
Molecular Human Targets of Bioactive Alkaloid-Type Compounds from Tabernaemontana cymose Jacq.

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
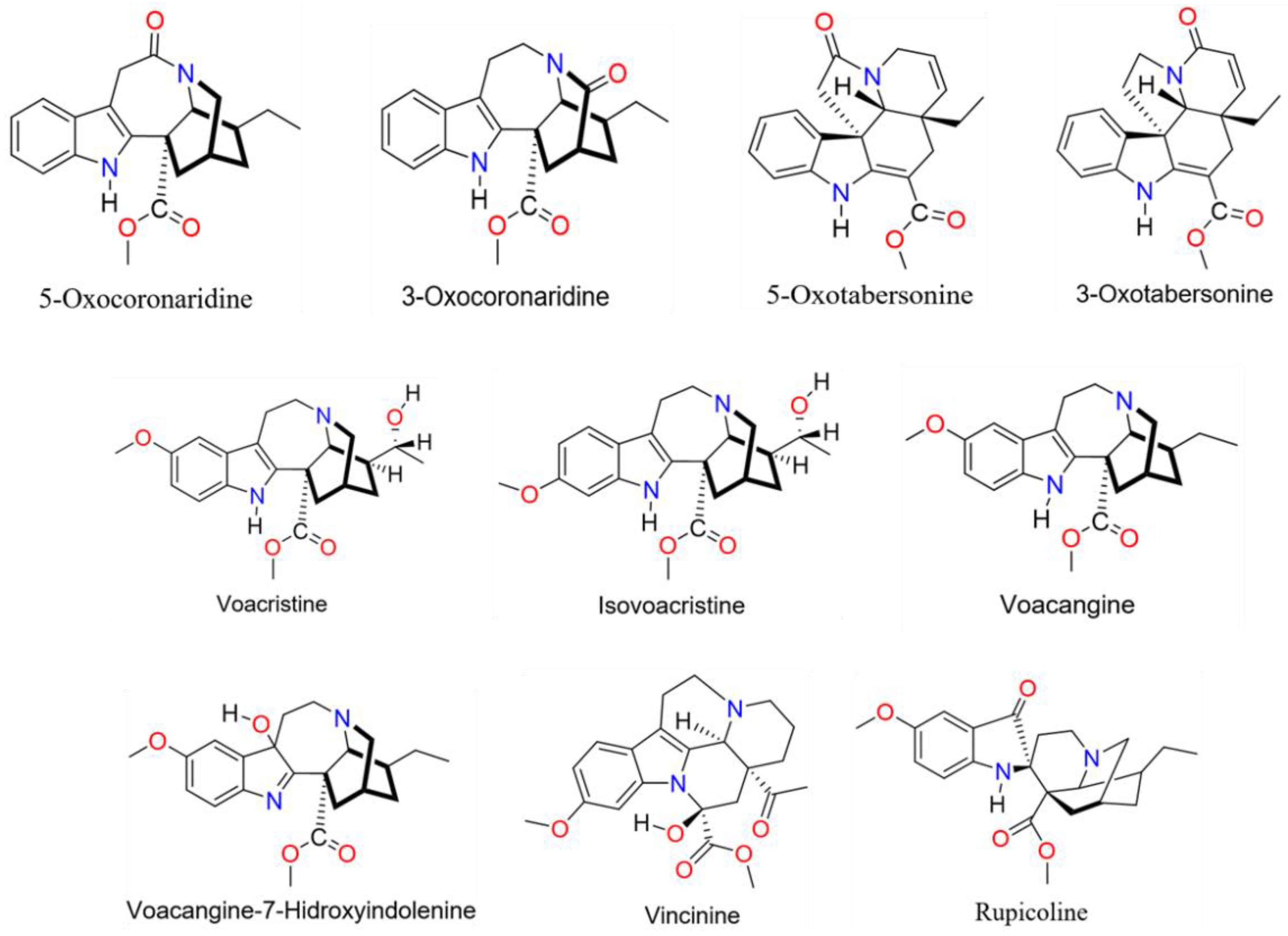
2.1. Plant Material and Compound Identification
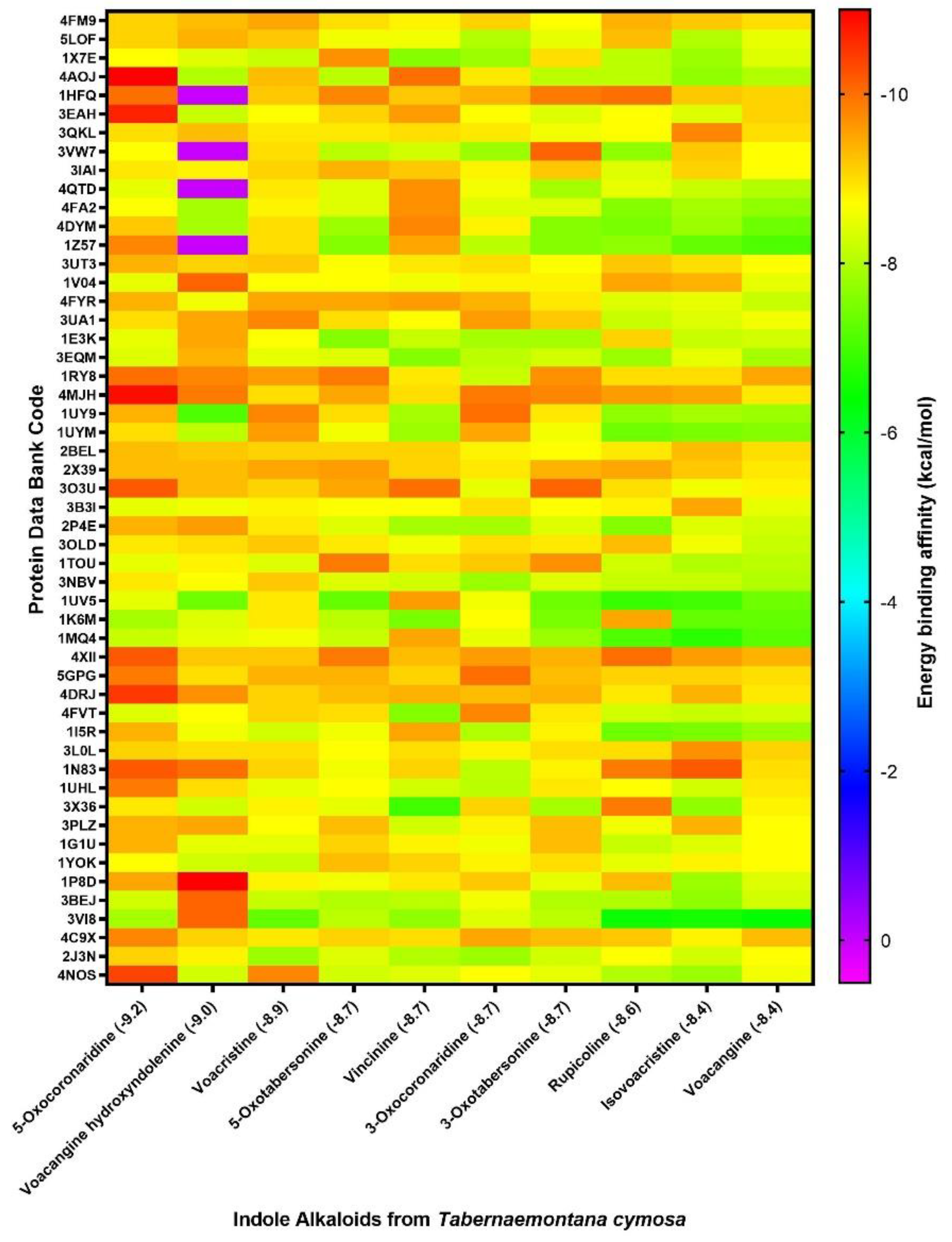
2.2. Virtual Screenig for Identified Alkaloids
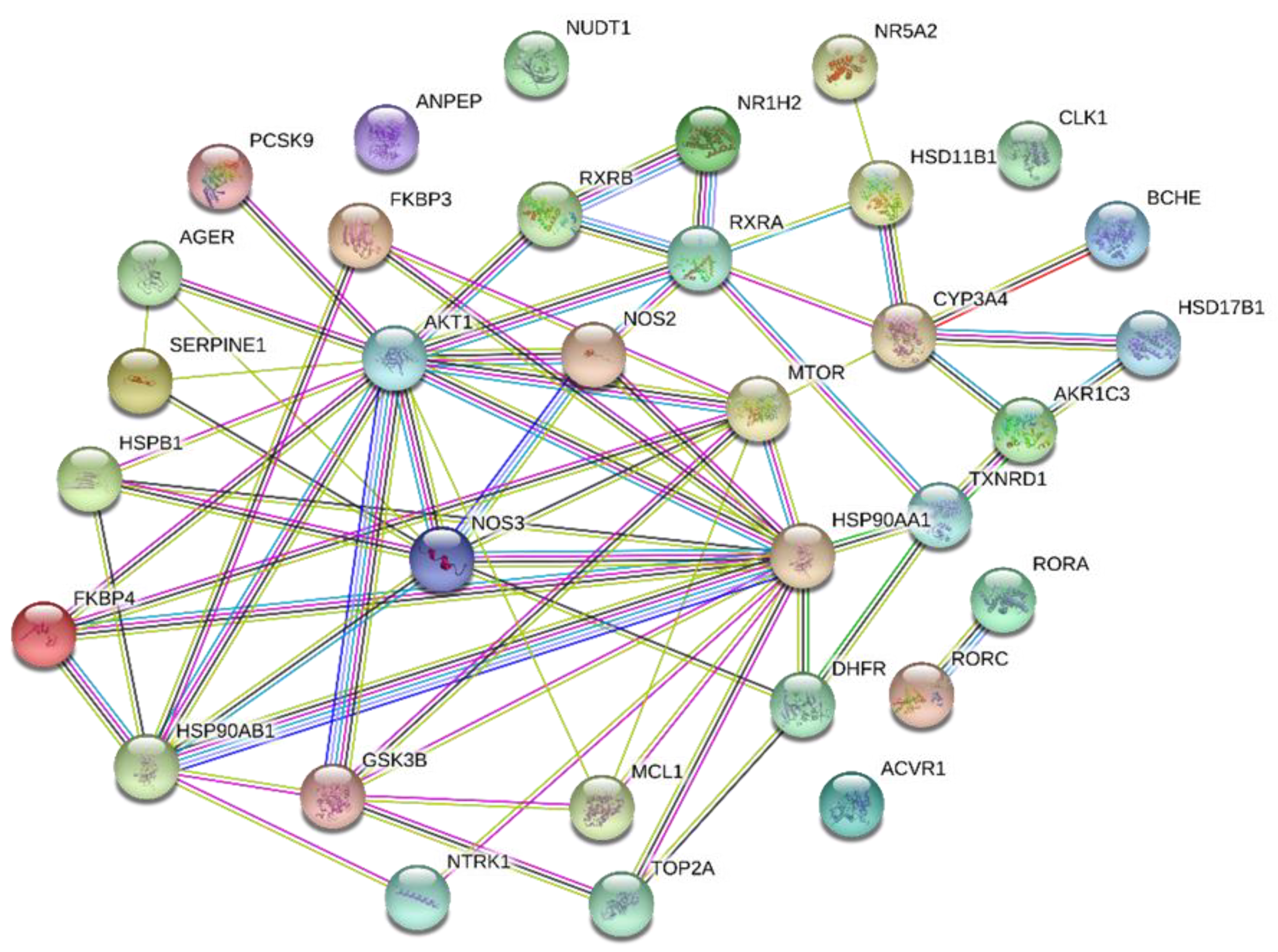
2.3. Protein–Protein Interactions (PPIs)
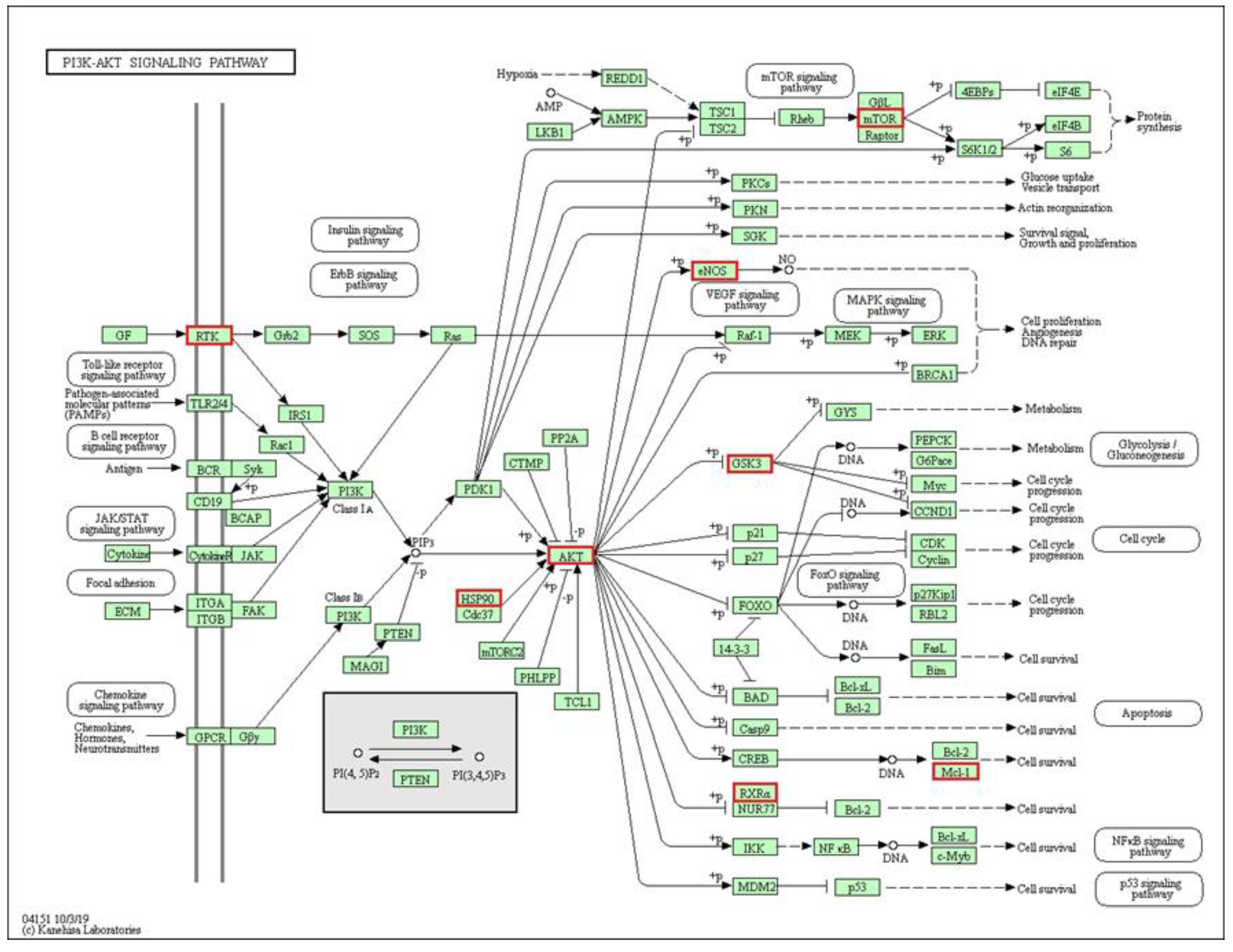
2.4. Genomic Association
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Establishment of the Database and Protein Optimization
4.3. Indole Alkaloids Identification and Ligand Preparation
4.4. Virtual Screening
4.5. Protein–Protein Interactions (PPIs)
4.6. Genomic Association
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Gillison, A.N. Plant Functional Indicators of Vegetation Response to Climate Change, Past Present and Future: I. Trends, Emerging Hypotheses and Plant Functional Modality. Flora 2019, 254, 12–30. [Google Scholar] [CrossRef]
- Sá, A.G.A.; Moreno, Y.M.F.; Carciofi, B.A.M. Plant Proteins as High-Quality Nutritional Source for Human Diet. Trends Food Sci. Technol. 2020, 97, 170–184. [Google Scholar] [CrossRef]
- Thakur, M.; Bhattacharya, S.; Khosla, P.K.; Puri, S. Improving Production of Plant Secondary Metabolites through Biotic and Abiotic Elicitation. J. Appl. Res. Med. Aromat. Plants 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Zaynab, M.; Fatima, M.; Abbas, S.; Sharif, Y.; Umair, M.; Zafar, M.H.; Bahadar, K. Role of Secondary Metabolites in Plant Defense against Pathogens. Microb. Pathog. 2018, 124, 198–202. [Google Scholar] [CrossRef]
- Chakraborty, P. Herbal Genomics as Tools for Dissecting New Metabolic Pathways of Unexplored Medicinal Plants and Drug Discovery. Biochim. Open 2018, 6, 9–16. [Google Scholar] [CrossRef]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernal, R.; Gradstein, S.; Celis, M. Catálogo de Plantas y Líquenes de Colombia; Universidad Nacional de Colombia: Bogotá, Colombia, 2016; ISBN 978-958-775-726-2. [Google Scholar]
- Gómez-Calderón, C.; Mesa-Castro, C.; Robledo, S.; Gómez, S.; Bolivar-Avila, S.; Diaz-Castillo, F.; Martínez-Gutierrez, M. Antiviral Effect of Compounds Derived from the Seeds of Mammea Americana and Tabernaemontana Cymosa on Dengue and Chikungunya Virus Infections. BMC Complement. Altern. Med. 2017, 17, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Caballero, K.; Pino-Benitez, N.; Pajaro, N.; Stashenko, E.; Olivero-Verbel, J. Plants Cultivated in Choco, Colombia, as Source of Repellents against Tribolium Castaneum (Herbst). J. Asia Pac. Entomol. 2014, 17, 753–759. [Google Scholar] [CrossRef]
- Stashenko, E.; Martínez, J.R. Study of Essential Oils Obtained from Tropical Plants Grown in Colombia. Essent. Oils Oils Nat. 2019. [Google Scholar] [CrossRef] [Green Version]
