
The Influence of 5′R and 5′S cdA and cdG on the Activity of BsmAI and SspI Restriction Enzymes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
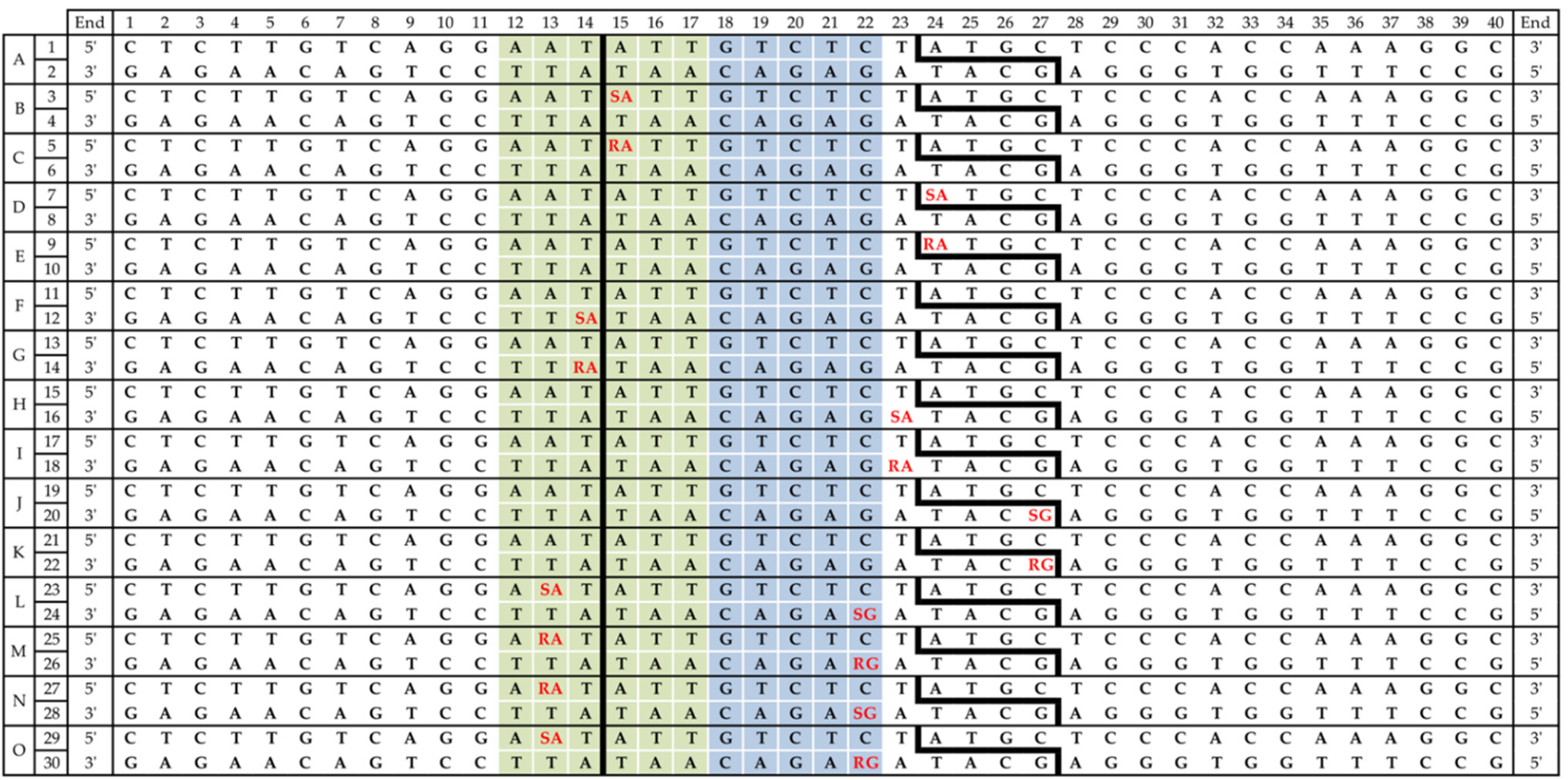
2.1. Substrate Oligonucleotide Synthesis and Purification
