
Identification of Bacteria Associated with Post-Operative Wounds of Patients with the Use of Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry Approach

 , , , and
, , , and 
Abstract
:1. Introduction
2. Results and Discussion
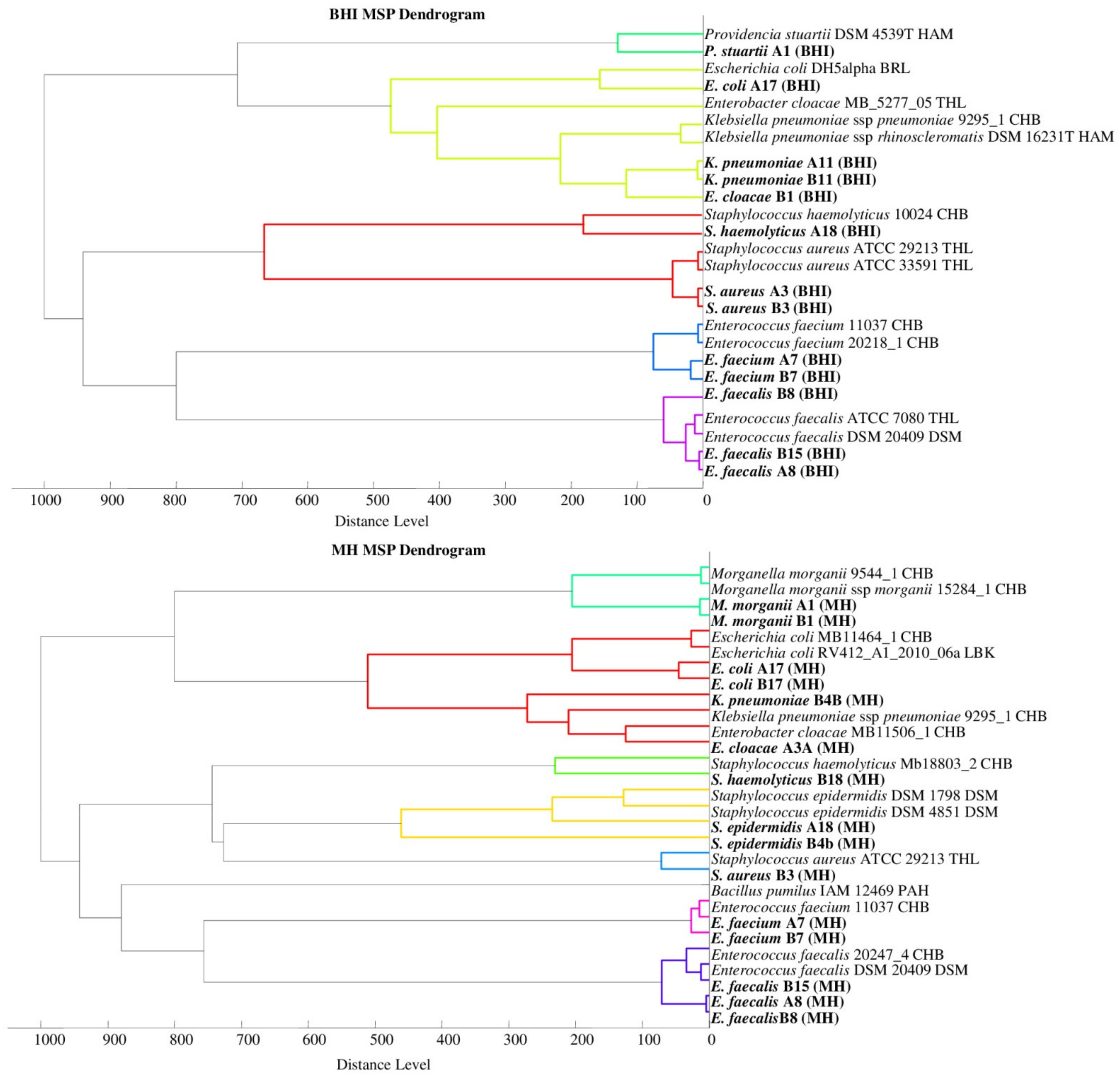
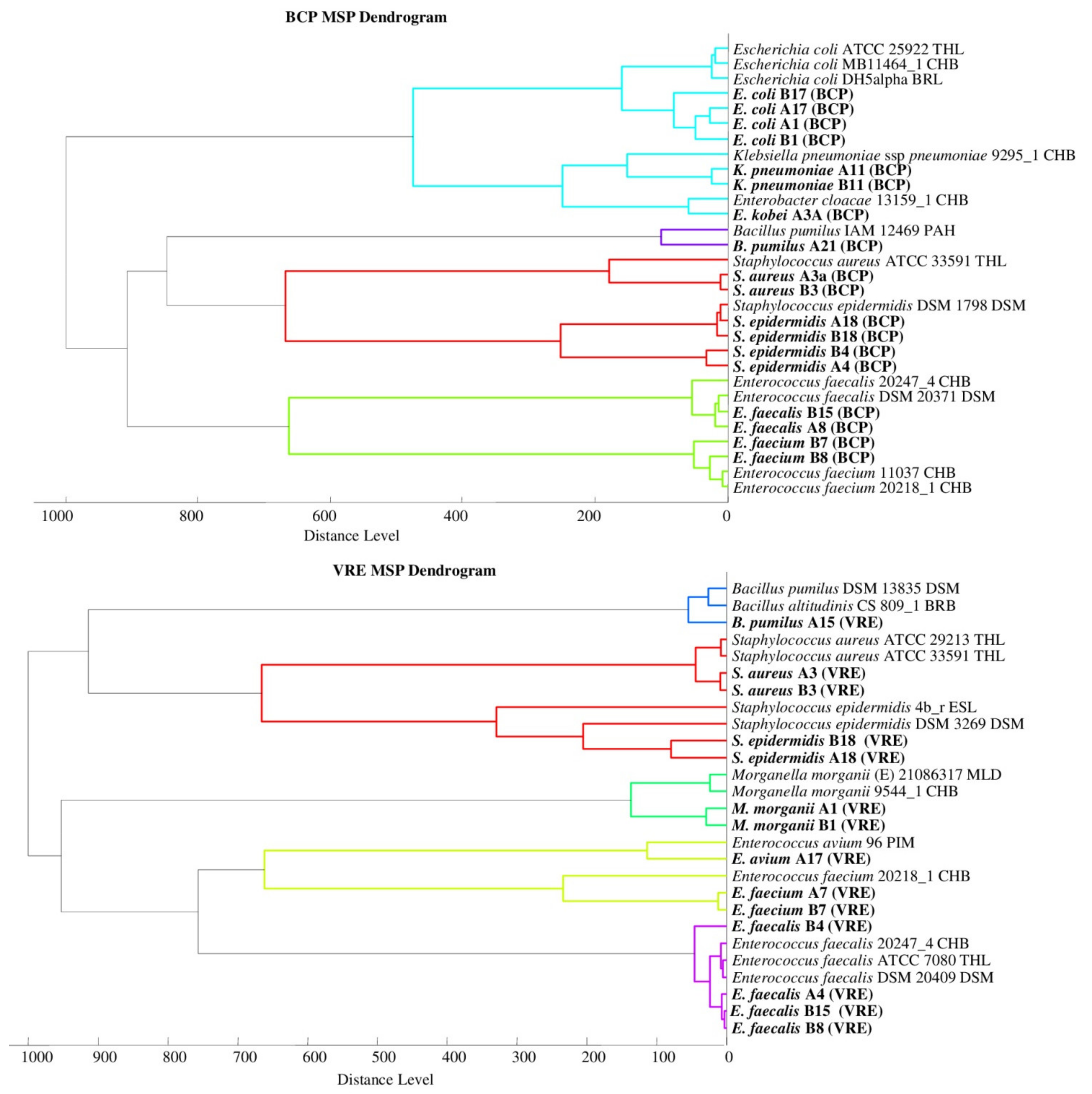
2.1. Bacterial Identification
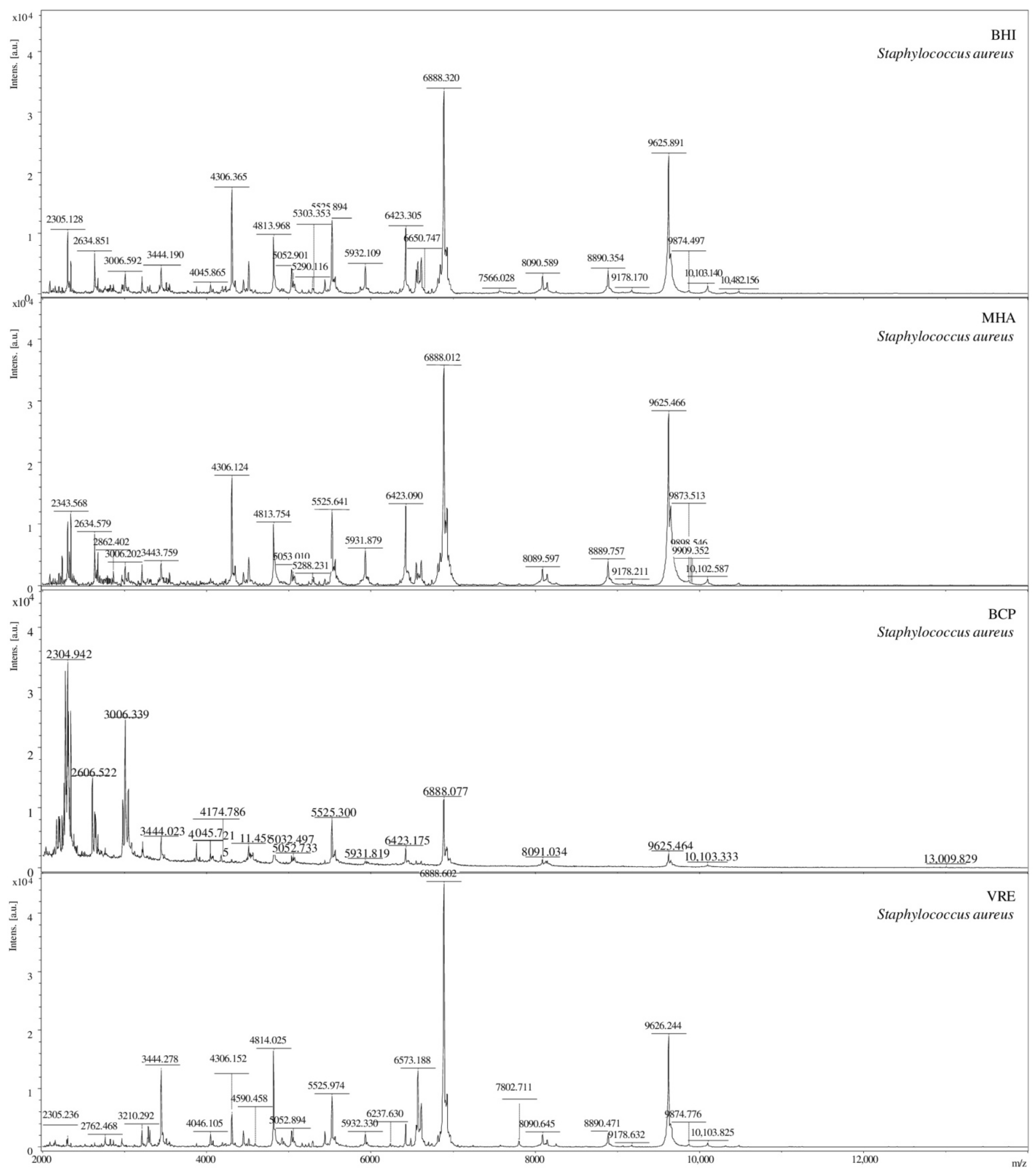
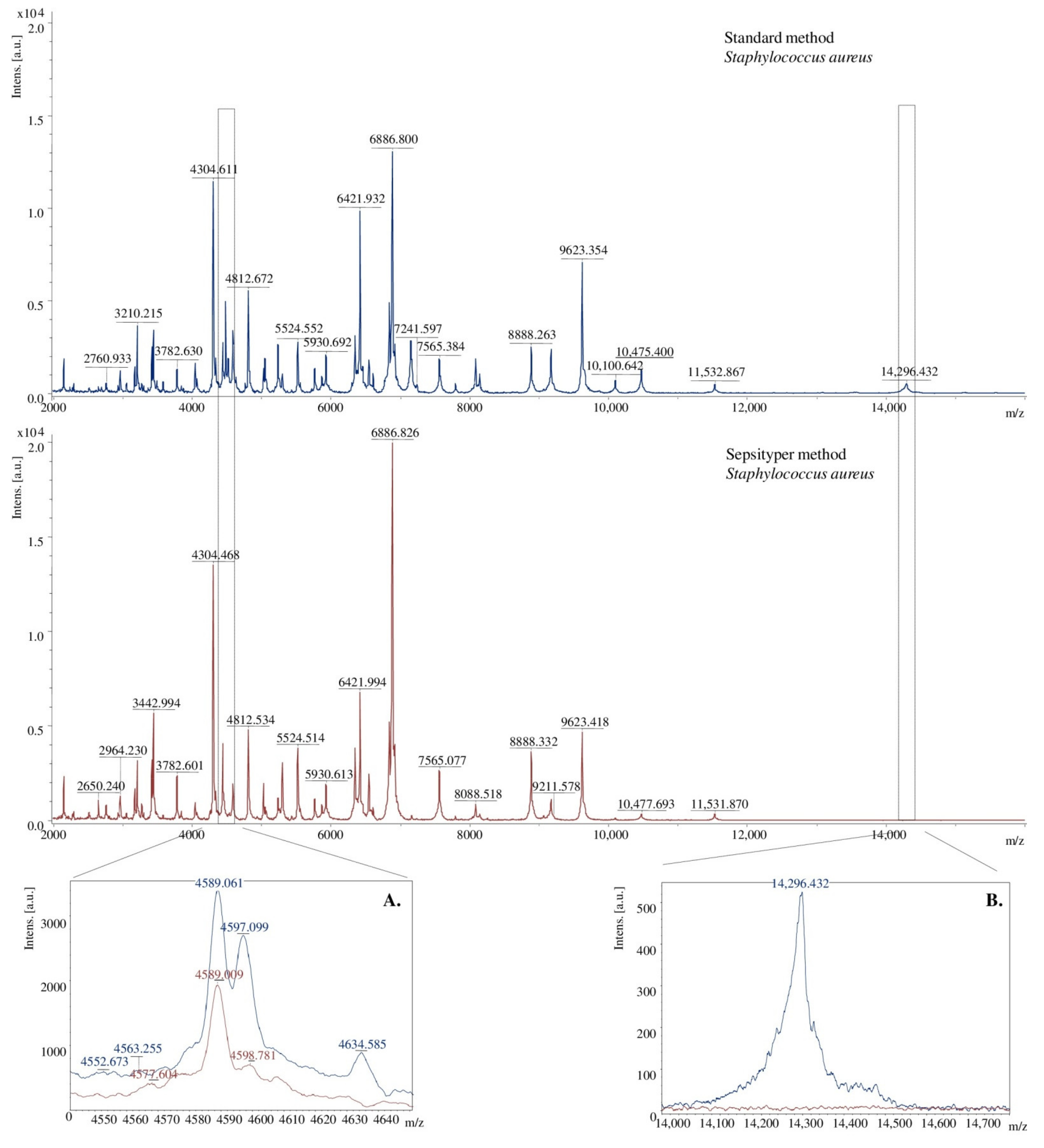
2.2. Mass Spectra Analysis
2.3. Identification of Proteins with Specific Peak to the Predicted Antibiotic Mechanism of Action and Bacterial Drug-Resistance
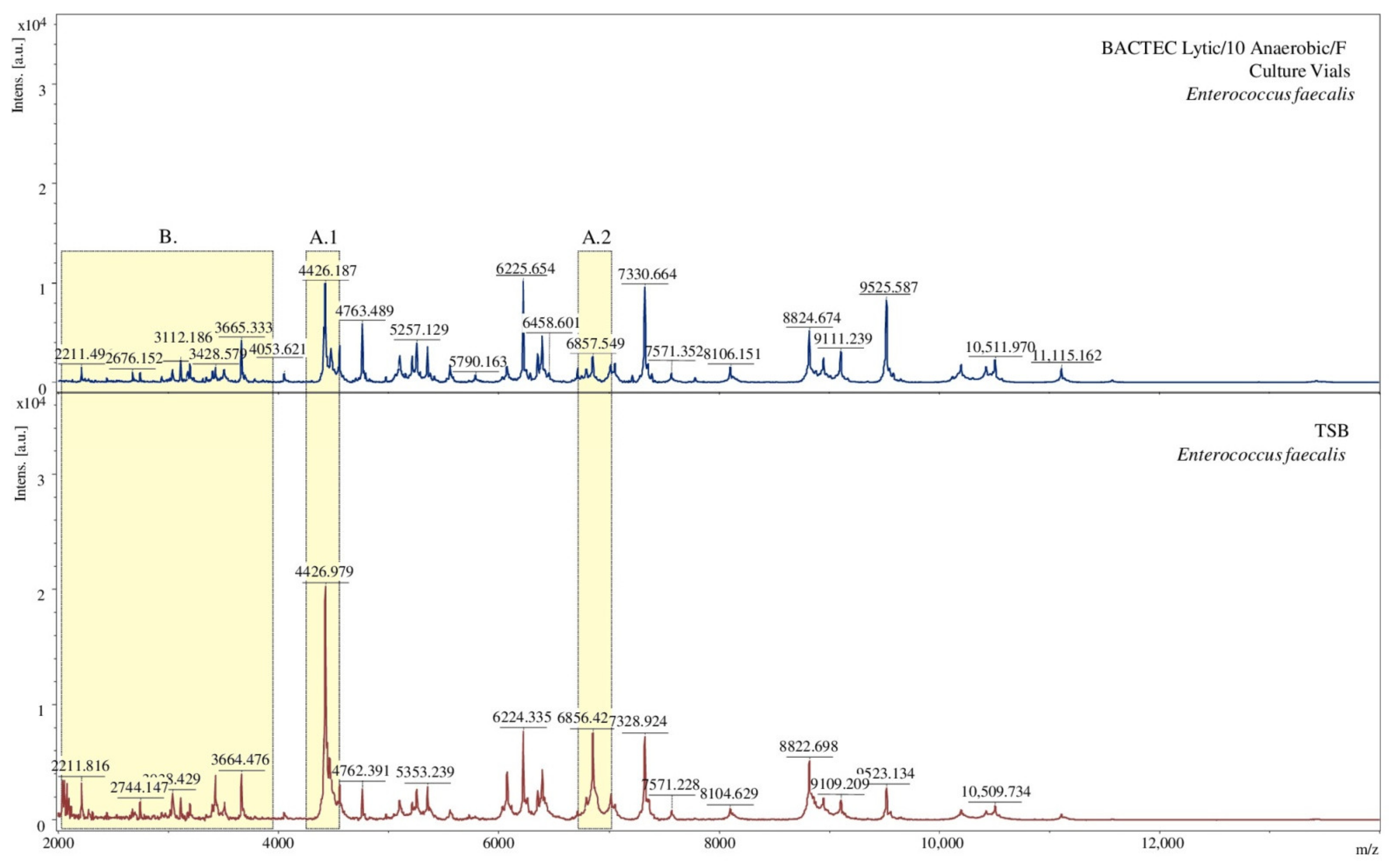
2.4. Selection of Sample Preparation Conditions to Determine Microorganisms Using the MALDI-TOF MS Technique
3. Experimental
3.1. Chemicals and Reagents
3.2. Patient and Sample Characterization
3.3. Isolation of Post-Operative Wound Bacteria
3.4. MALDI-TOF MS Analysis
3.5. Selection of Sample Preparation Conditions to Determine Microorganisms with MALDI-TOF MS
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Jackowski, M.; Szeliga, J.; Kłodzińska, E.; Buszewski, B. Application of capillary zone electrophoresis (CZE) to the determination of pathogenic bacteria for medical diagnosis. Anal. Bioanal. Chem. 2008, 391, 2153–2160. [Google Scholar] [CrossRef]
- Złoch, M.; Rodzik, A.; Pauter, K.; Szultka-Młynska, M.; Rogowska, A.; Kupczyk, W.; Buszewski, B. Problems with identifying and distinguishing salivary streptococci: A multi-instrumental approach. Future Microbiol. 2020, 15, 1157–1171. [Google Scholar] [CrossRef]
- Saffert, R.T.; Cunningham, S.A.; Ihde, S.M.; Jobe, K.E.; Mandrekar, J.; Patel, R. Comparison of Bruker Biotyper matrix-assisted laser desorption ionization-time of flight mass spectrometer to BD Phoenix automated microbiology system for identification of gram-negative bacilli. J. Clin. Microbiol. 2011, 49, 887–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, S.S.; Sercia, L.; Branda, J.A.; Burnham, C.A.; Bythow, M.; Ferraro, M.J.; Garner, O.B.; Ginocchio, C.C.; Jennemann, R.; Lewinski, M.A.; et al. Identification of Enterobacteriaceae by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry using the VITEK MS system. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1571–1578. [Google Scholar] [CrossRef] [PubMed]
- Kłodzińska, E.; Kupczyk, W.; Jackowski, M.; Buszewski, B. Capillary electrophoresis in the diagnosis of surgical site infections. Electrophoresis 2013, 34, 3206–3213. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.K.; Shukla, R.; Singh, P.; Kumar, R. Epidemiology and risk factors for surgical site infections in patients requiring orthopedic surgery. Eur. J. Orthop. Surg. Traumatol. 2015, 25, 251–254. [Google Scholar] [CrossRef]
- Pryor, K.O.; Fahey, T.J.; Lien, C.A.; Goldstein, P.A. Surgical Site Infection and the Routine Use of Perioperative Hyperoxia in a General Surgical Population: A Randomized Controlled Trial. J. Am. Med. Assoc. 2004, 291, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Gelaw, A.; Gebre-Selassie, S.; Tiruneh, M.; Mathios, E.; Yifru, S. Isolation of bacterial pathogens from patients with post-operative surgical site infections and possible sources of infections at the University of Gondar Hospital, Northwest Ethiopia. J. Environ. Occup. Sci. 2014, 3, 103–108. [Google Scholar] [CrossRef]
- Anguzu, J.R.; Olila, D. Drug sensitivity patterns of bacterial isolates from septic post-operative wounds in a regional referral hospital in Uganda. Afr. Health Sci. 2007, 7, 148–154. [Google Scholar] [CrossRef]
- Kramer, A.; Schwebke, I.; Kampf, G. How long do nosocomial pathogens persist on inanimate surfaces? A systematic review. BMC Infect. Dis. 2006, 6, 130. [Google Scholar] [CrossRef] [Green Version]
- Tahaesi, S.A.S.; Stájer, A.; Barrak, I.; Ostorházi, E.; Szabó, D.; Gajdács, M. Correlation between biofilm-formation and the antibiotic resistant phenotype in Staphylococcus aureus isolates: A laboratory-based study in Hungary and a review of the literature. Infect. Drug Resist. 2021, 14, 1155–1168. [Google Scholar] [CrossRef]
- Behzadi, P.; Baráth, Z.; Gajdács, M. It’s not easy being green: A narrative review on the microbiology virulence and therapeutic prospect of multidrug-resistant Pseudomonas aeruginosa. Antibiotics 2021, 10, 42. [Google Scholar] [CrossRef]