- Pabón, L.C.; Rodríguez, M.F.; Hernández-Rodríguez, P. Plantas medicinales que se comercializan en Bogotá (Colombia) para el tratamiento de enfermedades infecciosas. Bol. Latinoam. Y Caribe Plantas Med. Y Aromát. 2017, 16, 529–546. [Google Scholar]
- Majolo, F.; de Oliveira Becker Delwing, L.K.; Marmitt, D.J.; Bustamante-Filho, I.C.; Goettert, M.I. Medicinal Plants and Bioactive Natural Compounds for Cancer Treatment: Important Advances for Drug Discovery. Phytochem. Lett. 2019, 31, 196–207. [Google Scholar] [CrossRef]
- Srivastav, V.K.; Egbuna, C.; Tiwari, M. Chapter 1—Plant secondary metabolites as lead compounds for the production of potent drugs. In Phytochemicals as Lead Compounds for New Drug Discovery; Egbuna, C., Kumar, S., Ifemeje, J.C., Ezzat, S.M., Kaliyaperumal, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 3–14. ISBN 978-0-12-817890-4. [Google Scholar]
- Lee, S.-H.; Kim, J.-H. Kinetic and Thermodynamic Characteristics of Microwave-Assisted Extraction for the Recovery of Paclitaxel from Taxus Chinensis. Process Biochem. 2019, 76, 187–193. [Google Scholar] [CrossRef]
- Katiyar, C.; Gupta, A.; Kanjilal, S.; Katiyar, S. Drug Discovery from Plant Sources: An Integrated Approach. Ayu 2012, 33, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Mohs, R.C.; Greig, N.H. Drug Discovery and Development: Role of Basic Biological Research. Alzheimers Dement Transl. Res. Clin. Interv. 2017, 3, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Balunas, M.J.; Kinghorn, A.D. Drug Discovery from Medicinal Plants. Life Sci. 2005, 78, 431–441. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, J.; Pang, Z. Recent Insights for the Emerging COVID-19: Drug Discovery, Therapeutic Options and Vaccine Development. Asian J. Pharm. Sci. 2021, 16, 4–23. [Google Scholar] [CrossRef] [PubMed]
- WHO Coronavirus Disease (COVID-19) Dashboard. Available online: https://covid19.who.int (accessed on 9 February 2021).
- Ambrose, P.G. Antibacterial Drug Development Program Successes and Failures: A Pharmacometric Explanation. Curr. Opin. Pharmacol. 2017, 36, 1–7. [Google Scholar] [CrossRef]
- Lim, S.P. Dengue Drug Discovery: Progress, Challenges and Outlook. Antivir. Res. 2019, 163, 156–178. [Google Scholar] [CrossRef]
- Najafi, A.; Mahdian, N.; Yousefi, B.; Karbalaei, M.; Kermanjani, A.; Ezatpour, B.; Eslami, M. New Vaccine Candidates as a Scientific Solution against the Dream of Tuberculosis Vaccine. Rev. Med. Microbiol. 2020, 31, 1. [Google Scholar] [CrossRef]
- Ghorbani, M.; Farhoudi, R. Leishmaniasis in Humans: Drug or Vaccine Therapy? Drug Des. Dev. Ther. 2017, 12, 25–40. [Google Scholar] [CrossRef] [Green Version]
- Puhl, A.C.; Garzino Demo, A.; Makarov, V.A.; Ekins, S. New Targets for HIV Drug Discovery. Drug Discov. Today 2019, 24, 1139–1147. [Google Scholar] [CrossRef]
