S.; Wetekam, E.-M.; Crecelius, A.; Unkelbach, A.; Pünter, J. Stimulation of glucose utilization in 3T3 adipocytes and rat diaphragm in vitro by the sulfonylureas, glimepiride and glibenclamide, is correlated with modulations of the cAMP regulatory cascade. Biochem. Pharmacol. 1994, 48, 985–996. [Google Scholar] [CrossRef] [Green Version]
- Vertesy, L.; Beck, B.; Brönstrup, M.; Ehrlich, K.; Kurz, M.; Müller, G.; Schummer, D.; Seibert, G. Cyclipostins, novel hormone-sensitive lipase inhibitors from Streptomyces sp. DSM 13381. II. Isolation, structure elucidation and biological properties. J. Antibiotics 1992, 55, 480–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of the compounds Rimonabant, AM251, AM281, AM630 and NBD-FA are available from the authors. |











© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Müller, G.A.; Herling, A.W.; Wied, S.; Müller, T.D. CB1 Receptor-Dependent and Independent Induction of Lipolysis in Primary Rat Adipocytes by the Inverse Agonist Rimonabant (SR141716A). Molecules 2020, 25, 896. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25040896
Müller GA, Herling AW, Wied S, Müller TD. CB1 Receptor-Dependent and Independent Induction of Lipolysis in Primary Rat Adipocytes by the Inverse Agonist Rimonabant (SR141716A). Molecules. 2020; 25(4):896. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25040896
Chicago/Turabian StyleMüller, Günter A., Andreas W. Herling, Susanne Wied, and Timo D. Müller. 2020. "CB1 Receptor-Dependent and Independent Induction of Lipolysis in Primary Rat Adipocytes by the Inverse Agonist Rimonabant (SR141716A)" Molecules 25, no. 4: 896. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25040896