- Leoncini, E.; Ricciardi, W.; Cadoni, G.; Arzani, D.; Petrelli, L.; Paludetti, G. Adult height and head and neck cancer: A pooled analysis within the INHANCE Consortium. Eur. J. Epidemiol. 2014, 36, 1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajdács, M.; Urbán, E.; Stájer, A.; Baráth, Z. Antimicrobial resistance in the context of the sustainable development goals: A briev review. Eur. J. Investig. Health Psychol. Educ. 2021, 11, 71–82. [Google Scholar] [CrossRef]
- Goswami, N.N.; Trivedi, H.R.; Goswami, A.P.P.; Patel, T.K.; Tripathi, C.B. Antibiotic sensitivity profile of bacterial pathogens in post-operative wound infections at a tertiary care hospital in Gujarat, India. J. Pharmacol. Pharmacother. 2011, 2, 158–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauer, S.; Kliem, M. Mass spectrometry tools for the classification and identification of bacteria. Nat. Rev. Microbiol. 2010, 8, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Pomastowski, P.; Szultka, M.; Kupczyk, W.; Jackowski, M.; Buszewski, B. Evaluation of Intact Cell Matrix-Assisted Laser Desorption/Ionization Time of- Flight Mass Spectrometry for Capillary Electrophoresis Detection of Controlled Bacterial Clumping. J. Anal. Bioanal. Tech. 2015, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Mather, C.A.; Rivera, S.F.; Butler-Wu, S.M. Comparison of the Bruker Biotyper and Vitek MS matrix-assisted laser desorption ionization-time of flight mass spectrometry systems for identification of mycobacteria using simplified protein extraction protocols. J. Clin. Microbiol. 2014, 52, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Williams, T.L.; Andrzejewski, D.; Lay, J.O.; Musser, S.M. Experimental factors affecting the quality and reproducibility of MALDI TOF mass spectra obtained from whole bacteria cells. J. Am. Soc. Mass Spectrom. 2003, 14, 342–351. [Google Scholar] [CrossRef] [Green Version]
- Vargha, M.; Takáts, Z.; Konopka, A.; Nakatsu, C.H. Optimization of MALDI-TOF MS for strain level differentiation of Arthrobacter isolates. J. Microbiol. Methods 2006, 66, 399–409. [Google Scholar] [CrossRef]
- Becker, K.; Schubert, S. Editorial: MALDI-TOF MS Application for Susceptibility Testing of Microorganisms. Front. Microbiol. 2020, 11, 568891. [Google Scholar] [CrossRef] [PubMed]
- Szultka-Młyńska, M.; Pomastowski, P.; Buszewski, B. Application of solid phase microextraction followed by liquid chromatography-mass spectrometry in the determination of antibiotic drugs and their metabolites in human whole blood and tissue samples. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1086, 153–165. [Google Scholar] [CrossRef]
- Sharifzadeh, S.; Brown, N.W.; Shirley, J.D.; Bruce, K.E.; Winkler, M.E.; Carlson, E.E. Chemical Tools for Selective Activity Profiling of Bacterial Penicillin-Binding Proteins, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2020; Volume 638. [Google Scholar] [CrossRef]
- King, D.M.; Sobhanifar, S.; Strynadka, N.C.J. The mechanisms of resistance to β-lactam antibiotics. In Handbook of Antimicrobial Resistance; Gotte, M., Berghuis, A., Matlashewski, G., Sheppard, D., Wainberg, A., Eds.; Springer Science: New York, NY, USA, 2017; pp. 177–2017. [Google Scholar] [CrossRef]
- Champoux, J.J. DNA topoisomerases: Structure, function, and mechanism. Annu. Rev. Biochem. 2001, 70, 369–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacoby, G.A. Mechanisms of resistance to quinolones. Clin. Infect. Dis. 2005, 41, S120–S126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spížek, J.; Řezanka, T. Lincomycin, clindamycin and their applications. Appl. Microbiol. Biotechnol. 2004, 64, 455–464. [Google Scholar] [CrossRef]
- Abebe, M.; Tadesse, S.; Meseret, G.; Derbie, A. Type of bacterial isolates and antimicrobial resistance profile from different clinical samples at a Referral Hospital, Northwest Ethiopia: Five years data analysis. BMC Res. Notes 2019, 12, 568. [Google Scholar] [CrossRef]
- Saini, S.; Gupta, N.; Aparna, L.; Griwan, M.S. Surgical infections: A microbiological study. Braz. J. Infect. Dis. 2004, 8, 118–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Yang, H.; Chu, M.; Niu, X.; Huo, X.; Gao, Y. Klebsiella. In Beneficial Microbes in Agro-Ecology; Academic Press: Cambidge, MA, USA, 2020; pp. 233–257. [Google Scholar] [CrossRef]
