
Presence of Cholesterol in Non-Animal Organisms: Identification and Quantification of Cholesterol in Crude Seed Oil from Perilla frutescens and Dehydrated Pyropia tenera

Abstract
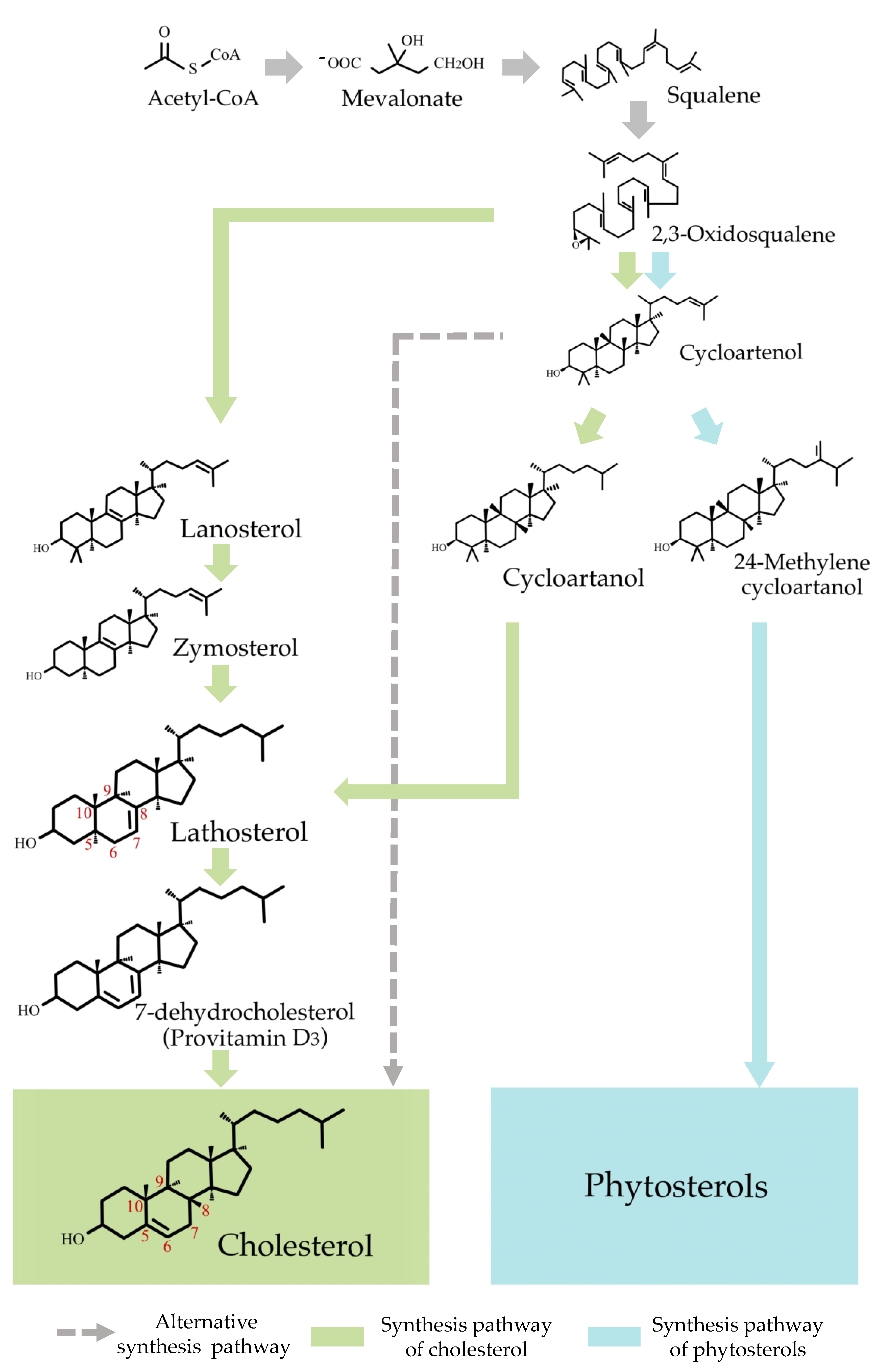
:1. Introduction
2. Results and Discussion
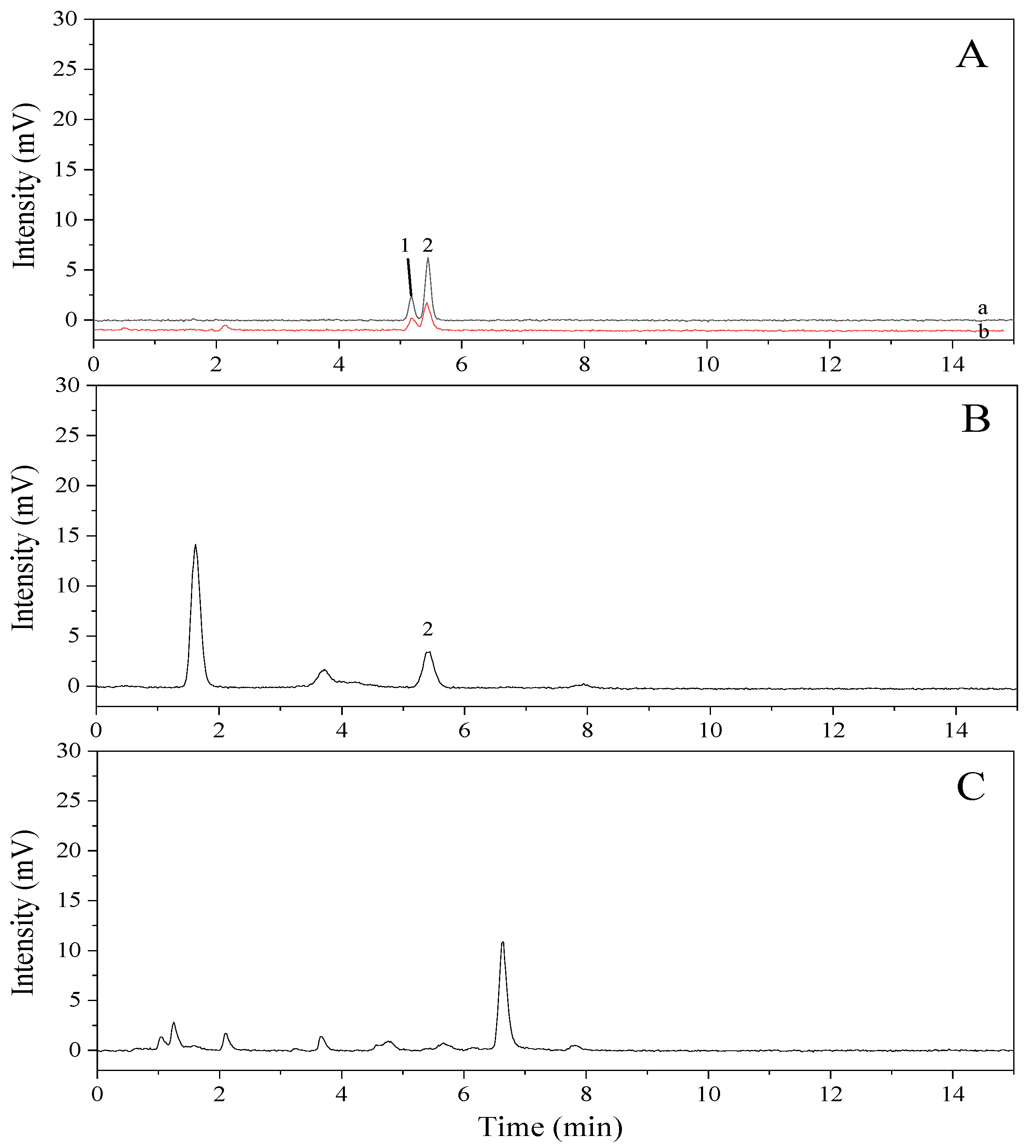
2.1. Determination of Cholesterol by Gas Chromatography (GC)
2.2. Determination of Cholesterol by High-Performance Liquid Chromatography (HPLC)
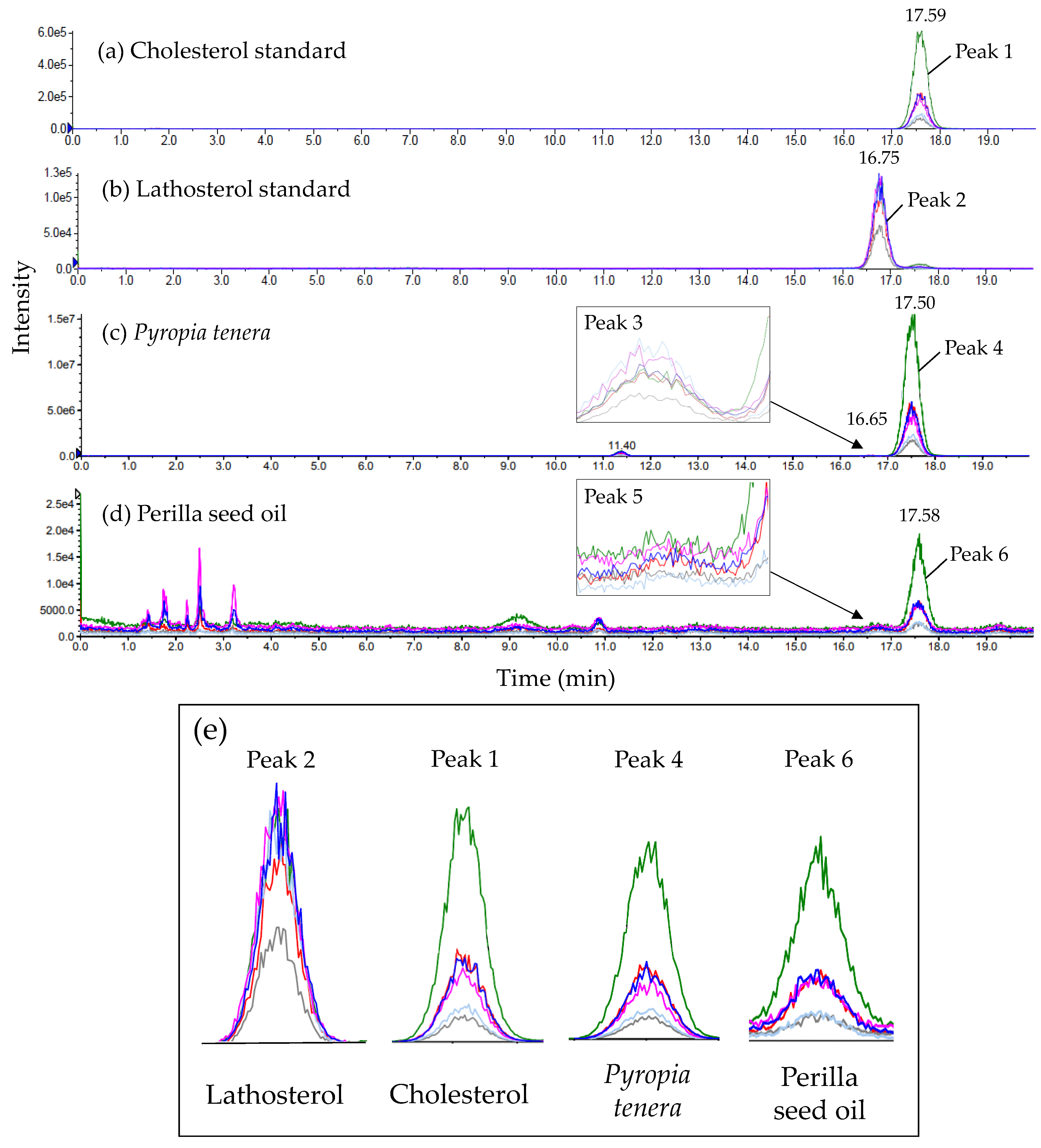
2.3. Identification of Cholesterol by Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
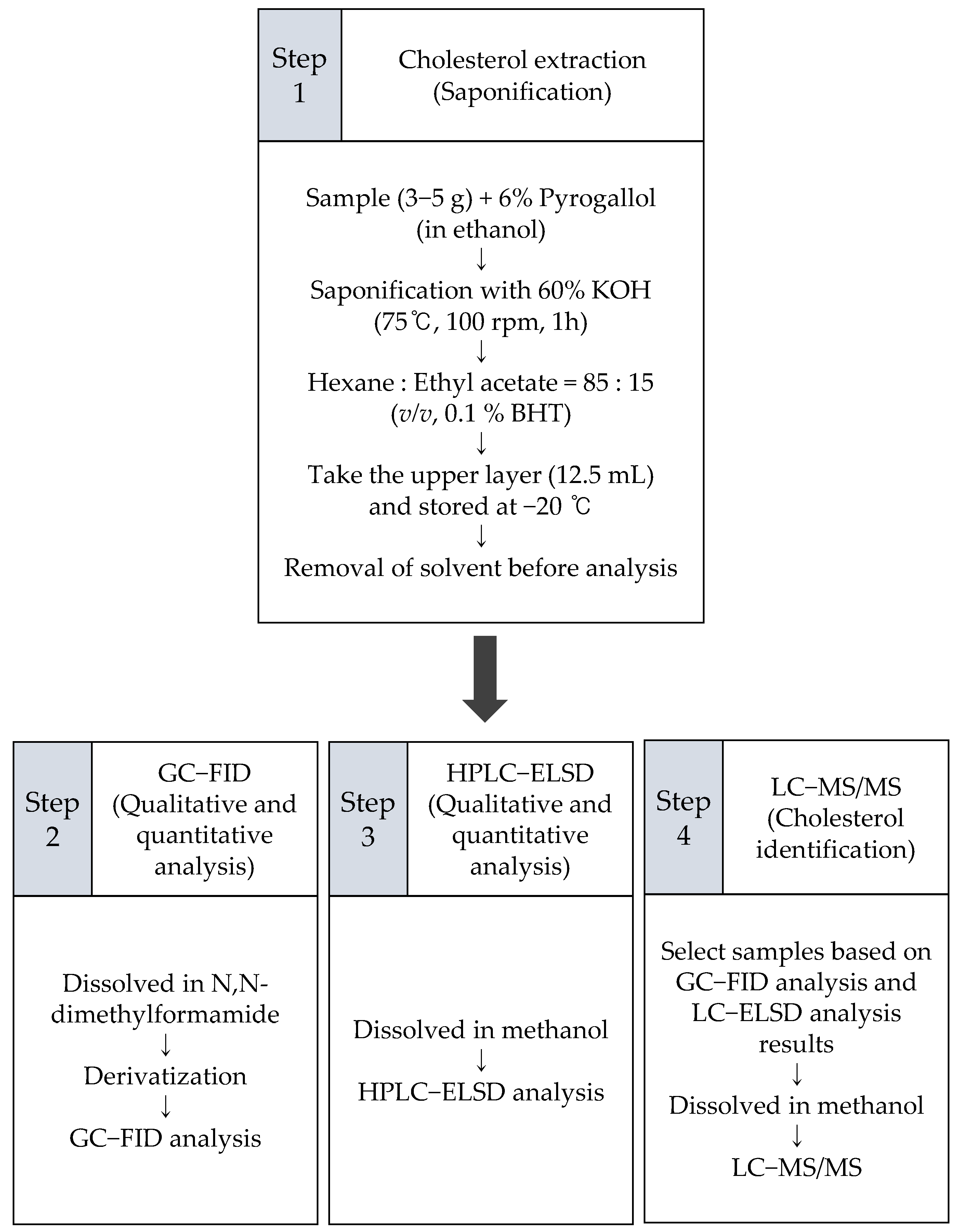
3. Material and Methods
3.1. Materials and Reagents
3.2. Extraction
3.3. Cholesterol Analysis Using Gas Chromatography-Flame Ionization Detector (GC-FID)