- Cabarcas-Montalvo, M.; Maldonado-Rojas, W.; Montes-Grajales, D.; Bertel-Sevilla, A.; Wagner-Döbler, I.; Sztajer, H.; Reck, M.; Flechas-Alarcon, M.; Ocazionez, R.; Olivero-Verbel, J. Discovery of Antiviral Molecules for Dengue: In Silico Search and Biological Evaluation. Eur. J. Med. Chem. 2016, 110, 87–97. [Google Scholar] [CrossRef]
- Johansson, J.; Larsson, M.H.; Hornberg, J.J. Predictive in Vitro Toxicology Screening to Guide Chemical Design in Drug Discovery. Curr. Opin. Toxicol. 2019, 15, 99–108. [Google Scholar] [CrossRef]
- Silva, D.R.; Sardi, J.D.C.O.; Freires, I.A.; Silva, A.C.B.; Rosalen, P.L. In Silico Approaches for Screening Molecular Targets in Candida Albicans: A Proteomic Insight into Drug Discovery and Development. Eur. J. Pharmacol. 2019, 842, 64–69. [Google Scholar] [CrossRef]
- Rognan, D. The Impact of in Silico Screening in the Discovery of Novel and Safer Drug Candidates. Pharmacol. Ther. 2017, 175, 47–66. [Google Scholar] [CrossRef]
- Yi, F.; Li, L.; Xu, L.; Meng, H.; Dong, Y.; Liu, H.; Xiao, P. In Silico Approach in Reveal Traditional Medicine Plants Pharmacological Material Basis. Chin. Med. 2018, 13, 33. [Google Scholar] [CrossRef] [Green Version]
- Vivo, M.D.; Cavalli, A. Recent Advances in Dynamic Docking for Drug Discovery. WIREs Comput. Mol. Sci. 2017, 7, e1320. [Google Scholar] [CrossRef]
- Murakami, Y.; Tripathi, L.P.; Prathipati, P.; Mizuguchi, K. Network Analysis and in Silico Prediction of Protein–Protein Interactions with Applications in Drug Discovery. Curr. Opin. Struct. Biol. 2017, 44, 134–142. [Google Scholar] [CrossRef]
- Villanueva, C.E.; Diaz-Castillo, F.; Gómez-Estrada, H. Preliminary Phytochemical Screening of Promising Plant Species of the Colombian Atlantic Coast. Rev. Cuba. Plantas Med. 2013, 18, 619–631. [Google Scholar]
- Fuentes, D.D.C.C.; Buendía, Y.C.O.; Mateus, L.C.O.; Díaz, R.D.S.; Castillo, F.D.; Ospina, L.A.F. Extractos del Caribe Colombiano con actividad antiproliferativa frente a líneas celulares de cáncer de pulmón. Rev. Cuba. Farm. 2019, 51. [Google Scholar]
- Rodríguez-Cavallo, E.; Guarnizo-Méndez, J.; Yépez-Terrill, A.; Cárdenas-Rivero, A.; Díaz-Castillo, F.; Méndez-Cuadro, D. Protein Carbonylation Is a Mediator in Larvicidal Mechanisms of Tabernaemontana Cymosa Ethanolic Extract. J. King Saud Univ. Sci. 2019, 31, 464–471. [Google Scholar] [CrossRef]
- Lounasmaa, M.; Tolvanen, A. Chapter 1 Eburnamine—Vincamine Alkaloids. In The Alkaloids: Chemistry and Pharmacology; Cordell, G.A., Ed.; Academic Press: Cambridge, MA, USA, 1992; Volume 42, pp. 1–116. [Google Scholar]
- Thomas, D.W.; Biemann, K. The Hydroxyindolenine Derivative of Voacangine, a New Indole Alkaloid from Voacanga Africana. Tetrahedron 1968, 24, 4223–4231. [Google Scholar] [CrossRef]
- Cava, M.P.; Tjoa, S.S.; Ahmed, Q.A.; Da Rocha, A.I. The Alkaloids of Tabernaemontana Riedelii and T. Rigida. J. Org. Chem. 1968, 33, 1055–1059. [Google Scholar] [CrossRef]