- Cuzon, G.; Naas, T.; Fortineau, N.; Nordmann, P. Novel chromogenic medium for detection of vancomycin-resistant Enterococcus faecium and Enterococcus faecalis. J. Clin. Microbiol. 2008, 46, 2442–2444. [Google Scholar] [CrossRef] [Green Version]
- Gangania, P.S.; Singh, V.A.; Ghimire, S.S. Bacterial Isolation and Their Antibiotic Susceptibility Pattern from Post-Operative Wound Infected Patients. Indian J. Microbiol. Res. 2015, 2, 231–235. [Google Scholar] [CrossRef]
- Akinkunmi, E.O.; Adesunkanmi, A.-R.; Lamikanra, A. Pattern of pathogens from surgical wound infections in a Nigerian hospital and their antimicrobial susceptibility profiles. Afr. Health Sci. 2014, 14, 802–809. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Zhou, Y.; Liu, X.; Cao, Y.; Hu, C.; Chen, Y. Extension and application of a database for the rapid identification of Vibrio using MALDI-TOF MS. Acta Oceanol. Sin. 2020, 39, 140–146. [Google Scholar] [CrossRef]
- Body, B.A.; Beard, M.A.; Slechta, E.S.; Hanson, K.E.; Barker, A.P.; Babady, N.E. Evaluation of the Vitek MS v3.0 matrix-Assisted laser desorption ionization–Time of flight mass spectrometry system for identification of mycobacterium and nocardia species. J. Clin. Microbiol. 2018, 56, e00237-18. [Google Scholar] [CrossRef] [Green Version]
- Van Belkum, A.; Welker, M.; Pincus, D.; Charrier, J.P.; Girard, V. Matrix-assisted laser desorption ionization time-of-flight mass spectrometry in clinical microbiology: What are the current issues? Ann. Lab. Med. 2017, 37, 475–483. [Google Scholar] [CrossRef]
- Microbiology by numbers. Nat. Rev. Microbiol. 2011, 9, 628. [CrossRef]
- Grenfell, R.C.; da Silva Junior, A.R.; Del Negro, G.; Munhoz, R.B.; Gimenes, V.M.; Assis, D.M.; Rockstroh, A.C.; Motta, A.L.; Rossi, F.; Juliano, L.; et al. Identification of Candida haemulonii complex species: Use of ClinProToolsTM to Overcome Limitations of the Bruker BiotyperTM, VITEK MSTM IVD, and VITEK MSTM RUO databases. Front. Microbiol. 2016, 7, 940. [Google Scholar] [CrossRef]
- Fonseca, E.L.; Ramos, N.V.; Andrade, B.G.N.; Morais, L.L.C.S.; Marin, M.F.A.; Vicente, A.C.P. A one-step multiplex PCR to identify Klebsiella pneumoniae, Klebsiella variicola, and Klebsiella quasipneumoniae in the clinical routine. Diagn. Microbiol. Infect. Dis. 2017, 87, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Garner, O.; Mochon, A.; Branda, J.; Burnham, C.A.; Bythrow, M.; Ferraro, M. Multi-centre evaluation of mass spectrometric identification of anaerobic bacteria using the VITEK® MS system. Clin. Microbiol. Infect. 2014, 20, 335–339. [Google Scholar] [CrossRef] [Green Version]
- Anderson, N.W.; Buchan, B.W.; Riebe, K.M.; Parsons, L.N.; Gnacinski, S.; Ledeboer, N.A. Effects of solid-medium type on routine identification of bacterial isolates by use of matrix-assisted laser desorption ionization-time of flight mass spectrometry. J. Clin. Microbiol. 2012, 50, 1008–1013. [Google Scholar] [CrossRef] [Green Version]
- Buskirk, A.D.; Hettick, J.M.; Chipinda, I.; Law, B.F.; Siegel, P.D.; Slaven, J.E. Fungal pigments inhibit the matrix-assisted laser desorption/ionization time-of-flight mass spectrometry analysis of darkly pigmented fungi. Anal. Biochem. 2011, 411, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Złoch, M.; Pomastowski, P.; Maślak, E.; Monedeiro, F.; Buszewski, B. Study on Molecular Profiles of Staphylococcus aureus Strains: Spectrometric Approach. Molecules 2020, 25, 4894. [Google Scholar] [CrossRef] [PubMed]
- Paradis, S.; Boissinot, M.; Paquette, N.; Bélanger, S.D.; Martel, E.A.; Boudreau, D.K. Phylogeny of the Enterobacteriaceae based on genes encoding elongation factor Tu and F-ATPase β-subunit. Int. J. Syst. Evol. Microbiol. 2005, 55, 2013–2025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, K.L.; Godfrey, P.; Griggs, A.; Kos, V.N.; Zucker, J.; Desjardins, C. Comparative genomics of enterococci: Variation in Enterococcus faecalis, clade structure in E. faecium, and defining characteristics of E. gallinarum and E. casseliflavus. MBio 2012, 3, e00318-11. [Google Scholar] [CrossRef] [Green Version]
- Queck, S.Y.; Khan, B.A.; Wang, R.; Bach, T.H.L.; Kretschmer, D.; Chen, L. Mobile genetic element-encoded cytolysin connects virulence to methicillin resistance in MRSA. PLoS Pathog. 2009, 5, e1000533. [Google Scholar] [CrossRef]
- Chatterjee, S.S.; Chen, L.; Joo, H.S.; Cheung, G.Y.C.; Kreiswirth, B.N.; Otto, M. Distribution and regulation of the mobile genetic element-encoded phenol-soluble modulin PSM-mec in methicillin-resistant Staphylococcus aureus. PLoS ONE 2011, 6, e28781. [Google Scholar] [CrossRef] [PubMed]