3.4. Cholesterol Analysis Using High-Performance Liquid Chromatography-Evaporative Light-Scattering Detector (HPLC-ELSD)
3.5. Cholesterol Analysis Using Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS) with Atmospheric Pressure Chemical Ionization (APCI)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Tsoupras, A.; Lordan, R.; Zabetakis, I. Inflammation, not Cholesterol, Is a Cause of Chronic Disease. Nutrients 2018, 10, 604. [Google Scholar] [CrossRef] [Green Version]
- Mouritsen, O.G.; Zuckermann, M.J. What’s so special about cholesterol? Lipids 2004, 39, 1101–1113. [Google Scholar] [CrossRef] [PubMed]
- Andrade, L.D.O. Understanding the role of cholesterol in cellular biomechanics and regulation of vesicular trafficking: The power of imaging. Biomed. Spectrosc. Imaging 2016, 5, S101–S117. [Google Scholar] [CrossRef] [Green Version]
- Sheriff, D.S.; Ali, E.F. Perspective on plasma membrane cholesterol efflux and spermatozoal function. J. Hum. Reprod. Sci. 2010, 3, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Pandit, S.A.; Bostick, D.; Berkowitz, M.L. Complexation of Phosphatidylcholine Lipids with Cholesterol. Biophys. J. 2004, 86, 1345–1356. [Google Scholar] [CrossRef] [Green Version]
- Rowlands, L.J.; Marks, A.; Sanderson, J.M.; Law, R.V. 17O NMR spectroscopy as a tool to study hydrogen bonding of cholesterol in lipid bilayers. Chem. Commun. 2020, 56, 14499–14502. [Google Scholar] [CrossRef]
- Clayton, P.T. Disorders of bile acid synthesis. In Inborn Metabolic Diseases; Fernandes, J., Saudubray, J.-M., van den Berghe, G., Walter, J.