- Prakash Chaturvedula, V.S.; Sprague, S.; Schilling, J.K.; Kingston, D.G.I. New Cytotoxic Indole Alkaloids from Tabernaemontana Calcarea from the Madagascar Rainforest. J. Nat. Prod. 2003, 66, 528–531. [Google Scholar] [CrossRef]
- Sharma, P.; Cordell, G.A. Heyneanine Hydroxyindolenine, a New Indole Alkaloid from Ervatamia Coronaria Var. Plena. J. Nat. Prod. 1988, 51, 528–531. [Google Scholar] [CrossRef]
- Kutney, J.P.; Perez, I. Studies on Natural Products from Cuban Plants. Alkaloids from Tabernaemontana Citrifolia. Helv. Chim. Acta 1982, 65, 2242–2250. [Google Scholar] [CrossRef]
- Achenbach, H.; Benirschke, M.; Torrenegra, R. Alkaloids and Other Compounds from Seeds of Tabernaemontana Cymosa. Phytochemistry 1997, 45, 325–335. [Google Scholar] [CrossRef]
- Van Beek, T.A.; Kuijlaars, F.L.C.; Thomassen, P.H.A.M.; Verpoorte, R.; Baerheim Svendsen, A. Antimicrobially Active Alkaloids from Tabernaemontana Pachysiphon. Phytochemistry 1984, 23, 1771–1778. [Google Scholar] [CrossRef]
- Pereira, P.S.; de Castro França, S.; de Oliveira, P.V.A.; de Souza Breves, C.M.; Pereira, S.I.V.; Sampaio, S.V.; Nomizo, A.; Dias, D.A. Chemical Constituents from Tabernaemontana Catharinensis Root Bark: A Brief NMR Review of Indole Alkaloids and in Vitro Cytotoxicity. Quím. Nova 2008, 31, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Nicola, C.; Salvador, M.; Escalona Gower, A.; Moura, S.; Echeverrigaray, S. Chemical Constituents Antioxidant and Anticholinesterasic Activity of Tabernaemontana Catharinensis. Sci. World J. 2013, 2013, e519858. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.; Starmer, G.A. The Isolation and Identification of the Major Alkaloid Present in Tabernaemontana Pachysiphon Stapf. Var Cumminsi (Stapf.) H. Huber. J. Pharm. Pharmacol. 1963, 15, 487. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Li, N.; Zhang, W.; Chen, J.; Chen, Z. Simultaneous Determination of Vinblastine and Its Monomeric Precursors Vindoline and Catharanthine in Catharanthus Roseus by Capillary Electrophoresis-Mass Spectrometry. J. Sep. Sci. 2011, 34, 2885–2892. [Google Scholar] [CrossRef]
- Arambewela, L.S.R.; Ranatunge, T. Indole Alkaloids from Tabernaemontana Divaricata. Phytochemistry 1991, 30, 1740–1741. [Google Scholar] [CrossRef]
- Jasiecki, J.; Jońca, J.; Żuk, M.; Szczoczarz, A.; Janaszak-Jasiecka, A.; Lewandowski, K.; Waleron, K.; Wasąg, B. Activity and Polymorphisms of Butyrylcholinesterase in a Polish Population. Chem. Biol. Interact 2016, 259, 70–77. [Google Scholar] [CrossRef]
- Sridharan, S.; Basu, A. Distinct Roles of MTOR Targets S6K1 and S6K2 in Breast Cancer. Int. J. Mol. Sci. 2020, 21, 1199. [Google Scholar] [CrossRef] [Green Version]