- Blankschien, M.D.; Potrykus, K.; Grace, E.; Choudhary, A.; Vinella, D.; Cashel, M. TraR, a homolog of a RNAP secondary channel interactor, modulates transcription. PLoS Genet. 2009, 5, e1000345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroda, M.; Ohta, T.; Uchiyama, I.; Baba, T.; Yuzawa, H.; Kobayashi, I.; Cui, L.; Oguchi, A.; Aoki, K.; Nagi, Y.; et al. Whole genome seqencing of meticillin-resistant Staphylococcus aureus. Lancet 2001, 357, 1225–1240. [Google Scholar] [CrossRef]
- Hensel, M.; Shea, J.E.; Gleeson, C.; Jones, M.D.; Dalton, E.; Holden, D.W. Simultaneous identification of bacterial virulence genes by negative selection. Science 1995, 269, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.L.; Dassa, E.; Orelle, C.; Chen, J. Structure, function and evolution of bacterial ATP-binding cassete system. Microbiol. Mol. Biol. Rev. 2008, 72, 317–364. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.H.; Guan, H.H.; Chen, C.J.; Huang, C.Y. Staphylococcus aureus single-stranded DNA-binding protein SsbA can bind but cannot stimulate PriA helicase. PLoS ONE 2017, 12, e0182060. [Google Scholar] [CrossRef] [Green Version]
- Hassan, K.A.; Skurray, R.A.; Brown, M.H. Active ecport proteins mediating drug resistance in staphylococci. J. Mol. Microbiol. Biotechnol. 2007, 12, 180–196. [Google Scholar] [CrossRef]
- Vetting, M.W.; Luiz, L.P.; Yu, M.; Hegde, S.S.; Magnet, S.; Roderick, S.L. Structure and functions of the GNAT superfamily of acetyltransferases. Arch. Biochem. Biophys. 2005, 433, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Palmqvist, N.; Foster, T.; Tarkowski, A.; Josefsson, E. Protein A is a virulence factor in Staphylococcus aureus arthritis and septic death. Microb. Pathog. 2002, 33, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, I.T.; Banerjei, L.; Myers, G.S.A.; Nelson, K.E.; Seshadri, R.; Read, T.D.; Fouts, D.E.; Eisen, J.A.; Gill, S.R.; Heidelberg, J.F.; et al. Role of mobile DNA in the evolution of vancomycin-resistant Enterococcus faecalis. Science 2003, 299, 2071–2074. [Google Scholar] [CrossRef] [Green Version]
- Tsakou, F.; Jersie-Christensen, R.; Jenssen, H.; Mojsoska, B. The role of proteomics in bacterial response to antibiotics. Pharmaceuticals 2020, 13, 214. [Google Scholar] [CrossRef] [PubMed]
- McCurtain, J.L.; Gilbertsen, A.J.; Evert, C.; Williams, B.J.; Hunter, R.C. Agmatine accumulation by Pseudomonas aeruginosa clinical isolates confers antibiotic tolerance and dampens host inflammation. J. Med. Microbiol. 2019, 68, 446–455. [Google Scholar] [CrossRef]
- Venkateswara Prasad, U.; Vasu, D.; Nada Kumar, Y.; Santhosh Kumar, P.; Yeswanth, S.; Swarupa, V.; Phaneendra, B.V.; Chaudhary, A.; Sarma, P.V.G.K. Cloning, expression and characterization of NADP-dependent isocitrate dehydrogenase vfrom Staphylococcus aureus. Appl. Biochem. Biotechnol. 2013, 169, 862–869. [Google Scholar] [CrossRef] [PubMed]
- Ostash, B.; Walker, S. Bacterial transglycosylase inhibitors. Curr. Opin. Chem. Biol. 2005, 9, 459–466. [Google Scholar] [CrossRef]
- Thongsomboon, W.; Serra, D.O.; Possling, A.; Hadjineophytou, C.; Hengge, R.; Cegelski, L. Phosphoetanolamine cellulose: A naturally produced chemically modified cellulose. Science 2018, 338, 334–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, M.F.; Woodgate, R. Translesion DNA polymerases. Cold Spring Harb. Perspect. Biol. 2015, 4, a010363. [Google Scholar] [CrossRef] [PubMed]
- Funnell, B.E. ParB partition proteins: Complex formation and spreading at bacterial and plasmid centromeres. Front. Mol. Biosci. 2016, 3, 44. [Google Scholar] [CrossRef] [Green Version]
- Ueta, M.; Ohniwa, R.L.; Yoshida, H.; Maki, Y.; Wada, C.; Wada, A. Role of HPF (hibernation promoting factor) in translational activity in Escherichia coli. J. Biochem. 2008, 143, 425–433. [Google Scholar] [CrossRef]
- Li, M.; Diep, B.A.; Villaruz, A.E.; Braughton, K.R.; Jiang, X.; DeLeo, F.R. Evolution of virulence in epidemic community-associated methicillin-resistant Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2009, 106, 5883–5888. [Google Scholar] [CrossRef] [Green Version]
- Talha, M.H.; Khazaal, S.S.; Al Hadraawy, M.K.; Mostafavi, S.K.S. Screening of antibiotic resistance genes and virulence determinants of Staphylococcus aureus from skin infections. Meta Gene 2020, 24, 100682. [Google Scholar] [CrossRef]