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 473–484. Available online: https://0-doi-org.brum.beds.ac.uk/10.1007/978-3-540-28785-8_34 (accessed on 18 May 2021).
- Mayes, P.A.; Botham, K.M. Cholesterol synthesis, transport and excretion. In Harper’s Illustrated Biochemistry; Robert, K.M., Daryl, K.G., Peter, A.M., Victor, W.R., Eds.; Tata McGraw Hill Education: New York City, NY, USA, 2003; p. 219. [Google Scholar]
- Miller, W.L. Molecular Biology of Steroid Hormone Synthesis. Endocr. Rev. 1988, 9, 295–318. [Google Scholar] [CrossRef]
- Gylling, H. Clinical utility of serum markers of cholesterol absorption and synthesis. Curr. Opin. Lipidol. 2014, 25, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Quintao, E.; Grundy, S.M.; Ahrens, E. Effects of dietary cholesterol on the regulation of total body cholesterol in man. J. Lipid Res. 1971, 12, 233–247. [Google Scholar] [CrossRef]
- Sweeney, J.P.; Weihrauch, J.L.; WeBirauch, J.L. Summary of available data for cholesterol in foods and methods for its determination. C R C Crit. Rev. Food Sci. Nutr. 1976, 8, 131–159. [Google Scholar] [CrossRef]
- Dietary Guidelines Advisory Committee. Scientific Report of the 2015 Dietary Guidelines Advisory Committee: Advisory Report to the Secretary of Health and Human Services and the Secretary of Agriculture; Agricultural Research Service: Washington, DC, USA, 2015; p. 58. Available online: https://health.gov/sites/default/files/2019-09/Scientific-Report-of-the-2015-Dietary-Guidelines-Advisory-Committee.pdf (accessed on 18 May 2021).
