
The Zebrafish Embryo as a Model to Test Protective Effects of Food Antioxidant Compounds

 , ,
, ,
Abstract
:1. Introduction
2. Results
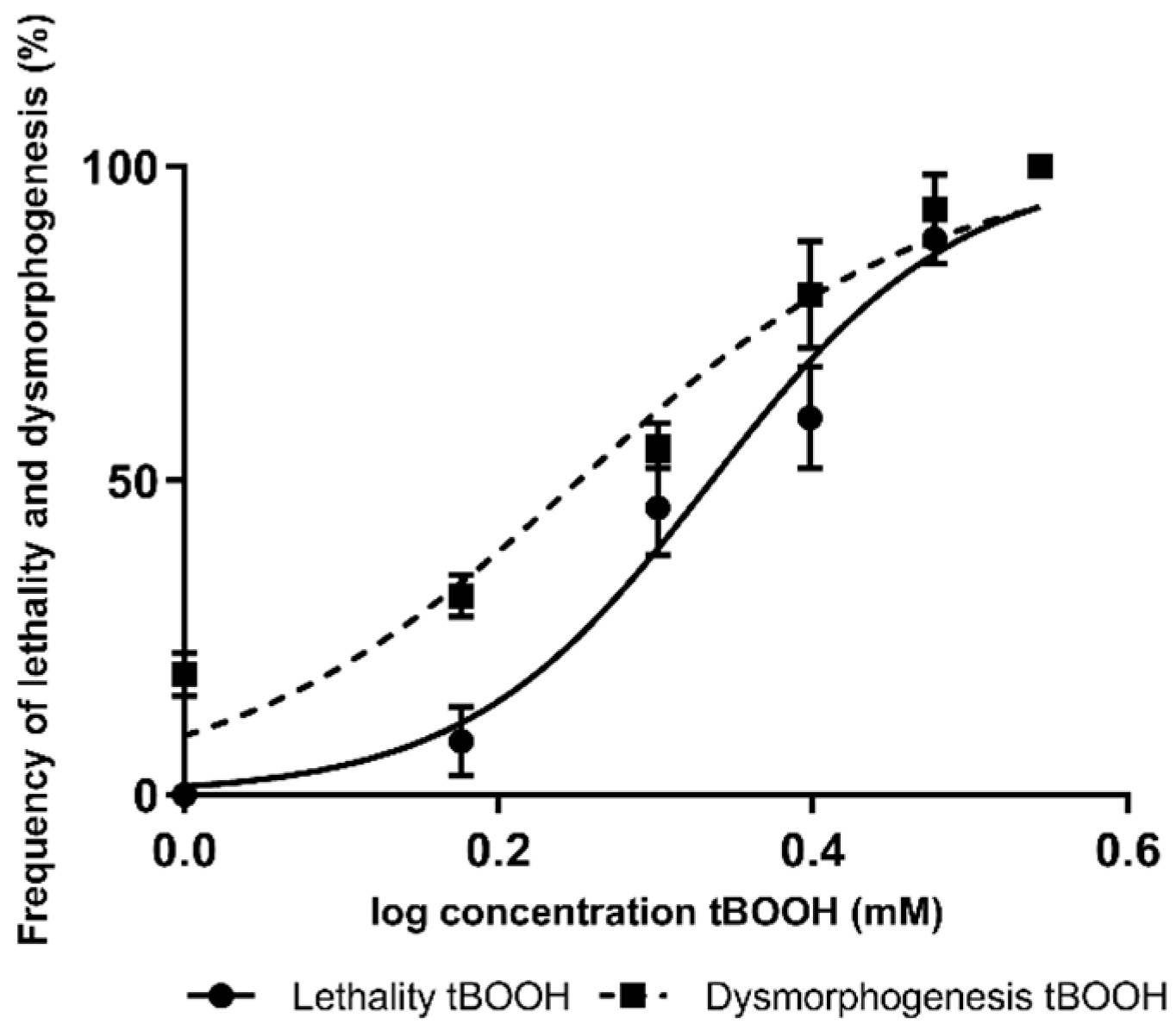
2.1. Concentration Effect Curves for tBOOH in Zebrafish Embryos
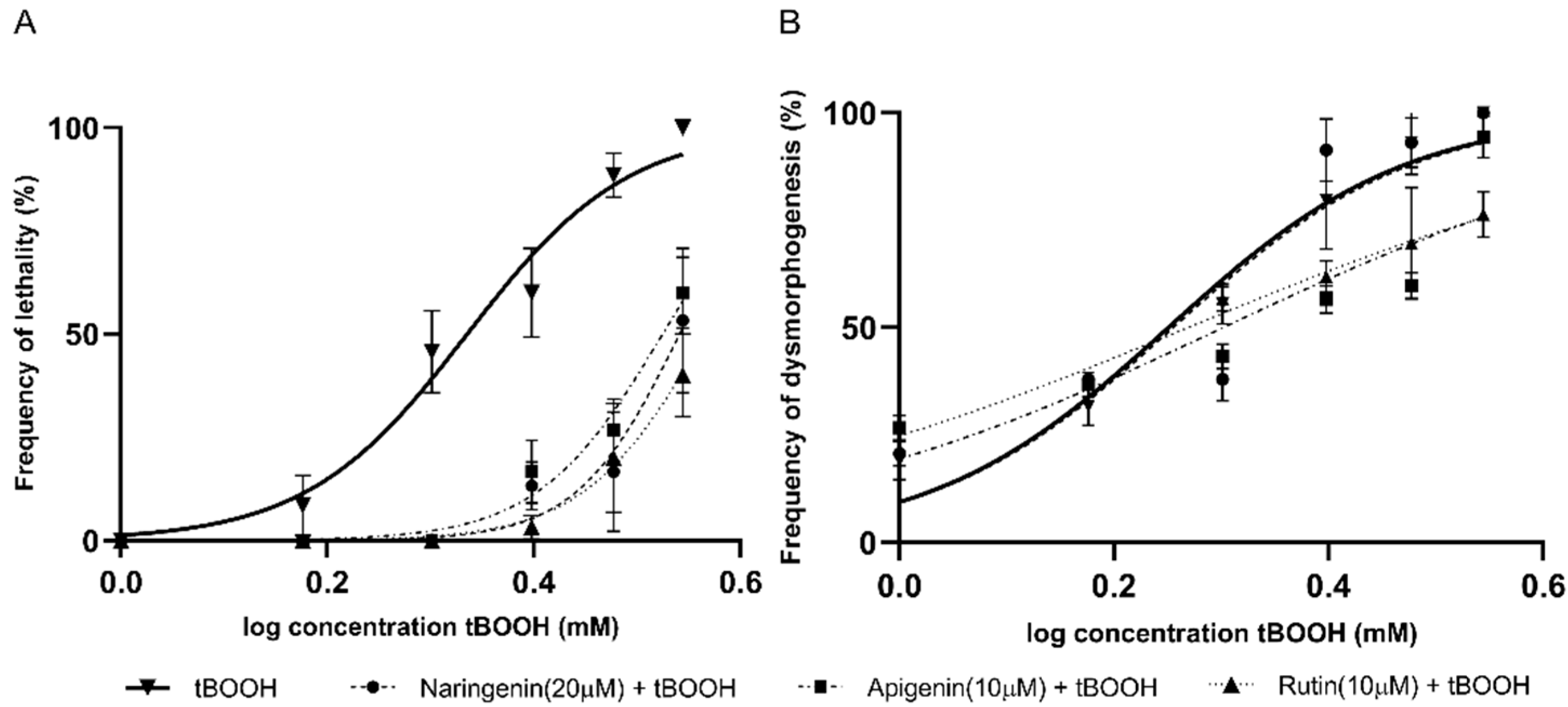
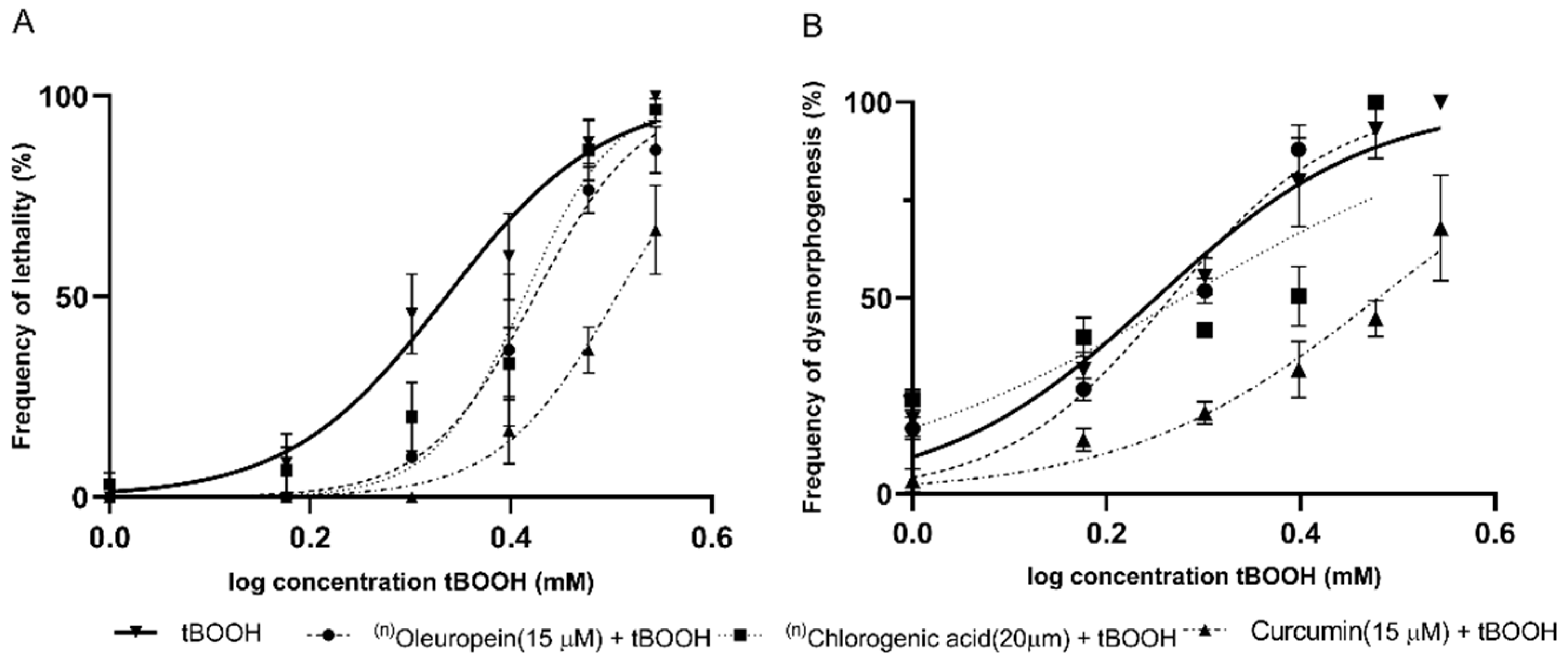
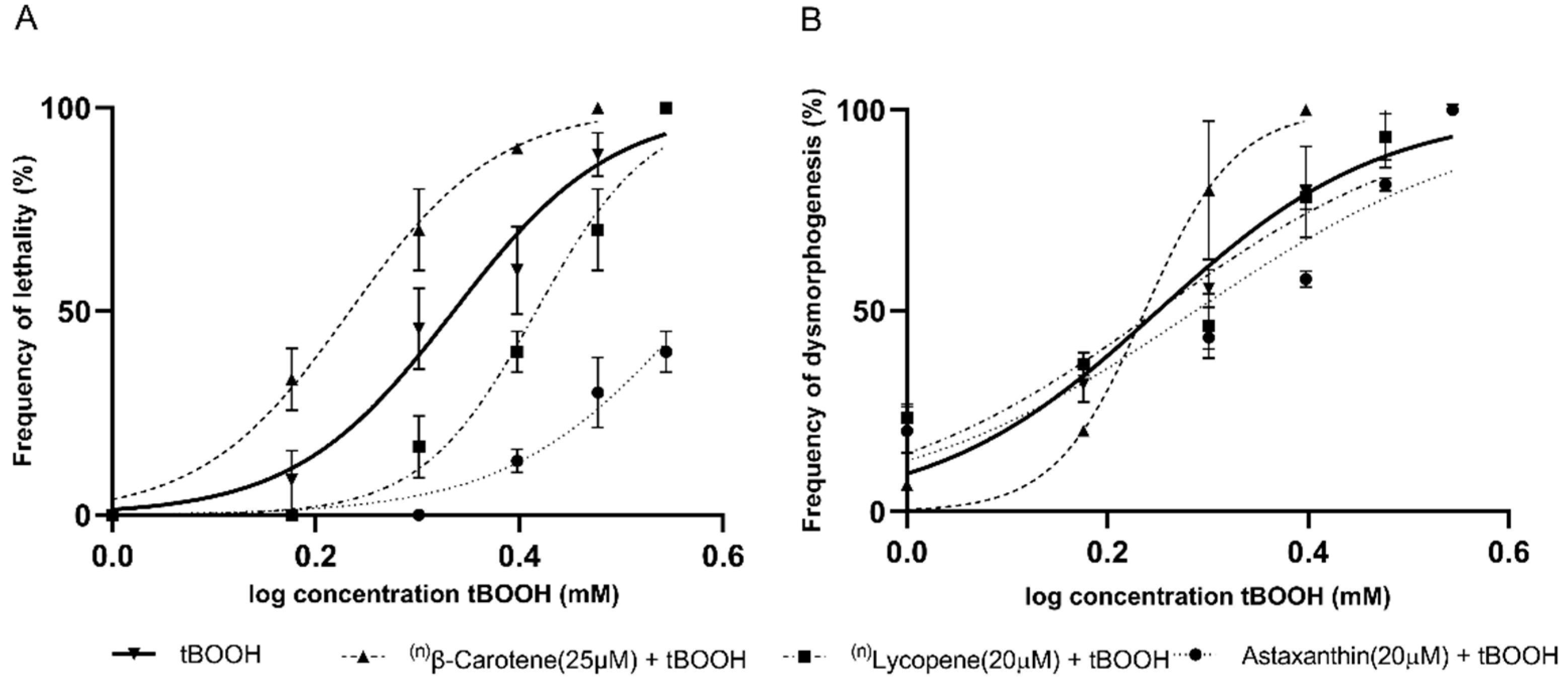
2.2. Identification of the Protective Effects of Antioxidant Compounds in Zebrafish Embryos
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Chemicals and Solution Preparation
4.3. Zebrafish Maintenance and Egg Production
4.4. Exposure of Zebrafish Embryos to Oxidative Stress (Tert-Butyl Hydroperoxide)
4.5. Determination of the Protective Effects of Antioxidant Compounds
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tan, B.L.; Norhaizan, M.E.; Liew, W.-P.-P. Nutrients and Oxidative Stress: Friend or Foe? Oxid. Med. Cell. Longev. 2018, 2018, 9719584. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Biochemistry of Oxidative Stress. Angew. Chemie Int. Ed. Engl. 1986, 25, 1058–1071. [Google Scholar] [CrossRef]
- Dröge, W. Free Radicals in the Physiological Control of Cell Function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef] [PubMed]
- Rendra, E.; Riabov, V.; Mossel, D.M.; Sevastyanova, T.; Harmsen, M.C.; Kzhyshkowska, J. Reactive oxygen species (ROS) in macrophage activation and function in diabetes. Immunobiology 2019, 224, 242–253. [Google Scholar] [CrossRef]
- Lin, M.T.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 2006, 443, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Therond, P. Oxidative stress and damages to biomolecules (lipids, proteins, DNA). Ann. Pharm. Fr. 2006, 64, 383–389. [Google Scholar] [CrossRef]
- Ermak, G.; Davies, K.J.A. Calcium and oxidative stress: From cell signaling to cell death. Mol. Immunol. 2002, 38, 713–721. [Google Scholar] [CrossRef]