- Periasamy, S.; Joo, H.S.; Duong, A.C.; Bach, T.H.L.; Tan, V.Y.; Chatterjee, S.S. How Staphylococcus aureus biofilms develop their characteristic structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1281–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dersch, P.; Khan, M.A.; Mühlen, S.; Görke, B. Roles of regulatory RNAs for antibiotic resistance in bacteria and their potential value as novel drug targets. Front. Microbiol. 2017, 8, 803. [Google Scholar] [CrossRef] [Green Version]
- Pauter, K.; Szultka-Młyńska, M.; Buszewski, B. Determination and Identification of Antibiotic Drugs and Bacterial Strains in Biological Samples. Molecules 2020, 25, 2556. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.Y.; Cai, J.C.; Zhou, H.W.; Zhang, R.; Chen, G.-X. Rapid detection of porins by matrix-assisted laser desorption/ionization-time of flight mass spectrometry. Front. Microbiol. 2015, 6, 784. [Google Scholar] [CrossRef]
- Sun, X.; Jia, H.L.; Xiao, C.L.; Yin, X.F.; Yang, X.Y.; Lu, J. Bacterial proteome of streptococcus pneumoniae through multidimensional separations coupled with LC-MS/MS. Omi. A J. Integr. Biol. 2011, 15, 477–482. [Google Scholar] [CrossRef]
- Drevinek, M.; Dresler, J.; Klimentova, J.; Pisa, L.; Hubalek, M. Evaluation of sample preparation methods for MALDI-TOF MS identification of highly dangerous bacteria. Lett. Appl. Microbiol. 2012, 55, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Burckhardt, I.; Last, K.; Zimmermann, S. Shorter incubation times for detecting multi-drug resistant bacteria in patient samples: Defining early imaging time points using growth kinetics and total laboratory automation. Ann. Lab. Med. 2018, 39, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Van den Bijllaardt, W.; Buitingm, A.G.; Mouton, J.W.; Muller, A.E. Shortening the incubation time for antimicrobial susceptibility testing by disk diffusion for Enterobacteriaceae: How short can it be and are the results accurate? Int. J. Antimicrob. Agents 2017, 49, 631–637. [Google Scholar] [CrossRef]
- Altun, O.; Botero-Kleiven, S.; Carlsson, S.; Ullberg, M.; Özenci, V. Rapid identification of bacteria from positive blood culture bottles by MALDI-TOF MS following short-term incubation on solid media. J. Med. Microbiol. 2015, 64, 1346–1352. [Google Scholar] [CrossRef] [PubMed]
- Oviaño, M.; Rodríguez-Sánchez, B.; Gómara, M.; Alcalá, L.; Zvezdanova, E.; Ruíz, A. Direct identification of clinical pathogens from liquid culture media by MALDI-TOF MS analysis. Clin. Microbiol. Infect. 2018, 24, 624–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haiko, J.; Savolainen, L.E.; Hilla, R.; Pätäri-Sampo, A. Identification of urinary tract pathogens after 3-hours urine culture by MALDI-TOF mass spectrometry. J. Microbiol. Methods 2016, 129, 81–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajdács, M.; Ábrók, M.; Lázár, A.; Terhes, G.; Urbán, E. Anaerobic blood culture positivity at a University Hospital in Hungary: A 5-year comparative retrospective study. Anaerobe 2020, 63, 102200. [Google Scholar] [CrossRef]
- Sanders, E.R. Aseptic laboratory techniques: Plating methods. JoVE 2012, 63, e3064. [Google Scholar] [CrossRef]
- Pomastowski, P.; Złoch, M.; Rodzik, A.; Ligor, M.; Kostrzewa, M.; Buszewski, B. Analysis of bacteria associated with honeys of different geographical and botanical origin using two different identification approaches: MALDI-TOF MS and 16S rDNA PCR technique. PLoS ONE 2019, 14, e0217078. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Selective Growth Media | |||||||||
| BHI | MH | ||||||||
| Sample Name | RAW | MSP | Sample Name | RAW | MSP | ||||
| Best Match | Score Value | Best Match | Score Value | Best Match | Score Value | Best Match | Score Value | ||
| B1a | Providencia stuartii DSM 4539T HAM | 2.10 | Providencia stuartii DSM 4539T HAM | 1.99 | B1a | Morganella morganii ssp morganii 15284_1 CHB | 2.04 | Morganella morganii ssp morganii 15284_1 CHB | 2.05 |
| B1b | Enterobacter cloacae MB_5277_05 THL | 1.91 | Enterobacter cloacae MB_5277_05 THL | 1.78 | B1b | Morganella morganii 9544_1 CHB | 1.93 | Morganella morganii 9544_1 CHB | 1.93 |
| B3a | Staphylococcus aureus ATCC 25923 THL | 2.35 | Staphylococcus aureus ATCC 33591 THL | 2.26 | B3aD | Enterobacter cloacae MB_5277_05 THL | 2.03 | Enterobacter cloacae MB11506_1 CHB | 1.99 |
