
Inhibitory Effect of Osthole from Cnidium monnieri (L.) Cusson on Fusarium oxysporum, a Common Fungal Pathogen of Potato

Abstract
:1. Introduction
2. Results
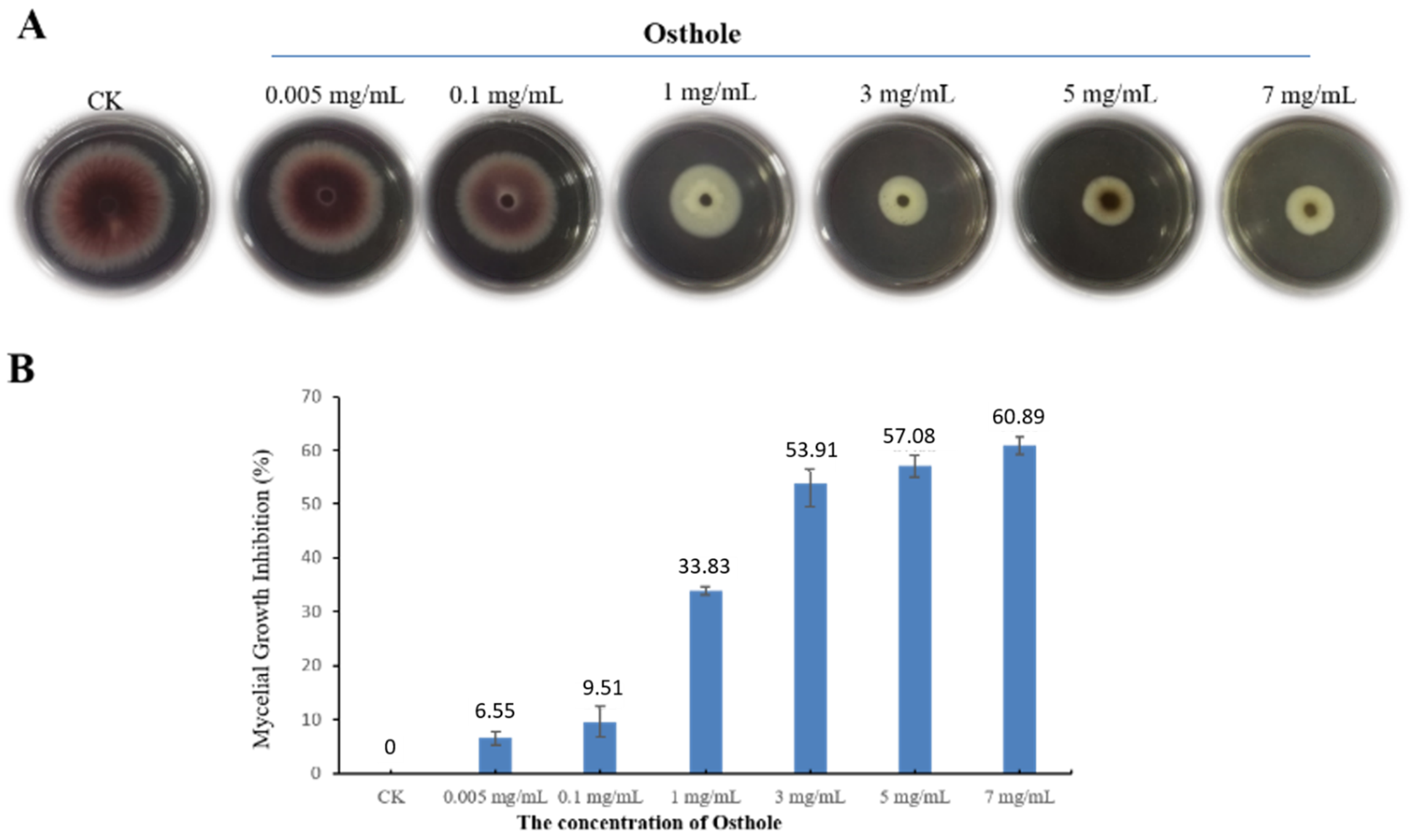
2.1. Osthole Effects on the Growth of F. oxysporum
2.2. The Dose-Dependent Inhibitory Activity of Osthole on the Growth of F. oxysporum
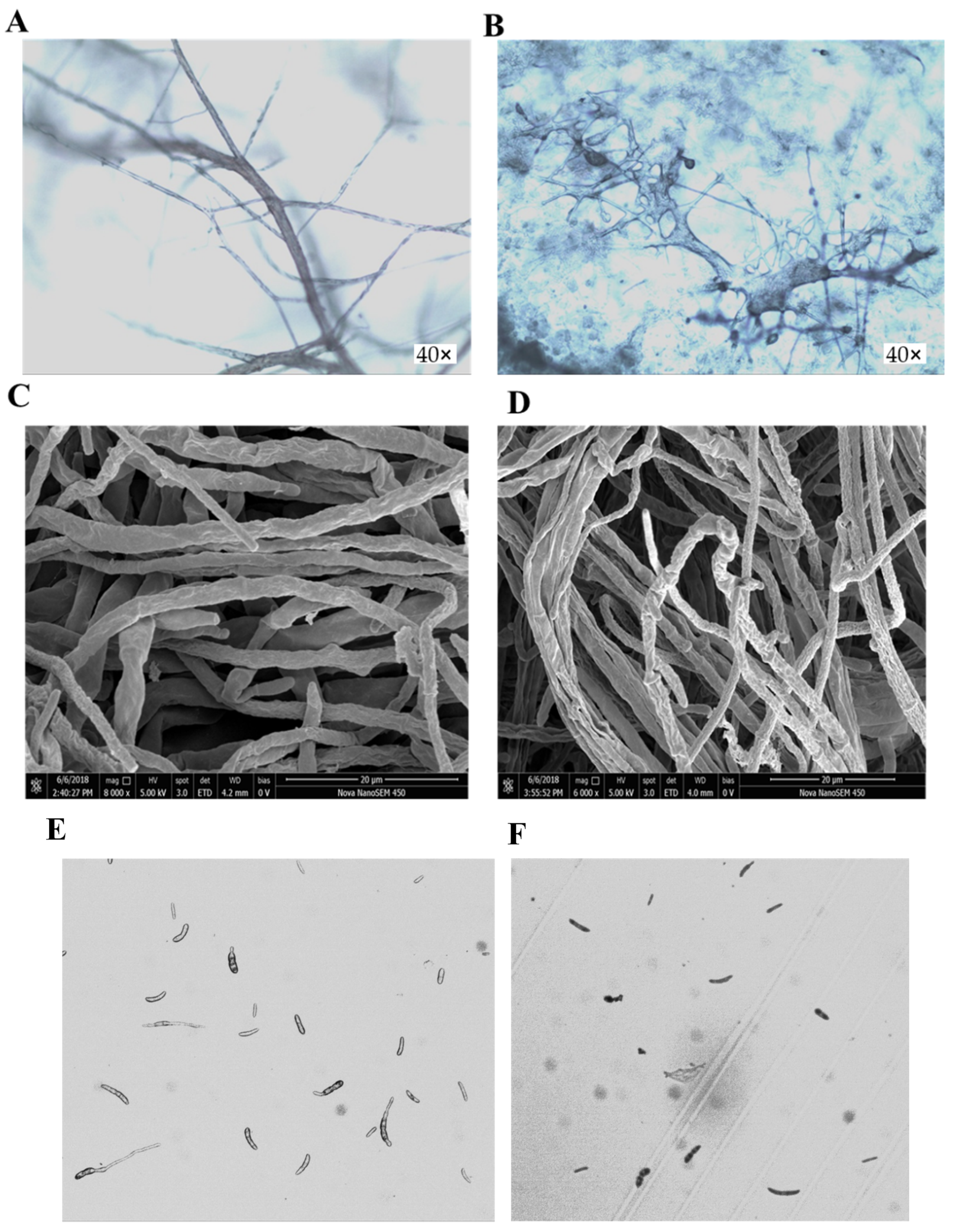
2.3. Effects of Osthole on Mycelia Morphology and Spore Germination of F. oxysporum
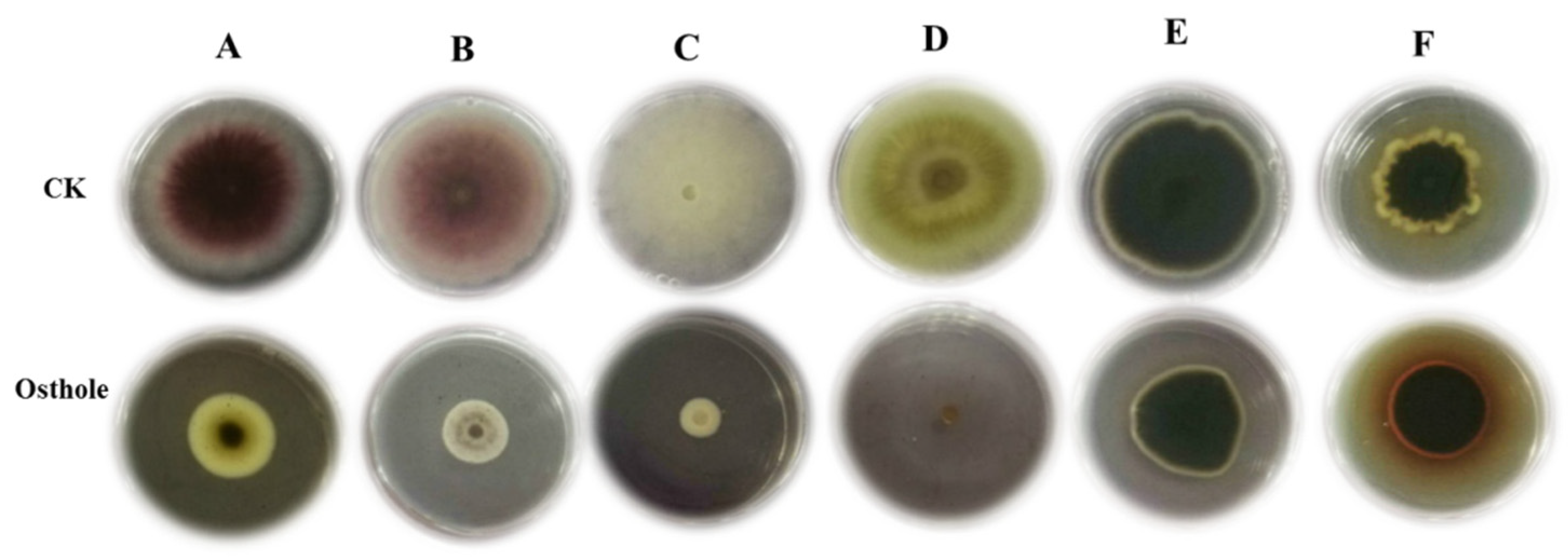
2.4. Inhibitory Activity of Osthole against Other Fungal Pathogens
3. Materials and Methods
3.1. Test Fungal Pathogens and Storage
3.2. Chemicals and Materials
3.3. Inhibitory Activity of Osthole on the Growth of F. oxysporum Schlecht
3.4. Mycelia Observation by Scanning Electron Microscopy (SEM)
3.5. Effects of Osthole on the Germination of F. oxysporum Schlecht Spores
3.6. Statistical Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Rakhimov, U.K.; Khakimov, A.K. Wilt of potatoes in Uzbekistan. Zashchita I Karantin Rastenii 2000, 3, 46. [Google Scholar]
- Peng, X.W.; Zhu, J.H. Species and Distribution of potato Fungal Diseases in Hebei Province, China. Chin. Potato J. 2008, 22, 31–33. [Google Scholar]
- An, X.M.; Hu, J.; Wu, J.H.; Liu, Z.H.; Meng, M.L. Overview of pathogen causing potato Fusarium Wilt. Chin. Potato J. 2017, 31, 302–306. [Google Scholar]
- Khedher, S.B.; Mejdoub-Trabelsi, B.; Tounsi, S. Biological potential of Bacillus subtilis V26 for the control of Fusarium wilt and tuber dry rot on potato caused by Fusarium species and the promotion of plant growth. Biol. Control 2020, 152, 104444. [Google Scholar] [CrossRef]
- Chen, Y.H.; Lu, M.H.; Guo, D.S.; Zhai, Y.Y.; Miao, D.; Yue, J.Y.; Yuan, C.H.; Zhao, M.M.; An, D.R. Antifungal Effect of Magnolol and Honokiol from Magnolia officinalis on Alternaria alternata Causing Tobacco Brown Spot. Molecules 2019, 24, 2140. [Google Scholar] [CrossRef] [Green Version]