- Wadhwa, R.; Gupta, R.; Maurya, P.K. Oxidative Stress and Accelerated Aging in Neurodegenerative and Neuropsychiatric Disorder. Curr. Pharm. Des. 2018, 24, 4711–4725. [Google Scholar] [CrossRef]
- Maritim, A.C.; Sanders, R.A.; Watkins, J.B. Diabetes, oxidative stress, and antioxidants: A review. J. Biochem. Mol. Toxicol. 2003, 17, 24–38. [Google Scholar] [CrossRef]
- Sinha, N.; Dabla, P.K. Oxidative stress and antioxidants in hypertension-a current review. Curr. Hypertens. Rev. 2015, 11, 132–142. [Google Scholar] [CrossRef]
- Klaunig, J.E. Oxidative Stress and Cancer. Curr. Pharm. Des. 2018, 24, 4771–4778. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef] [Green Version]
- Pietta, P.-G. Flavonoids as Antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 5th ed.; Oxford University Press: Oxford, UK, 2015; ISBN 9780198717478. [Google Scholar]
- Hollman, P.C.; Katan, M.B. Dietary flavonoids: Intake, health effects and bioavailability. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 1999, 37, 937–942. [Google Scholar] [CrossRef]
- Hertog, M.G.; Hollman, P.C.; Katan, M.B.; Kromhout, D. Intake of potentially anticarcinogenic flavonoids and their determinants in adults in The Netherlands. Nutr. Cancer 1993, 20, 21–29. [Google Scholar] [CrossRef]
- Martínez, S.; González, J.; Culebras, J.; Tuñón, M. Flavonoides: Propiedades y acciones antioxidantes. Nutr. Hosp. 2002, 17, 271–278. [Google Scholar]
- Arteaga, C.; Bustillos, A.; Gómez, J. Migración de neutrófilos en larvas de pez cebra expuestos a extractos de sofrito de tomate. Arch. Latinoam. Nutr. 2021, 70, 216–224. [Google Scholar] [CrossRef]
- Young, A.J.; Lowe, G.L. Carotenoids—Antioxidant properties. Antioxidants 2018, 7, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xavier, A.A.O.; Pérez-Gálvez, A. Carotenoids as a Source of Antioxidants in the Diet. Subcell. Biochem. 2016, 79, 359–375. [Google Scholar] [CrossRef] [PubMed]