- MFDS. Labeling Standards of Foods, Etc; Ministry of Food and Drug Safety: Osong, Korea, 2019; pp. 67–68. Available online: https://www.mfds.go.kr/eng/brd/m_15/view.do?seq=72427&srchFr=&srchTo=&srchWord=&srchTp=&itm_seq_1=0&itm_seq_2=0&multi_itm_seq=0&company_cd=&company_nm=&page=1 (accessed on 18 May 2021).
- Ghaleshahi, A.Z.; Ezzatpanah, H.; Rajabzadeh, G.; Ghavami, M. Comparison and analysis characteristics of flax, perilla and basil seed oils cultivated in Iran. J. Food Sci. Technol. 2020, 57, 1258–1268. [Google Scholar] [CrossRef]
- Uma Maheswari, M.; Reena, A. Phytochemical profiling of the red seaweed, Halymenia dilatata by GC-MS analysis. Int. J. Pharm. Sci. Res. 2017, 8, 167–172. [Google Scholar]
- Schwartz, H.; Ollilainen, V.; Piironen, V.; Lampi, A.-M. Tocopherol, tocotrienol and plant sterol contents of vegetable oils and industrial fats. J. Food Compos. Anal. 2008, 21, 152–161. [Google Scholar] [CrossRef]
- Govindan, M.; Hodge, J.D.; Brown, K.A.; Nuñez-Smith, M. Distribution of cholesterol in Caribbean marine algae. Steroids 1993, 58, 178–180. [Google Scholar] [CrossRef]
- Sánchez-Machado, D.I.; Lopez-Hernandez, J.; Paseiro-Losada, P.; López-Cervantes, J. An HPLC method for the quantification of sterols in edible seaweeds. Biomed. Chromatogr. 2004, 18, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Behrman, E.J.; Gopalan, V. Cholesterol and Plants. J. Chem. Educ. 2005, 82, 1791. [Google Scholar] [CrossRef]
- Sonawane, P.D.; Pollier, J.; Panda, S.; Szymanski, J.; Massalha, H.; Yona, M.; Unger, T.; Malitsky, S.; Arendt, P.; Pauwels, L.; et al. Plant cholesterol biosynthetic pathway overlaps with phytosterol metabolism. Nat. Plants 2016, 3, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Eller, F.J.; Moser, J.K.; Kenar, J.A.; Taylor, S.L. Extraction and Analysis of Tomato Seed Oil. J. Am. Oil Chem. Soc. 2010, 87, 755–762. [Google Scholar] [CrossRef] [Green Version]
- Giera, M.; Muller, C.; Bracher, F. Analysis and Experimental Inhibition of Distal Cholesterol Biosynthesis. Chromatographia 2014, 78, 343–358. [Google Scholar] [CrossRef]
- ChemSpider: Cholesterol. Available online: http://www.chemspider.com/Chemical-Structure.5775.html (accessed on 24 February 2021).
- ChemSpider: Lathosterol. Available online: http://www.chemspider.com/Chemical-Structure.59151.html (accessed on 24 February 2021).