| B3b | Staphylococcus aureus ATCC 25923 THL | 2.19 | Staphylococcus aureus ATCC 29213 THL | 1.99 | B3aM | Staphylococcus aureus ATCC 33591 THL | 2.15 | Staphylococcus aureus ATCC 29213 THL | 2.12 |
| B4a | Staphylococcus epidermidis 10547 CHB | 2.28 | - | 1.69 | B3b | Staphylococcus aureus ATCC 33591 THL | 2.23 | Staphylococcus aureus ATCC 29213 THL | 2.19 |
| B4b | Staphylococcus epidermidis ATCC 14990T THL | 1.90 | - | 1.53 | B4a | Staphylococcus epidermidis 10547 CHB | 1.99 | - | 1.51 |
| B7a | Enterococcus faecium 20218_1 CHB | 2.14 | Enterococcus faecium 20218_1 CHB | 2.15 | B4bD | Klebsiella pneumoniae ssp pneumoniae DSM 30104T HAM | 2.13 | Klebsiella pneumoniae ssp pneumoniae 9295_1 CHB | 1.90 |
| B7b | Enterococcus faecium 11037 CHB | 2.09 | Enterococcus faecium 11037 CHB | 2.09 | B4bM | Staphylococcus epidermidis 0547 CHB | 1.98 | Staphylococcus epidermidis DSM 4851 DSM | 1.74 |
| B8a | Enterococcus faecalis ATCC 7080 THL | 2.19 | Enterococcus faecalis ATCC 7080 THL | 2.14 | B7a | Enterococcus faecium 11037 CHB | 2.25 | Enterococcus faecium 11037 CHB | 2.25 |
| B8b | Enterococcus faecalis DSM 2570 DSM | 2.18 | Enterococcus faecalis DSM 20409 DSM | 1.91 | B7b | Enterococcus faecium 11037 CHB | 2.35 | Enterococcus faecium 11037 CHB | 2.29 |
| B9a | - | 1.45 | - | 1.39 | B8a | Enterococcus faecalis DSM 20409 DSM | 2.10 | Enterococcus faecalis DSM 20409 DSM | 2.03 |
| B11a | Klebsiella pneumoniae ssp ozaenae DSM 16358T HAM | 2.15 | Klebsiella pneumoniae ssp rhinoscleromatis DSM 16231T HAM | 2.06 | B8b | Enterococcus faecalis ATCC 7080 THL | 2.12 | Enterococcus faecalis 20247_4 CHB | 1.91 |
| B11b | Klebsiella pneumoniae ssp pneumoniae DSM 30104T HAM | 2.07 | Klebsiella pneumoniae ssp pneumoniae 9295_1 CHB | 2.04 | B11b | Klebsiella pneumoniae ssp pneumoniae 9295_1 CHB | 2.32 | Klebsiella pneumoniae ssp pneumoniae 9295_1 CHB | 2.32 |
| B15a | - | 1.39 | - | 1.33 | B15a | Bacillus pumilus IAM 12469 PAH | 1.74 | - | 1.64 |
| B15b | Enterococcus faecalis ATCC 7080 THL | 2.30 | Enterococcus faecalis DSM 20409 DSM | 2.26 | B15b | Enterococcus faecalis ATCC 7080 THL | 2.43 | Enterococcus faecalis DSM 20409 DSM | 2.35 |
| B17a | Escherichia coli ATCC 25922 THL | 2.25 | Escherichia coli DH5alpha BRL | 2.18 | B17a | Escherichia coli MB11464_1 CHB | 1.99 | Escherichia coli MB11464_1 CHB | 1.88 |
| B18a | Staphylococcus haemolyticus 10024 CHB | 2.23 | Staphylococcus haemolyticus 10024 CHB | 1.97 | B17b | Escherichia coli RV412_A1_2010_06a LBK | 1.95 | Escherichia coli RV412_A1_2010_06a LBK | 1.95 |
| B18b | Staphylococcus epidermidis CCM 4505 CCM | 2.10 | - | 1.68 | B18a | Staphylococcus epidermidis DSM 1798 DSM | 1.85 | Staphylococcus epidermidis DSM 1798 DSM | 1.89 |
| B21a | Bacillus pumilus IAM 12469 PAH | 1.77 | - | 1.68 | B18b | Staphylococcus haemolyticus Mb18803_2 CHB | 2.01 | Staphylococcus haemolyticus Mb18803_2 CHB | 1.93 |
| B21a (white) | Lactobacillus pentosus DSM 20199 DSM | 1.82 | - | 1.21 | B21a | - | 1.68 | - | 1.59 |
| Selective Growth Media | |||||||||
| BCP | VRE | ||||||||
| Sample Name | RAW | MSP | Sample name | RAW | MSP | ||||
| Best Match | Score Value | Best Match | Score Value | Best Match | Score Value | Best Match | Score Value | ||
| B1a | Escherichia coli ATCC 25922 THL | 2.22 | Escherichia coli ATCC 25922 THL | 2.09 | B1a | Morganella morgani 9544_1 CHB | 1.96 | Morganella morganii 9544_1 CHB | 1.80 |
| B1b | Escherichia coli MB11464_1 CHB | 2.15 | Escherichia coli MB11464_1 CHB | 2.10 | B1b | Morganella morgani (E) 21086317 MLD | 1.99 | Morganella morganii (E) 21086317 MLD | 1.89 |
| B3aD | Enterobacter kobei DSM 13645T DSM | 2.20 | Enterobacter cloacae 13159_1 CHB | 2.08 | B3a | Staphylococcus aureus ATCC 29213 THL | 2.47 | Staphylococcus aureus ATCC 29213 THL | 2.21 |
| B3aM | Staphylococcus aureus ATCC 33591 THL | 2.03 | Staphylococcus aureus ATCC 33591 THL | 1.88 | B3b | Staphylococcus aureus ATCC 29213 THL | 2.47 | Staphylococcus aureus ATCC 33591 THL | 2.25 |
| B3b | Staphylococcus aureus ATCC 33591 THL | 1.95 | Staphylococcus aureus ATCC 33591 THL | 1.84 | B4a | Enterococcus faecalis ATCC 7080 THL | 2.34 | Enterococcus faecalis DSM 20409 DSM | 2.33 |
| B4a | Staphylococcus epidermidis ATCC 14990T THL | 2.04 | Staphylococcus epidermidis DSM 1798 DSM | 1.84 | B4b | Enterococcus faecalis ATCC 7080 THL | 2.06 | Enterococcus faecalis ATCC 7080 THL | 1.83 |