- Fessler, M.B. The Challenges and Promise of Targeting the Liver X Receptors for Treatment of Inflammatory Disease. Pharmacol. Ther. 2018, 181, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, J.E.; Garro, A.; Pigni, N.B.; Agüero, M.B.; Roitman, G.; Slanis, A.; Enriz, R.D.; Feresin, G.E.; Bastida, J.; Tapia, A. Cholinesterase-Inhibitory Effect and in Silico Analysis of Alkaloids from Bulbs of Hieronymiella Species. Phytomedicine 2018, 39, 66–74. [Google Scholar] [CrossRef] [Green Version]
- Scotti, L.; Scotti, M.T. In Silico Studies Applied to Natural Products with Potential Activity Against Alzheimer’s Disease. In Computational Modeling of Drugs Against Alzheimer’s Disease; Roy, K., Ed.; Neuromethods; Springer: New York, NY, USA, 2018; pp. 513–531. ISBN 978-1-4939-7404-7. [Google Scholar]
- Hamid, H.A.; Ramli, A.N.M.; Yusoff, M.M. Indole Alkaloids from Plants as Potential Leads for Antidepressant Drugs: A Mini Review. Front. Pharmacol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Kallan, N.C.; Spencer, K.L.; Blake, J.F.; Xu, R.; Heizer, J.; Bencsik, J.R.; Mitchell, I.S.; Gloor, S.L.; Martinson, M.; Risom, T.; et al. Discovery and SAR of Spirochromane Akt Inhibitors. Bioorg. Med. Chem. Lett. 2011, 21, 2410–2414. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Zeng, M.; Pan, Z.; Wu, F.; Guo, L.; He, G. Discovery of Novel Akt1 Inhibitor Induces Autophagy Associated Death in Hepatocellular Carcinoma Cells. Eur. J. Med. Chem. 2020, 189, 112076. [Google Scholar] [CrossRef]
- Dai, Y.; Jin, S.; Li, X.; Wang, D. The Involvement of Bcl-2 Family Proteins in AKT-Regulated Cell Survival in Cisplatin Resistant Epithelial Ovarian Cancer. Oncotarget 2017, 8, 1354–1368. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, K.; Smith, R.S.; Hsieh, C.-M.; Sun, J.; Chao, J.; Liao, J.K. Activation of the Phosphatidylinositol 3-Kinase/Protein Kinase Akt Pathway Mediates Nitric Oxide-Induced Endothelial Cell Migration and Angiogenesis. Mol. Cell. Biol. 2003, 23, 5726–5737. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.-Y.; Ye, Z.-X.; Wang, X.-F.; Chang, J.; Yang, M.-W.; Zhong, H.-H.; Hong, F.-F.; Yang, S.-L. Nitric Oxide Bioavailability Dysfunction Involves in Atherosclerosis. Biomed. Pharmacother. 2018, 97, 423–428. [Google Scholar] [CrossRef]
- Chandarlapaty, S.; Scaltriti, M.; Angelini, P.; Ye, Q.; Guzman, M.; Hudis, C.A.; Norton, L.; Solit, D.B.; Arribas, J.; Baselga, J.; et al. Inhibitors of HSP90 Block P95-HER2 Signaling in Trastuzumab-Resistant Tumors and Suppress Their Growth. Oncogene 2010, 29, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Costa, T.E.M.M.; Raghavendra, N.M.; Penido, C. Natural Heat Shock Protein 90 Inhibitors in Cancer and Inflammation. Eur. J. Med. Chem. 2020, 189, 112063. [Google Scholar] [CrossRef]