- Kamenarska, Z.; Stefanov, K.; Stancheva, R.; Dimitrova-Konaklieva, S.; Popov, S. Comparative investigation on sterols from some Black Sea red algae. Nat. Prod. Res. 2006, 20, 113–118. [Google Scholar] [CrossRef]
- Patterson, G.W. The distribution of sterols in algae. Lipids 1971, 6, 120–127. [Google Scholar] [CrossRef]
- Watanabe, N.; Kimura, F.; Kojima, F.; Endo, Y.; Fujimoto, K.; Kikuchi, Y. Effect of Sterols in Dietary Fats on Whole Blood Viscosity of Stroke-Prone Spontaneously Hypertensive Rats (SHRSP). J. Oleo Sci. 2005, 54, 1–6. [Google Scholar] [CrossRef]
- Torri, L.; Bondioli, P.; Folegatti, L.; Rovellini, P.; Piochi, M.; Morini, G. Development of Perilla seed oil and extra virgin olive oil blends for nutritional, oxidative stability and consumer acceptance improvements. Food Chem. 2019, 286, 584–591. [Google Scholar] [CrossRef]
- Jeong, T.M.; Itoh, T.; Tamura, T.; Matsumoto, T. Analysis of sterol fractions from twenty vegetable oils. Lipids 1974, 9, 921–927. [Google Scholar] [CrossRef]
- Hojo, K.; Hakamata, H.; Kusu, F. Simultaneous determination of serum lathosterol and cholesterol by semi-micro high-performance liquid chromatography with electrochemical detection. J. Chromatogr. B 2011, 879, 751–755. [Google Scholar] [CrossRef] [PubMed]
- Ligor, M.; Kováčová, J.; Gadzała-Kopciuch, R.M.; Studzińska, S.; Bocian, S.; Lehotay, J.; Buszewski, B. Study of RP HPLC Retention Behaviours in Analysis of Carotenoids. Chromatogr. 2014, 77, 1047–1057. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.S. Analysis of polymer additives using LC/MS. Polym. Sci. Tech. 2011, 22, 188–193. [Google Scholar]
- Kang, M.J. Principles and Application of Mass Spectrometry. Polym. Sci. Tech. 2006, 17, 368–377. [Google Scholar]
- Block, C.; Wynants, L.; Kelchtermans, M.; De Boer, R.; Compernolle, F. Identification of polymer additives by liquid chromatography–mass spectrometry. Polym. Degrad. Stab. 2006, 91, 3163–3173. [Google Scholar] [CrossRef]
- Gachumi, G.; El-Aneed, A. Mass Spectrometric Approaches for the Analysis of Phytosterols in Biological Samples. J. Agric. Food Chem. 2017, 65, 10141–10156. [Google Scholar] [CrossRef]
- Honda, A.; Yamashita, K.; Miyazaki, H.; Shirai, M.; Ikegami, T.; Xu, G.; Numazawa, M.; Hara, T.; Matsuzaki, Y. Highly sensitive analysis of sterol profiles in human serum by LC-ESI-MS/MS. J. Lipid Res. 2008, 49, 2063–2073. [Google Scholar] [CrossRef] [Green Version]
- Menicatti, M.; Pallecchi, M.; Bua, S.; Vullo, D.; Mannelli, L.D.C.; Ghelardini, C.; Carta, F.; Supuran, C.T.; Bartolucci, G. Resolution of co-eluting isomers of anti-inflammatory drugs conjugated to carbonic anhydrase inhibitors from plasma in liquid chromatography by energy-resolved tandem mass spectrometry. J. Enzym. Inhib. Med. Chem. 2018, 33, 671–679. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, M.; Ioki, M.; Matsuura, H.; Hashimoto, A.; Kaya, K.; Nakajima, N.; Watanabe, M.M. Diverse steroidogenic pathways in the marine alga Aurantiochytrium. Environ. Boil. Fishes 2020, 32, 1631–1642. [Google Scholar] [CrossRef]
- Belcour, A.; Girard, J.; Aite, M.; Delage, L.; Trottier, C.; Marteau, C.; Leroux, C.; Dittami, S.M.; Sauleau, P.; Corre, E.; et al. Inferring Biochemical Reactions and Metabolite Structures to Understand Metabolic Pathway Drift. iScience 2020, 23, 100849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackay, D.S.; Gebauer, S.K.; Eck, P.K.; Baer, D.J.; Jones, P.J.H. Lathosterol-to-cholesterol ratio in serum predicts cholesterol-lowering response to plant sterol consumption in a dual-center, randomized, single-blind placebo-controlled trial. Am. J. Clin. Nutr. 2015, 101, 432–439. [Google Scholar] [CrossRef] [Green Version]