2.2. Mass Spectroscopy of Oligonucleotides
2.3. Preparation of 5′-End-Labeled Oligonucleotides
2.4. Oligonucleotide Hybridization
2.5. BsmAI and SspI Cleavage Assay
2.6. PAGE Electrophoresis
2.7. Data Presentation
3. Results and Discussion
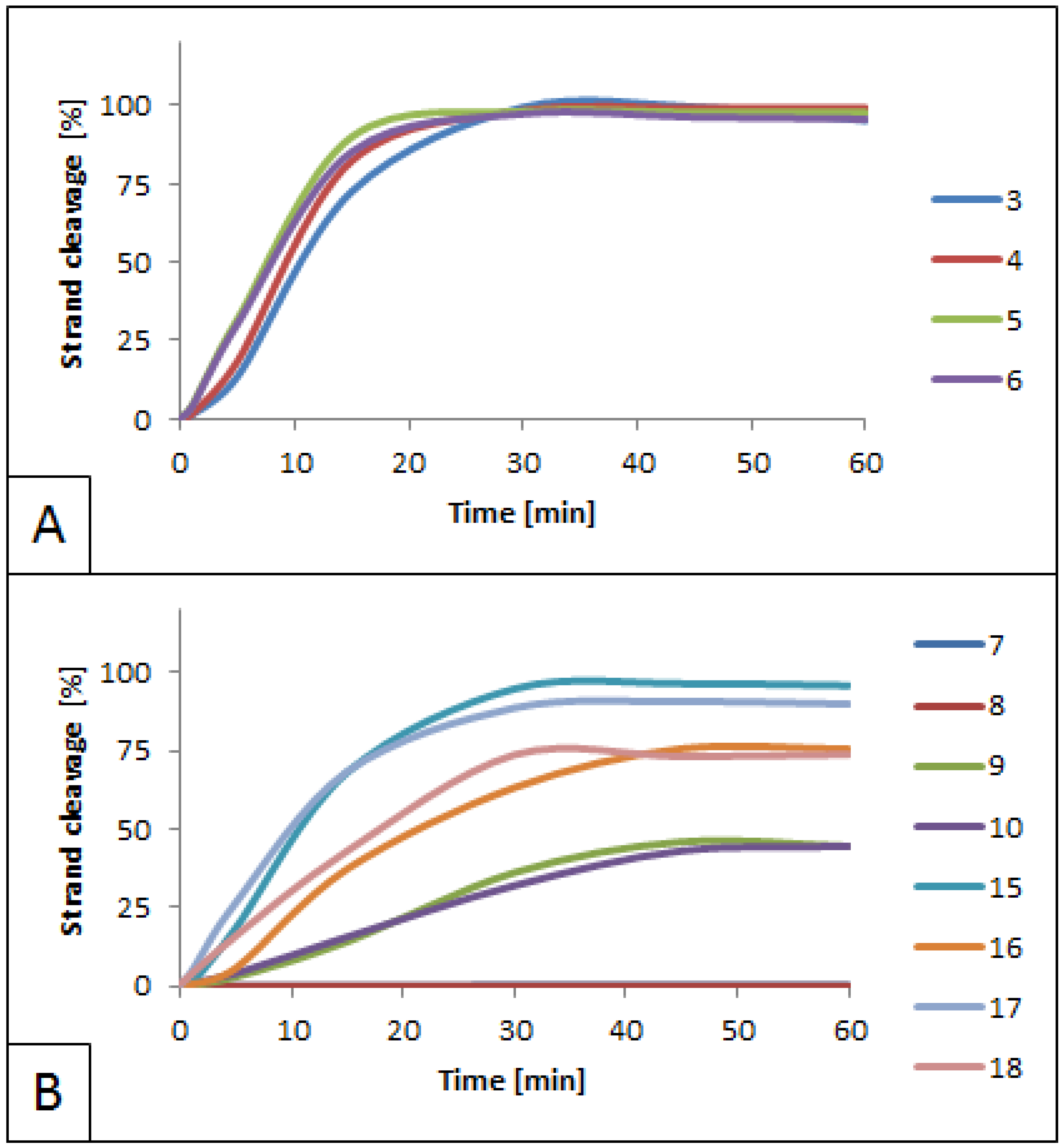