- Stahl, W.; Sies, H. Antioxidant activity of carotenoids. Mol. Asp. Med. 2003, 24, 345–351. [Google Scholar] [CrossRef]
- Boix, N.; Teixido, E.; Pique, E.; Llobet, J.M.; Gomez-Catalan, J. Modulation and Protection Effects of Antioxidant Compounds against Oxidant Induced Developmental Toxicity in Zebrafish. Antioxidants 2020, 9, 721. [Google Scholar] [CrossRef]
- Fenton, H.J.H. Oxidation of tartaric acid in presence of iron. J. Chem. Soc. Trans. 1894, 65, 899–910. [Google Scholar] [CrossRef] [Green Version]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free Radicals: Properties, Sources, Targets, and Their Implication in Various Diseases. Indian J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Pachón, M.S.; Villaño, D.; Troncoso, A.M.; García-Parrilla, M.C. Revisión de los métodos de evaluación de la actividad antioxidante in vitro del vino y valoración de sus efectos in vivo. Arch. Latinoam. Nutr. 2006, 56, 110–122. [Google Scholar] [PubMed]
- Cavia-Saiz, M.; Busto, M.D.; Pilar-Izquierdo, M.C.; Ortega, N.; Perez-Mateos, M.; Muñiz, P. Antioxidant properties, radical scavenging activity and biomolecule protection capacity of flavonoid naringenin and its glycoside naringin: A comparative study. J. Sci. Food Agric. 2010, 90, 1238–1244. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.; Singh, G.K.; Patel, D.K. A review on pharmacological and analytical aspects of naringenin. Chin. J. Integr. Med. 2014, 24, 551–560. [Google Scholar] [CrossRef]
- Rashmi, R.; Magesh, S.B.; Ramkumar, K.M.; Suryanarayanan, S.; SubbaRao, M.V. Antioxidant potential of naringenin helps to protect liver tissue from streptozotocin-induced damage. Rep. Biochem. Mol. Biol. 2017, 7, 76–84. [Google Scholar]