- Mi, Y.Y.; Byeong, J.C.; Jin, C.K. Recent Trends in Studies on Botanical Fungicides in Agriculture. Plant Pathol. 2013, 29, 1–9. [Google Scholar]
- Kijjoa, A.; Pinto, M.M.M.; Tantisewie, B.; Herz, W. A biphenyl type neolignan and biphenyl ether from Magnolia henryi. Phytochemistry 1989, 28, 1284–1286. [Google Scholar] [CrossRef]
- Du, C.M.; Wu, Y.H.; Zhao, X.X.; Zhu, C.Y.; Jiang, G.; Yan, X.M. Recent development in research of natural antiphytoviral substances. Acta Tabacaria Sin. 2004, 10, 34–40. [Google Scholar]
- Chen, Y.H.; Ru, B.L.; Zhai, Y.Y.; Li, J.; Cheng, J.L. Screening and inhibitory effects of plant extracts against Tobacco mosaic virus (TMV). J. Plant Prot. 2018, 45, 463–469. [Google Scholar]
- Zhao, L.; Feng, C.; Wu, K.; Chen, W.B.; Chen, Y.J.; Hao, X.A.; Wu, Y.F. Advances and prospects in biogenic substances against plant virus: A review. Pestic. Biochem. Phys. 2016, 135, 15–26. [Google Scholar] [CrossRef]
- Kitajima, J.; Aoki, Y.; Ishikawa, T.; Tanaka, Y. Monoterpenoid glucosides of Cnidium monnieri fruit. Chem. Pharm. Bull. 1999, 47, 639–642. [Google Scholar] [CrossRef] [Green Version]
- Oh, H.; Kim, J.S.; Song, E.K.; Cho, H.; Kim, D.H.; Park, S.E.; Lee, H.S.; Kim, Y.C. Sesquiterpenes with hepatoprotective activity from Cnidium monnieri on tacrine-induced cytotoxicity in Hep G2 cells. Planta Med. 2002, 68, 748–749. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.Y.; Zhou, M.; Liu, Y.; Zhang, G.L.; Luo, Y.G. Chromones and coumarins from the dried fructus of Cnidium monnieri. Fitoterapia 2011, 82, 767–771. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yang, A.; Lenon, G.B. Phytochemistry, Ethnopharmacology, Pharmacokinetics and Toxicology of Cnidium monnieri (L.) Cusson. Int. J. Mol. Sci. 2020, 21, 1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basnet, P.; Yasuda, I.; Kumagai, N.; Tohda, C.; Nojima, H.; Kuraishi, Y.; Komatsu, K. Inhibition of itch-scratch response by fruits of Cnidium monnieri in mice. Biol. Pharm. Bull. 2001, 24, 1012–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, J.J.; Xie, M.L.; Zhu, L.J. Treatment of osthol on osteoporosis in ovariectomized rats. Chin. Pharm. Bull. 2011, 27, 591–592. [Google Scholar]
- Matsuda, H.; Ido, Y.; Hirata, A.; Ino, Y.; Naruto, S.; Amamiya, T.; Kubo, M. Antipruritic effect of Cnidii monnieri Fructus (fruits of Cnidium monnieri Cusson). Biol. Pharm. Bull. 2002, 25, 260–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, H.; Tomohiro, N.; Ido, Y.; Kubo, M. Anti-allergic effects of Cnidii monnieri fructus (dried fruits of Cnidium monnieri) and its major component, osthol. Biol. Pharm. Bull. 2002, 25, 809–812. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.M.; Jia, M.; Li, H.Q.; Zhang, N.D.; Wen, X.; Rahman, K.; Zhang, Q.Y.; Qin, L.P. Cnidium monnieri: A Review of Traditional Uses, Phytochemical and Ethnopharmacological Properties. Am. J. Chin. Med. 2015, 43, 835–877. [Google Scholar] [CrossRef]
- Zhang, Z.R.; Leung, W.N.; Li, G.; Kong, S.K.; Lu, X.; Wong, Y.M.; Chan, C.W. Osthole Enhances Osteogenesis in Osteoblasts by Elevating Transcription Factor Osterix via cAMP/CREB Signaling In Vitro and In Vivo. Nutrients 2017, 9, 588. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.H.; Guo, D.S.; Lu, M.H.; Yue, J.Y.; Liu, Y.; Shang, C.M.; An, D.R.; Zhao, M.M. Inhibitory Effect of Osthole from Cnidium monnieri on Tobacco Mosaic Virus (TMV) Infection in Nicotiana glutinosa. Molecules 2019, 25, 65. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, Y.; Wang, C.; Wang, C.; Zhang, S.; Zhang, F. Fungicide selection in controlling potato blight. Guizhou Agric. Sci. 2014, 42, 43–45. [Google Scholar]