| B4b | Staphylococcus epidermidis ATCC 14990T THL | 2.14 | Staphylococcus epidermidis DSM 1798 DSM | 1.84 | B7a | Enterococcus faecium 20218_1 CHB | 1.97 | Enterococcus faecium 20218_1 CHB | 1.95 |
| B7a | Enterococcus faecium 11037 CHB | 2.19 | Enterococcus faecium 20218_1 CHB | 2.15 | B7b | Enterococcus faecium 20218_1 CHB | 1.95 | Enterococcus faecium 20218_1 CHB | 1.95 |
| B7b | Enterococcus faecium 11037 CHB | 2.05 | Enterococcus faecium 20218_1 CHB | 2.09 | B8b | Enterococcus faecalis ATCC 7080 THL | 2.20 | Enterococcus faecalis ATCC 7080 THL | 2.15 |
| B8a | Enterococcus faecalis DSM 20371 DSM | 1.98 | Enterococcus faecalis DSM 20371 DSM | 1.90 | B15a | Bacillus pumilus DSM 13835 DSM | 2.09 | Bacillus pumilus DSM 13835 DSM | 2.09 |
| B8b | Enterococcus faecium 11037 CHB | 2.32 | Enterococcus faecium 11037 CHB | 2.38 | B15b | Enterococcus faecalis ATCC 29212 CHB | 2.20 | Enterococcus faecalis 20247_4 CHB | 2.11 |
| B11a | Klebsiella pneumoniae ssp pneumoniae 9295_1 CHB | 2.22 | Klebsiella pneumoniae ssp pneumoniae 9295_1 CHB | 2.17 | B17a | Enterococcus avium 96 PIM | 1.87 | Enterococcus avium 96 PIM | 1.87 |
| B11b | Klebsiella pneumoniae ssp pneumoniae 9295_1 CHB | 2.10 | Klebsiella pneumoniae ssp pneumoniae 9295_1 CHB | 2.10 | B18a | Staphylococcus epidermidis 10547 CHB | 2.16 | Staphylococcus epidermidis DSM 3269 DSM | 1.92 |
| B15a | - | 1.61 | - | 1.61 | B18b | Staphylococcus epidermidis 10547 CHB | 1.92 | Staphylococcus epidermidis 4b_r ESL | 1.87 |
| B15b | Enterococcus faecalis 20247_4 CHB | 2.29 | Enterococcus faecalis 20247_4 CHB | 2.18 | B21a | Bacillus altitudis CS 809_1 BRB | 1.72 | - | 1.57 |
| B17a | Escherichia coli DH5alphaBRL | 2.19 | Escherichia coli DH5alpha BRL | 2.01 | |||||
| B17b | Escherichia coli DH5alpha BRL | 2.08 | Escherichia coli MB11464_1 CHB | 1.84 | |||||
| B18a | Staphylococcus epidermidis 10547 CHB | 2.08 | Staphylococcus epidermidis DSM 1798 DSM | 1.94 | |||||
| B18b | Staphylococcus epidermidis DSM 1798 DSM | 2.00 | Staphylococcus epidermidis DSM 1798 DSM | 2.13 | |||||
| B21a | Bacillus pumilus IAM 12469 PAH | 1.73 | - | 1.68 | |||||
| Bacteria from Biological Material | Reference Bacteria Strain |
|---|---|
| Bacillus | |
| B. altitudis VRE.B21a | B. altitudis CS 809_1 BRB |
| B. pumilus BCP.B21a | B. pumilus IAM 12469 PAH |
| B. pumilus VRE.B15a | B. pumilus DSM 13835 DSM |
| Enterobacter | |
| E. cloacae BHI.B1b | E. cloacae MB_5277_05 THL |
| E. kobei BCP.B3aD | E. kobei DSM 13645T DSM |
| Enterococcus | |
| E. avium VRE.B17a | E. avium 96 PIM |
| E. faecalis BCP.B8a | E. faecalis DSM 20371 DSM |
| E. faecalis BCP.B15b | E. faecalis 20247_4 CHB |
| E. faecalis BHI.B15b | E. faecalis ATCC 7080 THL |
| E. faecalis BHI.B8b | E. faecalis DSM 2570 DSM |
| E. faecalis VRE.B15b | E. faecalis ATCC 29212 CHB |
| E. faecalis MHA.B8a | E. faecalis DSM 20409 DSM |
| E. faecium BHI.B7b | E. faecium 11037 CHB |
| E. faecium BHI.B7a | E. faecium 20218_1 CHB |
| Escherichia | |
| E. coli BHI.B17a | E. coli ATCC 25922 THL |
| E. coli BCP.B1b | E. coli MB11464_1 CHB |
| E. coli BCP.B17a | E. coli DH5alpha BRL |
| E. coli MHA.B17a | E. coli RV412_A1_2010_06a LBK |
| Klebsiella | |
| K. pneumoniae BCP.B11a | K. pneumoniae ssp pneumoniae 9295_1 CHB |
| K. pneumoniae BHI.B11a | K. pneumoniae ssp ozaenae DSM 16358T HAM |
| K.pneumoniae BHI.B11b | K.pneumoniae ssp pneumoniae DSM 30104T HAM |
| Lactobacillus | |
| L. pentosus BHI.B21a(white) | L. pentosus DSM 20199 DSM |
| Morganella | |
| M. morgani MHA.B1b | M. morgani 9544_1 CHB |
| M. morgani VRE.B1b | M. morgani (E) 21086317 MLD |
| M. morganii MHA.B1a | M. morganii ssp morganii 15284_1 CHB |
| Providencia | |
| P. stuartii BHI.B1a | P. stuartii DSM 4539T HAM |
| Staphylococcus | |
| S. aureus BCP.B3aM | S. aureus ATCC_33591_THL |
| S. aureus BHI.B3b | S. aureus ATCC_25923_THL |
| S. epidermidis BCP.B4a | S. epidermidis ATCC 14990T THL |
| S. epidermidis BHI.B4a | S. epidermidis 10547 CHB |
| S. epidermidis BCP.B18b | S. epidermidis DSM 1798 DSM |
| S. epidermidis BHI. B18b | S. epidermidis CCM 4505 CCM |
| S. haemolyticus BHI.B18a | S. haemolyticus 10024 CHB |
| S. haemolyticus MHA.B18b | S. haemolyticus Mb18803_2 CHB |
| S. aureus /ATCC 29213 THL/(Antibiotic: Clindamycin) | |||||
| m/z | Control Sample | Test Sample | Protein | Putative Function | Ref. |