- Li, H.; Kang, T.; Qi, B.; Kong, L.; Jiao, Y.; Cao, Y.; Zhang, J.; Yang, J. Neuroprotective Effects of Ginseng Protein on PI3K/Akt Signaling Pathway in the Hippocampus of D-Galactose/AlCl3 Inducing Rats Model of Alzheimer’s Disease. J. Ethnopharmacol. 2016, 179, 162–169. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, J.S.; Massi, D.; Teng, M.W.L.; Mandala, M. PI3K-AKT-MTOR Inhibition in Cancer Immunotherapy, Redux. Semin. Cancer Biol. 2018, 48, 91–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrivastava, S.; Kulkarni, P.; Thummuri, D.; Jeengar, M.K.; Naidu, V.G.M.; Alvala, M.; Redddy, G.B.; Ramakrishna, S. Piperlongumine, an Alkaloid Causes Inhibition of PI3 K/Akt/MTOR Signaling Axis to Induce Caspase-Dependent Apoptosis in Human Triple-Negative Breast Cancer Cells. Apoptosis 2014, 19, 1148–1164. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Sun, J.; Xie, W.; Zhang, M.; Zhang, C.; Xu, X. Stemona Alkaloids Suppress the Positive Feedback Loop between M2 Polarization and Fibroblast Differentiation by Inhibiting JAK2/STAT3 Pathway in Fibroblasts and CXCR4/PI3K/AKT1 Pathway in Macrophages. Int. Immunopharmacol. 2019, 72, 385–394. [Google Scholar] [CrossRef]
- Liu, Y.; Yin, Y.; Zhang, Z.; Li, C.J.; Zhang, H.; Zhang, D.; Jiang, C.; Nomie, K.; Zhang, L.; Wang, M.L.; et al. Structural Optimization Elaborates Novel Potent Akt Inhibitors with Promising Anticancer Activity. Eur. J. Med. Chem. 2017, 138, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Zheng, T.-T.; Li, X.; Liang, Y.; Wang, L.-J.; Huang, Y.-C.; Xiao, H.-T. Plant-Derived Alkaloids: The Promising Disease-Modifying Agents for Inflammatory Bowel Disease. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naidoo, C.M.; Naidoo, Y.; Dewir, Y.H.; Murthy, H.N.; El-Hendawy, S.; Al-Suhaibani, N. Major Bioactive Alkaloids and Biological Activities of Tabernaemontana Species (Apocynaceae). Plants 2021, 10, 313. [Google Scholar] [CrossRef]
- Hussain, H.; Hussain, J.; Al-Harrasi, A.; Green, I.R. Chemistry and Biology of the Genus Voacanga. Pharm. Biol. 2012, 50, 1183–1193. [Google Scholar] [CrossRef] [PubMed]
- Silveira, D.; de Melo, A.M.M.F.; Magalhães, P.O.; Fonseca-Bazzo, Y.M. Chapter 7—Tabernaemontana Species: Promising Sources of New Useful Drugs. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 54, pp. 227–289. [Google Scholar]
- Zhou, S.-Y.; Zhou, T.-L.; Qiu, G.; Huan, X.; Miao, Z.-H.; Yang, S.-P.; Cao, S.; Fan, F.; Cai, Y.-S. Three New Cytotoxic Monoterpenoid Bisindole Alkaloids from Tabernaemontana Bufalina. Planta Med. 2018, 84, 1127–1133. [Google Scholar] [CrossRef] [Green Version]
- Singh, T.P.; Singh, O.M. Recent Progress in Biological Activities of Indole and Indole Alkaloids. Mini Rev. Med. Chem. 2018, 18, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.; Yamashita, H. Cytotoxic Effects of Diterpenoid Alkaloids Against Human Cancer Cells. Molecules 2019, 24, 2317. [Google Scholar] [CrossRef] [Green Version]
- Mejía-Gutiérrez, M.; Olivero-Verbel, J.; Quiroga, J.; Romo, P.; Castaño, S.; Fierro, L. Prediction of the Potential Biological Activity of Novel Spiropyrazolo[3,4-b]Pyridines and Spiropyrazolo[3,4-b]Pyridine-5,5′-Pyrimidines by a Ligand–Protein Inverse-Docking Approach. J. Mol. Gr. Model. 2020, 97, 107581. [Google Scholar] [CrossRef]