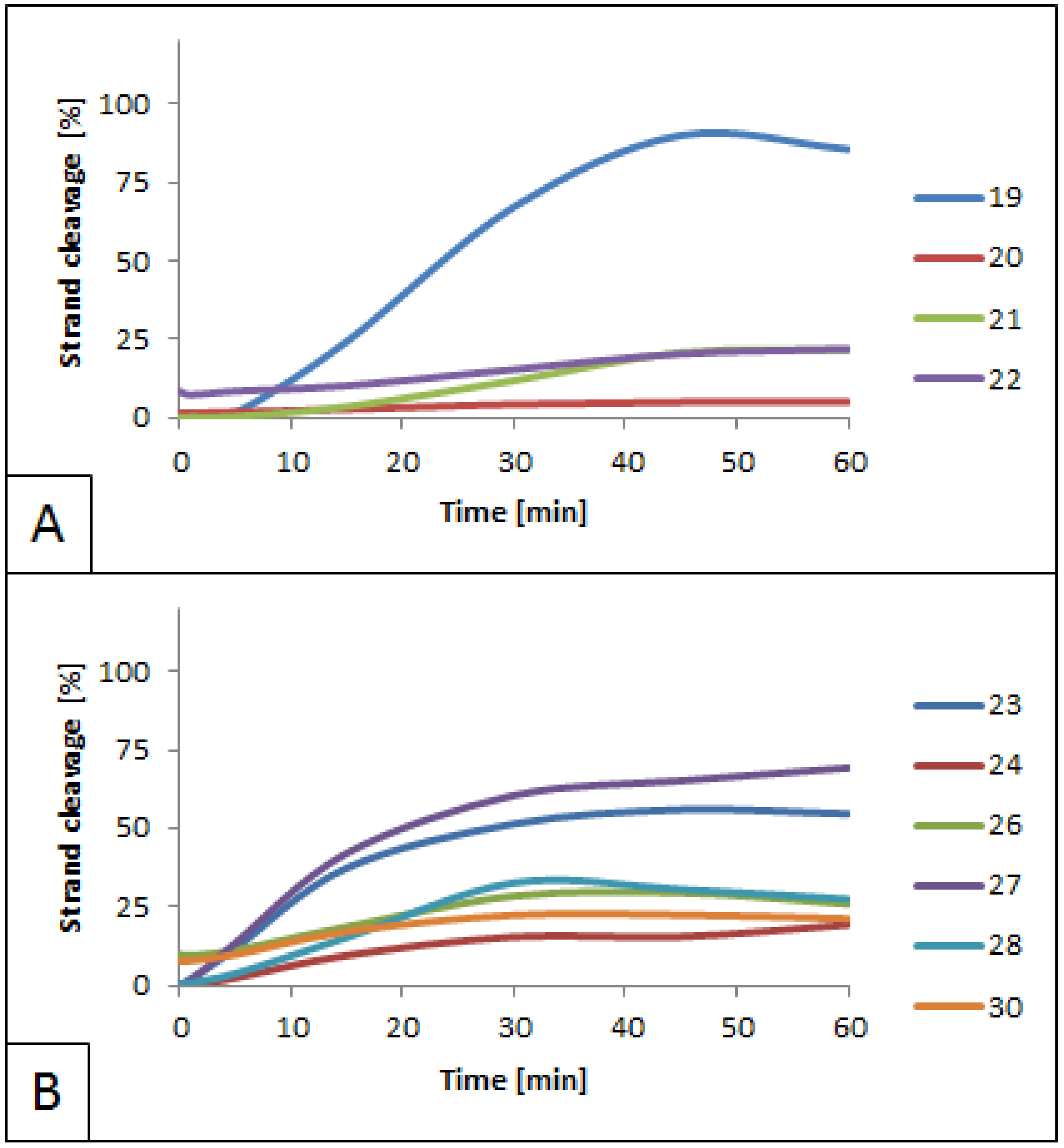
3.1. Influence of 5′S and 5′R cdPus on SspI Activity
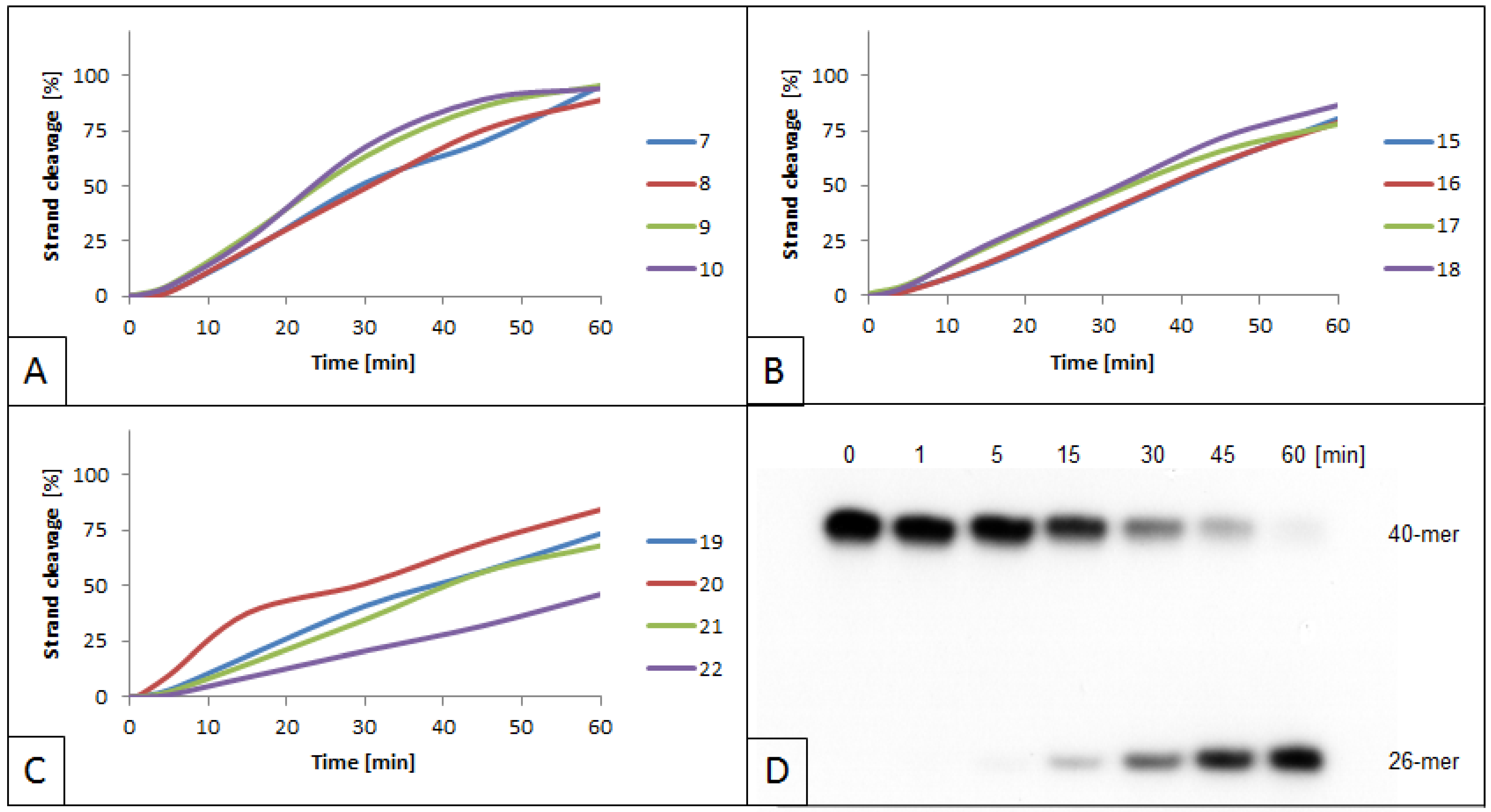
3.2. Influence of 5′S and 5′R cdA on BsmAI activity
3.3. Influence of 5′S and 5′R cdG on BsmAI Activity
4. Conclusions
- The activity of SspI was blocked by cdPus located within the enzyme’s recognition site.
- The inhibition of SspI by lesions placed outside the enzyme’s recognition site was inversely proportional to the distance between the lesions and the recognition site.