- Xue, Y.; Meng, M.; Hu, J.; Zhang, X.; Wang, X. Determination for Toxicity of Six Fungicides to Fusarium Wilt Pathogen of Potato in vitro. Chin. Potato J. 2012, 26, 228–230. [Google Scholar]
- Sun, P.P.; Jia, X.H.; Cui, J.C.; Tong, W.; Wang, W.H. Selection, Identification and Characterization of Actinomyces L-30 for theBiocontrol of Pear Gray Mold. Acta Hortic. Sin. 2016, 43, 2335–2346. [Google Scholar]
- Shan, H.Y.; Zhao, M.M.; Chen, D.X.; Chen, J.L.; Li, J.; Feng, Z.Z.; Ma, Z.Y.; An, D.R. Biocontrol of rice blast by the phenaminomethylacetic acid producer of Bacillus methylotrophicus strain BC79. Crop Prot. 2013, 44, 29–37. [Google Scholar] [CrossRef]
- Callahan, B.N.; Kammala, A.K.; Syed, M.; Yang, C.; Occhiuto, C.J.; Nellutla, R.; Chumanevich, A.P.; Oskeritzian, C.A.; Das, R.; Subramanian, H. Osthole, a Natural Plant Derivative Inhibits MRGPRX2 Induced Mast Cell Responses. Front. Immunol. 2020, 11, 703. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Qin, L.; He, W.; Van Puyvelde, L.; Maes, D.; Adams, A.; Zheng, H.; De Kimpe, N. Coumarins from Cnidium monnieri and their antiosteoporotic activity. Planta Medica 2007, 73, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Kwak, H.G.; Lim, H.B. Inhibitory effects of Cnidium monnieri fruit extract on pulmonary inflammation in mice induced by cigarette smoke condensate and lipopolysaccharide. Chin. J. Nat. Med. 2014, 12, 641–647. [Google Scholar] [CrossRef]
- Jing, C.L.; Zhao, J.; Han, X.B.; Huang, R.H.; Cai, D.S.; Zhang, C.S. Essential oil of Syringa oblata Lindl. as a potential biocontrol agent against tobacco brown spot caused by Alternaria alternata. Crop Prot. 2018, 104, 41–46. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Sharma, A.; Baek, K.H. Antibacterial mode of action of Cudrania tricuspidata fruit essential oil, affecting membrane permeability and surface characteristics of food-borne pathogens. Food Control 2013, 32, 582–590. [Google Scholar] [CrossRef]