- Montes-Grajales, D.; Bernardes, G.J.L.; Olivero-Verbel, J. Urban Endocrine Disruptors Targeting Breast Cancer Proteins. Chem. Res. Toxicol. 2016, 29, 150–161. [Google Scholar] [CrossRef] [Green Version]
- Maldonado-Rojas, W.; Olivero-Verbel, J.; Marrero-Ponce, Y. Computational Fishing of New DNA Methyltransferase Inhibitors from Natural Products. J. Mol. Gr. Model. 2015, 60, 43–54. [Google Scholar] [CrossRef]
- Montes-Grajales, D.; Olivero-Verbel, J. Computer-Aided Identification of Novel Protein Targets of Bisphenol A. Toxicol. Lett. 2013, 222, 312–320. [Google Scholar] [CrossRef]
- Diaz-Castillo, F.; Cardona, S.M.M.; Medina, M.C.; González, Y.P.; Gómez-Estrada, H. Larvicidal Activity of Ethanol Extracts of Tabernaemontana Cymosa and Trichilia Hirta against III and IV Stage Larvae of Aedes Aegypti (Diptera: Culicidae). Rev. Cuba. Plantas Med. 2012, 17, 256–267. [Google Scholar]
- Lavaud, C.; Massiot, G. The Iboga Alkaloids. Prog. Chem. Org. Nat. Prod. 2017, 105, 89–136. [Google Scholar] [CrossRef]
- Mekky, R.H.; Abdel-Sattar, E.; Segura-Carretero, A.; Contreras, M.D.M. Phenolic Compounds from Sesame Cake and Antioxidant Activity: A New Insight for Agri-Food Residues’ Significance for Sustainable Development. Foods 2019, 8, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Montes-Grajales, D.; Olivero-Verbel, J. Structure-Based Identification of Endocrine Disrupting Pesticides Targeting Breast Cancer Proteins. Toxicology 2020, 439, 152459. [Google Scholar] [CrossRef]
- Kelly-Smith, M.; Strain, G.M. STRING Data Mining of GWAS Data in Canine Hereditary Pigment-Associated Deafness. Vet. Anim. Sci. 2020, 9, 100118. [Google Scholar] [CrossRef] [PubMed]
- Shanmugapriya; Othman, N.; Sasidharan, S. Prediction of Genes and Protein-Protein Interaction Networking for MiR-221-5p Using Bioinformatics Analysis. Gene Rep. 2019, 16, 100426. [Google Scholar] [CrossRef]
- Wei, L.K.; Quan, L.S. Biomarkers for Ischemic Stroke Subtypes: A Protein-Protein Interaction Analysis. Comput. Biol. Chem. 2019, 83, 107116. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pick (N°) | Retention Time | m/z | Formula | Detected Molecule * | Reference |
|---|---|---|---|---|---|
| 1 | 9.77 | 399.1913 | C22H26O5N2 | Vincinine | [35] |
| 2 | 10.65 | 385.2122 | C22H28O4N2 | Rupicoline/voacangine | [36] |
| 3 | 10.88 | 353.1858 | C21H24O3N2 | D-Minovincine | [37] |
| 4 | 10.88 | 399.1913 | C22H26O5N2 | - | |
| 5 | 11.18 | 385.2118 | C22H28O4N2 | Voacangarine | [38] |
| 6 | 12.18 | 353.1858 | C21H24O3N2 | 6/5/3-oxocoronaridine | [39] |
| 7 | 12.45 | 383.1961 | C22H26O4N2 | 19-oxovoacangine/19-iso-oxovoacangine | [40] |
| 8 | 12.7 | 351.1701 | C21H22O3N2 | 3/5-Oxotabersonine | [41] |
| 9 | 12.97 | 383.1964 | C22H26O4N2 | 19-oxovoacangine/19-iso-oxovoacangine | [40] |