- The BsmAI activity was similar for all cdPus placed outside the enzyme’s recognition and incision sites—in most cases, the enzyme’s activity was slightly lower or similar to that in the control.
- The inhibition of BsmAI was lower for RcdA than for ScdA when lesions were located adjacent to the incision site of the enzyme—the results were similar for lesions located in different positions (position 24 toward the 3′-end of the leading strand and position 23 toward the 5′-end of the complementary strand).
- The presence of cdG (both diastereomers) next to the incision site of BsmAI lowered its activity more than cdA (both diastereomers).
- CdG showed a lower impact on the activity of BsmAI when it was located within the recognition site than next to the incision site.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Lindahl, T. Instability and decay of the primary structure of DNA. Nat. Cell Biol. 1993, 362, 709–715. [Google Scholar] [CrossRef]
- Ambekar, S.S. DNA: Damage and repair mechanisms in humans. Glob. J. Pharm. Pharm. Sci. 2017, 3, 555613. [Google Scholar] [CrossRef] [Green Version]
- Kuraoka, I.; Bender, C.; Romieu, A.; Cadet, J.; Wood, R.D.; Lindahl, T. Removal of oxygen free-radical-induced 5’,8-purine cyclodeoxynucleosides from DNA by the nucleotide excision-repair pathway in human cells. Proc. Natl. Acad. Sci. USA 2000, 97, 3832–3837. [Google Scholar] [CrossRef] [Green Version]
- Walmacq, C.; Wang, L.; Chong, J.; Scibelli, K.; Lubkowska, L.; Gnatt, A.; Brooks, P.J.; Wang, D.; Kashlev, M. Mechanism of RNA polymerase II bypass of oxidative cyclopurine DNA lesions. Proc. Natl. Acad. Sci. USA 2015, 112, E410–E419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatgilialoglu, C.; Ferreri, C.; Geacintov, N.E.; Krokidis, M.G.; Liu, Y.; Masi, A.; Shafirovich, V.; Terzidis, M.A.; Tsegay, P.S. 5′,8-Cyclopurine lesions in DNA damage: Chemical, analytical, biological, and diagnostic significance. Cells 2019, 8, 513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, C.; Dai, X.; Yuan, B.-F.; Wang, J.; Wang, J.; Brooks, P.J.; Niedernhofer, L.J.; Wang, Y. A quantitative assay for assessing the effects of DNA lesions on transcription. Nat. Chem. Biol. 2012, 8, 817–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaruga, P.; Dizdaroglu, M. 8,5′-Cyclopurine-2′-deoxynucleosides in DNA: Mechanisms of formation, measurement, repair and biological effects. DNA Repair 2008, 7, 1413–1425. [Google Scholar] [CrossRef]
- Brooks, P.J. The cyclopurine deoxynucleosides: DNA repair, biological effects, mechanistic insights, and unanswered ques-tions. Free Radic. Biol. Med. 2017, 107, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Suhasini, A.N.; Banerjee, T.; Sommers, J.A.; Kaplan, D.L.; Kuper, J.; Kisker, C.; Brosh, R.M. Impact of age-associated cyclopurine lesions on DNA repair helicases. PLoS ONE 2014, 9, e113293. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Das, R.S.; Basu, A.K.; Stone, M.P. Structures of (5′S)-8,5′-Cyclo-2′-deoxyguanosine mismatched with dA or dT. Chem. Res. Toxicol. 2012, 25, 478–490. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.M.; Jaruga, P.; Ramsey, C.R.; Gilman, N.K.; Green, V.M.; Rostad, S.W.; Emerman, J.T.; Dizdaroglu, M.; Malins, D.C. Structural alterations in breast stromal and epithelial DNA: The Influence of 8,5-cyclo-2-Deoxyadenosine. Cell Cycle 2006, 5, 1240–1244. [Google Scholar] [CrossRef] [Green Version]
- Jaruga, P.; Rozalski, R.; Jawien, A.; Migdalski, A.; Olinski, R.; Dizdaroglu, M. DNA damage products (5′R)- and (5′S)-8,5′-Cyclo-2′- deoxyadenosines as potential biomarkers in human urine for atherosclerosis. Biochemistry 2012, 51, 1822–1824. [Google Scholar] [CrossRef] [PubMed]