- Shukla, R.; Pandey, V.; Vadnere, G.P.; Lodhi, S. Chapter 18—Role of Flavonoids in Management of Inflammatory Disorders. In Bioactive Food as Dietary Interventions for Arthritis and Related Inflammatory Diseases; Watson, R.R., Preedy, V.R., Eds.; Academic Press: London, UK, 2019; pp. 293–322. ISBN 978-0-12-813820-5. [Google Scholar]
- Chen, Y.H.; Yang, Z.S.; Wen, C.C.; Chang, Y.S.; Wang, B.C.; Hsiao, C.A.; Shih, T.L. Evaluation of the structure-activity relationship of flavonoids as antioxidants and toxicants of zebrafish larvae. Food Chem. 2012, 134, 717–724. [Google Scholar] [CrossRef]
- Cushman, M.; Zhu, H.; Geahlen, R.L.; Kraker, A.J. Synthesis and Biochemical Evaluation of a Series of Aminoflavones as Potential Inhibitors of Protein-Tyrosine Kinases p56lck, EGFr, and p60v-src. J. Med. Chem. 1994, 37, 3353–3362. [Google Scholar] [CrossRef]
- Cioffi, G.; Pesca, M.S.; De Caprariis, P.; Braca, A.; Severino, L.; De Tommasi, N. Phenolic compounds in olive oil and olive pomace from Cilento (Campania, Italy) and their antioxidant activity. Food Chem. 2010, 121, 105–111. [Google Scholar] [CrossRef]
- Bulotta, S.; Corradino, R.; Celano, M.; D’Agostino, M.; Maiuolo, J.; Oliverio, M.; Procopio, A.; Iannone, M.; Rotiroti, D.; Russo, D. Antiproliferative and antioxidant effects on breast cancer cells of oleuropein and its semisynthetic peracetylated derivatives. Food Chem. 2011, 127, 1609–1614. [Google Scholar] [CrossRef]
- Han, J.; Talorete, T.P.N.; Yamada, P.; Isoda, H. Anti-proliferative and apoptotic effects of oleuropein and hydroxytyrosol on human breast cancer MCF-7 cells. Cytotechnology 2009, 59, 45–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, N.; Kitts, D.D. Role of chlorogenic acids in controlling oxidative and inflammatory stress conditions. Nutrients 2015, 8, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.