| 10 | 14.36 | 415.223 | C23H30O5N2 | 20-hidroxyconopharyngine | [42] |
| 11 | 15.44 | 397.1858 | C22H24O5N2 | - | |
| 12 | 16.05 | 353.1856 | C21H24O3N2 | 6/5/3-oxocoronaridine | [39] |
| 13 | 16.15 | 383.1923 | C22H26O4N2 | - | |
| 14 | 16.44 | 385.2118 | C22H28O4N2 | Voacristine/voacangine-7-hydroxyndolenine | [43] |
| 15 | 16.6 | 367.2012 | C22H26O3N2 | Voachalotine/corymantheine | [44] |
| 16 | 16.94 | 397.212 | C23H28O4N2 | - | |
| 17 | 17.06 | 385.2118 | C22H28O4N2 | Voacristine/isovoacristine | [43] |
| 18 | 17.27 | 399.2247 | C23H30O4N2 | Conopharyngine | [45] |
| 19 | 12.64 | 367.2013 | C22H26O3N2 | Voachalotine | [44] |
| 20 | 18.51 | 369.2179 | C22H28O3N2 | Voacangine/isovoacangine | [38] |
| 21 | 18.9 | 337.1908 | C21H24O2N2 | Catharantine | [46] |
| 22 | 18.9 | 369.2179 | C22H28O3N2 | Voacangine/isovoacangine | [38] |
| 23 | 20.19 | 383.2326 | C23H30O3N2 | N-Metil-isovoacangine | [47] |
| PDB_ ID | Predicted Binding Energy (kcal/mol) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 5-oxocoronaridine | Voacangine-7-hydroxyndolenine | Voacristine | 5-oxotabersonine | Vincinine | 3-oxocoronaridine | 3-oxotabersonine | Rupicoline | Isovoacristine | Voacangine | |
| 4AOJ * | −11.0 | −8.0 | −9.3 | −8.1 | −10.0 | −8.9 | −8.1 | −8.1 | −7.7 | −8.0 |
| 4MJH | −10.9 | −9.9 | −9.0 | −9.5 | −9.0 | −9.9 | −9.8 | −9.6 | −9.5 | −8.9 |
| 3EAH | −10.7 | −8.2 | −8.7 | −9.1 | −9.6 | −8.7 | −8.4 | −8.7 | −8.4 | −9.1 |
| 4DRJ | −10.5 | −9.7 | −9.1 | −9.3 | −9.4 | −9.3 | −9.4 | −8.9 | −9.4 | −8.9 |
| 4NOS | −10.4 | −8.3 | −9.8 | −8.3 | −8.4 | −8.7 | −8.5 | −8.0 | −7.8 | −8.6 |
| 1N83 | −10.2 | −10.0 | −9.1 | −8.6 | −9.1 | −8.1 | −8.8 | −9.9 | −10.2 | −9.0 |
| 3O3U | −10.2 | −9.3 | −9.1 | −9.5 | −10.0 | −8.5 | −10.1 | −9.0 | −8.6 | −8.8 |
| 4XII | −10.2 | −9.2 | −9.2 | −9.9 | −9.3 | −9.6 | −9.4 | −10.0 | −9.6 | −9.4 |
| 1HFQ | −10.0 | 0.0 | −9.2 | −9.8 | −9.2 | −9.4 | −9.9 | −10.0 | −9.2 | −9.1 |
| 1RY8 | −10.0 | −9.8 | −9.6 | −9.9 | −8.9 | −8.2 | −9.7 | −9.0 | −9.0 | −9.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveros-Díaz, A.; Olivero-Verbel, J.; Pájaro-González, Y.; Díaz-Castillo, F. Molecular Human Targets of Bioactive Alkaloid-Type Compounds from Tabernaemontana cymose Jacq. Molecules 2021, 26, 3765. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26123765
Oliveros-Díaz A, Olivero-Verbel J, Pájaro-González Y, Díaz-Castillo F. Molecular Human Targets of Bioactive Alkaloid-Type Compounds from Tabernaemontana cymose Jacq. Molecules. 2021; 26(12):3765. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26123765
Chicago/Turabian StyleOliveros-Díaz, Andrés, Jesús Olivero-Verbel, Yina Pájaro-González, and Fredyc Díaz-Castillo. 2021. "Molecular Human Targets of Bioactive Alkaloid-Type Compounds from Tabernaemontana cymose Jacq." Molecules 26, no. 12: 3765. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26123765