| 2656.8 | - | + | Phenol-soluble modulin PSM-alpha-3 | virulence factor | [46,47] |
| 3006.2 | - | + | TraR/DksA family transcriptional regulator | virulence, antibiotic resistance, pathogenesis | [48] |
| 4290.7 | + | - | 50S ribosomal protein L36 | builds ribosome, translation | [49] |
| 4365.1 | + | - | ABC transporter ATP-binding protein | [50,51] | |
| 4379.9 | + | - | Teichoic acid ABC transporter permease | [50,51] | |
| 4386.1 | + | - | Single-stranded DNA-binding protein | DNA replication | [52] |
| 4822.6 | + | - | Oligopeptide ABC superfamily ATP binding cassette transporter. binding protein (Protein CysJ) | [50,51] | |
| 5437.4 | + | - | 50S ribosomal protein L33. | builds ribosome, translation | [49] |
| 5872.7 | + | - | 50S ribosomal protein L33 | builds ribosome, translation | [49] |
| 6355.1 | - | + | Antibiotic transport-associated protein | [53] | |
| 6589.8 | - | + | Antibiotic resistance protein | [53] | |
| 6743.1 | - | + | Exported protein | [53] | |
| 7022.9 | + | - | ABC transporter ATP-binding protein | [50,51] | |
| 8111.7 | - | + | GCN5-related N-acetyltransferase (GNAT) | kanamycin and gentamycin resistance | [54] |
| 8908.0 | + | - | Compound ABC uptake transporter ATP-binding protein. | [50,51] | |
| 9898.5 | - | + | Immunoglobulin G binding protein A | binding protein A | [55] |
| 9909.4 | - | + | Protein A | pathogenic factor | [55] |
| E. faecalis /ATCC 7080 THL/(antibiotics: piperacylin and tazobactam) | |||||
| m/z | Control sample | Test sample | Protein | Putative Function | |
| 2443.3 | ‒ | + | Attenuator leader peptide | response to antibiotic | [56] |
| 6341.0 | + | ‒ | 50S ribosomal protein L30 | builds ribosome, translation | [57] |
| E. cloacae /MB_5277_05 THL/(antibiotic: clindamycin) | |||||
| m/z | Control sample | Test sample | Protein | Putative Function | |
| 3710.7 | ‒ | + | Agmatinase | catalysis of the reaction: agmatine + H2O = putrescine + urea | [58] |
| 5571.8 | + | ‒ | Isocitrate dehydrogenase (NADP(+)) | cofactor binding sites Mg2+, Mn2+; | [59] |
| 5598.8 | ‒ | + | Agmatinase | catalysis of the reaction: agmatine + H2O = putrescine + urea | [58] |
| 6417.9 | ‒ | + | Murein transglycosylase | break the glycosidic bond | [60] |
| 7620.8 | ‒ | + | Cellulose biosynthesis protein BcsF | necessary for biofilm formation | [61] |
| 9410.2 | ‒ | + | DNA polymerase V | part of the SOS response to DNA damage | [62] |
| 10,315.8 | + | ‒ | ParB partition protein | regulation of transcription | [63] |
| 10,883.9 | ‒ | + | Ribosome hibernation promoting factor HPF | primary metabolic process, regulation of translation | [64] |
| Bacteria Species | Time | Standard Method | Sepsityper Method | ||
|---|---|---|---|---|---|
| Vials | TSB | Vials | TSB | ||
| S. aureus | 4 h | 1.85–1.97 | 1.79–1.88 | 2.05–2.16 | |
| 6 h | 2.38–2.42 | 2.31–2.39 | 2.36–2.39 | 2.25–2.30 | |
| 24 h | 2.25–2.34 | 2.24–2.33 | 2.30–2.34 | 2.31–2.42 | |
| E. coli | 4 h | - | 2.35–2.37 | - | 2.31–2.38 |
| 6 h | - | 2.31–2.33 | - | 2.20–2.28 | |
| 24 h | - | 2.27–2.32 | - | 2.10–2.17 | |
| E. faecalis | 4 h | 2.33–2.35 | 2.26–2.36 | 2.28–2.32 | 2.06–2.18 |
| 6 h | 2.00–2.02 | 1.76–1.88 | 2.02–2.06 | ||
| 24 h | 2.39–2.42 | 2.45–2.47 | 2.19–2.24 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szultka-Młyńska, M.; Janiszewska, D.; Pomastowski, P.; Złoch, M.; Kupczyk, W.; Buszewski, B. Identification of Bacteria Associated with Post-Operative Wounds of Patients with the Use of Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry Approach. Molecules 2021, 26, 5007. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26165007
Szultka-Młyńska M, Janiszewska D, Pomastowski P, Złoch M, Kupczyk W, Buszewski B. Identification of Bacteria Associated with Post-Operative Wounds of Patients with the Use of Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry Approach. Molecules. 2021; 26(16):5007. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26165007
Chicago/Turabian StyleSzultka-Młyńska, Małgorzata, Daria Janiszewska, Paweł Pomastowski, Michał Złoch, Wojciech Kupczyk, and Bogusław Buszewski. 2021. "Identification of Bacteria Associated with Post-Operative Wounds of Patients with the Use of Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry Approach" Molecules 26, no. 16: 5007. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26165007