-Y.; Freedman, N.D.; Haiman, C.A.; Le Marchand, L.; Wilkens, L.R.; Setiawan, V.W. Association of Coffee Consumption With Total and Cause-Specific Mortality Among Nonwhite Populations. Ann. Intern. Med. 2017, 167, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Tajik, N.; Tajik, M.; Mack, I.; Enck, P. The potential effects of chlorogenic acid, the main phenolic components in coffee, on health: A comprehensive review of the literature. Eur. J. Nutr. 2017, 56, 2215–2244. [Google Scholar] [CrossRef] [PubMed]
- Poole, R.; Kennedy, O.J.; Roderick, P.; Fallowfield, J.A.; Hayes, P.C.; Parkes, J. Coffee consumption and health: Umbrella review of meta-analyses of multiple health outcomes. BMJ 2017, 359, j5024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanvir, E.M.; Hossen, M.S.; Hossain, M.F.; Afroz, R.; Gan, S.H.; Khalil, M.I.; Karim, N. Antioxidant Properties of Popular Turmeric (Curcuma longa) Varieties from Bangladesh. J. Food Qual. 2017, 2017, 8471785. [Google Scholar] [CrossRef] [Green Version]
- Fiedor, J.; Burda, K. Potential role of carotenoids as antioxidants in human health and disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [Green Version]
- Eggersdorfer, M.; Wyss, A. Carotenoids in human nutrition and health. Arch. Biochem. Biophys. 2018, 652, 18–26. [Google Scholar] [CrossRef]
- Padmanabhan, P.; Cheema, A.; Paliyath, G. Solanaceous Fruits Including Tomato, Eggplant, and Peppers, 1st ed.; Elsevier Ltd: London, UK, 2015. [Google Scholar]
- Kataja-Tuomola, M.K.; Kontto, J.P.; Männistö, S.; Albanes, D.; Virtamo, J.R. Effect of alpha-tocopherol and beta-carotene supplementation on macrovascular complications and total mortality from diabetes: Results of the ATBC Study. Ann. Med. 2010, 42, 178–186. [Google Scholar] [CrossRef]