- Karwowski, B.T. The influence of (5’R)- and (5’S)-5’,8-Cyclo-2’-Deoxyadenosine on UDG and hAPE1 activity. Tandem lesions are the base excision repair system’s nightmare. Cells 2019, 8, 1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boguszewska, K.; Szewczuk, M.; Kaźmierczak-Barańska, J.; Karwowski, B.T. How (5’S) and (5’R) 5’,8-Cyclo-2’-Deoxypurines affect base excision repair of clustered DNA damage in nuclear extracts of xrs5 cells? A bio-chemical study. Cells 2021, 10, 725. [Google Scholar] [CrossRef]
- Roberts, R.J.; Vincze, T.; Posfai, J.; Macelis, D. REBASE—Enzymes and genes for DNA restriction and modification. Nucleic Acids Res. 2007, 35, D269–D270. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.J.; Vincze, T.; Posfai, J.; Macelis, D. REBASE—A database for DNA restriction and modification: Enzymes, genes and genomes. Nucleic Acids Res. 2009, 38, D234–D236. [Google Scholar] [CrossRef] [PubMed]
- Bourniquel, A.; Bickle, T. Complex restriction enzymes: NTP-driven molecular motors. Biochimie 2002, 84, 1047–1059. [Google Scholar] [CrossRef]
- Chan, S.H.; Stoddard, B.L.; Xu, S.Y. Natural and engineered nicking endonucleases—From cleavage mechanism to engineering of strand-specificity. Nucleic Acids Res. 2011, 39, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Capalash, N.; Sharma, P. Restriction endonucleases: Natural and directed evolution. Appl. Microbiol. Biotechnol. 2012, 94, 583–599. [Google Scholar] [CrossRef]
- Chinen, A.; Naito, Y.; Handa, N.; Kobayashi, I. Evolution of sequence recognition by restriction-modification enzymes: Selective pressure for specificity decrease. Mol. Biol. Evol. 2001, 18, 276–277. [Google Scholar] [CrossRef] [Green Version]
- Karwowski, B.T.; Bellon, S.; O’Neill, P.; Lomax, M.E.; Cadet, J. Effects of (5′S)-5′,8-cyclo-2′-deoxyadenosine on the base excision repair of oxidatively generated clustered DNA damage. A biochemical and theoretical study. Org. Biomol. Chem. 2014, 12, 8671–8682. [Google Scholar] [CrossRef] [PubMed]
- Zahoor, M.; Abubakar, M.; Naim, S.; Arshed, Q. Molecular typing of field isolates from two outbreaks of infectious bursal disease virus from Pakistan. Veter. World 2011, 4, 297–300. [Google Scholar] [CrossRef]
- Stuchbury, G.; Münch, G. Optimizing the generation of stable neuronal cell lines via pre-transfection restriction enzyme digestion of plasmid DNA. Cytotechnology 2010, 62, 189–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hesson, J.C.; Lundström, J.O.; Halvarsson, P.; Erixon, P.; Collado, A. A sensitive and reliable restriction enzyme assay to distinguish between the mosquitoes Culex torrentium and Culex pipiens. Med. Veter. Èntomol. 2010, 24, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Morgan, R.D.; Chen, Z. A new type II restriction endonuclease, BsmA I, from Bacillus stearothermophilus. Nucleic Acids Res. 1990, 18, 686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Samuelson, J.C.; Zhou, J.; Dore, A.; Xu, S.-Y. Engineering strand-specific DNA nicking enzymes from the type IIS restriction endonucleases BsaI, BsmBI, and BsmAI. J. Mol. Biol. 2004, 337, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.J. How restriction enzymes became the workhorses of molecular biology. Proc. Natl. Acad. Sci. USA 2005, 102, 5905–5908. [Google Scholar] [CrossRef] [Green Version]
- Loenen, W.A.M.; Dryden, D.T.F.; Raleigh, E.A.; Wilson, G.G.; Murray, N.E. Highlights of the DNA cutters: A short history of the restriction enzymes. Nucleic Acids Res. 2014, 42, 3–19. [Google Scholar] [CrossRef] [Green Version]