- Cibickova, L.; Radomir, H.; Stanislav, M.; Norbert, C.; Helena, Z.; Daniel, J.; Alena, T.; Eva, B.; Vladimir, P. The influence of simvastatin, atorvastatin and high-cholesterol diet on acetylcholinesterase activity, amyloid beta and cholesterol synthesis in rat brain. Steroids 2009, 74, 13–19. [Google Scholar] [CrossRef]
- Lee, J.H.; Na Lee, H.; Shin, J.-A.; Chun, J.Y.; Lee, J.; Lee, K.-T. Content of Fat-Soluble Nutrients (Cholesterol, Retinol, and α-Tocopherol) in Different Parts of Poultry Meats according to Cooking Method. J. Korean Soc. Food Sci. Nutr. 2015, 44, 234–241. [Google Scholar] [CrossRef] [Green Version]
- Fu, R.; Joseph, M. LC/ELSD and LC/MS/MS of Cholesterol and Related Sterols on a Poroshell 120 Column. Application Note 2012; pp. 2–5. Available online: http://www.ingenieria-analitica.com/downloads/dl/file/id/2686/product/110/lc_elsd_and_lc_ms_ms_of_cholesterol_and_related_sterols_on_a_poroshell_120_column.pdf (accessed on 18 May 2021).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Instrument | Range a (mg/mL) | Calibration Curve b | R2 | LOD c | LOQ d | Pyropia tenera | Saccharina japonica | Undaria pinnatifida | Perilla Seed Oil |
|---|---|---|---|---|---|---|---|---|---|
| GC-FID | 0.0025−0.2 | 0.9989 | 0.10 | 0.28 | 14.6 | ND | ND | 0.3−0.5 e | |
| HPLC-ELSD | 0.005−0.1 | 0.9985 | 0.23 | 0.66 | 14.2 | ND | ND f | TR g |
| Compounds | MRM Transition | Collision Energy (V) | Peak Area | Relative Ratio a | |
|---|---|---|---|---|---|
| Precursor ion (m/z) | Product ion (m/z) | ||||
| Cholesterol standard | 369.5 | 81.0 | 65 | 3.3 | |
| 95.0 | 38 | 2.7 | |||
| 147.0 | 27 | 3.4 | |||
| 161.1 | 23 | 9.4 | |||
| 189.1 | 29 | 1.4 | |||
| 257.1 | 19 | 1.0 | |||
| Lathosterol standard | 369.5 | 81.0 | 65 | 5.8 | |
| 95.0 | 38 | 7.0 | |||
| 147.0 | 27 | 10.7 | |||
| 161.1 | 23 | 43.2 | |||
| 189.1 | 29 | 1.0 | |||
| 257.1 | 19 | 2.6 | |||
| Pyropiatenera | 369.5 | 81.0 | 65 | 3.2 | |
| 95.0 | 38 | 2.5 | |||
| 147.0 | 27 | 3.3 | |||
| 161.1 | 23 | 9.2 | |||
| 189.1 | 29 | 1.4 | |||
| 257.1 | 19 | 1.0 | |||
| Perilla seed oil | 369.5 | 81.0 | 65 | 3.4 | |
| 95.0 | 38 | 2.7 | |||
| 147.0 | 27 | 3.4 | |||
| 161.1 | 23 | 10.0 | |||
| 189.1 | 29 | 1.4 | |||
| 257.1 | 19 | 1.0 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, M.-J.; So, H.-J.; Hong, E.-S.; Shin, J.-A.; Lee, K.-T. Presence of Cholesterol in Non-Animal Organisms: Identification and Quantification of Cholesterol in Crude Seed Oil from Perilla frutescens and Dehydrated Pyropia tenera. Molecules 2021, 26, 3767. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26123767
Oh M-J, So H-J, Hong E-S, Shin J-A, Lee K-T. Presence of Cholesterol in Non-Animal Organisms: Identification and Quantification of Cholesterol in Crude Seed Oil from Perilla frutescens and Dehydrated Pyropia tenera. Molecules. 2021; 26(12):3767. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26123767
Chicago/Turabian StyleOh, Min-Ji, Hee-Jin So, Eun-Sik Hong, Jung-Ah Shin, and Ki-Teak Lee. 2021. "Presence of Cholesterol in Non-Animal Organisms: Identification and Quantification of Cholesterol in Crude Seed Oil from Perilla frutescens and Dehydrated Pyropia tenera" Molecules 26, no. 12: 3767. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26123767