- Ambati, R.R.; Moi, P.S.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications—A review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef] [PubMed]
- Dennery, P.A. Effects of oxidative stress on embryonic development. Birth Defects Res. C Embryo Today 2007, 81, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Selderslaghs, I.W.T.; Blust, R.; Witters, H.E. Feasibility study of the zebrafish assay as an alternative method to screen for developmental toxicity and embryotoxicity using a training set of 27 compounds. Reprod. Toxicol. 2012, 33, 142–154. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Teixidó, E.; Piqué, E.; Gómez-Catalán, J.; Llobet, J.M. Assessment of developmental delay in the zebrafish embryo teratogenicity assay. Toxicol. In Vitro 2013, 27, 469–478. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Lethality | Dysmorphogenesis | ||||||
|---|---|---|---|---|---|---|---|---|
| LC50 (mM) tBOOH (95% CI) | LC50 (mM) (Compound + tBOOH) (95% CI) | p-Value | Effect | EC50 (mM) tBOOH (95% CI) | EC50 (mM) (Compound + tBOOH) (95% CI) | p-Value | Effect | |
| Naringenin (20 μM) | 2.1 (2.0–2.2) | 3.4 (3.3–3.7) | <0.0001 | PE 1 | 1.7 (1.6–1.8) | 1.8 (1.5–2.0) | >0.05 | S/E 2 |
| Apigenin (10 μM) | 2.1 (2.0–2.2) | 3.3 (3.2–3.4) | <0.0001 | PE | 1.7 (1.6–1.8) | 2.0 (1.7–2.2) | 0.0006 | PE |
| Rutin (10 μM) | 2.1 (2.0–2.2) | 3.6 (3.5–3.9) | <0.0001 | PE | 1.7 (1.6–1.8) | 1.8 (1.7–2.0) | <0.0008 | PE |
| Oleuropein (15 μM) | 2.1 (2.0–2.2) | 2.6 (2.5–2.7) | <0.0001 | PE | 1.7 (1.6–1.8) | 1.8 (1.7–1.9) | >0.05 | S/E |
| Chlorogenic acid (20 μM) | 2.1 (2.0–2.2) | 2.5 (2.4–2.6) | 0.0028 | PE | 1.7 (1.6–1.8) | 1.8 (1.5–2.2) | >0.05 | S/E |
| Curcumin (15 μM) | 2.1 (2.0–2.2) | 3.2 (3.1–3.2) | <0.0001 | PE | 1.7 (1.6–1.8) | 3.0 (2.8–3.2) | <0.0001 | PE |
| Lycopene (20 μM) | 2.1 (2.0–2.2) | 2.6 (2.5–2.6) | 0.0003 | PE | 1.7 (1.6–1.8) | 1.7 (1.5–1.9) | 0.56 | S/E |