- Tzfira, T.; Weinthal, D.; Marton, I.; Zeevi, V.; Zuker, A.; Vainstein, A. Genome modifications in plant cells by custom-made restriction enzymes. Plant Biotechnol. J. 2012, 10, 373–389. [Google Scholar] [CrossRef]
- Romieu, A.; Gasparutto, D.; Cadet, J. Synthesis and characterization of oligonucleotides containing 5’,8- cyclopurine 2’-deoxyribonucleosides: (5’R)-5’,8-cyclo-2’-deoxyadenosine, (5’S)-5’,8-cyclo-2’-deoxyguanosine, and (5’R)-5’,8-cyclo-2’-deoxyguanosine. Chem. Res. Toxicol. 1999, 12, 412–421. [Google Scholar] [CrossRef]
- Pande, P.; Das, R.S.; Sheppard, C.Y.; Kow, W.; Basu, A.K. Repair efficiency of (5’S)-8,5’-cyclo-2’-deoxyguanosine and (5’S)-8,5’-cyclo-2’-deoxyadenosine depends on the complementary base. DNA Repair 2011, 11, 926–931. [Google Scholar] [CrossRef] [Green Version]
- Sage, E.; Harrison, L. Clustered DNA lesion repair in eukaryotes: Relevance to mutagenesis and cell survival. Mutat. Res. Mol. Mech. Mutagen. 2011, 711, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, P. The case for 8,5′-cyclopurine-2′-deoxynucleosides as endogenous DNA lesions that cause neurodegeneration in xeroderma pigmentosum. Neuroscience 2007, 145, 1407–1417. [Google Scholar] [CrossRef] [Green Version]
- Eccles, L.J.; Lomax, M.E.; O’Neill, P. Hierarchy of lesion processing governs the repair, double-strand break formation and mutability of three-lesion clustered DNA damage. Nucleic Acids Res. 2009, 38, 1123–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, L.; Brame, K.L.; Geltz, L.E.; Landry, A.M. Closely opposed apurinic/apyrimidinic sites are converted to double strand breaks in Escherichia coli even in the absence of exonuclease III, endonuclease IV, nucleotide excision repair and AP lyase cleavage. DNA Repair 2006, 5, 324–335. [Google Scholar] [CrossRef] [Green Version]
- Jasti, V.P.; Das, R.; Hilton, B.A.; Weerasooriya, S.; Zou, Y.; Basu, A.K. (5′S)-8,5′-Cyclo-2′-deoxyguanosine is a strong block to replication, a potent pol V-dependent mutagenic lesion, and is inefficiently repaired in Escherichia coli. Biochemistry 2011, 50, 3862–3865. [Google Scholar] [CrossRef] [PubMed]
- Pednekar, V.; Weerasooriya, S.; Jasti, V.P.; Basu, A.K. Mutagenicity and genotoxicity of (5′S)-8,5′-Cyclo-2′-deoxyadenosine in Escherichia coli and replication of (5′S)-8,5′-Cyclopurine-2′-deoxynucleosides in vitro by DNA polymerase IV, exo-free Klenow fragment, and Dpo4. Chem. Res. Toxicol. 2013, 27, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Tock, M.R.; Dryden, D.T. The biology of restriction and anti-restriction. Curr. Opin. Microbiol. 2005, 8, 466–472. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szewczuk, M.; Boguszewska, K.; Kaźmierczak-Barańska, J.; Karwowski, B.T. The Influence of 5′R and 5′S cdA and cdG on the Activity of BsmAI and SspI Restriction Enzymes. Molecules 2021, 26, 3750. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26123750
Szewczuk M, Boguszewska K, Kaźmierczak-Barańska J, Karwowski BT. The Influence of 5′R and 5′S cdA and cdG on the Activity of BsmAI and SspI Restriction Enzymes. Molecules. 2021; 26(12):3750. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26123750
Chicago/Turabian StyleSzewczuk, Michał, Karolina Boguszewska, Julia Kaźmierczak-Barańska, and Bolesław T. Karwowski. 2021. "The Influence of 5′R and 5′S cdA and cdG on the Activity of BsmAI and SspI Restriction Enzymes" Molecules 26, no. 12: 3750. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26123750