- Zhou, H.; Tao, N.; Jia, L. Antifungal activity of citral, octanal and α-terpineol against Geotrichum citri-aurantii. Food Control 2014, 37, 277–283. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Total Spore Number | Germinated Spore Number | Germination Rate (%) | Inhibition Rate (%) |
|---|---|---|---|---|
| CK | 39.80 ± 2.64 | 13.5 ± 1.95 | 33.92 ± 2.05 | |
| osthole | 34.60 ± 4.51 | 5 ± 3.06 | 14.45 ± 4.27 | 57.40 ± 3.75 |
| Pathogen | Mycelial Growth Inhibition Rate (%) |
|---|---|
| F. oxysporum Schlecht | 55.34 ± 4.32 |
| F. moniliforme Sheld | 70.66 ± 1.47 * |
| T. cucumeris Donk | 75.90 ± 0.73 |
| F. Valsa mali Miyabeet Yamada | 90.36 ± 2.88 |
| A. alternata Keissler | 37.60 ± 4.05 * |
| A. solani Jonesetgrout | 37.05 ± 0.97 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, H.; Chen, Y.; Guo, Q.; Wei, H.; Yue, J.; Zhou, H.; Zhao, M. Inhibitory Effect of Osthole from Cnidium monnieri (L.) Cusson on Fusarium oxysporum, a Common Fungal Pathogen of Potato. Molecules 2021, 26, 3818. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26133818
Zheng H, Chen Y, Guo Q, Wei H, Yue J, Zhou H, Zhao M. Inhibitory Effect of Osthole from Cnidium monnieri (L.) Cusson on Fusarium oxysporum, a Common Fungal Pathogen of Potato. Molecules. 2021; 26(13):3818. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26133818
Chicago/Turabian StyleZheng, Hongli, Yahan Chen, Qiuli Guo, Hong Wei, Jianying Yue, Hongyou Zhou, and Mingmin Zhao. 2021. "Inhibitory Effect of Osthole from Cnidium monnieri (L.) Cusson on Fusarium oxysporum, a Common Fungal Pathogen of Potato" Molecules 26, no. 13: 3818. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26133818