| β-carotene (25 μM) | 2.1 (2.0–2.2) | 1.7 (1.6–1.7) | 0.0001 | IL 3 | 1.7 (1.6–1.8) | 1.5 (1.5–1.6) | 0.01 | ID 4 |
| Astaxanthin (20 μM) | 2.1 (2.0–2.2) | 3.7 (3.5–3.9) | <0.0001 | PE | 1.7 (1.6–1.8) | 1.9 (1.7–2.1) | 0.0506 | S/E |
| Morphological Features | Morphological Abnormality | Example of Observed Characteristics. |
|---|---|---|
| Detachment of the tail; Tail | No tail, malformation of chorda or spinal cord. Tail necrosis, bent tail. |  |
| Optic system; Otic system; Brain | Abnormal pigmentation, asymmetric eyes. Formation of no, one, or more than two otoliths per sacculus. Brain necrosis, hemorrhage. |  |
| Heart | Pericardial edema, big heart, hemorrhage, abnormal chambers. |  |
| Head-body pigmentation; Tail pigmentation | Lack of pigmentation in the tail or body. |  |
| Movement | Spasms, abnormal movements, no movement at all. | The ZF embryo was touched and its movement was evaluated. |
| * Control | No signs of dysmorphogenesis. |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arteaga, C.; Boix, N.; Teixido, E.; Marizande, F.; Cadena, S.; Bustillos, A. The Zebrafish Embryo as a Model to Test Protective Effects of Food Antioxidant Compounds. Molecules 2021, 26, 5786. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26195786
Arteaga C, Boix N, Teixido E, Marizande F, Cadena S, Bustillos A. The Zebrafish Embryo as a Model to Test Protective Effects of Food Antioxidant Compounds. Molecules. 2021; 26(19):5786. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26195786
Chicago/Turabian StyleArteaga, Cristina, Nuria Boix, Elisabet Teixido, Fernanda Marizande, Santiago Cadena, and Alberto Bustillos. 2021. "The Zebrafish Embryo as a Model to Test Protective Effects of Food Antioxidant Compounds" Molecules 26, no. 19: 5786. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26195786